
Molecular Characterization of Imported and Autochthonous Dengue in Northeastern Spain

 ,
,  , , , add
Show full author list
, , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Nucleic Acid Extraction, DENV Genome Detection and DENV Serotyping
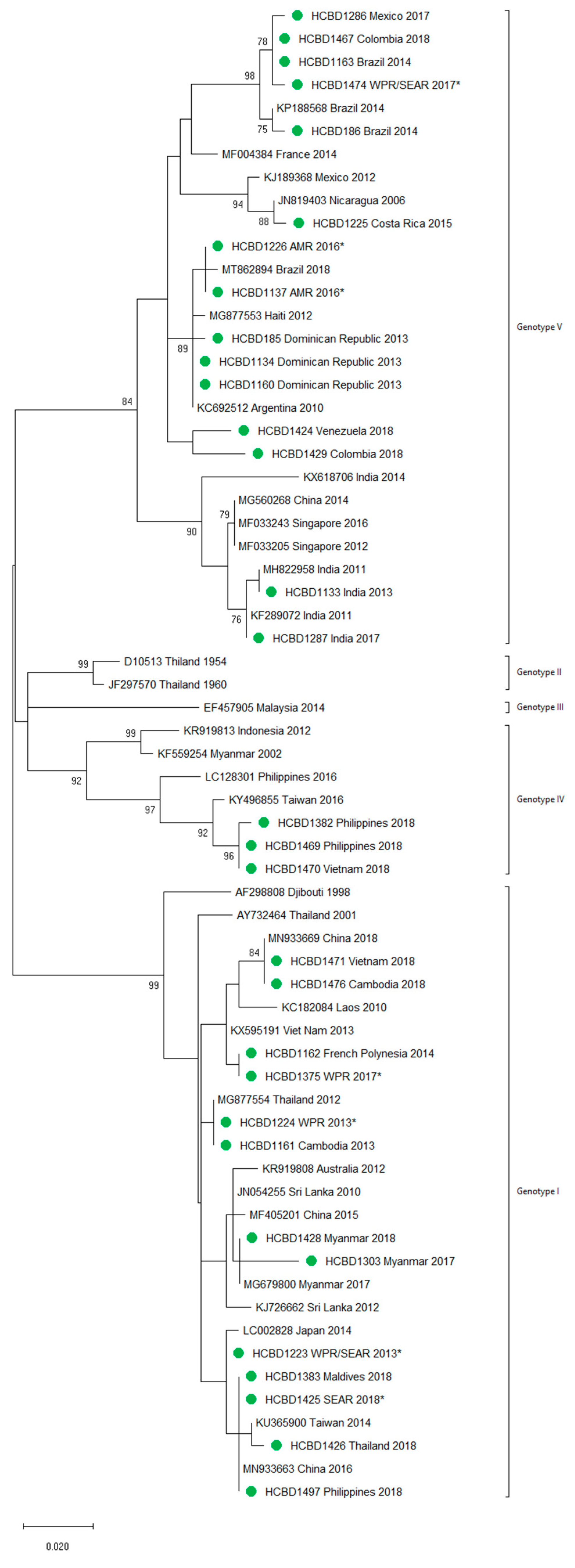
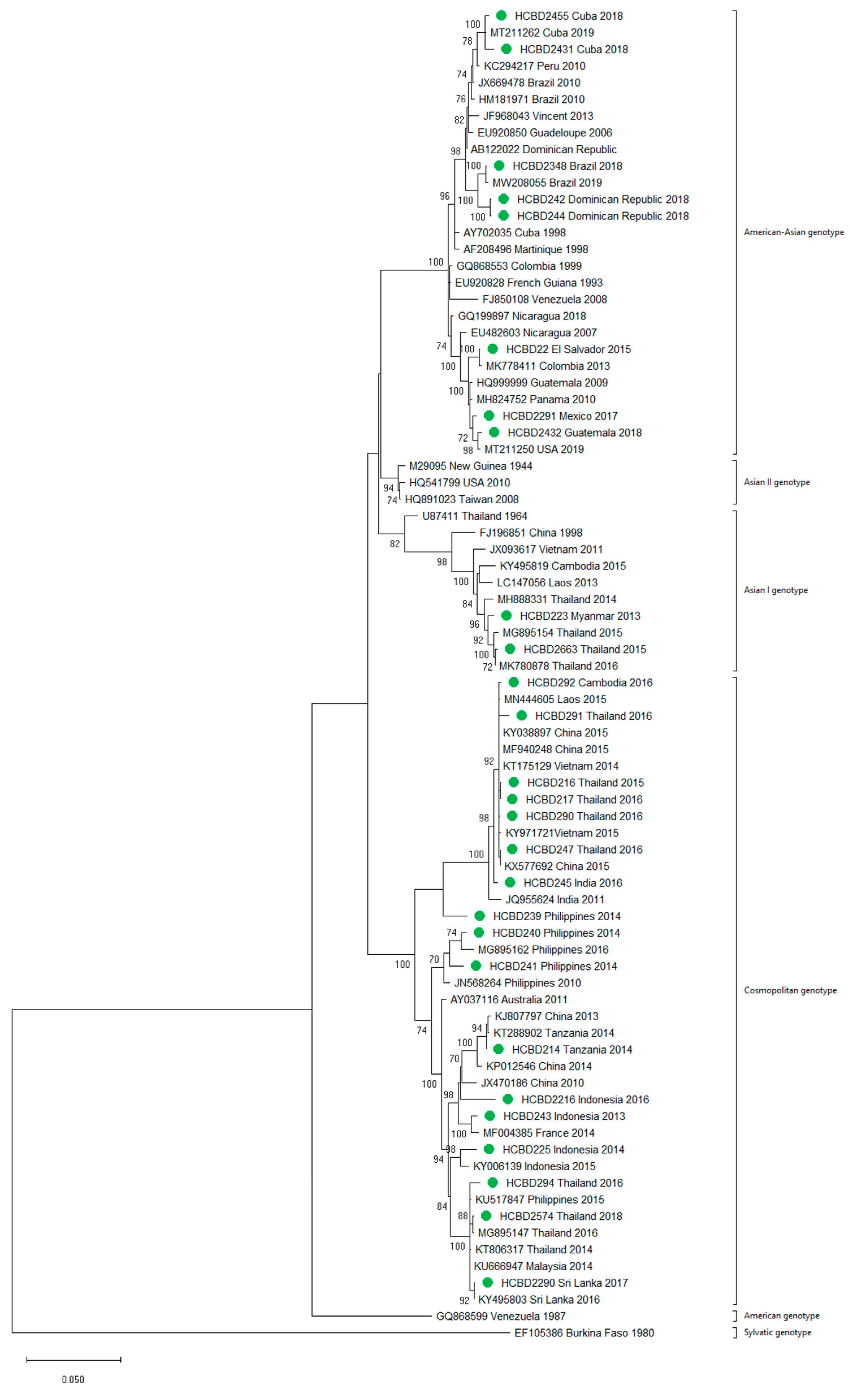
2.3. Genotyping of DENV and Phylogenetic Analysis
3. Results
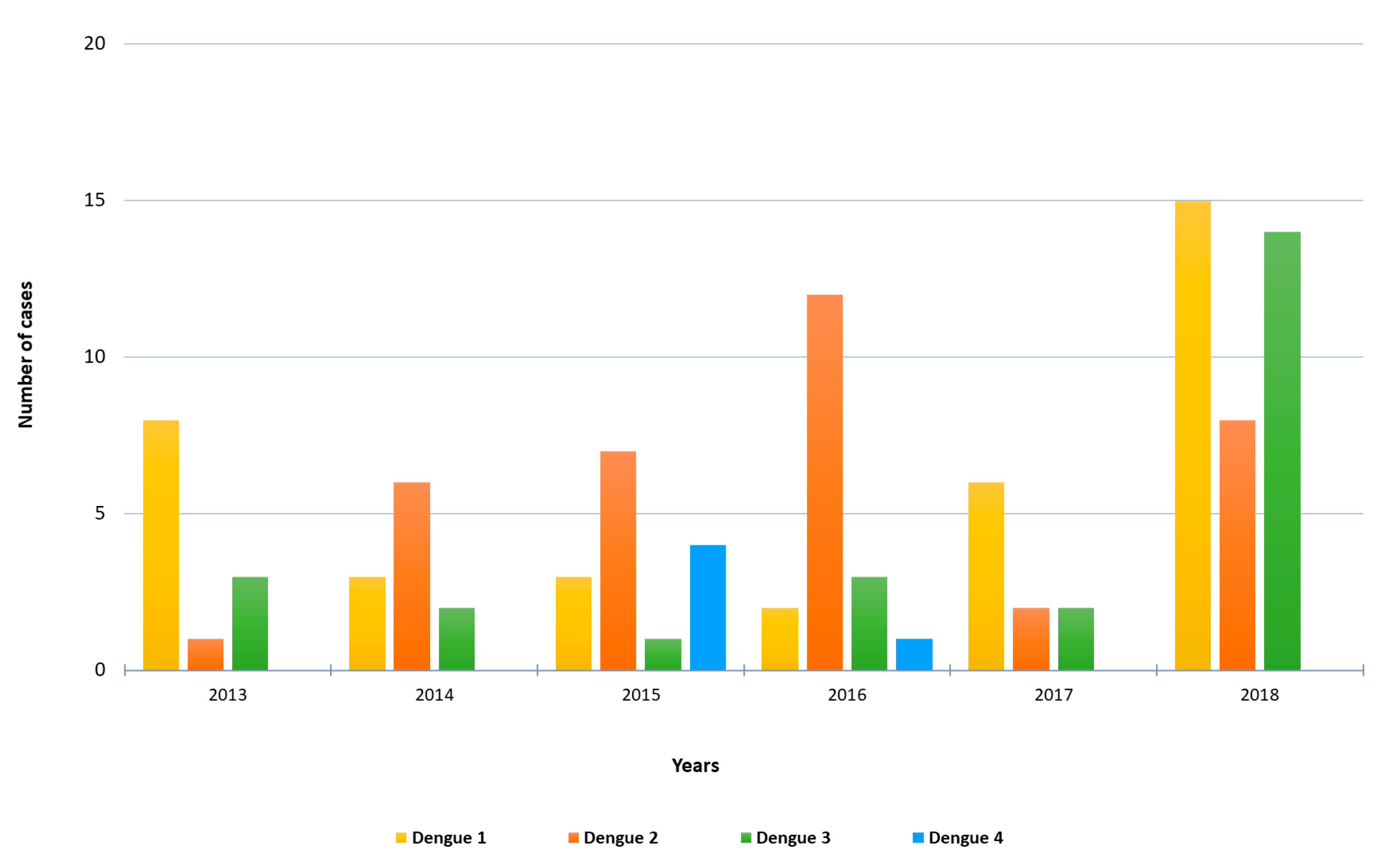
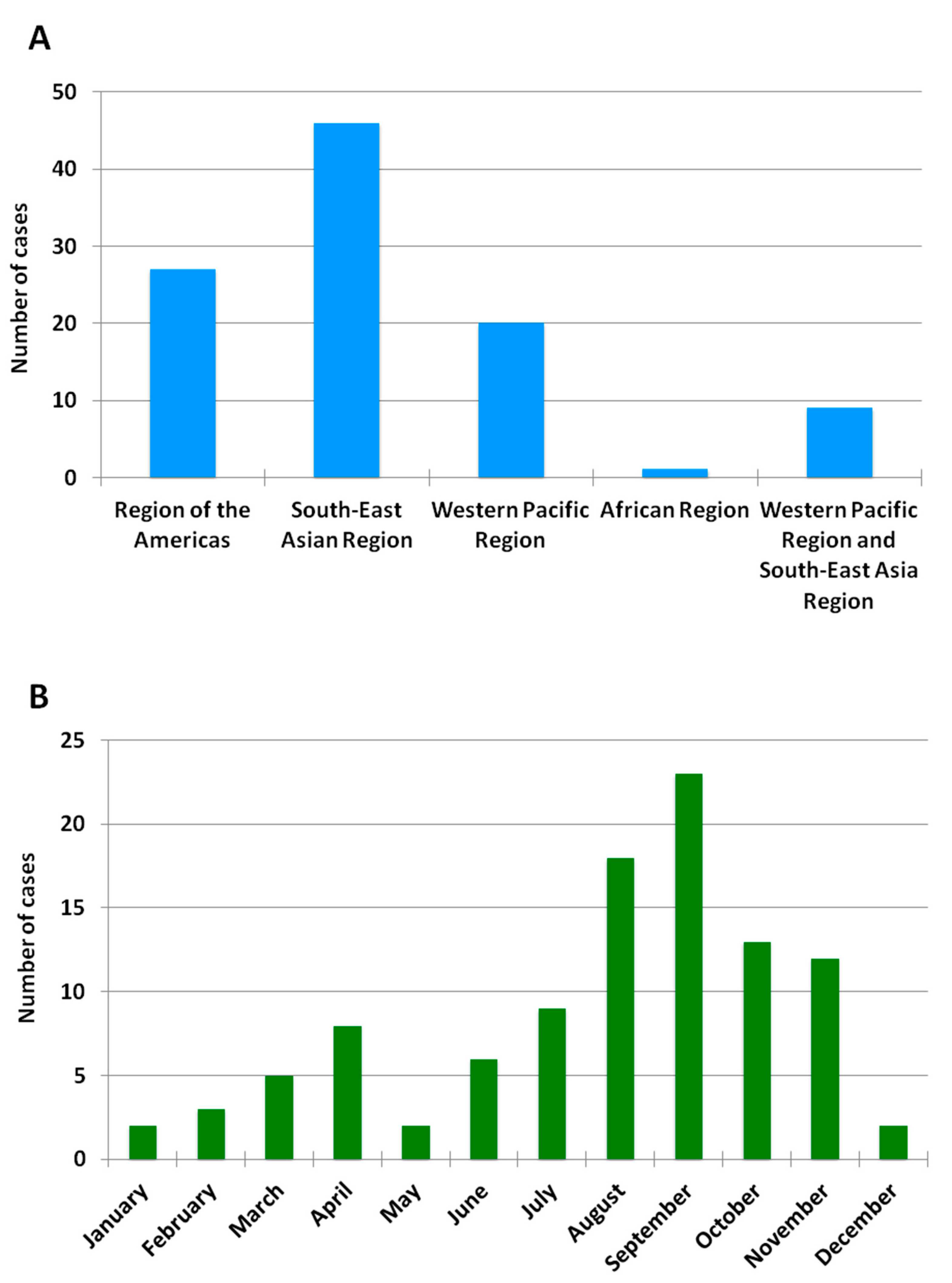
3.1. Imported Dengue Cases
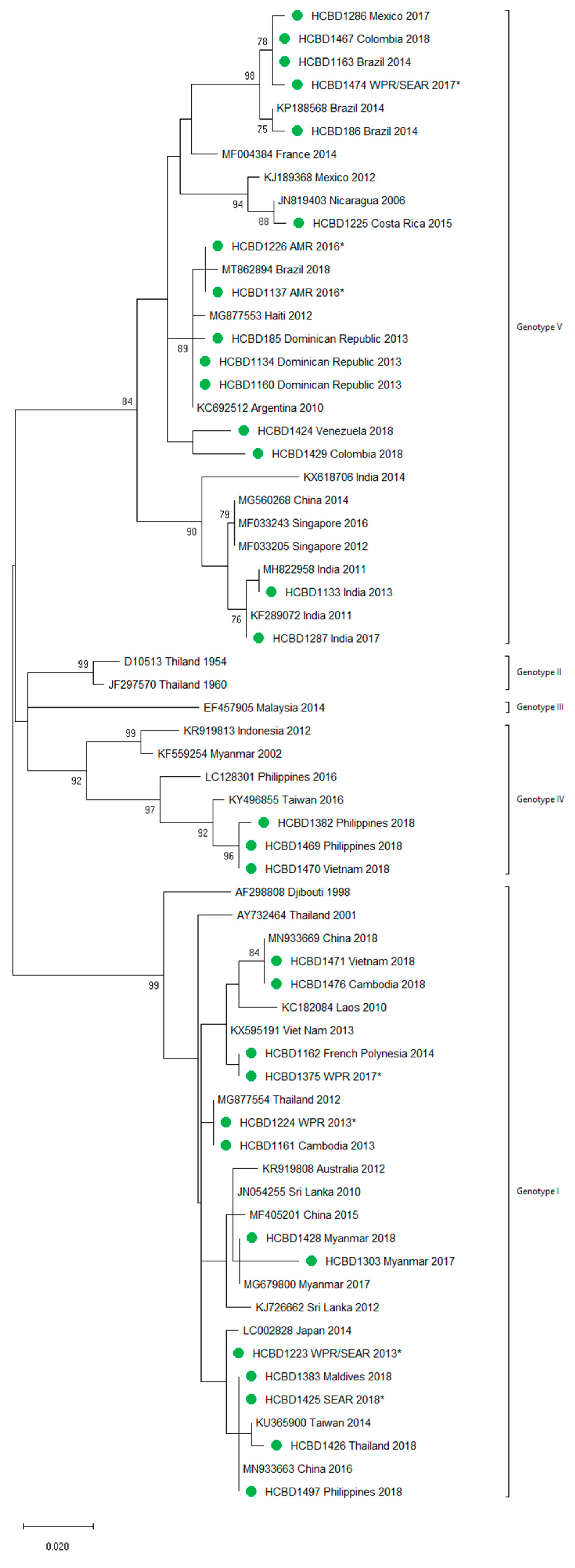
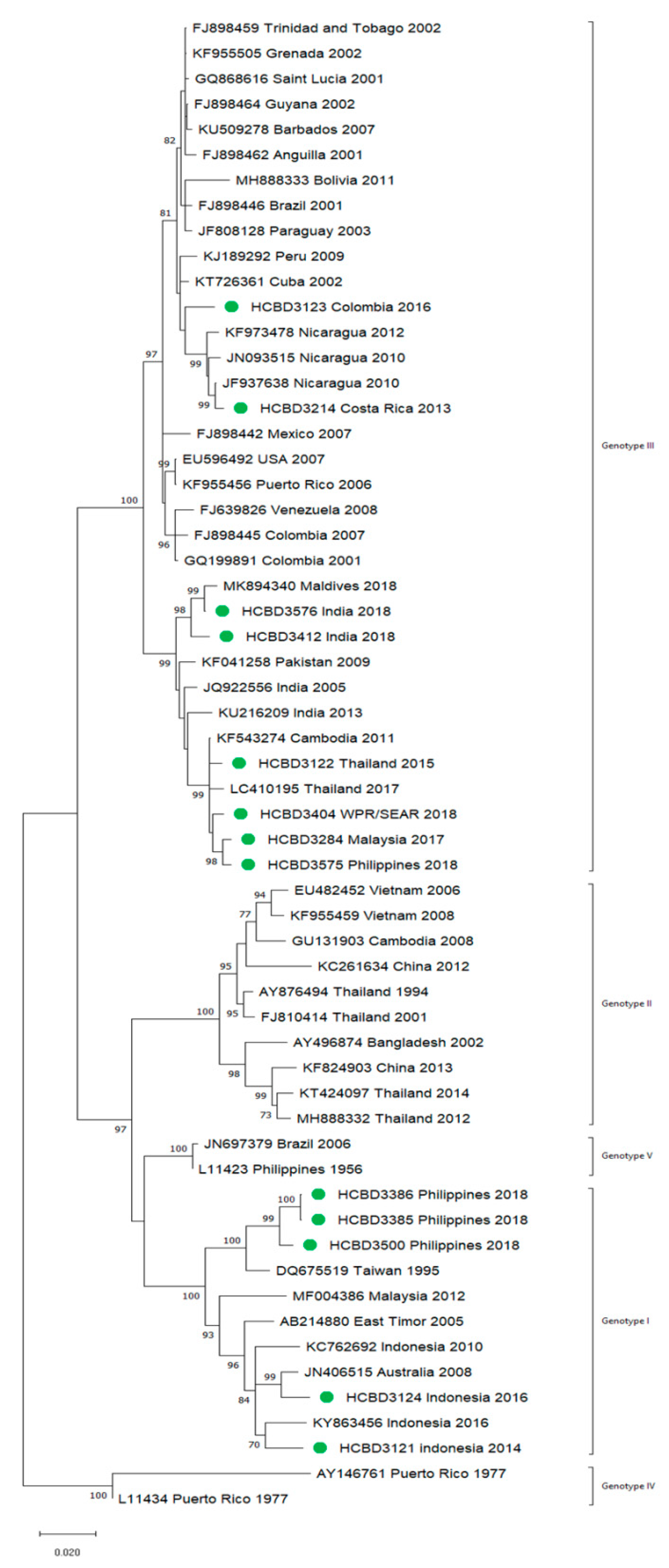
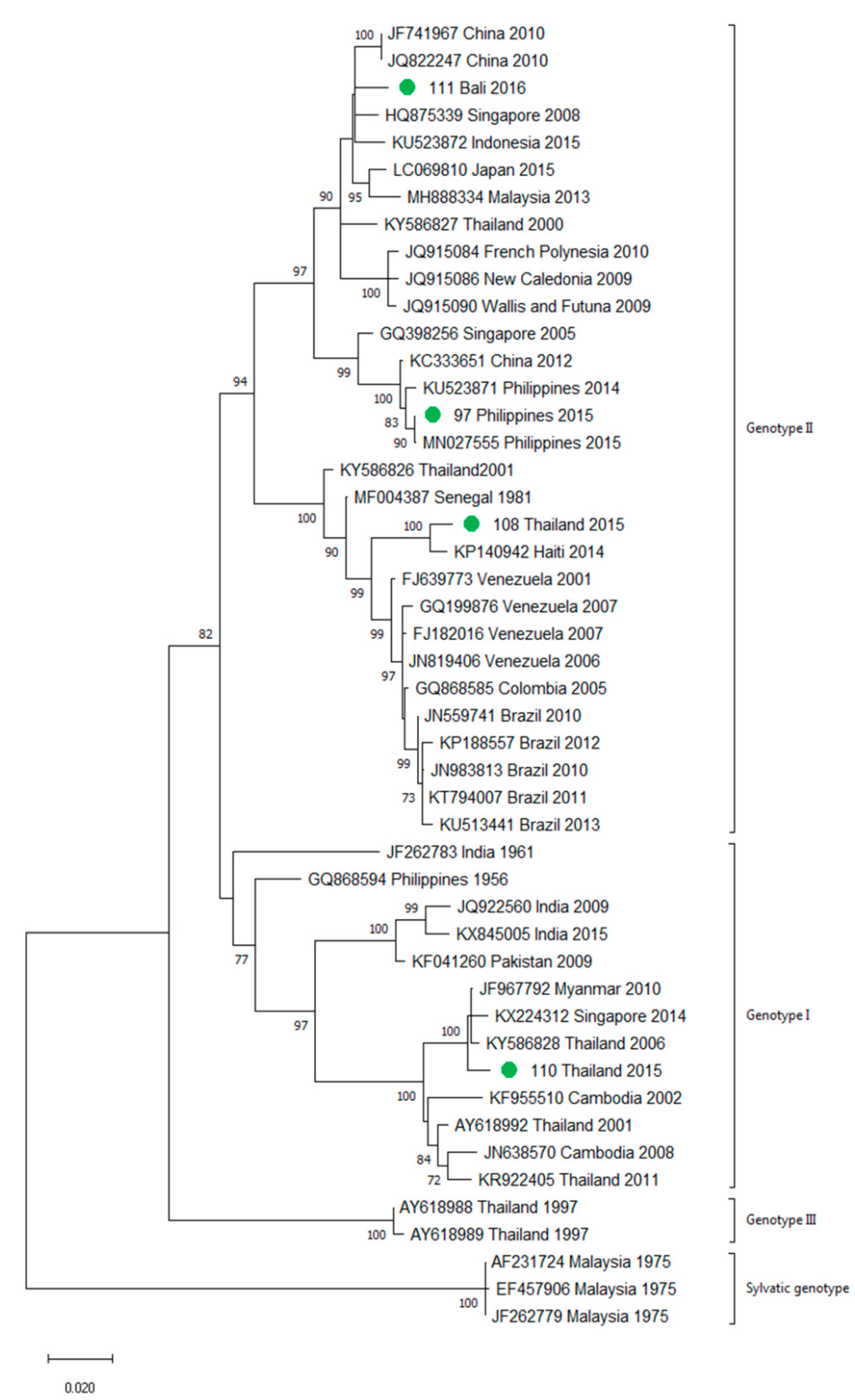
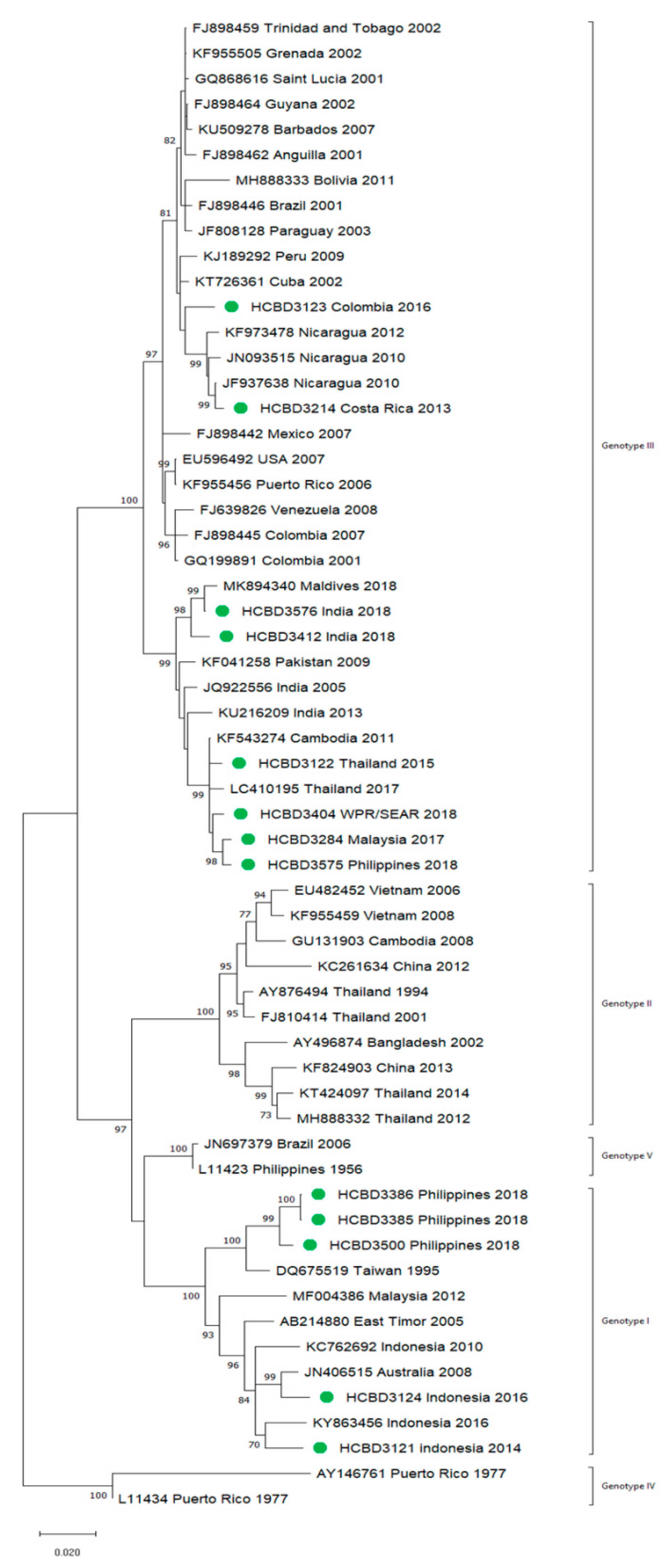
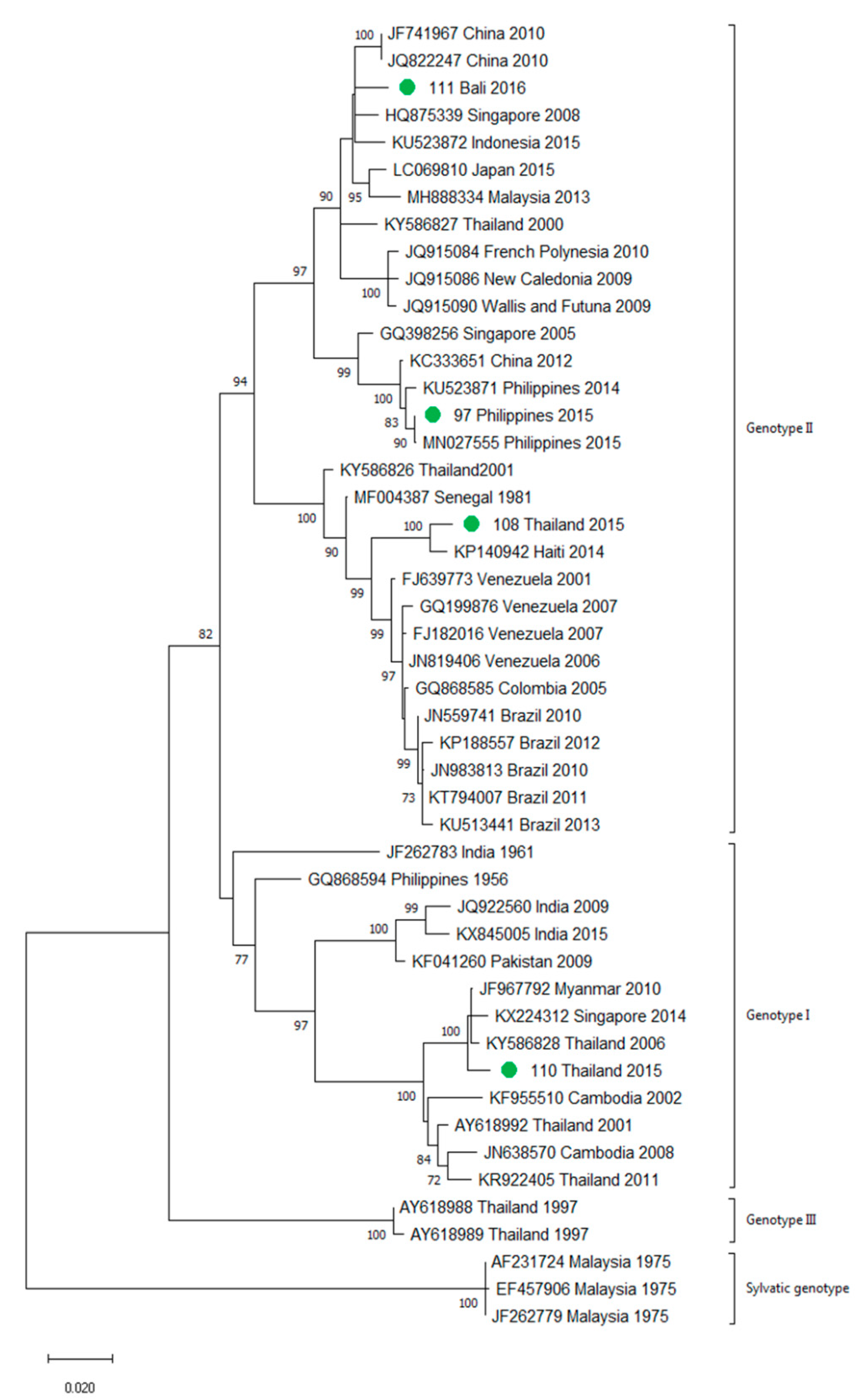
3.2. Molecular Characterization of Imported Dengue
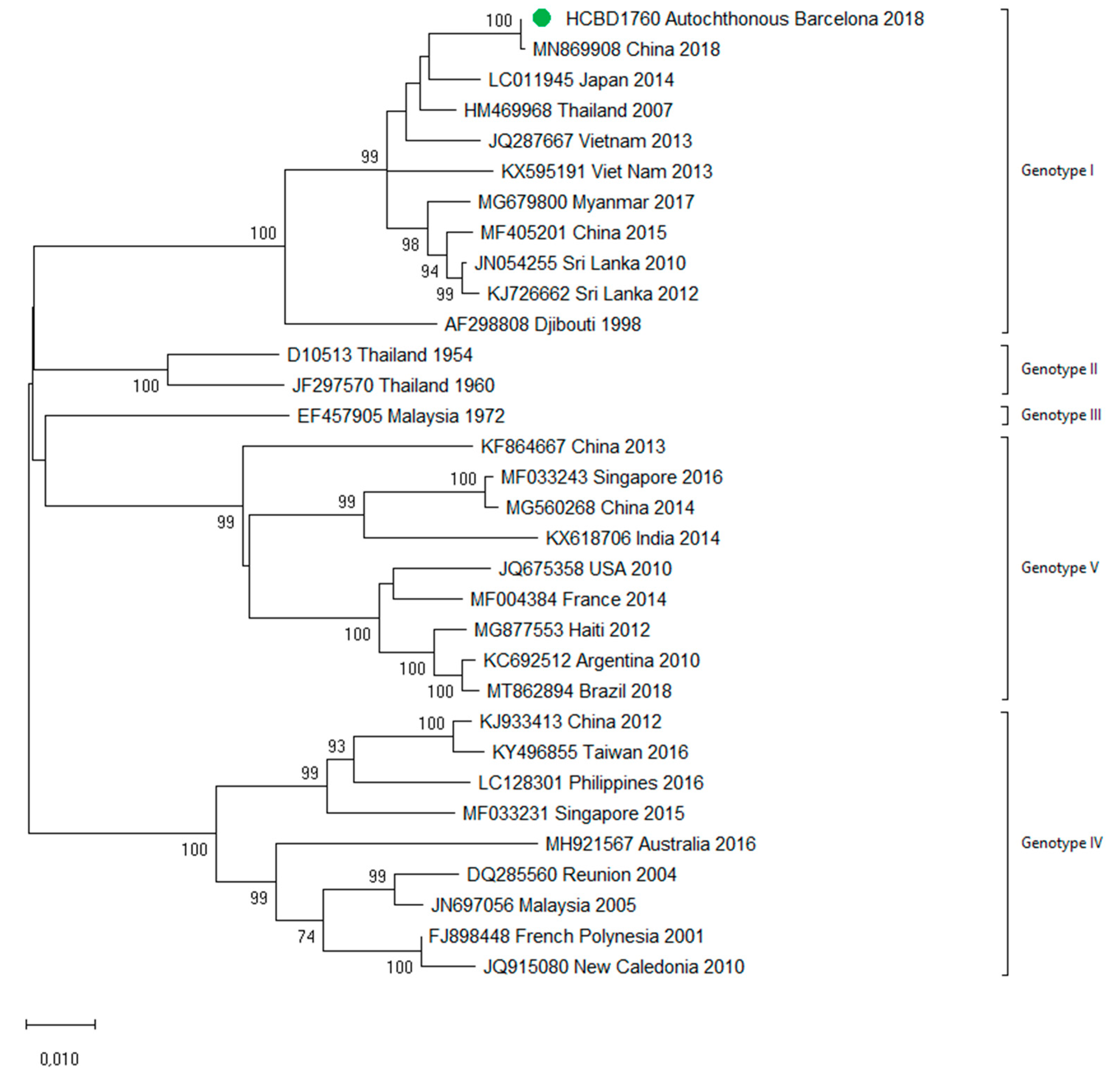
