
HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Specimens
2.2. Cell Lines and Culture
2.3. RNA Extraction and PCR Amplification of HERV Transcripts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Amplicon Size | Primers | Sequence (5′–3′) | Target Region or Reference 1 | Predicted Targets 2 |
|---|---|---|---|---|---|
| HERV-K Gag | 113 bp | Fwd | TTCCCGAGTACGTCTACAGTGA | [19] | 1 |
| Rev | GGTGTTTCTCATCAGGTGGAAT | ||||
| HERV-K Np9 | 113 bp | Fwd | GTTAGTCTACAGGTGTATCCA | GenBank Y17832.2 8836–8948 | 1 |
| Rev | CTGTCTCTTTTCCCTACATTTCC | ||||
| HERV-W Gag | 81 bp | Fwd | TCAGGTCAACAATAGGATGACAACA | [21] | 1 |
| Rev | CAATGAGGGTCTACACTGGGAACT | ||||
| HERV-E Pol | 98 bp | Fwd | CTCTACACAGTTAGGCTCG | Chr13 40,349,095–40,349,192 | 1 |
| Rev | GTGAAAATCCCCGCATAACC | ||||
| HERV_E Env | 168 bp | Fwd | ATTTGATGCTTGTGCAGCCA | [22] | 2 3 |
| Rev | TTCTTTTTCCAAGTAGCCCAAAT | ||||
| HERV_H Pol | 118 bp | Fwd | GCCAAACACATATACTCTCC | Chr8 8,066,673–8,066,790 | 1 |
| Rev | GAAGAGTGACTGGGATGAAG |
