
BK Polyomavirus—Biology, Genomic Variation and Diagnosis

 , , , ,
, , , ,
Abstract
1. Introduction
2. Description of BK Polyomavirus
3. Molecular Polymorphism of the BK Polyomavirus
4. Pathogenesis of BKPyV Infections
4.1. BKPyV Infection in Kidney Transplants Recipients
4.2. BKPyV Infection after Allogeneic Haematopoietic Stem-Cell Transplantation
4.3. Oncogenic Potential of BKPyV
4.4. Therapeutic Management
5. Methods of BK Polyomavirus Diagnosis
5.1. PCR
5.2. qPCR
5.3. Genotyping
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Pinto, M.; Dobson, S. BK and JC virus: A review. J. Infect. 2014, 68, S2–S8. [Google Scholar] [CrossRef]
- Ambalathingal, G.R.; Francis, R.S.; Smyth, M.J.; Smith, C.; Khanna, R. BK Polyomavirus: Clinical Aspects, Immune Regulation, and Emerging Therapies. Clin. Microbiol. Rev. 2017, 30, 503–528. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, C.; Orio, J.; Collette, S.; Senecal, L.; Hebert, M.J.; Renoult, E.; Tibbles, L.A.; Delisle, J.S. BK Polyomavirus and the Transplanted Kidney: Immunopathology and Therapeutic Approaches. Transplantation 2016, 100, 2276–2287. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Van Doorslaer, K.; Peretti, A.; Geoghegan, E.M.; Tisza, M.J.; An, P.; Katz, J.P.; Pipas, J.M.; McBride, A.A.; Camus, A.C.; et al. The Ancient Evolutionary History of Polyomaviruses. PLoS Pathog. 2016, 12, e1005574. [Google Scholar] [CrossRef]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New Human Papovavirus (BK) Isolated from Urine after Renal Transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef]
- Padgett, B.L.; Walker, D.L.; Zurhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of Papova-Like Virus from Human Brain with Progressive Multifocal Leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Vigil, D.; Konstantinov, N.K.; Barry, M.; Harford, A.M.; Servilla, K.S.; Kim, Y.H.; Sun, Y.; Ganta, K.; Tzamaloukas, A.H. BK nephropathy in the native kidneys of patients with organ transplants: Clinical spectrum of BK infection. World J. Transpl. 2016, 6, 472–504. [Google Scholar] [CrossRef] [PubMed]
- De Gascun, C.F.; Carr, M.J. Human Polyomavirus Reactivation: Disease Pathogenesis and Treatment Approaches. Clin. Dev. Immunol. 2013. [Google Scholar] [CrossRef]
- Prezioso, C.; Van Ghelue, M.; Moens, U.; Pietropaolo, V. HPyV6 and HPyV7 in urine from immunocompromised patients. Virol. J. 2021, 18, 24. [Google Scholar] [CrossRef]
- Allander, T.; Andreasson, K.; Gupta, S.; Bjerkner, A.; Bogdanovic, G.; Persson, M.A.A.; Dalianis, T.; Ramqvist, T.; Andersson, B. Identification of a third human polyomavirus. J. Virol. 2007, 81, 4130–4136. [Google Scholar] [CrossRef]
- Gaynor, A.M.; Nissen, M.D.; Whiley, D.M.; Mackay, I.M.; Lambert, S.B.; Wu, G.; Brennan, D.C.; Storch, G.A.; Sloots, T.P.; Wang, D. Identification of a novel polyomavirus from patients with acute respiratory tract infections. PLoS Pathog. 2007, 3, e64. [Google Scholar] [CrossRef]
- Feng, H.C.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Van der Meijden, E.; Janssens, R.W.A.; Lauber, C.; Bavinck, J.N.B.; Gorbalenya, A.E.; Feltkamp, M.C.W. Discovery of a New Human Polyomavirus Associated with Trichodysplasia Spinulosa in an Immunocompromized Patient. PLoS Pathog. 2010, 6, e1001024. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, R.M.; Pastrana, D.V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel Cell Polyomavirus and Two Previously Unknown Polyomaviruses Are Chronically Shed from Human Skin. Cell Host Microbe 2010, 7, 509–515. [Google Scholar] [CrossRef]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and characterization of a novel putative human polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef]
- Scuda, N.; Hofmann, J.; Calvignac-Spencer, S.; Ruprecht, K.; Liman, P.; Kuehn, J.; Hengel, H.; Ehlers, B. A Novel Human Polyomavirus Closely Related to the African Green Monkey-Derived Lymphotropic Polyomavirus. J. Virol. 2011, 85, 4586–4590. [Google Scholar] [CrossRef]