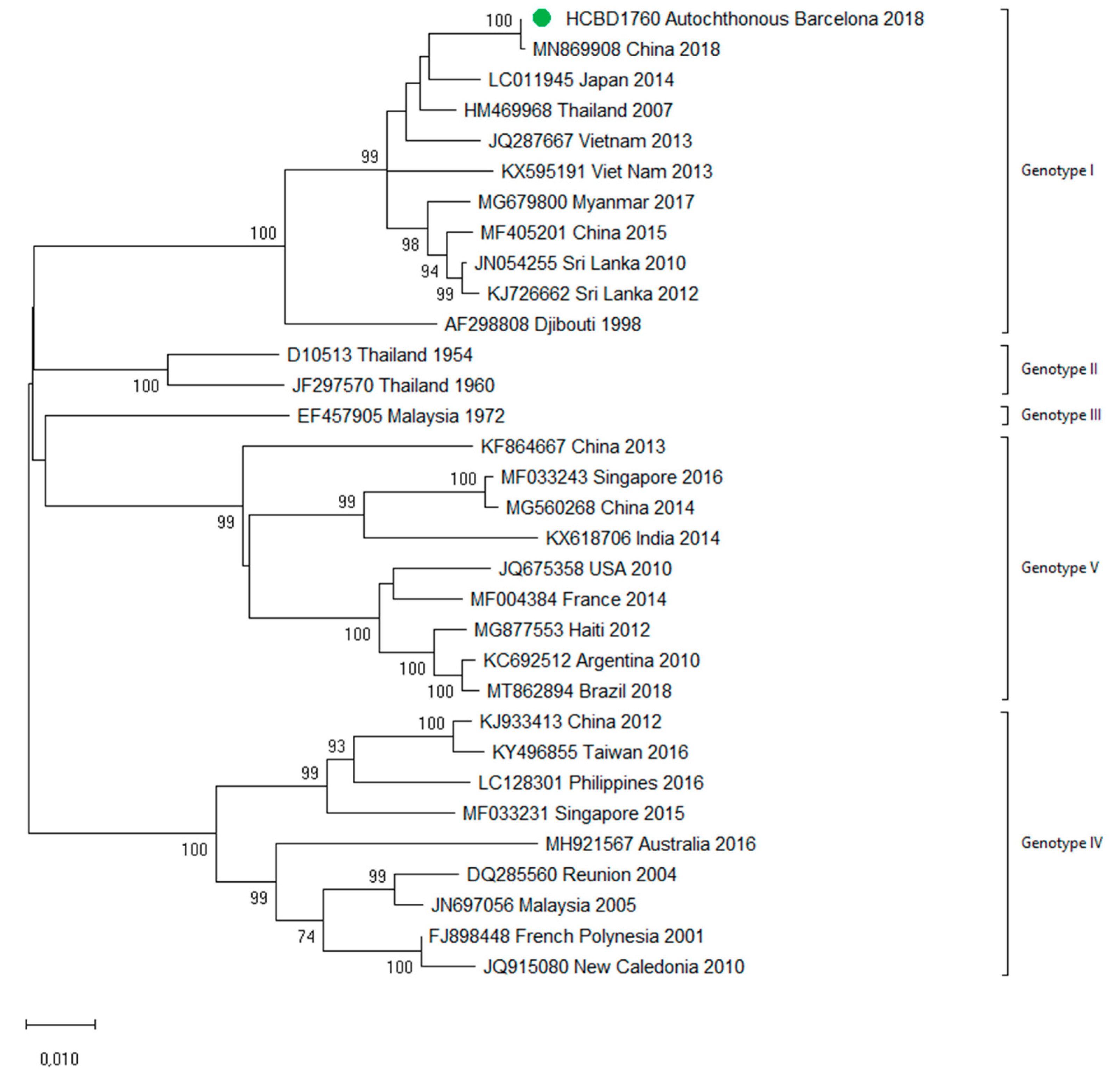
3.3. First Autochthonous Case in Northeastern Spain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. Dis. Prim. 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Kramer, I.M.; Pfeiffer, M.; Steffens, O.; Schneider, F.; Gerger, V.; Phuyal, P.; Braun, M.; Magdeburg, A.; Ahrens, B.; Groneberg, D.A.; et al. The ecophysiological plasticity of Aedes aegypti and Aedes albopictus concerning overwintering in cooler ecoregions is driven by local climate and acclimation capacity. Sci. Total. Environ. 2021, 778, 146128. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Scott, T.W.; Gubler, D.J. Consequences of the Expanding Global Distribution of Aedes albopictus for Dengue Virus Transmission. PLoS Negl. Trop. Dis. 2010, 4, e646. