
Exploring the Multifunctional Roles of Odontoglossum Ringspot Virus P126 in Facilitating Cymbidium Mosaic Virus Cell-to-Cell Movement during Mixed Infection

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Growth Conditions
2.2. RT-PCR Detection of CymMV and ORSV
2.3. Virus Isolation, Purification, Inoculation and RNA Extraction
2.4. Plasmid Construction
2.5. In Vitro Transcription
2.6. Tissue Blotting and RNA Blot Analysis
2.7. Agrobacterium Infiltration and GFP Imaging
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Construction of ORSV and CymMV Infectious cDNA Clones
3.2. Asymmetric Synergism of CymMV and ORSV Infection in N. benthamiana and Phalaenopsis Orchid
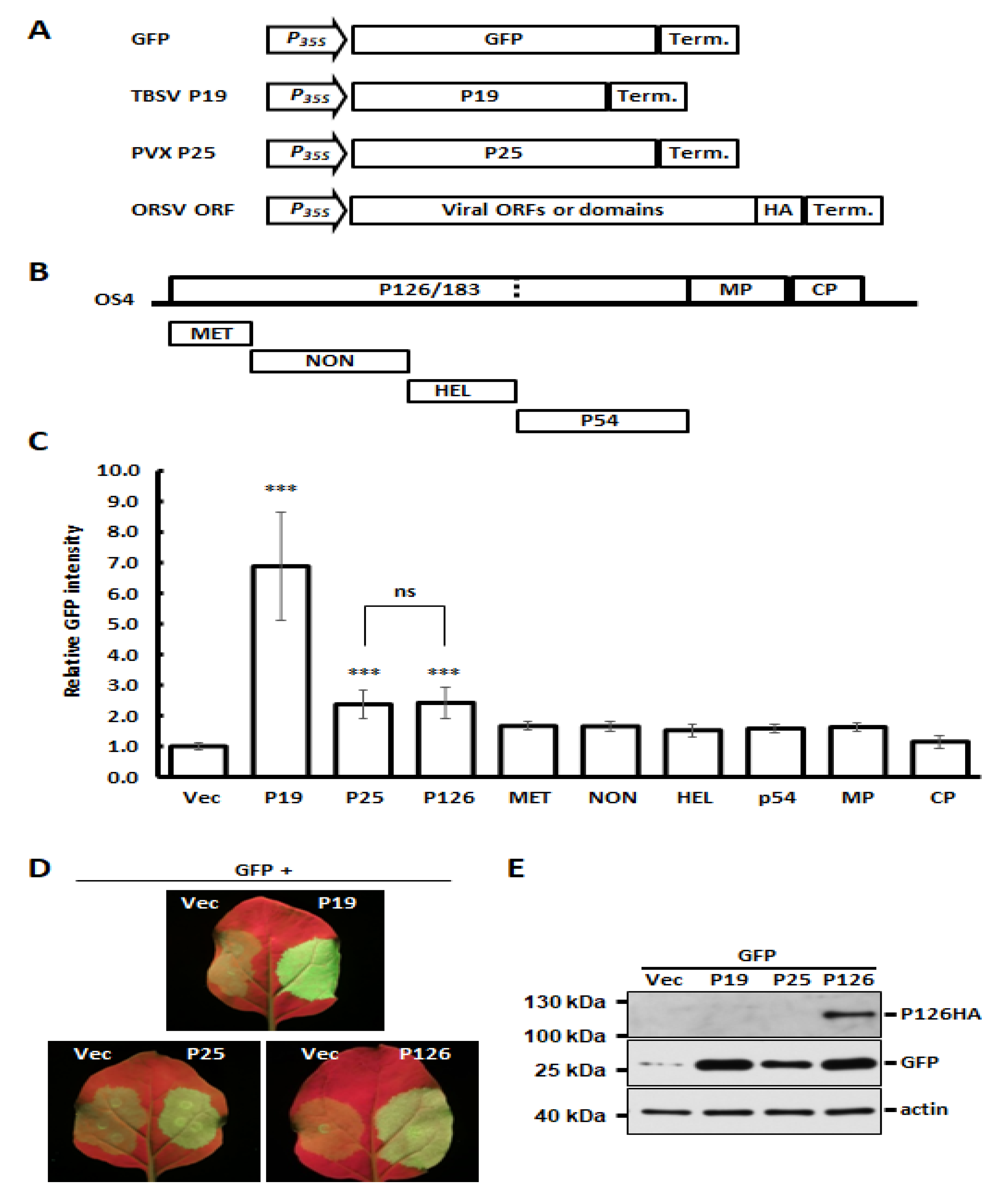
3.3. Identification of ORSV-Encoded P126 as a VSR
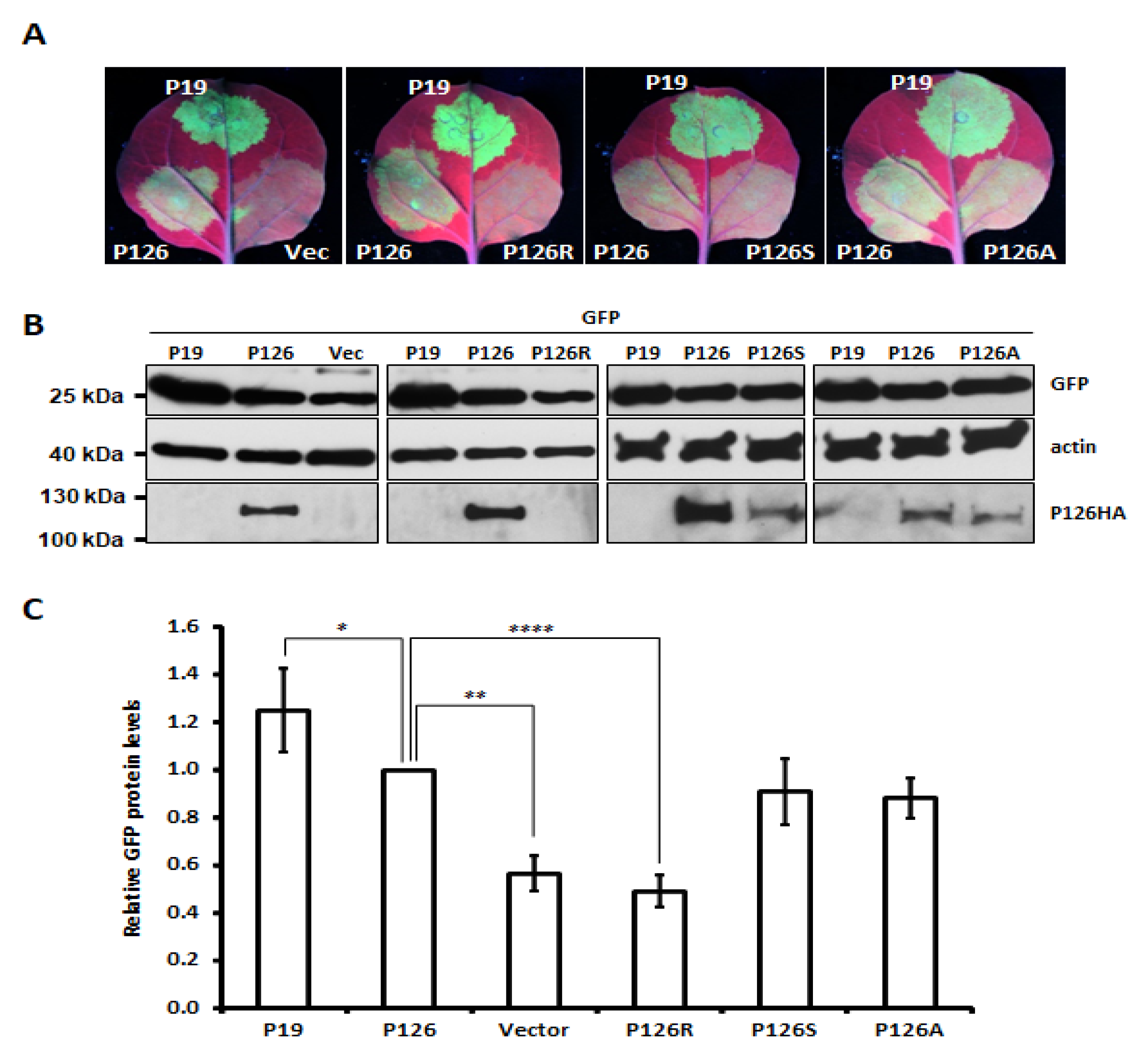
3.4. Single Amino Acid Substitution Abolishes ORSV P126 VSR Activity
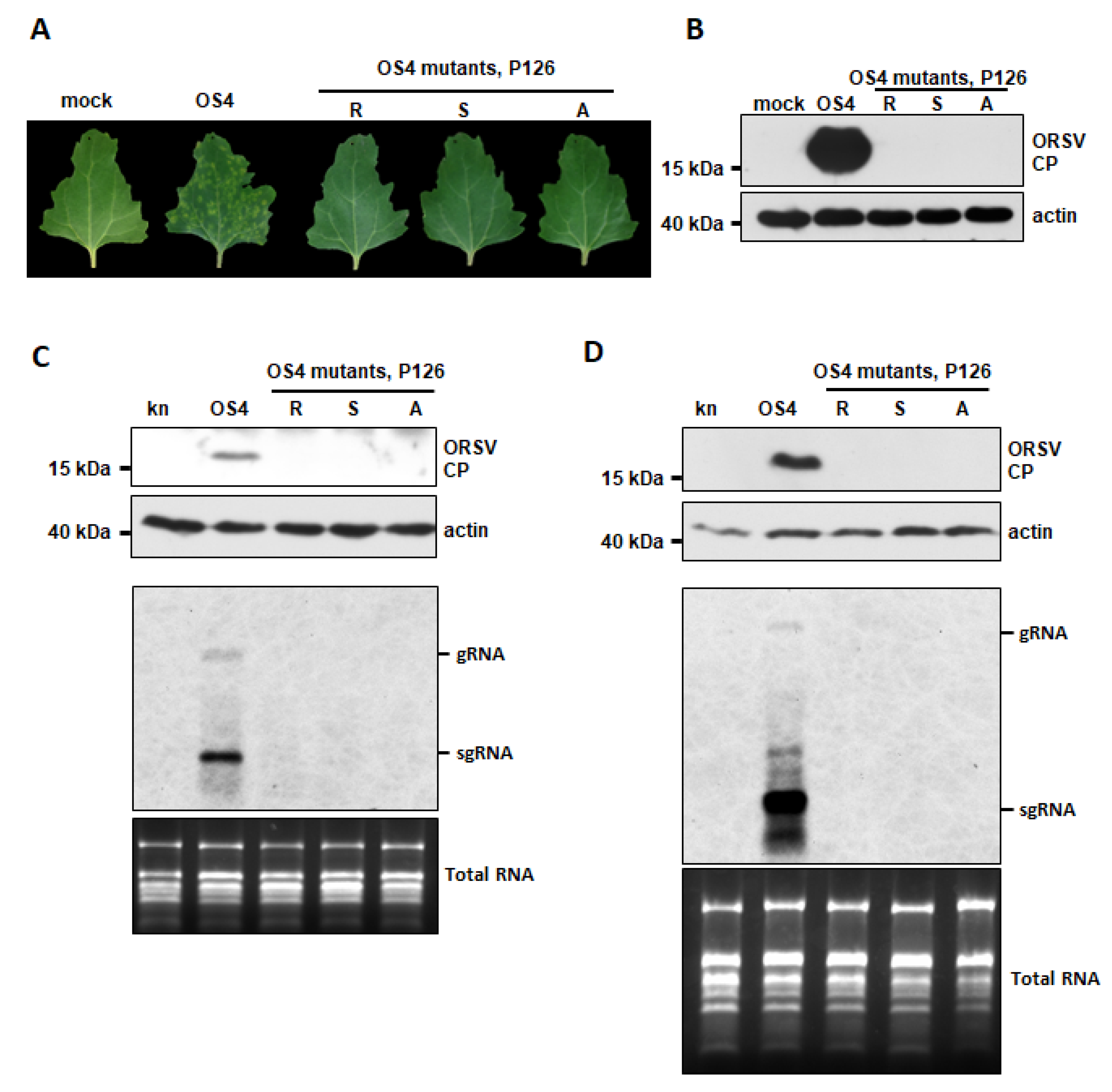
3.5. ORSV P126 VSR Activity Uncouples ORSV Replication
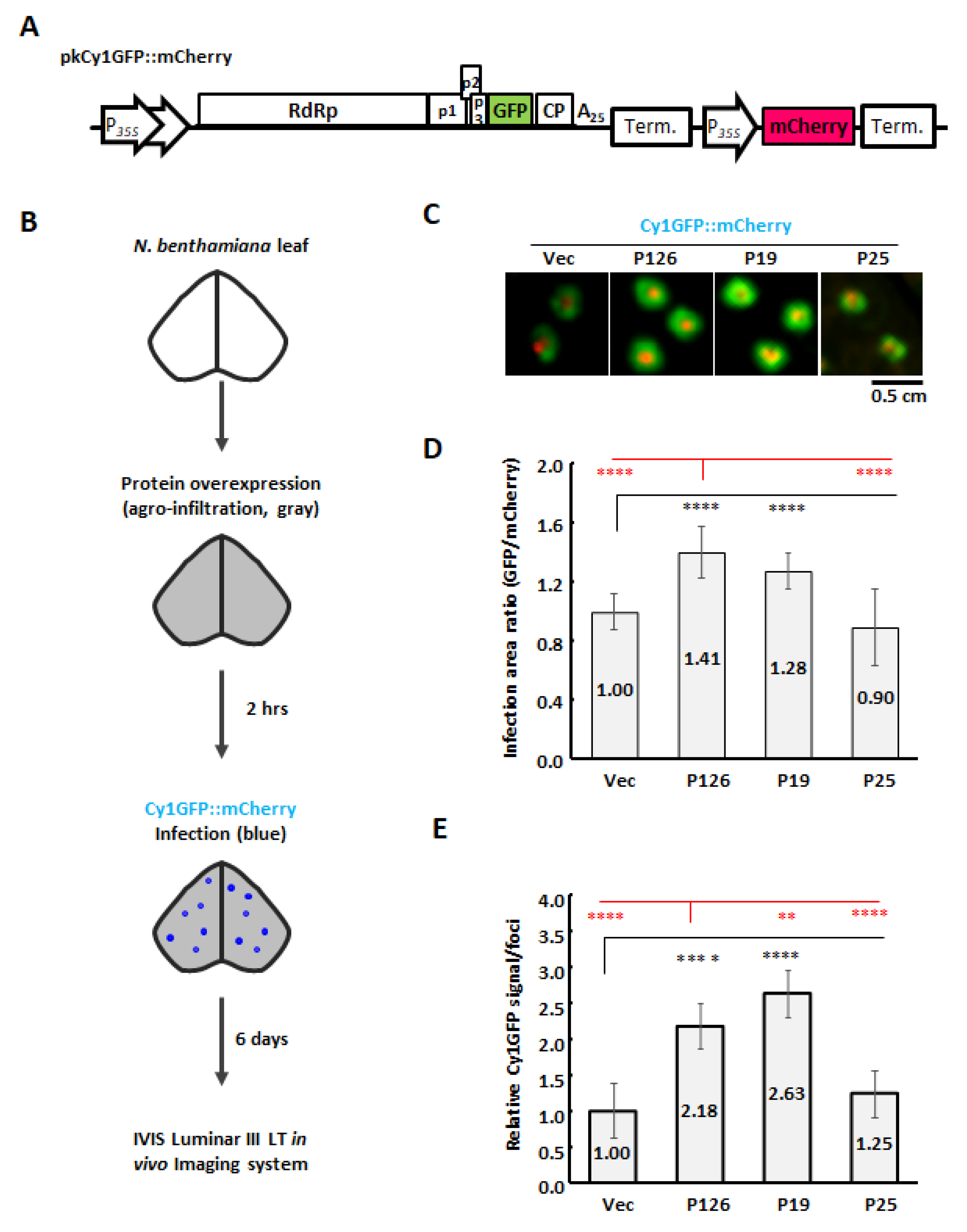
3.6. ORSV P126 Is the Synergistic Determinant That Enhances CymMV Cell-to-Cell Movement
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Syller, J. Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol. Plant Pathol. 2011, 13, 204–216. [Google Scholar] [CrossRef] [PubMed]
- García-Cano, E.; Resende, R.O.; Fernandez-Munoz, R.; Moriones, E. Synergistic Interaction Between Tomato chlorosis virus and Tomato spotted wilt virus Results in Breakdown of Resistance in Tomato. Phytopathology 2006, 96, 1263–1269. [Google Scholar] [CrossRef]
- Hu, W.-W.; Wong, S.-M.; Loh, C.-S.; Goh, C.-J. Synergism in replication of cymbidium mosaic potexvirus (CymMV) and odontoglossum ringspot tobamovirus (ORSV) RNA in orchid protoplasts. Arch. Virol. 1998, 143, 1265–1275. [Google Scholar] [CrossRef]
- Karyeija, R.; Kreuze, J.; Gibson, R.; Valkonen, J. Synergistic Interactions of a Potyvirus and a Phloem-Limited Crinivirus in Sweet Potato Plants. Virology 2000, 269, 26–36. [Google Scholar] [CrossRef]