- Buck, C.B.; Phan, G.Q.; Raiji, M.T.; Murphy, P.M.; McDermott, D.H.; McBride, A.A. Complete Genome Sequence of a Tenth Human Polyomavirus. J. Virol. 2012, 86, 10887. [Google Scholar] [CrossRef] [PubMed]
- Ondov, B.D.; Starrett, G.J.; Sappington, A.; Kostic, A.; Koren, S.; Buck, C.B.; Phillippy, A.M. Mash Screen: High-throughput sequence containment estimation for genome discovery. Genome Biol. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.S.; Reyes, A.; Antonio, M.; Saha, D.; Ikumapayi, U.N.; Adeyemi, M.; Stine, O.C.; Skelton, R.; Brennan, D.C.; Mkakosya, R.S.; et al. Discovery of STL polyomavirus, a polyomavirus of ancestral recombinant origin that encodes a unique T antigen by alternative splicing. Virology 2013, 436, 295–303. [Google Scholar] [CrossRef]
- Korup, S.; Rietscher, J.; Calvignac-Spencer, S.; Trusch, F.; Hofmann, J.; Moens, U.; Sauer, I.; Voigt, S.; Schmuck, R. Identification of a novel human polyomavirus in organs of the gastrointestinal tract. J. Neurovirol. 2013, 19, 286–287. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Pereira, M.; Rhodes, R.H.; An, P.; Pipas, J.M.; Jain, K.; Kapoor, A.; Briese, T.; Faust, P.L.; Lipkin, W.I. Identification of a Novel Polyomavirus in a Pancreatic Transplant Recipient with Retinal Blindness and Vasculitic Myopathy. J. Infect. Dis. 2014, 210, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Kamminga, S.; van der Meijden, E.; Feltkamp, M.C.W.; Zaaijer, H.L. Seroprevalence of fourteen human polyomaviruses determined in blood donors. PLoS ONE 2018, 13, e0206273. [Google Scholar] [CrossRef]
- Boothpur, R.; Brennan, D.C. Human polyoma viruses and disease with emphasis on clinical BK and JC. J. Clin. Virol. 2010, 47, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Moens, U.; Prezioso, C.; Pietropaolo, V. Genetic Diversity of the Noncoding Control Region of the Novel Human Polyomaviruses. Viruses 2020, 12, 1406. [Google Scholar] [CrossRef]
- Sharma, R.; Tzetzo, S.; Patel, S.; Zachariah, M.; Sharma, S.; Melendy, T. BK Virus in Kidney Transplant: Current Concepts, Recent Advances, and Future Directions. Exp. Clin. Transpl. 2016, 14, 377–384. [Google Scholar] [CrossRef][Green Version]
- Kean, J.M.; Rao, S.; Wang, M.; Garcea, R.L. Seroepidemiology of Human Polyomaviruses. PLoS Pathog. 2009, 5, e1000363. [Google Scholar] [CrossRef]
- Sharma, R.; Zachariah, M. BK Virus Nephropathy: Prevalence, Impact and Management Strategies. Int. J. Nephrol. Renovasc. Dis. 2020, 13, 187–192. [Google Scholar] [CrossRef]
- Stolt, A.; Sasnauskas, K.; Koskela, P.; Lehtinen, M.; Dillner, J. Seroepidemiology of the human polyomaviruses. J. Gen. Virol. 2003, 84, 1499–1504. [Google Scholar] [CrossRef]
- Egli, A.; Infanti, L.; Dumoulin, A.; Buser, A.; Samaridis, J.; Stebler, C.; Gosert, R.; Hirsch, H.H. Prevalence of Polyomavirus BK and JC Infection and Replication in 400 Healthy Blood Donors. J. Infect. Dis. 2009, 199, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Karimi Dehcheshmeh, L.; Makvandi, M.; Timori, A. Prevalence of Human Polyomavirus JC and BK in Normal Population. Asian Pac. J. Cancer 2020, 21, 2877–2882. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Steiger, J. Polyomavirus BK. Lancet Infect. Dis. 2003, 3, 611–623. [Google Scholar] [CrossRef]
- Hirsch, H.H. BK virus: Opportunity makes a pathogen. Clin. Infect. Dis. 2005, 41, 354–360. [Google Scholar] [CrossRef]
- Krumbholz, A.; Zell, R.; Egerer, R.; Sauerbrei, A.; Helming, A.; Gruhry, B.; Wutzler, P. Prevalence of BK virus subtype I in Germany. J. Med. Virol. 2006, 78, 1588–1598. [Google Scholar] [CrossRef]
- Krumbholz, A.; Bininda-Emonds, O.R.P.; Wutzler, P.; Zell, R. Evolution of four BK virus subtypes. Infect. Genet. Evol. 2008, 8, 632–643. [Google Scholar] [CrossRef]