2.4. Detection of HERV-K Gag Protein by Immunohistochemistry
3. Results
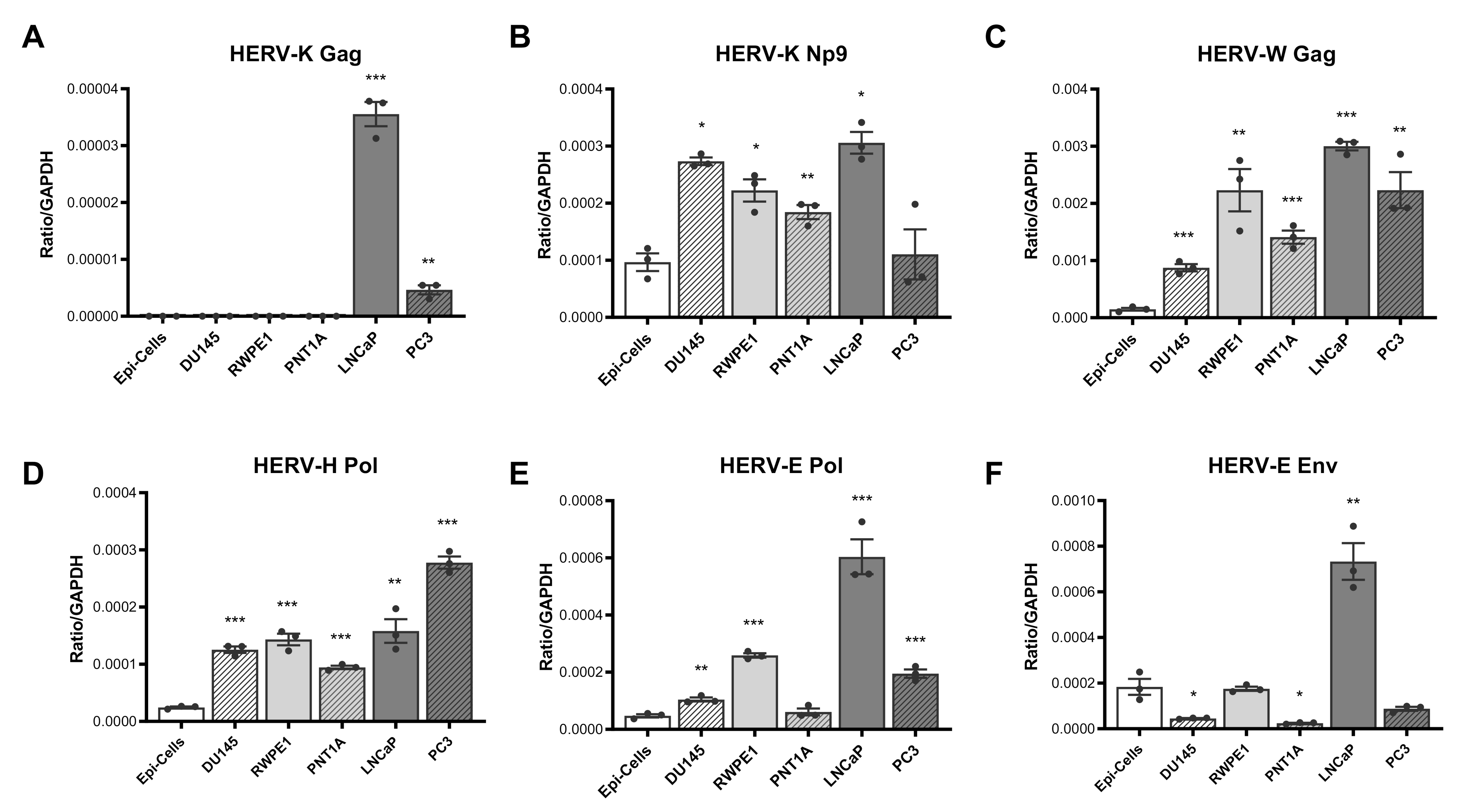
3.1. Expression of HERV Transcripts Is Significantly Upregulated in Prostate Cancer Cell Lines as Compared to Normal Epithelial Cells
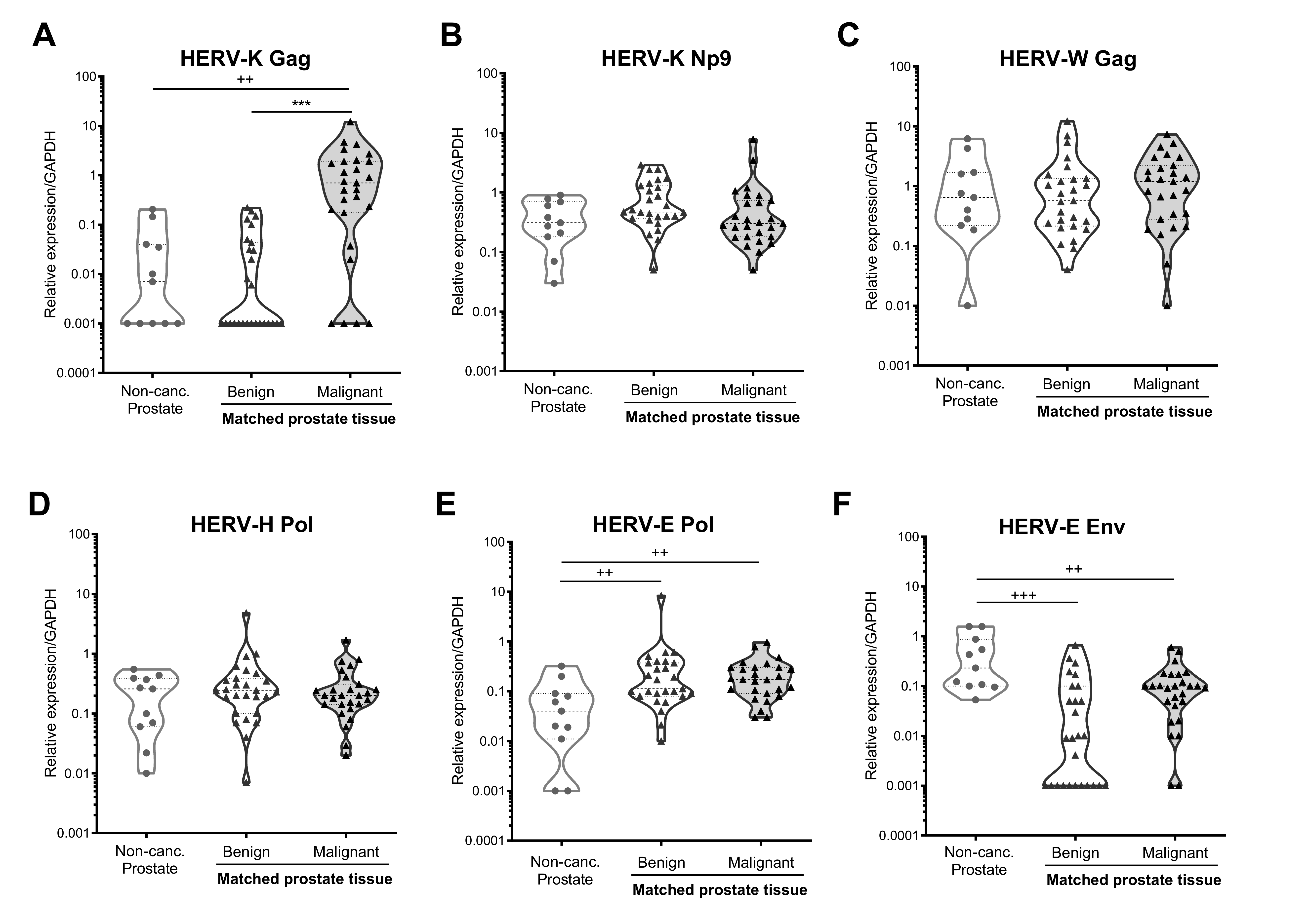
3.2. HERV-K Gag Expression Is Significantly Upregulated in Malignant, but Not Benign, Regions of Primary Prostate Tissue
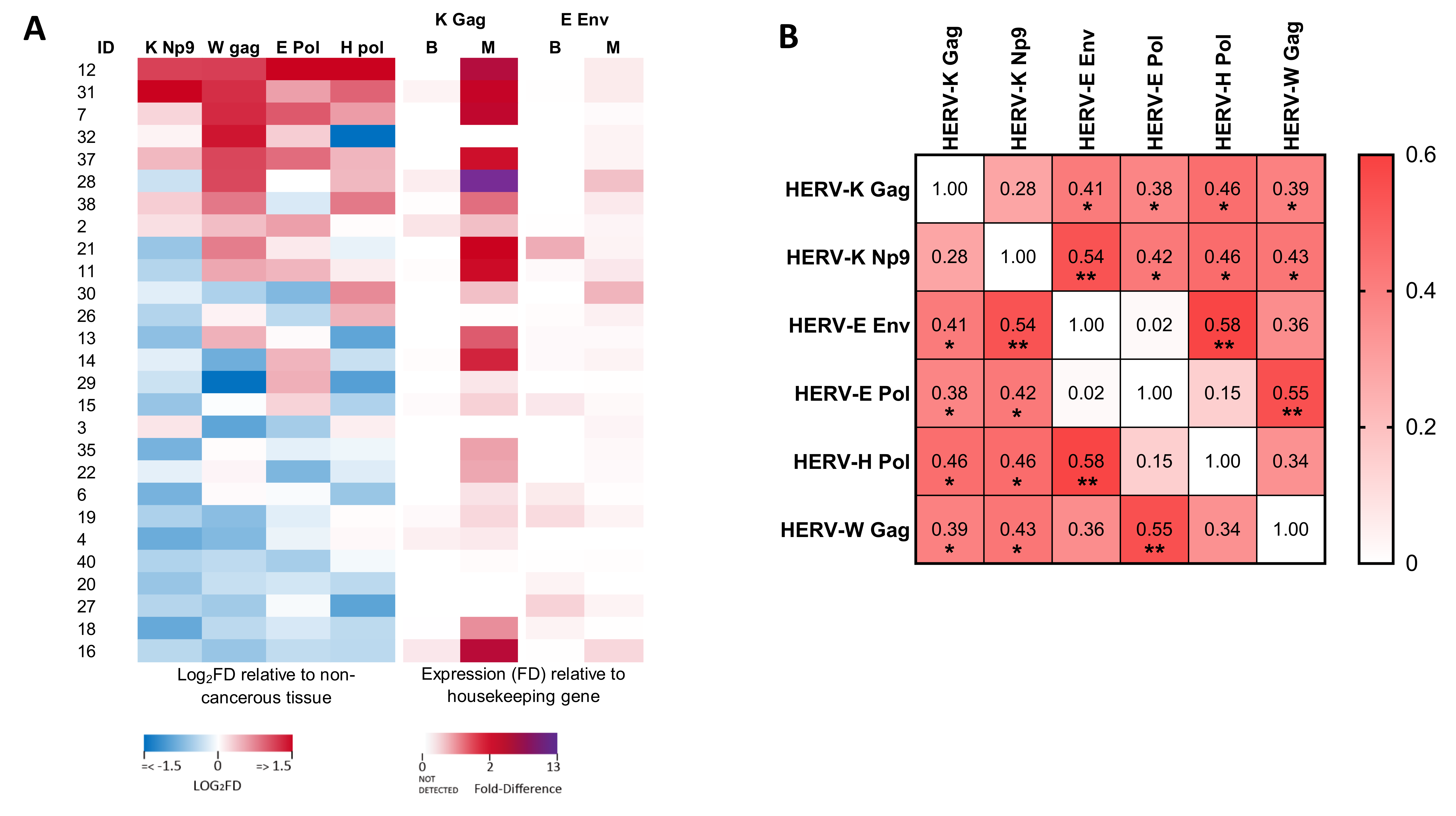