- Vance, V.B. Replication of Potato virus-X RNA is altered in coinfections with Potato virus-Y. Virology 1991, 182, 486–494. [Google Scholar] [PubMed]
- Ajjikuttira, P.; Loh, C.-S.; Wong, S.-M. Reciprocal function of movement proteins and complementation of long-distance movement of Cymbidium mosaic virus RNA by Odontoglossum ringspot virus coat protein. J. Gen. Virol. 2005, 86, 1543–1553. [Google Scholar] [CrossRef]
- Rao, A.L.N.; Cooper, B.; Deom, C.M. Defective movement of viruses in the family Bromoviridae is differentially comple-mented in Nicotiana benthamiana expressing tobamovirus or dianthovirus movement proteins. Phytopathology 1998, 88, 666–672. [Google Scholar] [CrossRef]
- Ryang, B.-S.; Kobori, T.; Matsumoto, T.; Kosaka, Y.; Ohki, S.T. Cucumber mosaic virus 2b protein compensates for restricted systemic spread of Potato virus Y in doubly infected tobacco. J. Gen. Virol. 2004, 85, 3405–3414. [Google Scholar] [CrossRef] [PubMed]
- Hannon, G.J. RNA interference. Nat. Cell Biol. 2002, 418, 244–251. [Google Scholar] [CrossRef]
- Ding, S.-W. RNA-based antiviral immunity. Nat. Rev. Immunol. 2010, 10, 632–644. [Google Scholar] [CrossRef]
- Li, F.; Wang, A. RNA-Targeted Antiviral Immunity: More Than Just RNA Silencing. Trends Microbiol. 2019, 27, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Pruss, G.; Ge, X.; Shi, X.M.; Carrington, J.; Vance, V.B. Plant viral synergism: The potyviral genome encodes a broad-range pathogenicity enhancer that transactivates replication of heterologous viruses. Plant Cell 1997, 9, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Cuellar, W.; Kreuze, J.; Rajamäki, M.-L.; Cruzado, K.R.; Untiveros, M.; Valkonen, J.P.T. Elimination of antiviral defense by viral RNase III. Proc. Natl. Acad. Sci. USA 2009, 106, 10354–10358. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.A.; Valkonen, J.; Rajamäki, M.