- Tremolada, S.; Delbue, S.; Larocca, S.; Carloni, C.; Elia, F.; Khalili, K.; Gordon, J.; Ferrante, P. Polymorphisms of the BK virus subtypes and their influence on viral in vitro growth efficiency. Virus Res. 2010, 149, 190–196. [Google Scholar] [CrossRef][Green Version]
- Randhawa, P.; Brennan, D.C. BK virus infection in transplant recipients: An overview and update. Am. J. Transpl. 2006, 6, 2000–2005. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.X.; Abend, J.R.; Johnson, S.F.; Imperiale, M.J. The role of polyomaviruses in human disease. Virology 2009, 384, 266–273. [Google Scholar] [CrossRef] [PubMed]
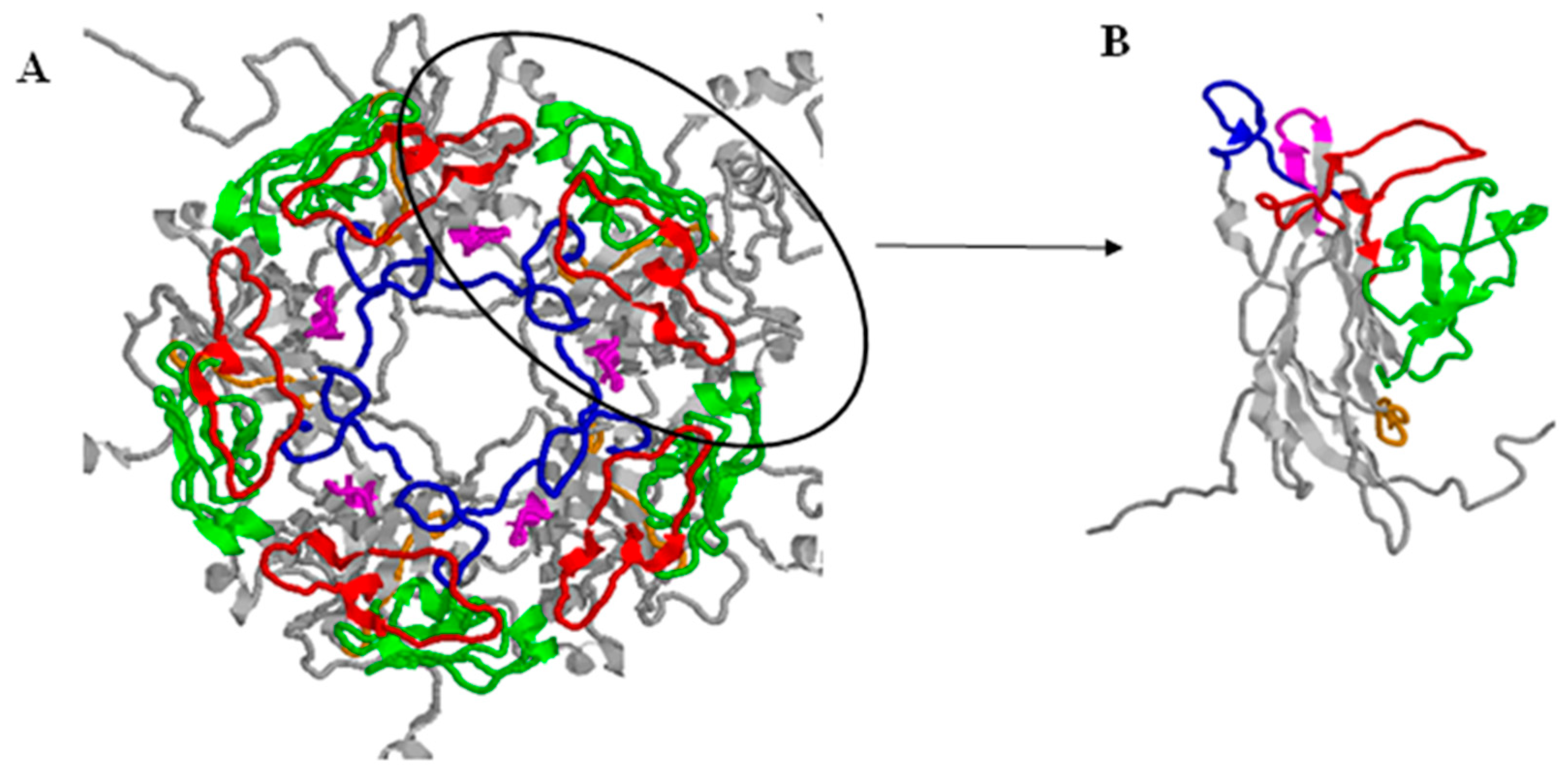
- Hurdiss, D.L.; Morgan, E.L.; Thompson, R.F.; Prescott, E.L.; Panou, M.M.; Macdonald, A.; Ranson, N.A. New Structural Insights into the Genome and Minor Capsid Proteins of BK Polyomavirus using Cryo-Electron Microscopy. Structure 2016, 24, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.Y.; Chen, H.Y.; Wang, M.L.; Chen, P.L.; Chang, C.F.; Chen, L.S.; Shen, C.H.; Ou, W.C.; Tsai, M.D.; Hsu, P.H.; et al. Global analysis of modifications of the human BK virus structural proteins by LC-MS/MS. Virology 2010, 402, 164–176. [Google Scholar] [CrossRef]
- Anzivino, E.; Zingaropoli, M.A.; Iannetta, M.; Pietropaolo, V.A.; Oliva, A.; Iori, F.; Ciardi, A.; Rodio, D.M.; Antonini, F.; Fedele, C.G.; et al. Archetype and Rearranged Non-coding Control Regions in Urothelial Bladder Carcinoma of Immunocompetent Individuals. Cancer Genom. Proteom. 2016, 13, 499–509. [Google Scholar] [CrossRef]
- Moens, U.; Van Ghelue, M. Polymorphism in the genome of non-passaged human polyomavirus BK: Implications for cell tropism and the pathological role of the virus. Virology 2005, 331, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Rinaldo, C.H.; Funk, G.A.; Egli, A.; Ramos, E.; Drachenberg, C.B.; Hirsch, H.H. Polyomavirus BK with rearranged noncoding control region emerge in vivo in renal transplant patients and increase viral replication and cytopathology. J. Exp. Med. 2008, 205, 841–852. [Google Scholar] [CrossRef]
- Liimatainen, H.; Weseslindtner, L.; Strassl, R.; Aberle, S.W.; Bond, G.; Auvinen, E. Next-generation sequencing shows marked rearrangements of BK polyomavirus that favor but are not required for polyomavirus-associated nephropathy. J. Clin. Virol. 2020, 122, 104215. [Google Scholar] [CrossRef] [PubMed]
- Jin, L. Rapid Genomic Typing of BK Virus Directly from Clinical Specimens. Mol. Cell Probes 1993, 7, 331–334. [Google Scholar] [CrossRef]
- Li, J.; Gibson, P.E.; Booth, J.C.; Clewley, J.P. Genomic Typing of BK Virus in Clinical Specimens by Direct Sequencing of Polymerase Chain-Reaction Products. J. Med. Virol. 1993, 41, 11–17. [Google Scholar] [CrossRef]
- Morel, V.; Martin, E.; Francois, C.; Helle, F.; Faucher, J.; Mourez, T.; Choukroun, G.; Duverlie, G.; Castelain, S.; Brochot, E. A Simple and Reliable Strategy for BK Virus Subtyping and Subgrouping. J. Clin. Microbiol. 2017, 55, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, H.; Saukko, P.J.; Tertti, R.; Metsarinne, K.P.; Carr, M.J.; Crowley, B.; Sakurada, K.; Zheng, H.-Y.; Kitamura, T.; Yogo, Y. Identification of a genomic subgroup of BK polyomavirus spread in European populations. J. Gen. Virol. 2006, 87, 3201–3208. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zheng, H.Y.; Zhong, S.; Ikegaya, H.; He, H.X.; Wei, W.; He, Y.Y.; Kobayashi, N.; Honjo, T.; Takasaka, T.; et al. Subtype IV of the BK polyomavirus is prevalent in East Asia. Arch. Virol. 2006, 151, 2419–2429. [Google Scholar] [CrossRef]
- Ikegaya, H.; Motani, H.; Saukko, P.; Sato, K.; Akutsu, T.; Sakurada, K. BK virus genotype distribution offers information of tracing the geographical origins of unidentitied cadaver. Forensic Sci. Int. 2007, 173, 41–46. [Google Scholar] [CrossRef]