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Autochthonous Transmission of Dengue Virus in EU/EEA, 2010-Present. Available online: https://www.ecdc.europa.eu/en/all-topics-z/dengue/surveillance-and-disease-data/autochthonous-transmission-dengue-virus-eueea. (accessed on 19 June 2021).
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention, and Control; WHO: Geneva, Switzerland, 2009; p. 147. [Google Scholar]
- Yacoub, S.; Mongkolsapaya, J.; Screaton, G. Recent advances in understanding dengue. F1000Research 2016, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Genet. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Vasilakis, N. Dengue-Quo tu et quo vadis? Viruses 2011, 3, 1562–1608. [Google Scholar] [CrossRef]
- Pollett, S.; Melendrez, M.; Berry, I.M.; Duchene, S.; Salje, H.; Cummings, D.; Jarman, R. Understanding dengue virus evolution to support epidemic surveillance and counter-measure development. Infect. Genet. Evol. 2018, 62, 279–295. [Google Scholar] [CrossRef]
- Rico-Hesse, R. Microevolution and virulence of dengue viruses. Adv. Virus Res. 2003, 59, 315–341. [Google Scholar]
- Wittke, V.; Robb, T.; Thu, H.; Nisalak, A.; Nimmannitya, S.; Kalayanrooj, S.; Vaughn, D.; Endy, T.; Holmes, E.; Aaskov, J. Extinction and Rapid Emergence of Strains of Dengue 3 Virus during an Interepidemic Period. Virology 2002, 301, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Vasilakis, N. Molecular evolution of dengue viruses: Contributions of phylogenetics to understanding the history and epidemiology