3.3. Analysis of the Relationship between the Expression of Different HERV Transcripts in Malignant Regions of the Prostate
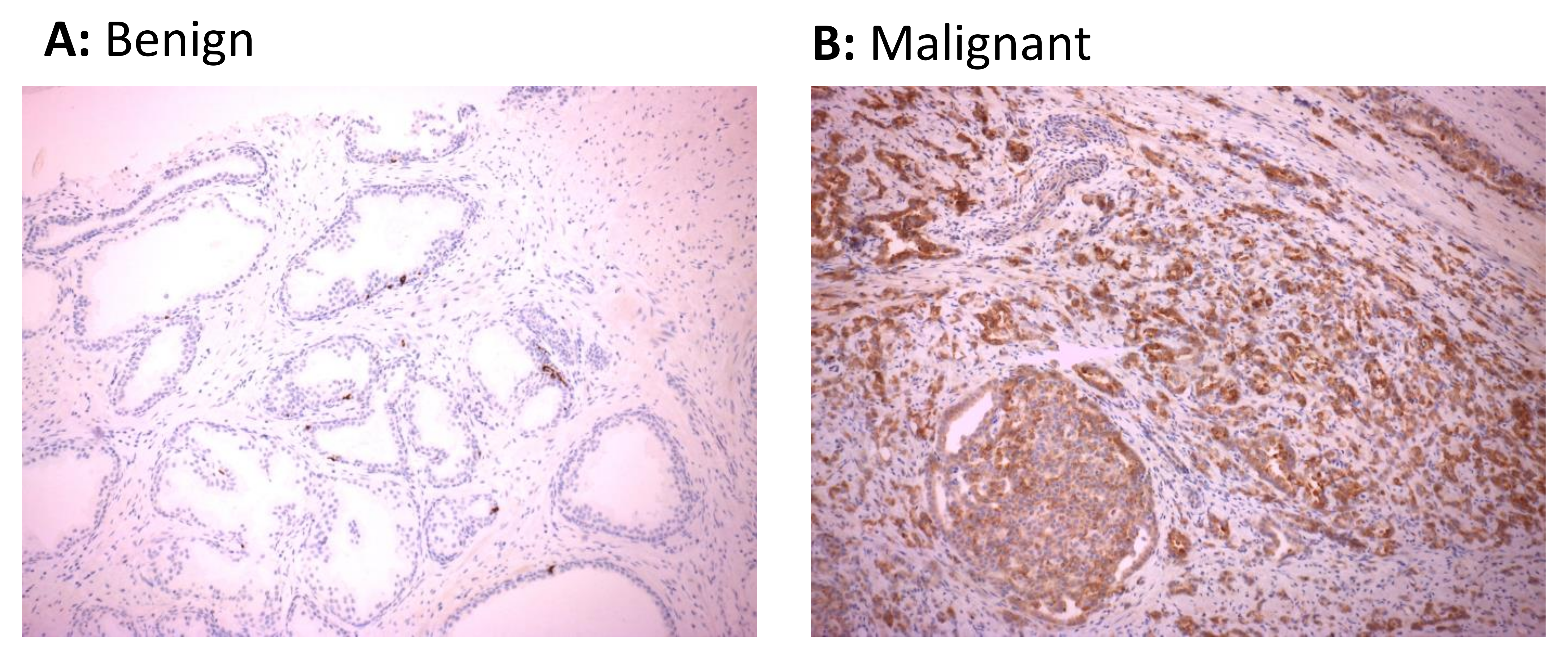
3.4. HERV-K Gag Protein Is Detected Primarily in Malignant Regions of the Prostate in Men with Prostate Cancer
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Australian Institute of Health and Welfare. Cancer Data in Australia. Available online: https://www.aihw.gov.au/reports/cancer/cancer-data-in-australia/contents/cancer-summary-data-visualisation (accessed on 12 February 2021).