- Nishimoto, Y.; Zheng, H.Y.; Zhong, S.; Ikegaya, H.; Chen, Q.; Sugimoto, C.; Kitamura, T.; Yogo, Y. An Asian origin for subtype IV BK virus based on phylogenetic analysis. J. Mol. Evol. 2007, 65, 103–111. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Nishimoto, Y.; Chen, Q.; Hasegawa, M.; Zhong, S.; Ikegaya, H.; Ohno, N.; Sugimoto, C.; Takasaka, T.; Kitamura, T.; et al. Relationships between BK virus lineages and human populations. Microbes Infect. 2007, 9, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Randhawa, P.S.; Ikegaya, H.; Chen, Q.; Zheng, H.Y.; Suzuki, M.; Takeuchi, T.; Shibuya, A.; Kitamura, T.; Yogo, Y. Distribution patterns of BK polyomavirus (BKV) subtypes and subgroups in American, European and Asian populations suggest co-migration of BKV and the human race. J. Gen. Virol. 2009, 90, 144–152. [Google Scholar] [CrossRef]
- Wong, A.S.Y.; Cheng, V.C.C.; Yuen, K.Y.; Kwong, Y.L.; Leung, A.Y.H. High frequency of polyoma BK virus shedding in the gastrointestinal tract after hematopoietic stem cell transplantation: A prospective and quantitative analysis. Bone Marrow Transpl. 2009, 43, 43–47. [Google Scholar] [CrossRef]
- Reploeg, M.D.; Storch, G.A.; Clifford, D.B. BK virus: A clinical review. Clin. Infect. Dis. 2001, 33, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Weyandt, T.B.; Frisque, R.J. Identification of archetype and rearranged forms of BK virus in leukocytes from healthy individuals. J. Med. Virol. 2000, 60, 353–362. [Google Scholar] [CrossRef]
- Pezeshgi, A.; Ghods, A.; Keivani, H.; Asgari, M. Incidence of BK Virus Nephropathy (BKVN) in Renal Transplant Recipients. Int. J. Organ. Transpl. Med. 2012, 3, 115–118. [Google Scholar]
- Balba, G.P.; Javaid, B.; Timpone, J.G., Jr. BK Polyomavirus Infection in the Renal Transplant Recipient. Infect. Dis. Clin. N. Am. 2013, 27, 271–283. [Google Scholar] [CrossRef]
- Saade, A.; Styczynski, J.; Cesaro, S.; on behalf of Infectious Disease Working party of EBMT. BK virus infection in allogeneic hematopoietic cell transplantation: An update on pathogenesis, immune responses, diagnosis and treatments. J. Infect. 2020, 81, 372–382. [Google Scholar] [CrossRef]
- Mazzoni, E.; Pellegrinelli, E.; Mazziotta, C.; Lanzillotti, C.; Rotondo, J.C.; Bononi, I.; Iaquinta, M.R.; Manfrini, M.; Vesce, F.; Tognon, M.; et al. Mother-to-child transmission of oncogenic polyomaviruses BKPyV, JCPyV and SV40. J. Infect. 2020, 80, 563–570. [Google Scholar] [CrossRef]
- Geddes, C.C.; Gunson, R.; Mazonakis, E.; Wan, R.; Thomson, L.; Clancy, M.; Carman, W.F. BK viremia surveillance after kidney transplant: Single-center experience during a change from cyclosporine-to lower-dose tacrolimus-based primary immunosuppression regimen. Transpl. Infect. Dis. 2011, 13, 109–116. [Google Scholar] [CrossRef]
- Schwarz, A.; Linnenweber-Held, S.; Heim, A.; Framke, T.; Haller, H.; Schmitt, C. Viral Origin, Clinical Course, and Renal Outcomes in Patients with BK Virus Infection after Living-Donor Renal Transplantation. Transplantation 2016, 100, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.; Raggub, L.; Linnenweber-Held, S.; Adams, O.; Schwarz, A.; Heim, A. Donor origin of BKV replication after kidney transplantation. J. Clin. Virol. 2014, 59, 120–125. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Randhawa, P.S.; AST Infectious Diseases Community of Practice. BK polyomavirus in solid organ transplantation—Guidelines from the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transpl. 2019, 33, e13528. [Google Scholar] [CrossRef]
- Binggeli, S.; Egli, A.; Schaub, S.; Binet, I.; Mayr, M.; Steiger, J.; Hirsch, H.H. Polyomavirus BK-specific cellular immune response to VP1 and large T-antigen in kidney transplant recipients. Am. J. Transpl. 2007, 7, 1131–1139. [Google Scholar] [CrossRef]
- Comoli, P.; Binggeli, S.; Ginevri, F.; Hirsch, H.H. Polyomavirus-associated nephropathy: Update on BK virus-specific immunity. Transpl. Infect. Dis. 2006, 8, 86–94. [Google Scholar] [CrossRef]
