1. Introduction
Land preservation, a key strategy of conservation biology, is the effort to protect habitat from human-caused impacts such as fragmentation, development, species loss, and the encroachment of invasive exotic plants [
1,
2]. The establishment and range expansion of exotic plants are closely associated with human disturbances such as logging, mining, road construction, agriculture, horticulture, and development [
3,
4,
5]. This link between exotic plants and anthropogenic disturbance is the result of both direct and indirect effects. Not only are humans often responsible for the disturbances that facilitate exotic plant invasion, they often are directly responsible for the dispersal of exotic plants into these areas through propagule transport [
2,
6,
7]. Invasive exotic plants tend to be pioneer species with high reproductive rates and effective propagule dispersal mechanisms [
8], traits amenable to colonizing newly disturbed areas [
9]. Disturbed areas tend to harbor exotic plants and can act as propagule sources [
3,
10,
11]. As a result, contiguously forested areas near disturbed habitats are more likely to become invaded than interior forests [
12]. Even within preserved lands, roads and private lands can be pervasive, creating a patchwork of edge habitat (e.g., roads, fields, recent logging, and developments). Forest roads in particular are important vectors for the spread of invasive exotic plants [
11,
13]. This link between anthropogenic disturbance and exotic plants has justified setting aside large tracts of land to maintain ecological integrity [
14,
15]. However, despite efforts to limit anthropogenic disturbance, natural disturbances such as tree canopy windthrow, fire, and flooding continue unabated. Given natural disturbances will continue unabated: Is removal of human disturbances through land preservation effective in limiting exotic plants?
Invasive exotic plants pose a serious threat to native ecosystems through mechanisms such as competition, allelopathy, and habitat manipulation [
16,
17,
18]. Exotic plants can outcompete and displace native plants, as is the case for species such as tree-of-heaven (
Ailanthus altissima (Mill.) Swingle) and Japanese stiltgrass (
Microstegium vimineum (Trin.) A. Camus) [
11,
19]. Furthermore, exotic plants may decrease biodiversity, which can have cascading impacts across species and trophic levels [
20,
21,
22,
23]. Invasion can disrupt ecological communities and may limit native tree re-establishment following a disturbance [
24,
25]. Given the tendency of invasive plants to take advantage of available resources, disturbed areas present a prime opportunity for invasive plants to exploit.
Canopy disturbances in forests tend to increase the amount of sunlight reaching the forest floor, promoting new plant growth and forest heterogeneity through variable forest structure and new species composition [
5,
15,
26]. Canopy disturbances have the potential to affect all forests; however, forests without a preservation mandate are commonly salvage logged following natural disturbance events [
27]. Salvage logging increases exotic plant abundance due to more light reaching the forest floor, along with more exposed soil [
26,
27]. Preserved lands on the other hand present an opportunity to observe whether exotic plants opportunistically invade following natural disturbance despite the absence of anthropogenic disturbance.
In this study we focus on two natural disturbance events, severe storm systems Hurricane Sandy and a wind event known as the 29 June Derecho, which impacted the Central Appalachians in 2012 and caused a patchwork of tree windthrow and canopy gaps. These events generated a prime opportunity to investigate species responses to forest canopy disturbances in preserved lands. We used data from National Park Service (NPS) long-term vegetation monitoring plots [
28], located in areas affected by storm windthrow, to answer the following two questions. First, in preserved lands, does natural disturbance facilitate exotic vascular plant species invasion? We compared vegetation measured in plots before and after the 2012 forest canopy windthrow from severe storms to quantify changes in species richness and cover as a result of natural disturbance. We hypothesized that canopy disturbance and exotic plant colonization were positively correlated. Secondly, does proximity to edge habitat increase the abundance of exotic plants in naturally disturbed preserved areas? To address this question, we studied the interaction between plot proximity to edge habitat and plant species composition in disturbed and undisturbed areas. We hypothesized that canopy gaps with more adjacent edge habitats would have greater exotic plant cover pre-disturbance and a proportionally greater increase in these plants post-disturbance correlating to the amount of edge.
2. Materials and Methods
2.1. Study Areas
The study areas were located in four United States National Park Service (NPS) units in the Central Appalachian Mountains: Bluestone National Scenic River (BLUE) in West Virginia (WV), Delaware Water Gap National Recreation Area (DEWA) in Pennsylvania (PA) and New Jersey (NJ), Gauley River National Recreation Area (GARI) in WV, and New River Gorge National River (NERI) in WV. The dominant forest types included dry oak-hickory-heath and mesic sugar maple-beech-basswood forests, though these parks cover a wide range of vegetation types, land-use histories, and baseline exotic plant prevalence [
29,
30,
31,
32]. DEWA, located along the Delaware River, has had longer and more intensive agricultural use than the other NPS units, as well as having a much greater human population density both historically (post-European colonization) and currently. These factors have led to a higher proportion of exotic plants in DEWA compared to the NPS units in West Virginia [
33].
These four NPS units contain 282 long-term vegetation monitoring plots that were established to monitor the long-term health of these forests and are intended to be sampled in perpetuity. The intensive sampling design prevents all 282 plots from being sampled in one growing season, and, therefore, the plots are sampled on a rotating panel design over a four-year period with one quarter of all plots sampled each year between 2007 and 2010, and resampled between 2011 and 2014 with a third round of sampling beginning in 2015 [
28]. A detailed justification of the sampling design and data collection methods can be found in Perles et al. 2014a [
28]. Here we only present an overview of the sampling methods. Protocols from the US Forest Service’s Forest Inventory Analysis program [
34] and four NPS Inventory and Monitoring protocols from networks in the eastern U.S. were fundamental in establishing these methods [
35,
36,
37,
38].