- Prostate Cancer Foundation of Australia. Prostate Cancer in Australia—What Do the Numbers Tell Us? Available online: https://www.prostate.org.au/news-media/news/prostate-cancer-in-australia-what-do-the-numbers-tell-us/ (accessed on 12 February 2021).
- Glasziou, P.P.; Jones, M.A.; Pathirana, T.; Barratt, A.L.; Bell, K.J. Estimating the magnitude of cancer overdiagnosis in Australia. Med. J. Aust. 2020, 212, 163–168. [Google Scholar] [CrossRef]
- Geis, F.K.; Goff, S.P. Silencing and transcriptional regulation of endogenous retroviruses: An overview. Viruses 2020, 12, 884. [Google Scholar] [CrossRef] [PubMed]
- Buscher, K.; Hahn, S.; Hofmann, M.; Trefzer, U.; Ozel, M.; Sterry, W.; Lower, J.; Lower, R.; Kurth, R.; Denner, J. Expression of the human endogenous retrovirus-K transmembrane envelope, Rec and Np9 proteins in melanomas and melanoma cell lines. Melanoma Res. 2006, 16, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Schulte, A.M.; Lai, S.; Kurtz, A.; Czubayko, F.; Riegel, A.T.; Wellstein, A. Human trophoblast and choriocarcinoma expression of the growth factor pleiotrophin attributable to germ-line insertion of an endogenous retrovirus. Proc. Natl. Acad. Sci. USA 1996, 93, 14759–14764. [Google Scholar] [CrossRef] [PubMed]
- Jern, P.; Coffin, J.M. Effects of retroviruses on host genome function. Ann. Rev. Genet. 2008, 42, 709–732. [Google Scholar] [CrossRef]
- Grabski, D.F.; Hu, Y.; Sharma, M.; Rasmussen, S.K. Close to the bedside: A systematic review of endogenous retroviruses and their impact in oncology. J. Surg. Res. 2019, 240, 145–155. [Google Scholar] [CrossRef]
- Levet, S.; Charvet, B.; Bertin, A.; Deschaumes, A.; Perron, H.; Hober, D. Human endogenous retroviruses and type 1 diabetes. Curr. Diab. Rep. 2019, 19, 141. [Google Scholar] [CrossRef]
- Wang-Johanning, F.; Radvanyi, L.; Rycaj, K.; Plummer, J.B.; Yan, P.; Sastry, K.J.; Piyathilake, C.J.; Hunt, K.K.; Johanning, G.L. Human endogenous retrovirus K triggers an antigen-specific immune response in breast cancer patients. Cancer Res. 2008, 68, 5869–5877. [Google Scholar] [CrossRef] [PubMed]
- Rycaj, K.; Plummer, J.B.; Yin, B.; Li, M.; Garza, J.; Radvanyi, L.; Ramondetta, L.M.; Lin, K.; Johanning, G.L.; Tang, D.G.; et al. Cytotoxicity of human endogenous retrovirus K-specific T cells toward autologous ovarian cancer cells. Clin. Cancer Res. 2015, 21, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Hong, Z.; Liu, H.; Chen, X.; Ding, L.; Liu, Z.; Zhou, F.; Yuan, Y. Human endogenous retroviruses-K (HML-2) expression is correlated with prognosis and progress of hepatocellular carcinoma. Biomed. Res. Int. 2016, 2016, 8201642. [Google Scholar] [CrossRef]
- Wang-Johanning, F.; Li, M.; Esteva, F.J.; Hess, K.R.; Yin, B.; Rycaj, K.; Plummer, J.B.; Garza, J.G.; Ambs, S.; Johanning, G.L. Human endogenous retrovirus type K antibodies and mRNA as serum biomarkers of early-stage breast cancer. Int. J. Cancer 2014, 134, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Golan, M.; Hizi, A.; Resau, J.H.; Yaal-Hahoshen, N.; Reichman, H.; Keydar, I.; Tsarfaty, I. Human endogenous retrovirus (HERV-K) reverse transcriptase as a breast cancer prognostic marker. Neoplasia 2008, 10, 521–533. [Google Scholar] [CrossRef]