-L.; Lehto, K. The 2b Silencing Suppressor of a Mild Strain of Cucumber mosaic virus Alone Is Sufficient for Synergistic Interaction with Tobacco mosaic virus and Induction of Severe Leaf Malformation in 2b-Transgenic Tobacco Plants. Mol. Plant Microbe Interact. 2011, 24, 685–693. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Harries, P.A.; Palanichelvam, K.; Bhat, S.; Nelson, R.S. Tobacco mosaic virus 126-kDa Protein Increases the Susceptibility of Nicotiana tabacum to Other Viruses and Its Dosage Affects Virus-Induced Gene Silencing. Mol. Plant Microbe Interact. 2008, 21, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.; Griffiths, M. Manual of Orchids; Timber Press: Portland, OR, USA, 2005. [Google Scholar]
- Chang, C.; Chen, Y.-C.; Hsu, Y.-H.; Wu, J.-T.; Hu, C.-C.; Chang, W.-C.; Lin, N.-S. Transgenic resistance to Cymbidium mosaic virus in Dendrobium expressing the viral capsid protein gene. Transgenic. Res. 2005, 14, 41–46. [Google Scholar] [CrossRef]
- Zettler, F.W.; Ko, N.J.; Wisler, G.C.; Elliott, M.S.; Wong, S.M. Viruses of orchids and their control. Plant Dis. 1990, 74, 621–626. [Google Scholar] [CrossRef]
- Wong, S.; Chng, C.; Lee, Y.; Tan, K.; Zettler, F. Incidence of cymbidium mosaic and odontoglossum ringspot viruses and their significance in orchid cultivation in Singapore. Crop. Prot. 1994, 13, 235–239. [Google Scholar] [CrossRef]
- Moraes, L.A.; Krause-Sakate, R.; Pavan, M.A. Incidence and characterization of viruses infecting orchids in São Paulo state, Brazil. Trop. Plant Pathol. 2017, 42, 126–131. [Google Scholar] [CrossRef]
- Liu, F.; Han, Y.; Li, W.; Shi, X.; Xu, W.; Lin, M. Incidence of Cymbidium mosaic virus and Odontoglossum ringspot virus affecting Oncidium orchids in Hainan Island, China. Crop. Prot. 2013, 54, 176–180. [Google Scholar] [CrossRef]
- McMillan, R.T.; Palmateer, A.; Vendrame, W.A. Survey for Cymbidium mosaic and Odontoglossum ring spot viruses in domestic and international orchids. Phytopathology 2006, 96, S76. [Google Scholar]