Monitoring plots were selected from a regular grid of possible sampling points using a Generalized Random-Tessellation Stratified design [
39] to ensure a balanced sample while still maintaining a random design. Points that fell on managed lands (i.e., parking lots, lawns, roads, etc.) were removed, as were sites that had >30 degree slope. Selected plot locations were installed and sampled in a four-year rotating panel design, meaning that a quarter of the total plots were installed each year from 2007 to 2010 and resampled between 2011 and 2014 with a third round of sampling beginning in 2015. All plots were resampled within two weeks of the date of original installation to capture a similar suite of the flora present at that time of year.
In October 2012 Hurricane Sandy brought high winds, heavy rains, and flooding to parts of the Mid-Atlantic including DEWA, while the 2012 Derecho (June 2012) swept across the upper Mid-West and Mid-Atlantic including BLUE, GARI, and NERI. A total of 18 plots (6.3% of all plots) were disturbed by these storms, one in BLUE, five in DEWA, seven in GARI, and five in NERI.
2.2. Field Methods
Plots were comprised of a 15-m radius circle, within which an array of forest condition measurements were sampled including live tree basal area (LBA), coarse woody debris (CWD), and vascular understory plant richness and cover. Plots were marked with metal spikes to ensure that the same areas were sampled each visit. A center point marked with rebar and a metal cap was installed, along with 25 cm galvanized metal nails that mark the ends of transects along the 0, 60, 120, 180, 240, and 300 degree azimuths from plot center.
Along the six transects, 12 1 m × 1 m quadrats were sampled to capture cover of all vascular plants by species covering an area under 2 m in height. In each quadrat, each species was given a cover class (0%, 1–2%, 2–5%, 5–10%, 10–25%, 25–50%, 50–75%, 75–95%, and 95–100%), and the midpoints of these cover classes were used in the analysis to represent species cover. The midpoint value of each cover class represents the mean of the two values at each end of each range. The average total cover of all species for a plot was calculated by summing the cover midpoints from all species within each quadrat and then averaging the summed values across the 12 quadrats. Total cover for a quadrat or average total cover for a plot may exceed 100% (or 1 m2) due to overlap of species canopies. Average species richness for each plot was calculated by determining species richness in each quadrat and then averaging those values across the 12 quadrats.
All plant taxa were classified as native or exotic and those designations were used to calculate native and exotic cover and richness using the formulas described above. Invasive exotic species can be difficult to define, let alone classify into a system to rank invasiveness. Therefore, we did not differentiate the degree of invasiveness between the different species and pooled exotic species in order to simplify the study. However, nearly all of the exotic plant species found in the disturbed plots are considered moderately or highly invasive according to NatureServe’s Invasive Species Impact Rank (I-Rank) classification system (
Appendix A,
Table A8), which provides some context for the invasiveness of the exotic plants encountered in this study. An I-Rank protocol was developed to rank invasive exotic plant species primarily by their detrimental impact to biodiversity [
40].
CWD was sampled along the six transects for fallen wood greater than 7.5 cm in diameter at its intersection with each transect. Huber’s formula [
41] was used to estimate CWD volume from line intersect transect data using the diameter of each CWD piece measured at the point of intersection with the transect line [
42]. This calculation was slope-corrected by converting the slope-length of each line intersect transect to its equivalent horizontal distance [
43]. Inserting a correction factor into the formula allows calculation of
CWD volume per hectare (m
3∙ha
−1) as
where %
Slope is percent slope of the transect,
Slope_length is the slope length of the transect in meters (i.e., 15 m), and
Diami is the recorded diameter in centimeters of each piece of coarse woody debris. CWD volume was then averaged across the six transects to generate average CWD volume per plot, expressed as a volume (m
3∙ha
−1).
Trees were measured for diameter at breast height (dbh) in cm, which is 1.37 m from the base of the tree on the uphill side, and scribed 5 cm below dbh to ensure precise measurements between sampling periods. Trees greater than 10 cm at dbh and within 15 m horizontal distance of the plot center were sampled. DEWA had five undisturbed plots without any trees present and those plots were excluded from the analyses because there were no trees that could have been blown over. Live tree basal area in square meters was calculated from dbh (cm) using the following formula below. Plot live basal area was calculated as the sum of the basal area of all the live trees in that plot. This value was converted to m
2∙ha
−1 by dividing by the plot area (707 m
2) and converting to hectares:
2.3. Disturbance Criteria
Plots were identified as disturbed if they experienced windstorm disturbance damage (i.e., stem breakage or tip-up) since the time of last sampling to >25% of all trees in a plot, or a disturbance (i.e., trees and tree tops falling into a plot) to >25% of the soil surface, understory vegetation, or canopy trees. These thresholds were coarse measurements designed to record disturbances that may have influenced the plot structure.