of the preeminent arboviral disease. Infect. Genet. Evol. 2009, 9, 523–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyke, A.T.; Moore, P.R.; Taylor, C.T.; Hall-Mendelin, S.; Cameron, J.N.; Hewitson, G.R.; Pukallus, D.S.; Huang, B.; Warrilow, D.; Hurk, A.F.V.D. Highly divergent dengue virus type 1 genotype sets a new distance record. Sci. Rep. 2016, 6, 22356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutsuna, S.; Kato, Y.; Moi, M.L.; Kotaki, A.; Ota, M.; Shinohara, K.; Kobayashi, T.; Yamamoto, K.; Fujiya, Y.; Mawatari, M.; et al. Autochthonous Dengue Fever, Tokyo, Japan, 2014. Emerg. Infect. Dis. 2015, 21, 517–520. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wu, T.; Liu, B.; Nelly, R.M.J.; Fu, Y.; Kang, X.; Chen, C.; Huang, Z.; Wu, B.; Wang, J.; et al. The Origin and Molecular Epidemiology of Dengue Fever in Hainan Province, China, 2019. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Franco, L.; Pagan, I.; Delcor, N.S.; Schunk, M.; Neumayr, A.; Molero, F.; Potente, A.; Hatz, C.; Wilder-Smith, A.; Sanchez-Seco, M.P.; et al. Molecular epidemiology suggests Venezuela as the origin of the dengue outbreak in Madeira, Portugal in 2012–2013. Clin. Microbiol. Infect. 2015, 21, 713.e5–713.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Instituto de Salud Carlos. Red Nacional de Vigilancia Epidemiológica. Resultados de la vigilancia Epidemiológico de las enfermedades transmisibles. Informe anual. Años 2017–2018. Available online: https://repisalud.isciii.es/handle/20.500.12105/11822 (accessed on 19 June 2021).