- Hu, J.S.; Ferreira, S.; Wang, M.; Xu, M.Q. Detection of Cymbidium mosaic virus, Odontoglossum ringspot virus, Tomato spotted wilt virus, and potyviruses infecting orchids in Hawaii. Plant Dis. 1993, 77, 464–468. [Google Scholar] [CrossRef]
- Pearson, M.N.; Cole, J.S. Further Observations on the Effects of Cymbidium mosaic virus and Odontoglossum ringspot virus on the Growth of Cymbidium Orchids. J. Phytopathol. 1991, 131, 193–198. [Google Scholar] [CrossRef]
- Ryu, K.H.; Park, W.M. The complete nucleotide sequence and genome organization of odontoglossum ringspot tobamovirus RNA. Arch. Virol. 1995, 140, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Yu, H.H.; Wong, S.M. Mutation of Phe50 to Ser50 in the 126/183-kDa proteins of Odontoglossum ringspot virus abolishes virus replication but can be complemented and restored by exact reversion. J. Gen. Virol. 2004, 85, 2447–2457. [Google Scholar] [CrossRef]
- Rabindran, S.; Robertson, C.; Achor, D.; Holt, C.A.; Dawson, W.O.; German-Retana, S. Odontoglossum ringspot virus host range restriction in Nicotiana sylvestris maps to the replicase gene. Mol. Plant Pathol. 2005, 6, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.S.; Liu, J.; Cheng, N.-H.; Folimonov, A.; Hou, Y.-M.; Bao, Y.; Katagi, C.; Carter, S.A.; Nelson, R.S. The Tobacco mosaic virus 126-kDa Protein Associated with Virus Replication and Movement Suppresses RNA Silencing. Mol. Plant Microbe Interact. 2004, 17, 583–592. [Google Scholar] [CrossRef]
- Kubota, K.; Tsuda, S.; Tamai, A.; Meshi, T. Tomato mosaic virus replication protein suppresses virus-targeted posttranscrip-tional gene silencing. J. Virol. 2003, 77, 11016–11026. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Lin, S.-S.; Hung, T.-H.; Li, T.-K.; Lin, N.-C.; Shen, T.-L. Multiple Domains of the Tobacco mosaic virus p126 Protein Can Independently Suppress Local and Systemic RNA Silencing. Mol. Plant Microbe Interact. 2012, 25, 648–657. [Google Scholar] [CrossRef]