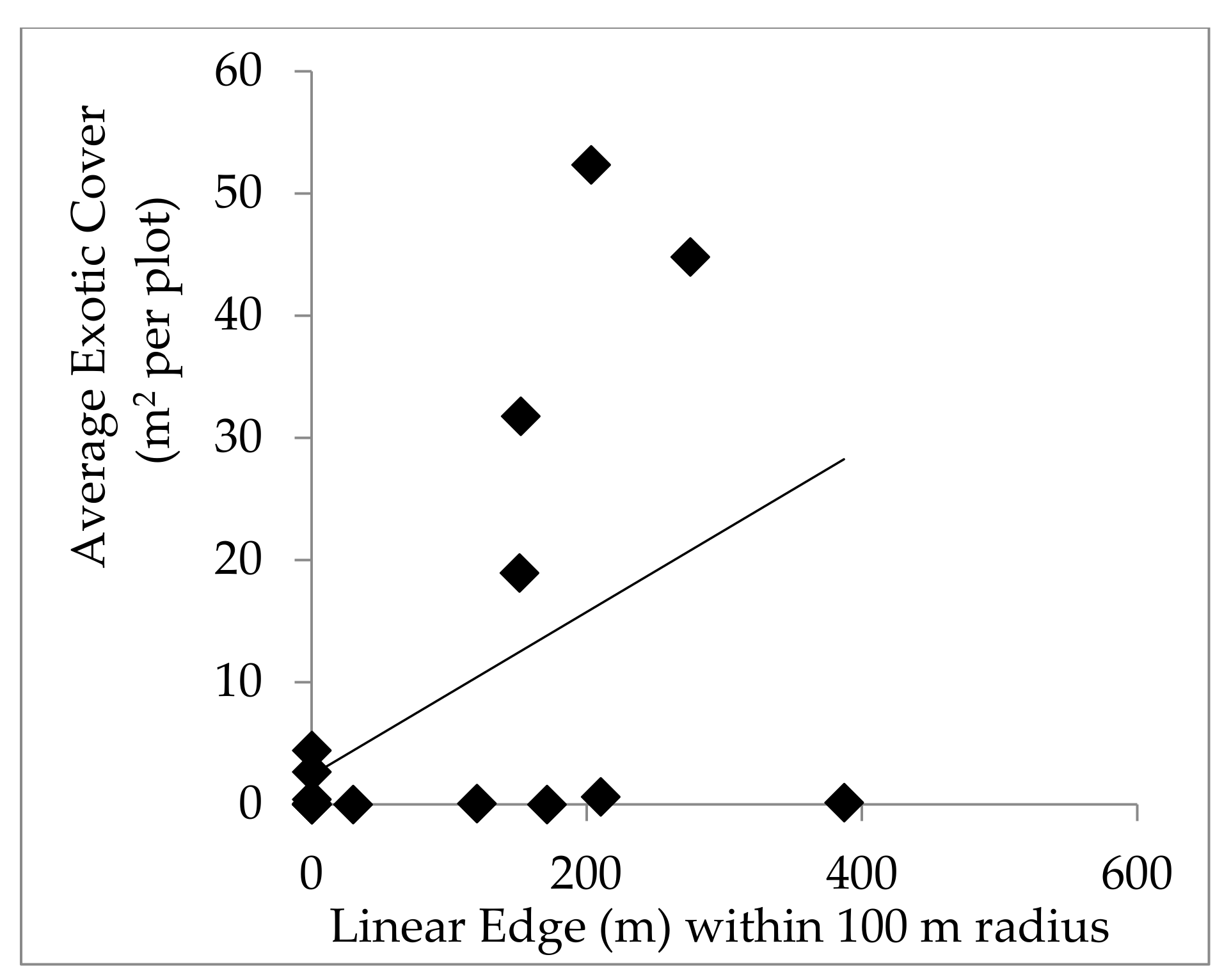
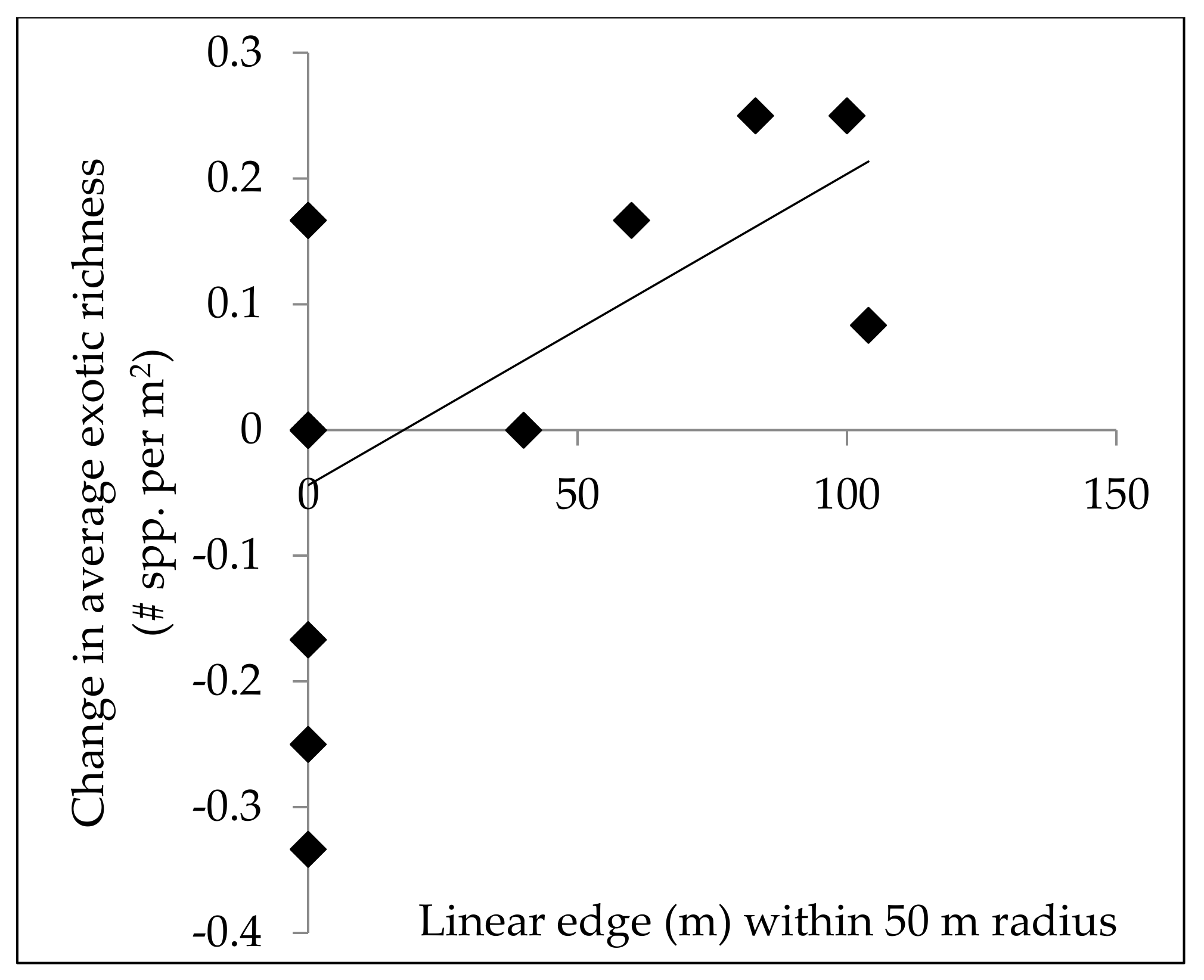
Edge habitat is ideal for invasive exotic plants due to high light availability, and can act as a source of propagule pressure for the surrounding area, so we quantified the amount of edge habitat for disturbed plots to relate to invasive exotic plant richness and abundance. The amount of linear edge surrounding each disturbed plot was measured using Google Earth Pro’s “Ruler” tool. Linear edge was defined as the edges of roads, fields, and other human developments. Measurements of total linear edge within 50 m, 100 m, and 250 m of the plot center were recorded. These distances were chosen based on research from Mortensen et al. 2009 [
11]. To determine if roads had a distinct impact on exotic plant recruitment in plots, the edge habitat along roads was analyzed separately, as well as grouped with other sources of edge habitat.
2.4. Data Analyses
Vegetation monitoring data from the NPS were used to study forest changes following disturbance. These data are made publicly available here:
https://irma.nps.gov/DataStore/. Data prior to disturbance from disturbed and undisturbed plots were compared to establish if disturbed plots were representative of NPS unit conditions. The amount of linear edge habitat to exotic plant cover and richness in plots that were later disturbed was compared to determine if edge habitat was associated with exotic plant abundance.
Vegetation responses to disturbance were compared by assessing post-disturbance forest structure (LBA and CWD) in disturbed versus undisturbed plots to confirm that disturbance impacted disturbed plots. Changes in plant species richness and cover in disturbed plots were compared from pre- to post-disturbance sampling; amount of linear edge habitat was then compared to these changes to determine if edge habitat was associated with exotic plant colonization.
The statistical programs Minitab and Microsoft Excel were used to analyze LBA, CWD, plant species richness, plant cover, and edge habitat data. Tukey’s method was used to calculate differences of means for LBA, CWD, and native and exotic plant richness. Native and exotic plant cover were analyzed using a general linear mixed model ANOVA in Minitab. NPS unit, plot condition (disturbed or undisturbed), and disturbance event (pre- or post-disturbance) were fixed factors in the ANOVA and interactions between NPS unit and plot condition, NPS unit and disturbance events, and plot condition and disturbance events were added into the model. Significant differences of means between LBA, CWD, native and exotic plant richness, as well as native and exotic plant cover, were identified at a = 0.05.
