A Lectin Disrupts Vector Transmission of a Grapevine Ampelovirus


Abstract
1. Introduction
2. Materials and Methods
2.1. Membrane Feeding Transmission Assays
2.2. Virion Localization Assays
2.3. Virus Transmission
3. Results
3.1. Mealybug Transmission of GLRaV-3 through an Artificial Diet Membrane System
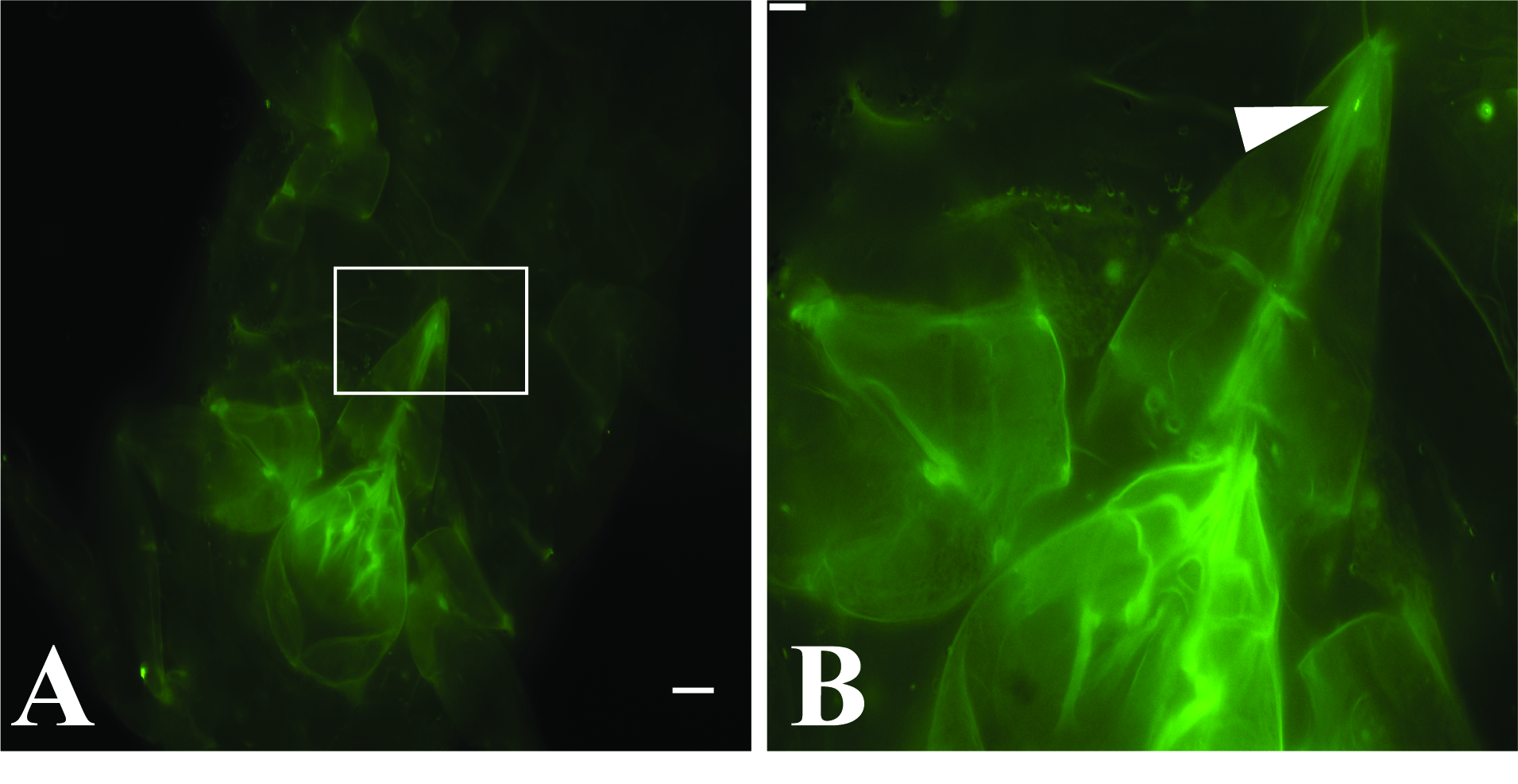
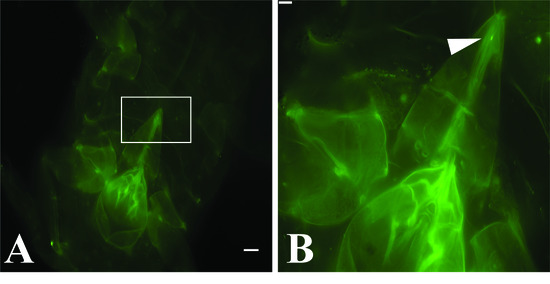
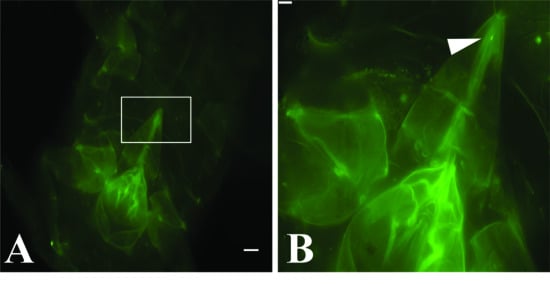
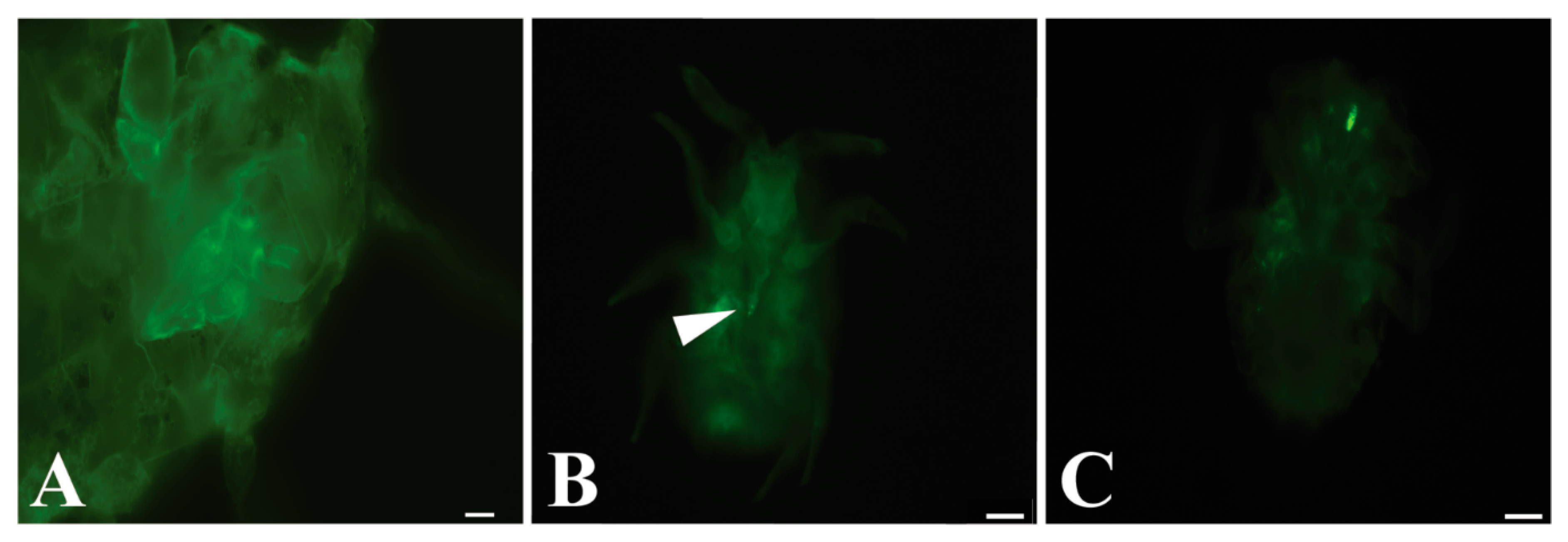
3.2. GLRaV-3 Virions Are Retained in the Mouthparts of Vectors
3.3. GLRaV-3-Transmission Is Blocked by a Lectin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, J.C.; Falk, B.W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 2006, 44, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Uzest, M.; Gargani, D.; Drucker, M.; Hébrard, E.; Garzo, E.; Candresse, T.; Fereres, A.; Blanc, S. A protein key to plant virus transmission at the tip of the insect vector stylet. Proc. Natl. Acad. Sci. USA 2007, 104, 17959–17964. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.; Drucker, M.; Uzest, M. Localizing viruses in their insect vectors. Annu. Rev. Phytopathol. 2014, 52, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.G.; Pichon, E.; van Munster, M.; Monsion, B.; Deshoux, M.; Gargani, D.; Calevro, F.; Jimenez, J.; Moreno, A.; Krenz, B.; et al. Identification of plant virus receptor candidates in the stylets of their aphid vectors. J. Virol. 2018, 92, e00432-e18. [Google Scholar] [CrossRef]
- Chen, A.Y.S.; Walker, G.P.; Carter, D.; Ng, J.C.K. A virus capsid component mediates virion retention and transmission by its insect vector. Proc. Natl. Acad. Sci. USA 2011, 108, 16777–16782. [Google Scholar] [CrossRef]