- Csorba, T.; Bovi, A.; Dalmay, T.; Burgyan, J. The p122 subunit of Tobacco mosaic virus replicase is a potent silencing suppressor and compromises both small interfering RNA- and microRNA-mediated pathways. J. Virol. 2007, 81, 11768–11780. [Google Scholar] [CrossRef]
- Kurihara, Y.; Inaba, N.; Kutsuna, N.; Takeda, A.; Tagami, Y.; Watanabe, Y. Binding of tobamovirus replication protein with small RNA duplexes. J. Gen. Virol. 2007, 88, 2347–2352. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.M.; Mahtani, P.H.; Lee, K.C.; Yu, H.H.; Tan, Y.; Neo, K.K.; Chan, Y.; Wu, M.; Chng, C.G. Cymbidium mosaic potexvirus RNA: Complete nucleotide sequence and phylogenetic analysis. Arch. Virol. 1997, 142, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-C.; Chen, C.-E.; Tsai, M.-H.; Wang, H.-I.; Su, H.-J.; Yeh, H.-H. Cymbidium mosaic potexvirus isolate-dependent host movement systems reveal two movement control determinants and the coat protein is the dominant. Virology 2009, 388, 147–159. [Google Scholar] [CrossRef][Green Version]
- Pai, H.; Jean, W.; Lee, Y.; Chang, Y.A.; Lin, N. Genome-wide analysis of small RNAs from Odontoglossum ringspot virus and Cymbidium mosaic virus synergistically infecting Phalaenopsis. Mol. Plant Pathol. 2019, 21, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-Y.; Pai, H.; Hou, L.-Y.; Lee, S.-C.; Lin, T.-T.; Chang, C.-H.; Hsu, F.-C.; Hsu, Y.-H.; Lin, N.-S. Dual resistance of transgenic plants against Cymbidium mosaic virus and Odontoglossum ringspot virus. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Lee, S.-C.; Chang, Y.-C. Multiplex RT-PCR detection of two orchid viruses with an internal control of plant nad5 mRNA. Plant Pathol. Bull. 2006, 15, 187–196. [Google Scholar]
- Lin, N.S.; Chen, C.C. Association of Bamboo mosaic virus (BoMV) and BoMV-specific electron-dense crystalline bodies with chloroplasts. Phytopathology 1991, 81, 1551–1555. [Google Scholar] [CrossRef]