Amount of linear edge was compared by linear regression to plant cover and richness prior to disturbance and to the changes in plant cover (post-disturbance minus pre-disturbance) and richness for disturbed plots. Pre- and post-disturbance plant cover in disturbed and undisturbed plots were compared by linear regression, and the slope of those lines compared to the equation y = x that represents no change in cover or richness from pre- to post-disturbance.
2.5. Limitations
This study used data from an NPS long-term vegetation monitoring program that were not originally designed to capture in-depth details about the impacts of natural and stochastic disturbance events. However, pre-disturbance forest vegetation data from these plots presented a unique and exciting opportunity to investigate the impacts of these storms. The opportunistic nature of the study leads to particular limitations. For example, the monitoring protocol allowed for a coarse identification of canopy disturbance and does not capture gap size, which would be a better metric for the study of vegetation response to natural canopy disturbance. In addition, disturbance impacts would ideally be compared between disturbed and control plots, however we were unable to identify control plots that matched disturbed plots in all ways except disturbance, therefore we compared the 18 disturbed plots to the population of 262 monitoring plots. The limited number of disturbed plots should also be considered in the interpretation of results. Post-disturbance sampling happened over four field seasons following the 2012 storms. We did not separate plots by the time relative to disturbance because the number of plots in each time period varied widely. Plots were sampled from <1 to 3 years following disturbance, but one year had as many as ten disturbed plots while another year only had one disturbed plot. Years since disturbance was initially included as a covariate for most analyses, however it was removed in the final models because it explained little of the model variation. Ultimately, the time between the disturbance events and plot sampling was relatively short (<3 years). These findings only offer a view into initial vegetation responses and further work exploring long-term changes would certainly help demonstrate if these trends continue as the canopy re-establishes overtime. Finally, the inventory protocol uses a two-dimensional estimate of percent cover by species, which could be highly influenced by coarse woody debris dropping from the canopy from the wind disturbance.