- Killiny, N.; Harper, S.; Alfaress, S.; El Mohtar, C.; Dawson, W. Minor coat and heat shock proteins are involved in the binding of citrus tristeza virus to the foregut of its aphid vector, Toxoptera citricida. Appl. Environ. Microbiol. 2016, 82, 6294–6302. [Google Scholar] [CrossRef]
- Maree, H.J.; Almeida, R.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.; Martelli, G.P. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4, 82. [Google Scholar] [CrossRef]
- Tsai, C.W.; Chau, J.; Fernandez, L.; Bosco, D.; Daane, K.M.; Almeida, R.P.P. Transmission of Grapevine leafroll-associated virus 3 by the Vine Mealybug (Planococcus ficus). Phytopathology 2008, 98, 1093–1098. [Google Scholar] [CrossRef]
- Daane, K.; Bentley, W.; Walton, V.; Malakar-Kuenen, R.; Millar, J.; Ingels, C.; Weber, E.; Gispert, C. New controls investigated for vine mealybug. Calif. Agric. 2006, 60, 31–38. [Google Scholar] [CrossRef]
- Herrbach, E.; Alliaume, A.; Prator, C.; Daane, K.; Cooper, M.; Almeida, R. Vector transmission of grapevine leafroll-associated viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Berlin/Heidelberg, Germany, 2017; pp. 483–503. [Google Scholar]
- Heriot, A. The crumena of the Coccidae and the Adelges. J. Entomol. Soc. BC 1936, 33, 22–24. [Google Scholar]