- Monge, S.; García-Ortúzar, V.; Hernández, B.L.; Pérez, M.Á.L.; Delacour-Estrella, S.; Sánchez-Seco, M.P.; Martinez, B.F.; Miguel, L.G.S.; García-Fulgueiras, A.; Moros, M.J.S. Characterization of the first autochthonous dengue outbreak in Spain (August–September 2018). Acta Trop. 2020, 205, 105402. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Autochthonous Cases of Dengue in Spain and France; European Centre for Disease Prevention and Control (ECDC): Solna Municipality, Sweden, 2019. [Google Scholar]
- European Centre for Disease Prevention and Control (ECDC). Rapid Risk Assessment-Sexual Transmission of Dengue in Spain; European Centre for Disease Prevention and Control (ECDC): Solna Municipality, Sweden, 2019. [Google Scholar]
- Moureau, G.; Temmam, S.; Gonzalez, J.-P.; Charrel, R.; Grard, G.; De Lamballerie, X. A Real-Time RT-PCR Method for the Universal Detection and Identification of Flaviviruses. Vector-Borne Zoonotic Dis. 2007, 7, 467–478. [Google Scholar] [CrossRef]
- Domingo, C.; Palacios, G.; Niedrig, M.; Cabrerizo, M.; Jabado, O.; Reyes, N.; Lipkin, W.I.; Tenorio, A. A new tool for the diagnosis and molecular surveillance of dengue infections in clinical samples. Dengue Bull. 2004, 28, 87–95. [Google Scholar]
- Domingo, C.; Niedrig, M.; Gascon, J.; Palacios, G.; Reyes, N.; Malo, M.J.; Wichmann, O.; Ruiz, J.; Schultze, D.; Schunk, M.; et al. Molecular Surveillance of Circulating Dengue Genotypes Through European Travelers. J. Travel Med. 2011, 18, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Meng, M.; Kumar, R.; Wu, Y.; Huang, J.; Lian, N.; Deng, Y.; Lin, S. Fatal Dengue Acquired in Florida. N. Engl. J. Med. 2021, 384, 2257–2259. Available online: http://www.ncbi.nlm.nih.gov/pubmed/32293753. (accessed on 10 June 2021).
- Aranda, C.; Martinez, M.J.; Montalvo, T.; Eritja, R.; Navero-Castillejos, J.; Herreros, E.; Marqués, E.; Escosa, R.; Corbella, I.; Bigas, E.; et al. Arbovirus surveillance: First dengue virus detection in local Aedes albopictus mosquitoes in Europe, Catalonia, Spain, 2015. Eurosurveillance 2018, 23, 1700837. [Google Scholar] [CrossRef] [Green Version]
- Luz, M.A.V.; Nabeshima, T.; Moi, M.L.; Dimamay, M.T.A.; Pangilinan, L.S.; Dimamay, M.P.S.; Matias, R.R.; Mapua, C.A.; Buerano, C.C.; de Guzman, F.; et al. An Epidemic of Dengue Virus Serotype-4 during the 2015–2017: The Emergence of a Novel Genotype IIa of DENV-4 in the Philippines. Jpn. J. Infect. Dis. 2019, 72, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harapan, H.; Michie, A.; Sasmono, R.T.; Imrie, A. Dengue: A Minireview. Viruses 2020, 12, 829. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A Review of the Invasive Mosquitoes in Europe: Ecology, Public Health Risks, and Control Options. Vector-Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [Green Version]
- European Centre for Disease Prevention and Control (ECDC). Autochthonous Transmission of Chikungunya Virus in EU/EEA, 2007–Present. Available online: https://www.ecdc.europa.eu/en/all-topics-z/chikungunya-virus-disease/surveillance-threats-and-outbreaks/autochthonous. (accessed on 26 June 2021).
- Giron, S.; Franke, F.; Decoppet, A.; Cadiou, B.; Travaglini, T.; Thirion, L.; Durand, G.; Jeannin, C.; L’Ambert, G.; Grard, G.; et al. Vector-borne transmission of Zika virus in Europe, southern France, August 2019. Eurosurveillance 2019, 24, 1900655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzarini, L.; Barzon, L.; Foglia, F.; Manfrin, V.; Pacenti, M.; Pavan, G.; Rassu, M.; Capelli, G.; Montarsi, F.; Martini, S.; et al. First autochthonous dengue outbreak in Italy, August 2020. Eurosurveillance 2020, 25, 2001606. [Google Scholar] [CrossRef] [PubMed]
- Mariconti, M.; Obadia, T.; Mousson, L.; Malacrida, A.; Gasperi, G.; Failloux, A.-B.; Yen, P.-S. Estimating the risk of arbovirus transmission in Southern Europe using vector competence data. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Dengu-Annual Epidemiological Report for 2019. Available online: https://www.ecdc.europa.eu/en/publications-data/dengue-annual-epidemiological-report-2019 (accessed on 26 June 2021).
- European Centre for Disease Prevention and Control (ECDC). Chikungunya Virus Disease-Annual Epidemiological Report for 2019. Available online: https://www.ecdc.europa.eu/en/publications-data/chikungunya-virus-disease-annual-epidemiological-report-2019 (accessed on 11 June 2019).