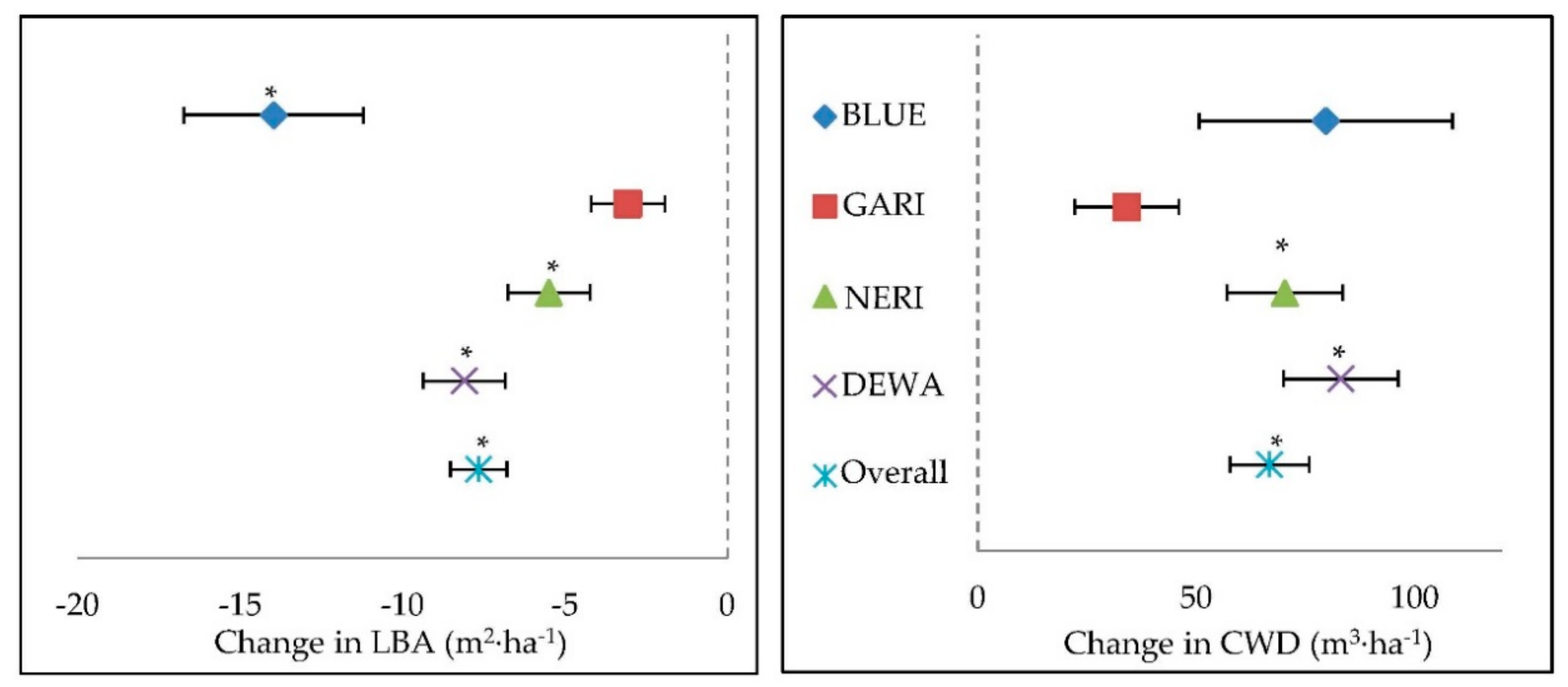
4. Discussion
Canopy disturbance in preserved forests caused a reduction in live tree basal area and an increase in coarse woody debris from fallen trees and broken limbs. Canopy damage increases light penetrating the canopy, which has the potential to change the forest structure and species composition [
5,
15,
26]. We hypothesized that exotic plant cover and richness would increase in the understory of these forests following disturbances as most of eastern North America’s invasive exotic plants are disturbance adapted [
3,
5,
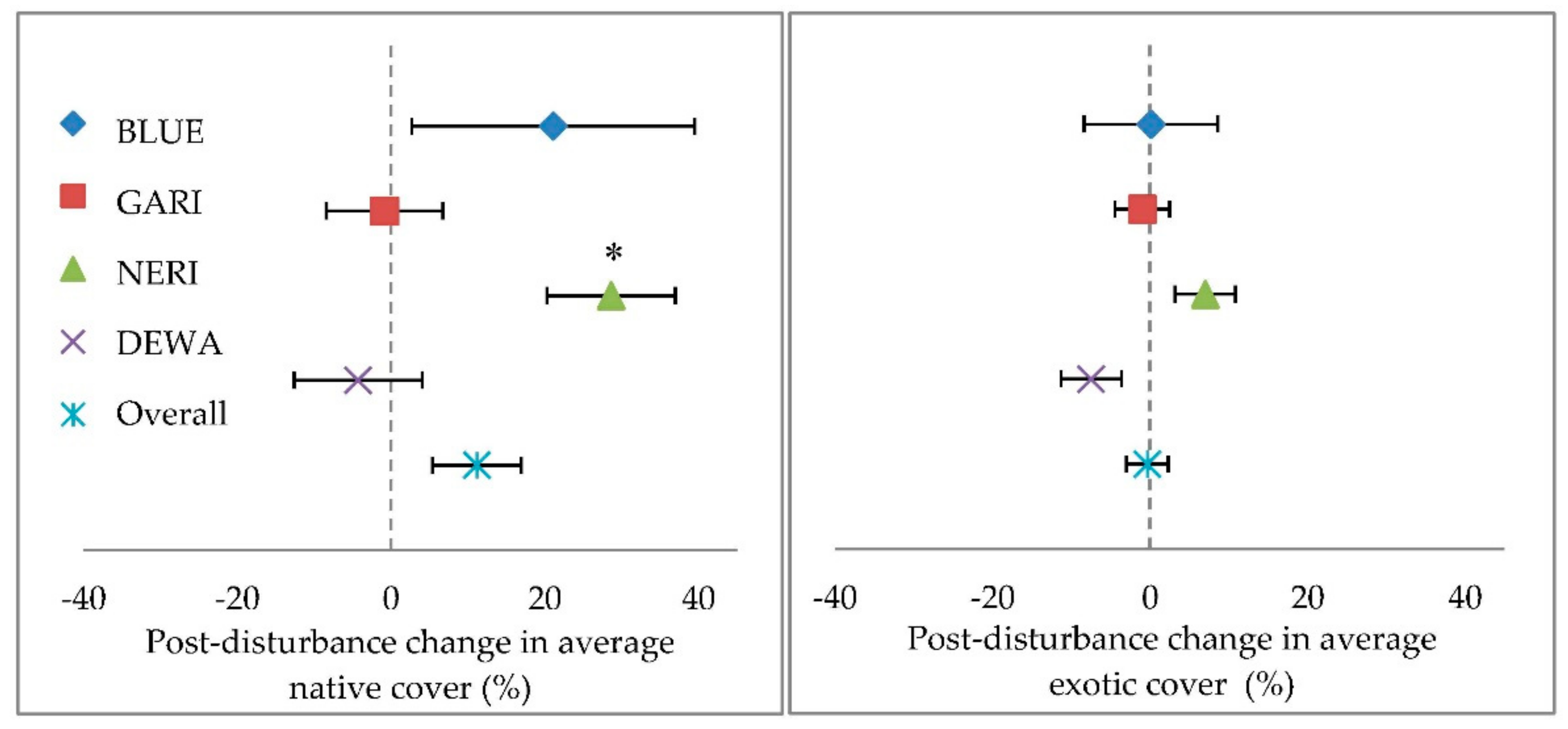
9]. However, exotic plant richness and cover in these disturbed forests did not increase relative to undisturbed forests in the three years following disturbance. Native plant cover and richness increased to a greater extent in response to canopy disturbance than that of exotic plants, though native plant cover change was highly variable between both disturbed and undisturbed plots.
In the majority of plots, exotic plant cover or richness did not increase following canopy disturbance despite the decrease in forest basal area and increase in coarse woody debris. The acute increase of
Glechoma hederacea at one plot underscores the stochasticity of plant responses to disturbance, while the dramatic reduction in exotic plant cover observed in the three most heavily invaded plots before disturbance suggests that fallen tree canopies and branches crushed or covered these plants. Consequentially, the fallen woody debris may have created ground-level shade and cover that prevented the generally more shade-intolerant exotics from establishing in the forest understory. In contrast, the increase in average native plant cover following disturbance may reflect the more shade-tolerant nature of the extant native plants that are common in the understory of forests in the study region [
4,
30,
31,
32].