- Liou, M.-R.; Huang, Y.-W.; Hu, C.-C.; Lin, N.-S.; Hsu, Y.-H. A dual gene-silencing vector system for monocot and dicot plants. Plant Biotechnol. J. 2013, 12, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-C.; Chen, H.-H.; Tsai, W.-C.; Chen, W.-H.; Su, H.-J.; Chang, D.C.-N.; Yeh, H.-H. Strategies for Functional Validation of Genes Involved in Reproductive Stages of Orchids. Plant Physiol. 2007, 143, 558–569. [Google Scholar] [CrossRef]
- Bendahmane, A.; Farnham, G.; Moffett, P.; Baulcombe, D.C. Constitutive gain-of-function mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. Plant J. 2002, 32, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.S.; Hsu, Y.H.; Hsu, H.T. Immunological detection of plant viruses and a mycoplasma-like organism by direct tissue blotting on nitrocellulose membranes. Phytopathology 1990, 80, 824–828. [Google Scholar] [CrossRef]
- Lin, N.S.; Lee, Y.S.; Lin, B.; Lee, C.W.; Hsu, Y.H. The open reading frame of bamboo mosaic potexvirus satellite RNA is not essential for its replication and can be replaced with a bacterial gene. Proc. Natl. Acad. Sci. USA 1996, 93, 3138–3142. [Google Scholar] [CrossRef]
- Palani, P.V.; Lin, N.-S. Northern Analysis of Viral Plus- and Minus-Strand RNAs. Curr. Protoc. Microbiol. 2007, 16, 16E.3.1–16E.3.32. [Google Scholar]
- Johansen, L.K.; Carrington, J.C. Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacte-rium-mediated transient expression system. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Stephan, D.; Slabber, C.; George, G.; Ninov, V.; Francis, K.P.; Burger, J.T. Visualization of plant viral suppressor silencing activity in intact leaf lamina by quantitative fluorescent imaging. Plant Methods 2011, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, B.R.T.; Nicolini, C.; Lucinda, N.; Orílio, A.F.; Nagata, T. A simplified approach to construct infectious cDNA clones of a tobamovirus in a binary vector. J. Virol. Methods 2014, 198, 32–36. [Google Scholar] [CrossRef]