- Uzest, M.; Gargani, D.; Dombrovsky, A.; Cazevieille, C.; Cot, D.; Blanc, S. The “acrostyle”: A newly described anatomical structure in aphid stylets. Arthropod Struct. Dev. 2010, 39, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Cid, M.; Fereres, A. Characterization of the probing and feeding behavior of Planococcus citri (Hemiptera: Pseudococcidae) on grapevine. Ann. Entomol. Soc. Am. 2010, 103, 404–417. [Google Scholar] [CrossRef]
- Prator, C.A.; Kashiwagi, C.M.; Voncina, D.; Almeida, R.P.P. Infection and colonization of Nicotiana benthamiana by Grapevine leafroll-associated virus 3. Virology 2017, 510, 60–66. [Google Scholar] [CrossRef]
- Klaassen, V.; Boeshore, M.; Dolja, V.V.; Falk, B.W. Partial characterization of the lettuce infectious yellows virus genomic RNAs, identification of the coat protein gene and comparison of its amino acid sequence with those of other filamentous RNA plant viruses. J. Gen. Virol. 1994, 75, 1525–1533. [Google Scholar] [CrossRef]
- Sharma, A.M.; Wang, J.; Duffy, S.; Zhang, S.; Wong, M.K.; Rashed, A.; Cooper, M.L.; Daane, K.M.; Almeida, R.P.P. Occurrence of grapevine leafroll-associated virus complex in Napa Valley. PLoS ONE 2011, 6, e26227. [Google Scholar] [CrossRef]
- Killiny, N.; Rashed, A.; Almeida, R.P.P. Disrupting the transmission of a vector-borne plant pathogen. Appl. Environ. Microbiol. 2012, 78, 638–643. [Google Scholar] [CrossRef]
- Heinze, G.; Schemper, M. A solution to the problem of separation in logistic regression. Stat. Med. 2002, 21, 2409–2419. [Google Scholar] [CrossRef]
- Heinze, G.; Ploner, M.; Dunkler, D.; Southworth, H. Logistf: Firth’s Bias Reduced Logistic Regression (R Package Version 1.21). 2013. Available online: http://finzi.psych.upenn.edu/library/logistf/html/logistf-package.html (accessed on 6 May 2020).