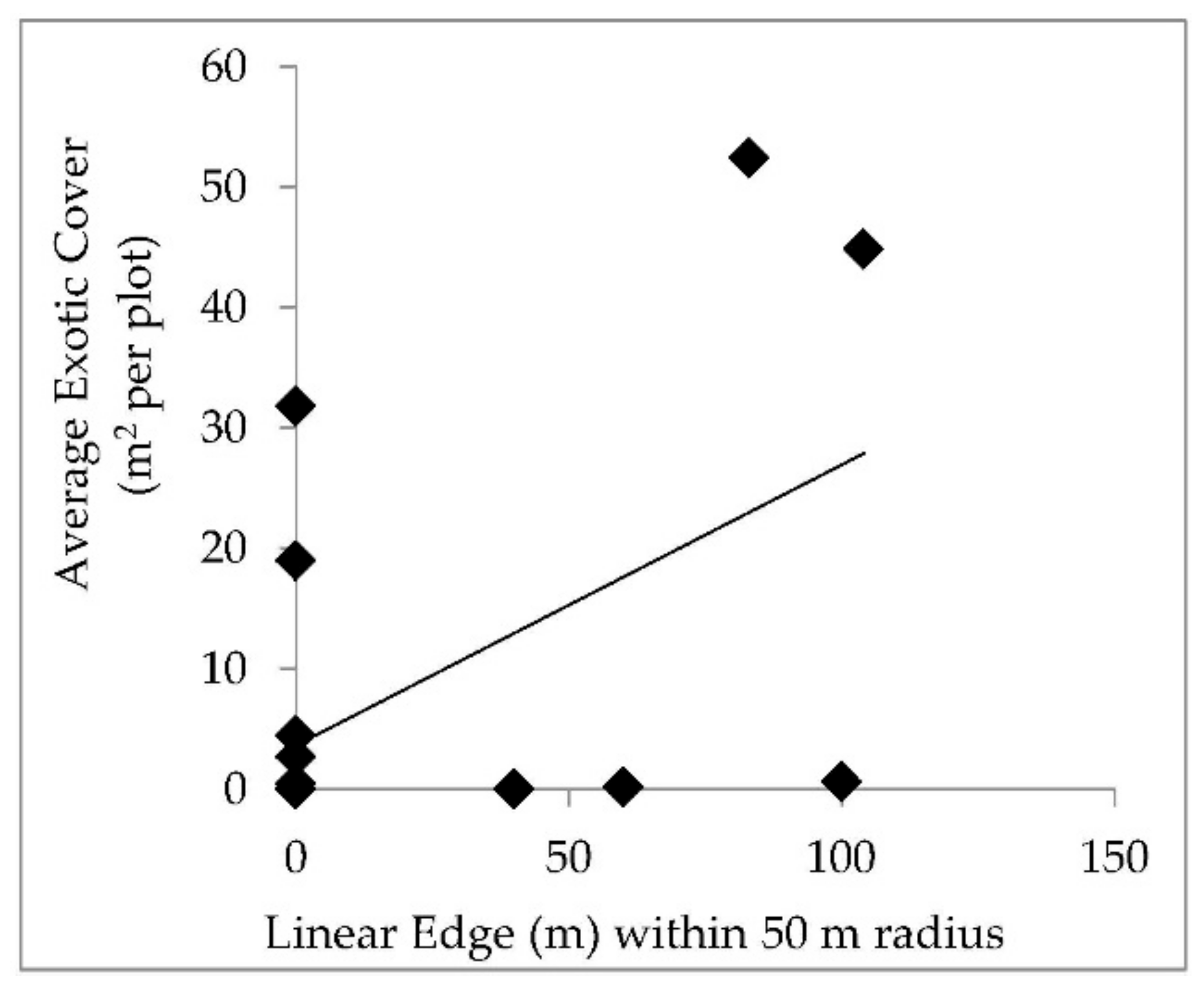
The suggestion that windthrow did not create conditions that favor exotic species is further supported by the lack of a positive relationship between linear edge and exotic plant cover or richness following disturbance events, which contradicts expectations for our second hypothesis. Baseline exotic plant cover independent of the recent storm disturbance was correlated to linear edge within 50 m and 100 m radii surrounding plots. Yet, plots that likely have more exotic propagule pressure and seed banks did not experience increased exotic cover following disturbance. The short window between the disturbance and sampling of these forests may not have been enough time for plant cover to respond to canopy disturbance. The positive relationship between species richness and nearby forest edge habitat (within 50 m) following disturbance may reflect the propagule pressure coming from the edge habitat, which lead to the appearance of new exotic species in the plots.
Invasive exotic plants are generally considered to be fast colonizers of disturbed land [
8,
44]. Despite this, it is possible that the speed at which these plants colonize disturbed sites may not have been rapid enough to be detected during this study. The majority of these plots were sampled within two years of the disturbance events. Trends observed in this window of time following the disturbance may not necessarily reflect the future trajectory of these disturbed forests. Over time, we except that propagule pressure will play a major role in the expansion of exotic plants and will continue to affect these forests in the future. Seed sources include existing edge habitat, river systems that cut through each of these NPS units, pre-existing and now expanding human populations in surrounding lands widely planted with exotic horticultural plants, and increasing recreational visitation to parks [
45]. Further complicating matters, lag times in exotic plant invasions that operate on timescales ranging from years to decades are difficult to account for, but can have an important role in the establishment and expansion of invasive exotic plants [
46]. Once exotic species become established, they can maintain a foothold as the forest recovers [
3,
5,
8,
47]. Future sampling of these long-term vegetation monitoring plots will not only aid in understanding the trajectory of plant community assembly of these forests, but could assess the power of the short-term responses observed here as indicators for future composition of preserved forests following natural disturbance.
Continued study of variability in vegetation community, soils, topography, and land use history of the disturbed sites may lend insight in the short-term responses reported here, as well as document vegetative changes that will occur over the long term in these forests. The six plots in GARI and NERI that contained zero exotic species before and after disturbance also merit further investigation. Less adjacent edge habitat and relatively light historical land-use disturbance for these parks are likely contributing factors in preventing exotic species establishment. However, investigating the link between higher native diversity and increased resistance to invasion, as observed by Gilbert and Lechowicz 2005 [
48] and others, may help to understand which factors contribute forest resistance to exotic plant encroachment [
49,
50,
51]. Understanding which forest habitats and conditions are most vulnerable is critical to informing management of these forests following disturbance events [
52].