- Turpen, T.H.; Turpen, A.M.; Weinzettl, N.; Kumagai, M.H.; Dawson, W.O. Transfection of whole plants from wounds inocu-lated with Agrobacterium tumefaciens containing cDNA of Tobacco mosaic virus. J. Virol. Methods 1993, 42, 227–239. [Google Scholar] [CrossRef]
- Qu, F.; Morris, T.J. Suppressors of RNA silencing encoded by plant viruses and their role in viral infections. FEBS Lett. 2005, 579, 5958–5964. [Google Scholar] [CrossRef]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Genet. 2013, 11, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Lederer, C.; Baulcombe, D.C. A Viral Movement Protein Prevents Spread of the Gene Silencing Signal in Nicotiana benthamiana. Cell 2000, 103, 157–167. [Google Scholar] [CrossRef]
- Wang, X.; Kelman, Z.; Culver, J.N. Helicase ATPase activity of the Tobacco mosaic virus 126-kDa protein modulates replicase complex assembly. Virology 2010, 402, 292–302. [Google Scholar] [CrossRef]
- Ishibashi, K.; Nishikiori, M.; Ishikawa, M. Interactions Between Tobamovirus Replication Proteins and Cellular Factors: Their Impacts on Virus Multiplication. Mol. Plant Microbe Interact. 2010, 23, 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Burgyan, J.; Martelli, G.P. Molecular Biology of Tombusviridae. Adv. Clin. Chem. 1994, 44, 381–428. [Google Scholar]
- Powers, J.G.; Sit, T.L.; Qu, F.; Morris, T.J.; Kim, K.-H.; Lommel, S.A. A Versatile Assay for the Identification of RNA Silencing Suppressors Based on Complementation of Viral Movement. Mol. Plant Microbe Interact. 2008, 21, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Hirashima, K.; Watanabe, Y. RNA Helicase Domain of Tobamovirus Replicase Executes Cell-to-Cell Movement Possibly through Collaboration with Its Nonconserved Region. J. Virol. 2003, 77, 12357–12362. [Google Scholar] [CrossRef][Green Version]