- Childress, S.A.; Harris, K.F. Localization of virus-like particles in the foreguts of viruliferous Graminella nigrifrons leafhoppers carrying the semi-persistent maize chlorotic dwarf virus. J. Gen. Virol. 1989, 70, 247–251. [Google Scholar] [CrossRef]
- Murant, A.; Roberts, I.; Elnagar, S. Association of virus-like particles with the foregut of the aphid Cavariella aegopodii transmitting the semi-persistent viruses anthriscus yellows and parsnip yellow fleck. J. Gen. Virol. 1976, 31, 47–57. [Google Scholar] [CrossRef]
- Ammar, E.; Nault, L. Maize chlorotic dwarf viruslike particles associated with the foregut in vector and nonvector leafhopper species. Phytopathology 1991, 81, 444–448. [Google Scholar] [CrossRef]
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. C. R. Biol. 2010, 333, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Labroussaa, F.; Zeilinger, A.R.; Almeida, R.P.P. Blocking the transmission of a noncirculative vector-borne plant pathogenic bacterium. Mol. Plant Microbe Interact. 2016, 29, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.; Herrler, G.; Klenk, H.D. Sialic acid receptors of viruses. Top. Curr. Chem. 2015, 367, 1–28. [Google Scholar] [PubMed]
- Whitfield, A.E.; Rotenberg, D. Disruption of insect transmission of plant viruses. Curr. Opin. Insect Sci. 2015, 8, 79–87. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Recipient | Plants Infected/Plants Inoculated | Control |
|---|---|---|---|
| V. vinifera diet | V. vinifera | 12/94 | 0/18 |
| N. benthamiana | 1/187 | 0/37 | |
| N. benthamiana diet | V. vinifera | 1/84 | 0/17 |
| N. benthamiana | 2/125 | 0/25 |
| Source | Stylet | Cibarium | Both | Total | |
|---|---|---|---|---|---|
| Artificial diet | Number of P. ficus labeled | 19 | 11 | 1 | 31 |
| P. ficus controls * | 0 | 0 | 0 | 149 | |
| Total P. ficus viewed | 794 | ||||
| GLRaV-3 vine cuttings | Number of P. ficus labeled | 4 | 4 | 0 | 8 |
| P. ficus controls † | 0 | 0 | 0 | 35 | |
| Total P. ficus viewed | 333 |
| (a) | |||||
| Treatment | 1 | 2 | 3 | Total Positive | |
| WGA | 0/15 | 0/15 | 0/15 | 0/45 | |
| Casein | 1/15 | 1/15 | 2/15 | 4/45 | |
| Sucrose | 4/16 | 3/15 | 2/15 | 9/46 | |
| (b) | |||||
| Treatment | Estimate | SE | χ2 Statistic | PValue | |
| Intercept | −1.204 | 0.517 | 6.331 | 0.011 | |
| Casein | −0.828 | 0.608 | 1.963 | 0.161 | |
| WGA | −3.103 | 1.429 | 10.492 | 0.001 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prator, C.A.; Almeida, R.P.P. A Lectin Disrupts Vector Transmission of a Grapevine Ampelovirus. Viruses 2020, 12, 843. https://doi.org/10.3390/v12080843
Prator CA, Almeida RPP. A Lectin Disrupts Vector Transmission of a Grapevine Ampelovirus. Viruses. 2020; 12(8):843. https://doi.org/10.3390/v12080843
Chicago/Turabian StylePrator, Cecilia A., and Rodrigo P. P. Almeida. 2020. "A Lectin Disrupts Vector Transmission of a Grapevine Ampelovirus" Viruses 12, no. 8: 843. https://doi.org/10.3390/v12080843
APA StylePrator, C. A., & Almeida, R. P. P. (2020). A Lectin Disrupts Vector Transmission of a Grapevine Ampelovirus. Viruses, 12(8), 843. https://doi.org/10.3390/v12080843