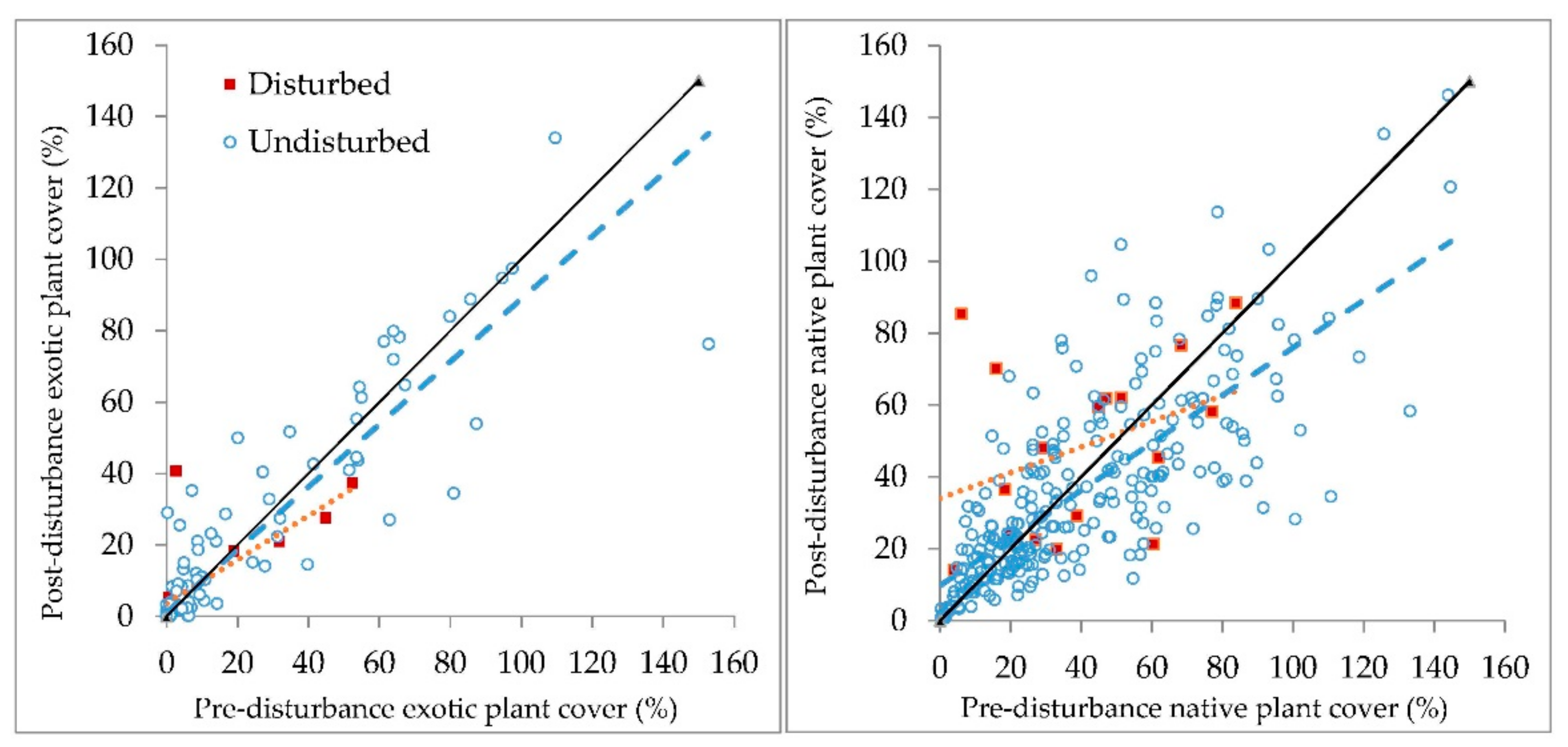
In contrast to exotic plant species in this study, native plants showed high temporal and spatial variability. For example, native plant cover in undisturbed plots measured before 2012 only explains 57% of the variance of native plant cover when the plots were resampled (
Figure 4). This variability may be the result of year-to-year variability in native plant cover that could be driven by factors such as annual variability in weather or population dynamics of annual species; the richness of understory plants in similar forests have high temporal variability [
53]. Abiotic measures of light, gap size, temperature, and moisture would offer much-needed insight into the nature of the understory plant responses observed.
If windthrown tree tops and branches shading the forest floor limit the expansion of exotic plants into disturbed areas of these forests, these benefits would be lost by salvage logging. Salvage logging is common practice on forested lands following disturbance events [
26,
54]. Not only are tree trunks removed in salvage logging operations, but tree limbs and canopies are generally stacked in piles allowing much more light to hit the forest floor than in a non-salvage logged site. Salvage logging also tends to bring in invasive exotic plant seeds directly through logging equipment [
44,
55]. Fallen trees play an important role in forest resiliency following disturbance [
26,
56]. In our disturbed plots we measured increases in coarse woody debris on the forest floor following disturbance and no rapid response of invasive species. These plots are within National Park Service land, where salvage logging is typically not employed to protect the parks’ natural resources.
Natural disturbance events such as wildfires, flooding, and intense storms in forests are expected to increase with continued changing of the climate [
56,
57]. As land managers work to promote ecological integrity within managed lands, understanding the impacts of projected increases in disturbance regimes [
57] is critical. Increased temperatures and changes in annual precipitation will likely further complicate native forest vegetation recovery following disturbance.
Protected areas comprise 14.8% of the Earth’s land area and 13.9% of the United States [
58]. However, conservation practices do not always translate to preservation. In the eastern United States, many of the conserved forested areas owned by local, state, or federal agencies are still actively managed for timber and other anthropogenic uses. The findings of this study underscore the importance of how preserved lands are managed during forest recovery following natural disturbance events. Lands set aside from direct anthropogenic exploitation may be resistant to exotic plant invasive following disturbance because of the initial shade provided by the debris.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