- Lee, Y.J.; Zheng, J.; Kolitsopoulos, Y.; Chung, D.; Amigues, I.; Son, T.; Choo, K.; Hester, J.; Giralt, S.A.; Glezerman, I.G.; et al. Relationship of BK Polyoma Virus (BKV) in the Urine with Hemorrhagic Cystitis and Renal Function in Recipients of T Cell-Depleted Peripheral Blood and Cord Blood Stem Cell Transplantations. Biol. Blood Marrow Transpl. 2014, 20, 1204–1210. [Google Scholar] [CrossRef][Green Version]
- Najafabadi, M.M.; Soleimani, M.; Ahmadvand, M.; Zomorrod, M.S.; Mousavi, S.A. Treatment protocols for BK virus associated hemorrhagic cystitis after hematopoietic stem cell transplantation. Am. J. Blood Res. 2020, 10, 217–230. [Google Scholar]
- Umeda, K.; Kato, I.; Kawaguchi, K.; Tasaka, K.; Kamitori, T.; Ogata, H.; Mikami, T.; Hiramatsu, H.; Saito, R.; Ogawa, O.; et al. High incidence of BK virus-associated hemorrhagic cystitis in children after second or third allogeneic hematopoietic stem cell transplantation. Pediatr. Transpl. 2018, 22. [Google Scholar] [CrossRef]
- Dropulic, L.K.; Jones, R.J. Polyomavirus BK infection in blood and marrow transplant recipients. Bone Marrow Transpl. 2008, 41, 11–18. [Google Scholar] [CrossRef]
- Laskin, B.L.; Denburg, M.R.; Furth, S.L.; Moatz, T.; Altrich, M.; Kleiboeker, S.; Lutzko, C.; Zhu, X.; Blackard, J.T.; Jodele, S.; et al. The Natural History of BK Polyomavirus and the Host Immune Response after Stem Cell Transplantation. Clin. Infect. Dis. 2020, 71, 3044–3054. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, S.; Dalianis, T.; Rinaldo, C.H.; Koskenvuo, M.; Pegoraro, A.; Einsele, H.; Cordonnier, C.; Hirsch, H.H. ECIL guidelines for the prevention, diagnosis and treatment of BK polyomavirus-associated haemorrhagic cystitis in haematopoietic stem cell transplant recipients. J. Antimicrob. Chemother. 2018, 73, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Gilis, L.; Morisset, S.; Billaud, G.; Ducastelle-Lepretre, S.; Labussiere-Wallet, H.; Nicolini, F.E.; Barraco, F.; Detrait, M.; Thomas, X.; Tedone, N.; et al. High burden of BK virus-associated hemorrhagic cystitis in patients undergoing allogeneic hematopoietic stem cell transplantation. Bone Marrow Transpl. 2014, 49, 664–670. [Google Scholar] [CrossRef]
- Lunde, L.E.; Dasaraju, S.; Cao, Q.; Cohn, C.S.; Reding, M.; Bejanyan, N.; Trottier, B.; Rogosheske, J.; Brunstein, C.; Warlick, E.; et al. Hemorrhagic cystitis after allogeneic hematopoietic cell transplantation: Risk factors, graft source and survival. Bone Marrow Transpl. 2015, 50, 1432–1437. [Google Scholar] [CrossRef]
- Cesaro, S.; Tridello, G.; Pillon, M.; Calore, E.; Abate, D.; Tumino, M.; Carucci, N.; Varotto, S.; Cannata, E.; Pegoraro, A.; et al. A Prospective Study on the Predictive Value of Plasma BK Virus-DNA Load for Hemorrhagic Cystitis in Pediatric Patients after Stem Cell Transplantation. J. Pediatr. Infect. Dis. Soc. 2015, 4, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, S.; Facchin, C.; Tridello, G.; Messina, C.; Calore, E.; Biasolo, M.A.; Pillon, M.; Varotto, S.; Brugiolo, A.; Mengoli, C.; et al. A prospective study of BK-virus-associated haemorrhagic cystitis in paediatric patients undergoing allogeneic haematopoietic stem cell transplantation. Bone Marrow Transpl. 2008, 41, 363–370. [Google Scholar] [CrossRef]
- Leung, A.Y.H.; Chan, M.; Kwong, Y.L. Genotyping of the noncoding control region of BK virus in patients with haemorrhagic cystitis after allogeneic haematopoietic stem cell transplantation. Bone Marrow Transpl. 2005, 35, 531–532. [Google Scholar] [CrossRef][Green Version]
- Onda, Y.; Kanda, J.; Hanaoka, N.; Watanabe, M.; Arai, Y.; Hishizawa, M.; Kondo, T.; Yamashita, K.; Nagao, M.; Fujimoto, T.; et al. Possible nosocomial transmission of virus-associated hemorrhagic cystitis after allogeneic hematopoietic stem cell transplantation. Ann. Hematol. 2021, 100, 753–761. [Google Scholar] [CrossRef]