- Reis, B.S.; Jungbluth, A.A.; Frosina, D.; Holz, M.; Ritter, E.; Nakayama, E.; Ishida, T.; Obata, Y.; Carver, B.; Scher, H.; et al. Prostate cancer progression correlates with increased humoral immune response to a human endogenous retrovirus GAG protein. Clin. Cancer Res. 2013, 19, 6112–6125. [Google Scholar] [CrossRef]
- Ishida, T.; Obata, Y.; Ohara, N.; Matsushita, H.; Sato, S.; Uenaka, A.; Saika, T.; Miyamura, T.; Chayama, K.; Nakamura, Y.; et al. Identification of the HERV-K gag antigen in prostate cancer by SEREX using autologous patient serum and its immunogenicity. Cancer Immun. 2008, 8, 15. [Google Scholar]
- Wang-Johanning, F.; Frost, A.R.; Jian, B.; Azerou, R.; Lu, D.W.; Chen, D.T.; Johanning, G.L. Detecting the expression of human endogenous retrovirus E envelope transcripts in human prostate adenocarcinoma. Cancer 2003, 98, 187–197. [Google Scholar] [CrossRef]
- Wallace, T.A.; Downey, R.F.; Seufert, C.J.; Schetter, A.; Dorsey, T.H.; Johnson, C.A.; Goldman, R.; Loffredo, C.A.; Yan, P.; Sullivan, F.J.; et al. Elevated HERV-K mRNA expression in PBMC is associated with a prostate cancer diagnosis particularly in older men and smokers. Carcinogenesis 2014, 35, 2074–2083. [Google Scholar] [CrossRef]
- Goering, W.; Ribarska, T.; Schulz, W.A. Selective changes of retroelement expression in human prostate cancer. Carcinogenesis 2011, 32, 1484–1492. [Google Scholar] [CrossRef]
- Wang, K.; Li, H.; Xu, Y.; Shao, Q.; Yi, J.; Wang, R.; Cai, W.; Hang, X.; Zhang, C.; Cai, H.; et al. MFEprimer-3.0: Quality control for PCR primers. Nucleic Acids Res. 2019, 47, W610–W613. [Google Scholar] [CrossRef]
- Nellaker, C.; Yao, Y.; Jones-Brando, L.; Mallet, F.; Yolken, R.H.; Karlsson, H. Transactivation of elements in the human endogenous retrovirus W family by viral infection. Retrovirology 2006, 3, 44. [Google Scholar] [CrossRef]
- Andersson, A.C.; Yun, Z.; Sperber, G.O.; Larsson, E.; Blomberg, J. ERV3 and related sequences in humans: Structure and RNA expression. J. Virol. 2005, 79, 9270–9284. [Google Scholar] [CrossRef] [PubMed]
- Agoni, L.; Guha, C.; Lenz, J. Detection of human endogenous retrovirus K (HERV-K) transcripts in human prostate cancer cell lines. Front. Oncol. 2013, 3, 180. [Google Scholar] [CrossRef]
- Curty, G.; Marston, J.L.; de Mulder Rougvie, M.; Leal, F.E.; Nixon, D.F.; Soares, M.A. Human endogenous retrovirus K in cancer: A potential biomarker and immunotherapeutic target. Viruses 2020, 12, 726. [Google Scholar] [CrossRef] [PubMed]
- Denne, M.; Sauter, M.; Armbruester, V.; Licht, J.D.; Roemer, K.; Mueller-Lantzsch, N. Physical and functional interactions of human endogenous retrovirus proteins Np9 and rec with the promyelocytic leukemia zinc finger protein. J. Virol. 2007, 81, 5607–5616. [Google Scholar] [CrossRef]