- Hirashima, K.; Watanabe, Y. Tobamovirus Replicase Coding Region Is Involved in Cell-to-Cell Movement. J. Virol. 2001, 75, 8831–8836. [Google Scholar] [CrossRef]
- Liu, J.; Blancaflor, E.; Nelson, R.S. The Tobacco Mosaic Virus 126-Kilodalton Protein, a Constituent of the Virus Replication Complex, Alone or within the Complex Aligns with and Traffics along Microfilaments. Plant Physiol. 2005, 138, 1853–1865. [Google Scholar] [CrossRef]
- Kawakami, S.; Watanabe, Y.; Beachy, R.N. Tobacco mosaic virus infection spreads cell to cell as intact replication complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 6291–6296. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-C.; Pai, H.; Huang, Y.-W.; He, M.-H.; Song, Y.-L.; Kuo, S.-Y.; Chang, W.-C.; Hsu, Y.-H.; Lin, N.-S. Exploring the Multifunctional Roles of Odontoglossum Ringspot Virus P126 in Facilitating Cymbidium Mosaic Virus Cell-to-Cell Movement during Mixed Infection. Viruses 2021, 13, 1552. https://doi.org/10.3390/v13081552
Lee S-C, Pai H, Huang Y-W, He M-H, Song Y-L, Kuo S-Y, Chang W-C, Hsu Y-H, Lin N-S. Exploring the Multifunctional Roles of Odontoglossum Ringspot Virus P126 in Facilitating Cymbidium Mosaic Virus Cell-to-Cell Movement during Mixed Infection. Viruses. 2021; 13(8):1552. https://doi.org/10.3390/v13081552
Chicago/Turabian StyleLee, Shu-Chuan, Hsuan Pai, Ying-Wen Huang, Meng-Hsun He, Yun-Lin Song, Song-Yi Kuo, Wen-Chi Chang, Yau-Heiu Hsu, and Na-Sheng Lin. 2021. "Exploring the Multifunctional Roles of Odontoglossum Ringspot Virus P126 in Facilitating Cymbidium Mosaic Virus Cell-to-Cell Movement during Mixed Infection" Viruses 13, no. 8: 1552. https://doi.org/10.3390/v13081552
APA StyleLee, S.-C., Pai, H., Huang, Y.-W., He, M.-H., Song, Y.-L., Kuo, S.-Y., Chang, W.-C., Hsu, Y.-H., & Lin, N.-S. (2021). Exploring the Multifunctional Roles of Odontoglossum Ringspot Virus P126 in Facilitating Cymbidium Mosaic Virus Cell-to-Cell Movement during Mixed Infection. Viruses, 13(8), 1552. https://doi.org/10.3390/v13081552