- Kato, J.; Mori, T.; Suzuki, T.; Ito, M.; Li, T.C.; Sakurai, M.; Yamane, Y.; Yamazaki, R.; Koda, Y.; Toyama, T.; et al. Nosocomial BK polyomavirus infection causing hemorrhagic cystitis among patients with hematological malignancies after hematopoietic stem cell transplantation. Am. J. Transpl. 2017, 17, 2428–2433. [Google Scholar] [CrossRef]
- Cheungpasitporn, W.; Thongprayoon, C.; Craici, I.M.; Sharma, K.; Chesdachai, S.; Khoury, N.J.; Ettore, A.S. Reactivation of BK polyomavirus during pregnancy, vertical transmission, and clinical significance: A meta-analysis. J. Clin. Virol. 2018, 102, 56–62. [Google Scholar] [CrossRef]
- Yin, W.Y.; Lee, M.C.; Lai, N.S.; Lu, M.C. BK virus as a potential oncovirus for bladder cancer in a renal transplant patient. J. Formos. Med. Assoc. 2015, 114, 373–374. [Google Scholar] [CrossRef]
- Starrett, G.J.; Buck, C.B. The case for BK polyomavirus as a cause of bladder cancer. Curr. Opin. Virol. 2019, 39, 8–15. [Google Scholar] [CrossRef]
- El-Mouallem, N.J.; Paul, A.K. BK Virus-Associated Urinary Bladder Cancer in a Kidney Transplant Recipient: A Case Report and Review of the Pathogenesis. Am. J. Hematol. 2017, 13, 15–20. [Google Scholar]
- Bertz, S.; Ensser, A.; Stoehr, R.; Eckstein, M.; Apel, H.; Mayr, D.; Buettner-Herold, M.; Gaisa, N.T.; Comperat, E.; Wullich, B.; et al. Variant morphology and random chromosomal integration of BK polyomavirus in posttransplant urothelial carcinomas. Mod. Pathol. 2020, 33, 1433–1442. [Google Scholar] [CrossRef]
- Lavien, G.; Alger, J.; Preece, J.; Alexiev, B.A.; Alexander, R.B. BK Virus-Associated Invasive Urothelial Carcinoma with Prominent Micropapillary Carcinoma Component in a Cardiac Transplant Patient: Case Report and Review of Literature. Clin. Genitourin. Cancer 2015, 13, e397–e399. [Google Scholar] [CrossRef]
- Jin, Y.; Zhou, Y.; Deng, W.; Wang, Y.; Lee, R.J.; Liu, Y.; Elias, N.; Hu, Y.; Luo, M.-H.; Liu, R.; et al. Genome-wide profiling of BK polyomavirus integration in bladder cancer of kidney transplant recipients reveals mechanisms of the integration at the nucleotide level. Oncogene 2021, 40, 46–54. [Google Scholar] [CrossRef]
- Kenan, D.J.; Mieczkowski, P.A.; Latulippe, E.; Cote, I.; Singh, H.K.; Nickeleit, V. BK polyomavirus genomic integration and large T antigen expression: Evolving paradigms in human oncogenesis. Am. J. Transpl. 2017, 17, 1674–1680. [Google Scholar] [CrossRef]
- Kenan, D.J.; Mieczkowski, P.A.; Burger-Calderon, R.; Singh, H.K.; Nickeleit, V. The oncogenic potential of BK-polyomavirus is linked to viral integration into the human genome. J. Pathol. 2015, 237, 379–389. [Google Scholar] [CrossRef]
- Muller, D.C.; Ramo, M.; Naegele, K.; Ribi, S.; Wetterauer, C.; Perrina, V.; Quagliata, L.; Vlajnic, T.; Ruiz, C.; Balitzki, B.; et al. Donor-derived, metastatic urothelial cancer after kidney transplantation associated with a potentially oncogenic BK polyomavirus. J. Pathol. 2018, 244, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Liu, Y.N.; Deng, W.F.; Fu, F.X.; Yan, S.S.; Yang, H.W.; Liu, R.; Geng, J.; Xu, J.; Wu, Y.; et al. Viral integration in BK polyomavirus-associated urothelial carcinoma in renal transplant recipients: Multistage carcinogenesis revealed by next-generation virome capture sequencing. Oncogene 2020, 39, 5734–5742. [Google Scholar] [CrossRef]
- Sawinski, D.; Trofe-Clark, J. BK Virus Nephropathy. Clin. J. Am. Soc. Nephrol. 2018, 13, 1893–1896. [Google Scholar] [CrossRef]
- Funahashi, Y. BK Virus-Associated Nephropathy after Renal Transplantation. Pathogens 2021, 10, 150. [Google Scholar] [CrossRef] [PubMed]
- Mullis, K.B. Target Amplification for DNA Analysis by the Polymerase Chain-Reaction. Ann. Biol. Clin. 1990, 48, 579–582. [Google Scholar]
- Sawinski, D.; Goral, S. BK virus infection: An update on diagnosis and treatment. Nephrol. Dial. Transpl. 2015, 30, 209–217. [Google Scholar] [CrossRef]