- Kudo-Saito, C.; Yura, M.; Yamamoto, R.; Kawakami, Y. Induction of immunoregulatory CD271+ cells by metastatic tumor cells that express human endogenous retrovirus H. Cancer Res. 2014, 74, 1361–1370. [Google Scholar] [CrossRef]
- Zhao, J.; Rycaj, K.; Geng, S.; Li, M.; Plummer, J.B.; Yin, B.; Liu, H.; Xu, X.; Zhang, Y.; Yan, Y.; et al. Expression of human endogenous retrovirus type K envelope protein is a novel candidate prognostic marker for human breast cancer. Genes Cancer 2011, 2, 914–922. [Google Scholar] [CrossRef]
- Hahn, S.; Ugurel, S.; Hanschmann, K.M.; Strobel, H.; Tondera, C.; Schadendorf, D.; Lower, J.; Lower, R. Serological response to human endogenous retrovirus K in melanoma patients correlates with survival probability. AIDS Res. Hum. Retrovir. 2008, 24, 717–723. [Google Scholar] [CrossRef]
- Kleiman, A.; Senyuta, N.; Tryakin, A.; Sauter, M.; Karseladze, A.; Tjulandin, S.; Gurtsevitch, V.; Mueller-Lantzsch, N. HERV-K(HML-2) GAG/ENV antibodies as indicator for therapy effect in patients with germ cell tumors. Int. J. Cancer 2004, 110, 459–461. [Google Scholar] [CrossRef]




| Sample ID | Age | Gleason Score | HERV K Gag Detected by IHC 1 | HERV K Gag Detected by qRT-PCR 4 | ||
|---|---|---|---|---|---|---|
| Benign | Cancerous | Benign | Cancerous | |||
| 71 | 59 | 7 | − | +/++ | ND | 12.3 |
| 72 | 66 | 7 | − | +/++ 2 | ND | 1.2 |
| 74 | 53 | 7 | − | − | ND | ND |
| 75 | 65 | 7 | −3 | − | ND | ND |
| 76 | 62 | 7 | − | + | ND | ND |
| 77 | 47 | 8 | − | − | 0.15 | 12.1 |
| 78 | 61 | 6 | −3 | + | ND | 0.2 |
| 79 | 69 | 7 | − | ++/+++ | ND | 0.5 |
| 80 | 65 | 7 | − | − | ND | 2.7 |
| 83 | 54 | 7 | − | +/+++ | ND | ND |
| 84 | 65 | 7 | − | − 3 | 0.4 | 1.2 |
| 85 | 66 | 7 | − | +/++ | ND | 10.1 |
| 86 | 65 | 8 | − | − | ND | 0.75 |
| 87 | 69 | 7 | −3 | + | 0.01 | 4.89 |
| 88 | 70 | 9 | − | ++/+++ | ND | 1.9 |
| 94 | 66 | 8 | +2 | + | 0.02 | 1.15 |
| 97 | 65 | 8 | − | ++/+++ | ND | ND |
| 98 | 64 | 7 | − | ++ | ND | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rezaei, S.D.; Hayward, J.A.; Norden, S.; Pedersen, J.; Mills, J.; Hearps, A.C.; Tachedjian, G. HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer. Viruses 2021, 13, 449. https://doi.org/10.3390/v13030449
Rezaei SD, Hayward JA, Norden S, Pedersen J, Mills J, Hearps AC, Tachedjian G. HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer. Viruses. 2021; 13(3):449. https://doi.org/10.3390/v13030449
Chicago/Turabian StyleRezaei, Simin D., Joshua A. Hayward, Sam Norden, John Pedersen, John Mills, Anna C. Hearps, and Gilda Tachedjian. 2021. "HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer" Viruses 13, no. 3: 449. https://doi.org/10.3390/v13030449
APA StyleRezaei, S. D., Hayward, J. A., Norden, S., Pedersen, J., Mills, J., Hearps, A. C., & Tachedjian, G. (2021). HERV-K Gag RNA and Protein Levels Are Elevated in Malignant Regions of the Prostate in Males with Prostate Cancer. Viruses, 13(3), 449. https://doi.org/10.3390/v13030449