- Neumayr, A.; Muñoz, J.; Schunk, M.; Bottieau, E.; Cramer, J.; Calleri, G.; López-Vélez, R.; Angheben, A.; Zoller, T.; Visser, L.; et al. Sentinel surveillance of imported dengue via travellers to Europe 2012 to 2014: TropNet data from the DengueTools Research Initiative. Eurosurveillance 2017, 22, 30433. [Google Scholar] [CrossRef] [Green Version]
- Duong, V.; Lambrechts, L.; Paul, R.E.; Ly, S.; Lay, R.S.; Long, K.C.; Huy, R.; Tarantola, A.; Scott, T.W.; Sakuntabhai, A.; et al. Asymptomatic humans transmit dengue virus to mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 14688–14693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, A.F.; Machado, L.C.; Oidtman, R.J.; Siconelli, M.J.L.; Tran, Q.M.; Fauver, J.R.; Carvalho, R.D.D.O.; Dezordi, F.Z.; Pereira, M.R.; de Castro-Jorge, L.A.; et al. Lying in wait: The resurgence of dengue virus after the Zika epidemic in Brazil. Nat. Commun. 2021, 12, 2619. [Google Scholar] [CrossRef] [PubMed]
- Leder, K.; Torresi, J.; Libman, M.D.; Cramer, J.P.; Castelli, F.; Schlagenhauf, P.; Wilder-Smith, A.; Wilson, M.E.; Keystone, J.S.; Schwartz, E.; et al. GeoSentinel Surveillance of illness in returned travelers, 2007–2011. Ann. Intern. Med. 2013, 158, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Collantes, F.; Delacour, S.; Alarcón-Elbal, P.M.; Ruiz-Arrondo, I.; Delgado, J.A.; Torrell-Sorio, A.; Bengoa, M.; Eritja, R.; Miranda, M. Ángel; Molina, R.; et al. Review of ten-years presence of Aedes albopictus in Spain 2004–2014: Known distribution and public health concerns. Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.T.; Chew, L.Z.X.; Choo, E.L.W.; Dickens, B.S.L.; Ong, J.; Aik, J.; Ng, L.C.; Cook, A.R. Increased dengue transmissions in Singapore Attributable to SARS-CoV-2 social distancing measures. J. Infect. Dis. 2021, 223, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.; Wilder-Smith, A. What Is the Impact of Lockdowns on Dengue? Curr. Infect. Dis. Rep. 2021, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Olive, M.-M.; Baldet, T.; Devillers, J.; Fite, J.; Paty, M.-C.; Paupy, C.; Quénel, P.; Quillery, E.; Raude, J.; Stahl, J.-P.; et al. The COVID-19 pandemic should not jeopardize dengue control. PLoS Negl. Trop. Dis. 2020, 14, e0008716. [Google Scholar] [CrossRef] [PubMed]
- Microbe, T.L. Arboviruses and COVID-19: The need for a holistic view. Lancet Microbe 2020, 1, e136. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Localization | Ref. | ||||
|---|---|---|---|---|---|---|---|
| Envelope gene | DENV-1 | RT-PCR | F | EGENE1-S | CCGAAACGTGGATGTCCTCTGARGG | 756–780 | [24] |
| R | EGENE-R | TCCTCCCATGCCTTCCCRATGG | 2553–2574 | ||||
| Nested PCR | F | F Nested | ATAGGAACATCCATYACYCAG | 866–887 | Designed de novo | ||
| R | EGENE/NS-RR | TGRAAYTTRTAYTGYTCTGTCC | 2502–2523 | [24] | |||
| DENV-2 | RT-PCR | F | EGENE2-S | CTGAAACATGGATGTCATCAGAAGG | 758–782 | [24] | |
| R | RRT 2 | GCYGARGCYARYTTTGARGGRG | 2533-2555 | Modified from [24] | |||
| Nested PCR | F | F Nested | ATGGCRGCDATYYTGGCDYAY | 844–865 | Designed de novo | ||
| R | R Nested | CGKGARTTCATYCCTATCCATGT | 2348–2371 | Designed de novo | |||
| DENV-3 | RT-PCR | F | EGENE3-S | CTCAAACCTGGATGTCGGCTGARGG | 756–780 | [24] | |
| R | RRT 3 | ATYCCRCAVACTCCATTYTYCC | 2561–2583 | Modified from [24] | |||
| Nested PCR | F | F NESTED | ATGYTGGTCACYCCATCCATG | 911–932 | Designed de novo | ||
| R | R NESTED | TTGTAYTGYTCTGTCCARGTRTG | 2511–2534 | Designed de novo | |||
| DENV-4 | RT-PCR | F | EGENE4-S | CTGAGACATGGATGTCATCGGAAGG | 760–784 | [24] | |
| R | RRT 4 | CACAGACCCCHTCTTTGTGRGC | 2567–2589 | Modified from [24] | |||
| Nested PCR | F | F NESTED | TACTCAGRAABCCAGGATTYGC | 869–890 | Designed de novo | ||
| R | R NESTED | YTCCATGACACYRCACAACCC | 2478–2470 | Designed de novo | |||
| E/NS1 junction | DENV-1 | RT-PCR | F | FRTC 1 | TGSYTGAGACYCARCAYGGNAC | 1869–1890 | Modified from [23] |
| R | RRTC 1 | YTCRTTTGATATYTGYYTCCAC | 2620–2641 | ||||
| Nested PCR | F | FNC 1 | GRAAATGTTYGARGCHACYGCCC | 2130–2153 | Modified from [24] | ||
| R | RNC 1 | TCYTCCCAYGCYYTYCCRATGG | 2553–2574 | ||||
| DENV-2 | RT-PCR | F | FRTC 2 | TAGCWRRRACRCARCATGGAAC | 1871–1889 | Modified from [23] | |
| R | RRTC 2 | CAGTTCYGGWGYTATYTGYYTCCAC | 2622–2646 | ||||
| Nested PCR | F | FNC 2 | CARTYARYATAGAAGCAGARCC | 2027–2048 | Modified from [24] | ||
| R | RNC 2 | GCYGAWGCYARYTTTGRRGGRG | 2534–2555 | ||||
| DENV-3 | RT-PCR | F | FRTC 3 | TYTCHGARACRCARCAYGGRAC | 1863–1884 | Modified from [23] | |
| R | RRTC 3 | BARYTCATTRGCTAYTTGCTTCCAY | 2614–2638 | ||||
| Nested PCR | F | FNC 3 | GRRAARATGTTYGAGRCSMCYG | 2125–2146 | Modified from [24] | ||
| R | RNC 3 | ATYCCRCAVACTCCATTYTYCC | 2562–2583 | ||||
| DENV-4 | RT-PCR | F | FRTC 4 | TGGCAGAAACACARCATGGRAC | 1873–1894 | Modified from [23] | |
| R | RRTC 4 | YARYTCRTTRGTTATTTGYTTCCAC | 2624–2648 | ||||
| Nested PCR | F | FNC4 | ATYGGYAAGATGTTYGAGTCY | 2130–2150 | Modified from [24] | ||
| R | RNC 4 | CACAGACCCCHTCTTTGTGRGC | 2568–2589 | ||||
| Serotype | Genotypes Detected | Number of Cases |
|---|---|---|
| DENV-1 | I | 15 |
| IV | 3 | |
| V | 16 | |
| DENV-2 | Asian I | 3 |
| American/Asian | 9 | |
| Cosmopolitan | 21 | |
| DENV-3 | I | 8 |
| III | 10 | |
| DENV-4 | I | 1 |
| III | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navero-Castillejos, J.; Benitez, R.; Torner, N.; Muñoz, J.; Camprubí-Ferrer, D.; Peiró-Mestres, A.; Sulleiro, E.; Silgado, A.; Gonzalo, V.; Falgueras, T.; et al. Molecular Characterization of Imported and Autochthonous Dengue in Northeastern Spain. Viruses 2021, 13, 1910. https://doi.org/10.3390/v13101910
Navero-Castillejos J, Benitez R, Torner N, Muñoz J, Camprubí-Ferrer D, Peiró-Mestres A, Sulleiro E, Silgado A, Gonzalo V, Falgueras T, et al. Molecular Characterization of Imported and Autochthonous Dengue in Northeastern Spain. Viruses. 2021; 13(10):1910. https://doi.org/10.3390/v13101910
Chicago/Turabian StyleNavero-Castillejos, Jessica, Rosa Benitez, Nuria Torner, José Muñoz, Daniel Camprubí-Ferrer, Aida Peiró-Mestres, Elena Sulleiro, Aroa Silgado, Verónica Gonzalo, Teresa Falgueras, and et al. 2021. "Molecular Characterization of Imported and Autochthonous Dengue in Northeastern Spain" Viruses 13, no. 10: 1910. https://doi.org/10.3390/v13101910
APA StyleNavero-Castillejos, J., Benitez, R., Torner, N., Muñoz, J., Camprubí-Ferrer, D., Peiró-Mestres, A., Sulleiro, E., Silgado, A., Gonzalo, V., Falgueras, T., Alejo-Cancho, I., Roldán, M., Plasencia, V., Albarracin, R., Perez, J., Navarro, A., Calderón, A., Rubio, R., Navarro, M., ... Martínez, M. J. (2021). Molecular Characterization of Imported and Autochthonous Dengue in Northeastern Spain. Viruses, 13(10), 1910. https://doi.org/10.3390/v13101910