- Pastrana, D.V.; Ray, U.; Magaldi, T.G.; Schowalter, R.M.; Cuburu, N.; Buck, C.B. BK Polyomavirus Genotypes Represent Distinct Serotypes with Distinct Entry Tropism. J. Virol. 2013, 87, 10105–10113. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Lei, Y.; He, X.; Zhong, L.; Yu, X.; Huang, B. Sample Processing Effect on BK Virus Detection by Real-Time PCR in Urine Samples. Clin. Lab. 2016, 62, 833–837. [Google Scholar] [CrossRef]
- Hu, C.; Huang, Y.; Su, J.; Wang, M.; Zhou, Q.; Zhu, B. The prevalence and isolated subtypes of BK polyomavirus reactivation among patients infected with human immunodeficiency virus-1 in southeastern China. Arch. Virol. 2018, 163, 1463–1468. [Google Scholar] [CrossRef]
- Vaezjalali, M.; Azimi, H.; Hosseini, S.M.; Taghavi, A.; Goudarzi, H. Different Strains of BK Polyomavirus: VP1 Sequences in a Group of Iranian Prostate Cancer Patients. Urol. J. 2018, 15, 44–48. [Google Scholar]
- Moriyama, T.; Sorokin, A. BK virus (BKV): Infection, propagation, quantitation, purification, labeling, and analysis of cell entry. Curr. Protoc. Cell Biol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.; Medeiros, M.; Dadhania, D.; Muthukumar, T.; Kracker, D.; Kong, J.M.; Epstein, S.R.; Sharma, V.K.; Seshan, S.V.; Li, B.; et al. Noninvasive diagnosis of BK virus nephritis by measurement of messenger RNA for BK virus VP1 in urine. Transplantation 2002, 74, 987–994. [Google Scholar] [CrossRef]
- Mbianda, C.; El-Meanawy, A.; Sorokin, A. Mechanisms of BK virus infection of renal cells and therapeutic implications. J. Clin. Virol. 2015, 71, 59–62. [Google Scholar] [CrossRef]
- Boldorini, R.; Allegrini, S.; Miglio, U.; Paganotti, A.; Veggiani, C.; Mischitelli, M.; Monga, G.; Pietropaolo, V. Genomic Mutations of Viral Protein 1 and BK Virus Nephropathy in Kidney Transplant Recipients. J. Med. Virol. 2009, 81, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
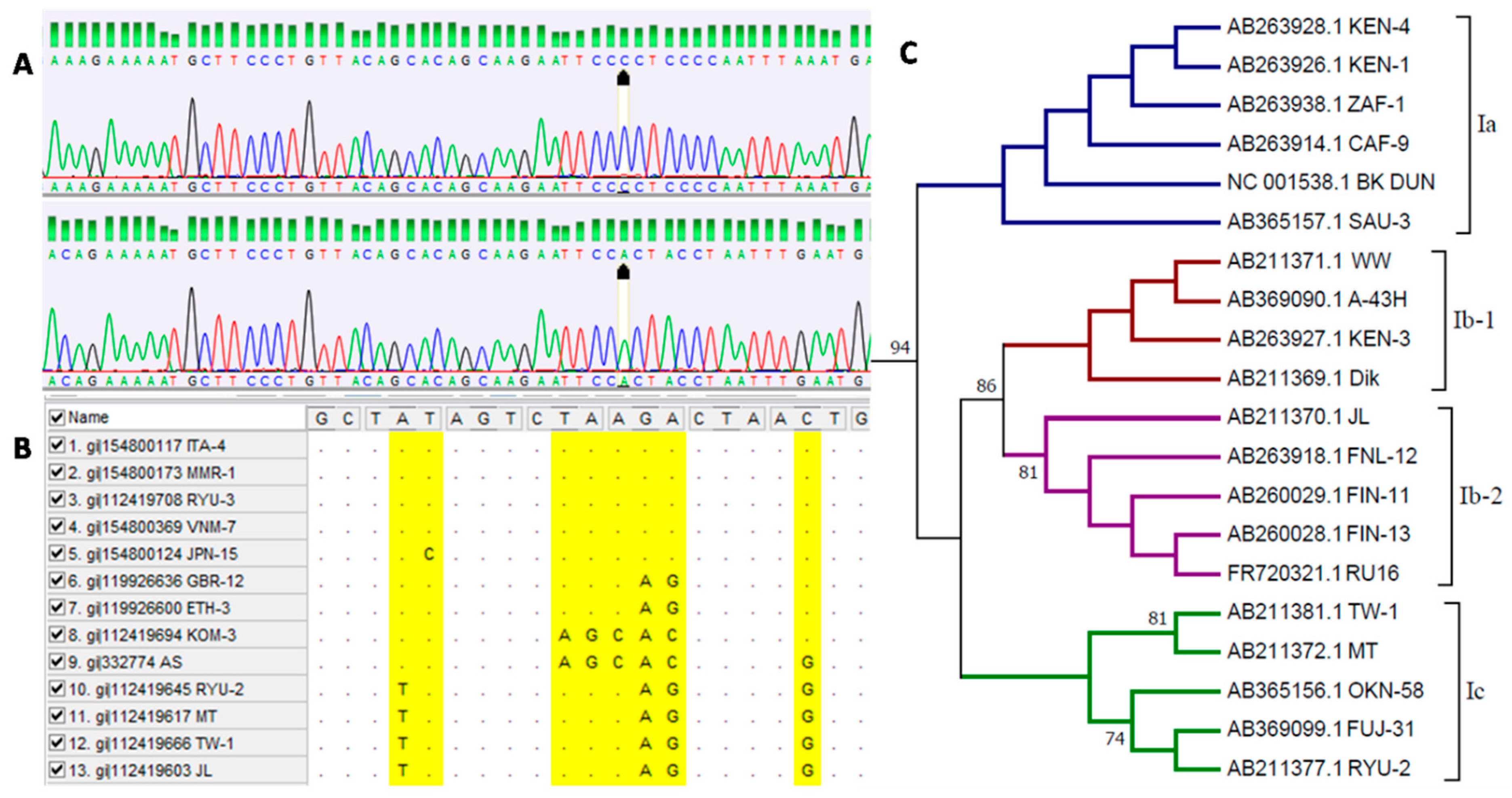
- Luo, C.; Bueno, M.; Kant, J.; Martinson, J.; Randhawa, P. Genotyping Schemes for Polyomavirus BK, Using Gene-Specific Phylogenetic Trees and Single Nucleotide Polymorphism Analysis. J. Virol. 2009, 83, 2285–2297. [Google Scholar] [CrossRef][Green Version]
- Seif, I.; Khoury, G.; Dhar, R. Genome of Human Papovavirus BKV. Cell 1979, 18, 963–977. [Google Scholar] [CrossRef]
- Carpenter, M.L.; Tan, S.K.; Watson, T.; Bacher, R.; Nagesh, V.; Watts, A.; Bentley, G.; Weber, J.; Huang, C.; Sahoo, M.K.; et al. Metagenomic next-generation sequencing for identification and quantitation of transplant-related DNA viruses. J. Clin. Microbiol. 2019, 57, e01113-19. [Google Scholar] [CrossRef]
- Luo, C.Q.; Hirsch, H.H.; Kant, J.; Randhawa, P. VP-1 Quasispecies in Human Infection with Polyomavirus BK. J. Med. Virol. 2012, 84, 152–161. [Google Scholar] [CrossRef]
- Varella, R.B.; Zalona, A.C.J.; Diaz, N.C.; Zalis, M.G.; Santoro-Lopes, G. BK polyomavirus genotypes Ia and Ib1 exhibit different biological properties in renal transplant recipients. Virus Res. 2018, 243, 65–68. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Ikegaya, H.; Takasaka, T.; Matsushima-Ohno, T.; Sakurai, M.; Kanazawa, I.; Kishida, S.; Nagashima, K.; Kitamura, T.; Yogo, Y. Characterization of the VP1 loop mutations widespread among JC polyomavirus isolates associated with progressive multifocal leukoencephalopathy. Biochem. Biophys. Res. Commun. 2005, 333, 996–1002. [Google Scholar] [CrossRef]
- McIlroy, D.; Halary, F.; Bressollette-Bodin, C. Intra-patient viral evolution in polyomavirus-related diseases. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374. [Google Scholar] [CrossRef]
- Scribano, S.; Guerrini, M.; Arvia, R.; Guasti, D.; Nardini, P.; Romagnoli, P.; Giannecchini, S. Archetype JC polyomavirus DNA associated with extracellular vesicles circulates in human plasma samples. J. Clin. Virol. 2020, 128, 104435. [Google Scholar] [CrossRef]
- Randhawa, P.S.; Khaleel-Ur-Rehman, K.; Swalsky, P.A.; Vats, A.; Scantlebury, V.; Shapiro, R.; Finkelstein, S. DNA sequencing of viral capsid protein VP-1 region in patients with BK virus interstitial nephritis. Transplantation 2002, 73, 1090–1094. [Google Scholar] [CrossRef]
- McIlroy, D.; Honemann, M.; Nguyen, N.K.; Barbier, P.; Peltier, C.; Rodallec, A.; Halary, F.; Przyrowski, E.; Liebert, U.; Hourmant, M.; et al. Persistent BK Polyomavirus Viruria is Associated with Accumulation of VP1 Mutations and Neutralization Escape. Viruses 2020, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Verhalen, B.; Starrett, G.J.; Harris, R.S.; Jiang, M. Functional Upregulation of the DNA Cytosine Deaminase APOBEC3B by Polyomaviruses. J. Virol. 2016, 90, 6379–6386. [Google Scholar] [CrossRef] [PubMed]
- Peretti, A.; Geoghegan, E.M.; Pastrana, D.V.; Smola, S.; Feld, P.; Sauter, M.; Lohse, S.; Ramesh, M.; Lim, E.S.; Wang, D.; et al. Characterization of BK Polyomaviruses from Kidney Transplant Recipients Suggests a Role for APOBEC3 in Driving In-Host Virus Evolution. Cell Host Microbe 2018, 23, 628–635.e7. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variable Region | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype/Subtype | Accession Number | 61 | 62 | 66 | 68 | 69 | 71 | 73 | 74 | 75 | 77 | 82 |
| Ia | NC_001538.1_BK_DUN | E | N | F | L | K | S | E | N | D | S | E |
| AB263926.1_KEN-1 | . | . | . | . | . | . | . | . | . | . | . | |
| Ib-1 | AB211371.1_WW | . | . | . | . | . | . | K | . | . | . | . |
| AB211369.1_Dik | . | . | . | . | . | . | . | . | . | . | . | |
| Ib-2 | AB260029.1_FIN-11 | . | . | . | . | . | . | . | . | . | . | D |
| AB260028.1_FIN-13 | . | . | . | . | . | . | . | . | . | . | D | |
| AB211370.1_JL | . | . | . | . | . | . | . | . | . | . | . | |
| Ic | AB211381.1_TW-1 | . | . | . | . | . | . | . | . | . | . | . |
| AB211377.1_RYU-2 | . | . | . | . | . | . | . | . | . | . | Q | |
| AB211372.1_MT | . | . | . | . | . | . | . | . | . | . | . | |
| II | AB263916.1_ETH-3 | . | . | Y | . | . | T | . | . | A | D | D |
| JN793996.1 KT40 | D | . | Y | . | . | T | . | . | A | D | D | |
| AB301101.1 J2B-11 | D | . | Y | . | . | T | . | . | A | D | D | |
| III | M23122.1_AS | D | . | Y | Q | H | . | . | . | A | E | D |
| JN192440.1 SJH-LG-310 | . | . | Y | Q | H | . | . | . | A | E | D | |
| AB365139.1 NEA-27 | D | H | Y | Q | H | T | . | . | A | D | D | |
| AB365130.1 FUK-22 | D | H | Y | Q | H | T | . | . | A | D | D | |
| IV | AB211388.1_KOM-7 | N | D | Y | . | R | T | . | T | A | D | D |
| AB211387.1_KOM-2 | N | D | Y | . | R | T | . | T | A | D | D | |
| AB211391.1_TW-3 | N | D | Y | . | R | T | . | T | A | D | D | |
| AB211390.1_THK-8 | N | D | Y | . | R | T | . | T | A | N | D | |
| AB211389.1_RYU-3 | N | D | Y | . | R | T | . | T | A | D | D | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furmaga, J.; Kowalczyk, M.; Zapolski, T.; Furmaga, O.; Krakowski, L.; Rudzki, G.; Jaroszyński, A.; Jakubczak, A. BK Polyomavirus—Biology, Genomic Variation and Diagnosis. Viruses 2021, 13, 1502. https://doi.org/10.3390/v13081502
Furmaga J, Kowalczyk M, Zapolski T, Furmaga O, Krakowski L, Rudzki G, Jaroszyński A, Jakubczak A. BK Polyomavirus—Biology, Genomic Variation and Diagnosis. Viruses. 2021; 13(8):1502. https://doi.org/10.3390/v13081502
Chicago/Turabian StyleFurmaga, Jacek, Marek Kowalczyk, Tomasz Zapolski, Olga Furmaga, Leszek Krakowski, Grzegorz Rudzki, Andrzej Jaroszyński, and Andrzej Jakubczak. 2021. "BK Polyomavirus—Biology, Genomic Variation and Diagnosis" Viruses 13, no. 8: 1502. https://doi.org/10.3390/v13081502
APA StyleFurmaga, J., Kowalczyk, M., Zapolski, T., Furmaga, O., Krakowski, L., Rudzki, G., Jaroszyński, A., & Jakubczak, A. (2021). BK Polyomavirus—Biology, Genomic Variation and Diagnosis. Viruses, 13(8), 1502. https://doi.org/10.3390/v13081502