1. Introduction
When a competition index is used to describe an individual tree’s social status and quantify the surrounding environment, it is labeled as one-sided competition [
1]. A two-step procedure is generally used for constructing one-sided, distance-dependent, or position-dependent competition indices [
2]. The first step determines the count and which trees are labeled as competitors, while the second step determines their size and distance. The closer and larger the competitor tree is to the subject tree, the larger is the index. Several authors have indicated the superiority of selecting neighbor trees for the first step with angle count sampling (also known as point sampling, variable plot sampling, or the Bitterlich method [
3,
4]). Other authors [
5,
6] have presented alternative methods for selection of competitor trees in forests of
Eucalyptus globulus and
Betula pendula, respectively. It is affirmed, however, that any fixed-radius method proves disadvantageous when it is utilized in long-term growth studies [
2].
Spurr [
7] defined a modified angle summation estimate of tree competition that is dependent upon the spatial distribution and relative size of trees counted at a point. In particular, Spurr [
7] contended that the Bitterlich [
8] angle count method presents a drawback in that a cluster of large trees close to the sample point could present a tally identical to the count of relatively smaller trees distributed around the periphery of a wide circle. Spurr [
7] proposed measuring the angles subtended by sample trees surrounding the test point to account for the density influence of large stems and stems closer to the sample point. To accomplish this, the Spurr [
7] methodology treats each sample tree as a borderline tree and computes a basal area type or borderline factor for the individual tree. The borderline factor (
BLF) can be expressed as
BLF = (d/
R)2/4, where
d is diameter at breast height (cm) and
R is the distance to the center of the tree (m) from the point (plot) center. While the Spurr [
7] angle summation technique has found some popularity as an individual tree competition index for growth and yield models [
9], its original formulation cannot be used to duplicate a random sample of stand density in terms of basal area. Rouvinen and Kuuluvainen [
10] derived an angle summation competition index that is based on the value of arctan (
d/
R). The advantage of the index by [
10] is that the measurement equals π/2 as
R → 0, whereas
BLF → ∞ as
R → 0.
It has been suggested that the separate measurements of
d and
R could be omitted for determining the
BLF by using the Wide Scale Relaskop or Telerelaskop to directly measure the subtended angle of the sample tree formed by the ratio
d/
R [
11]. As displayed in
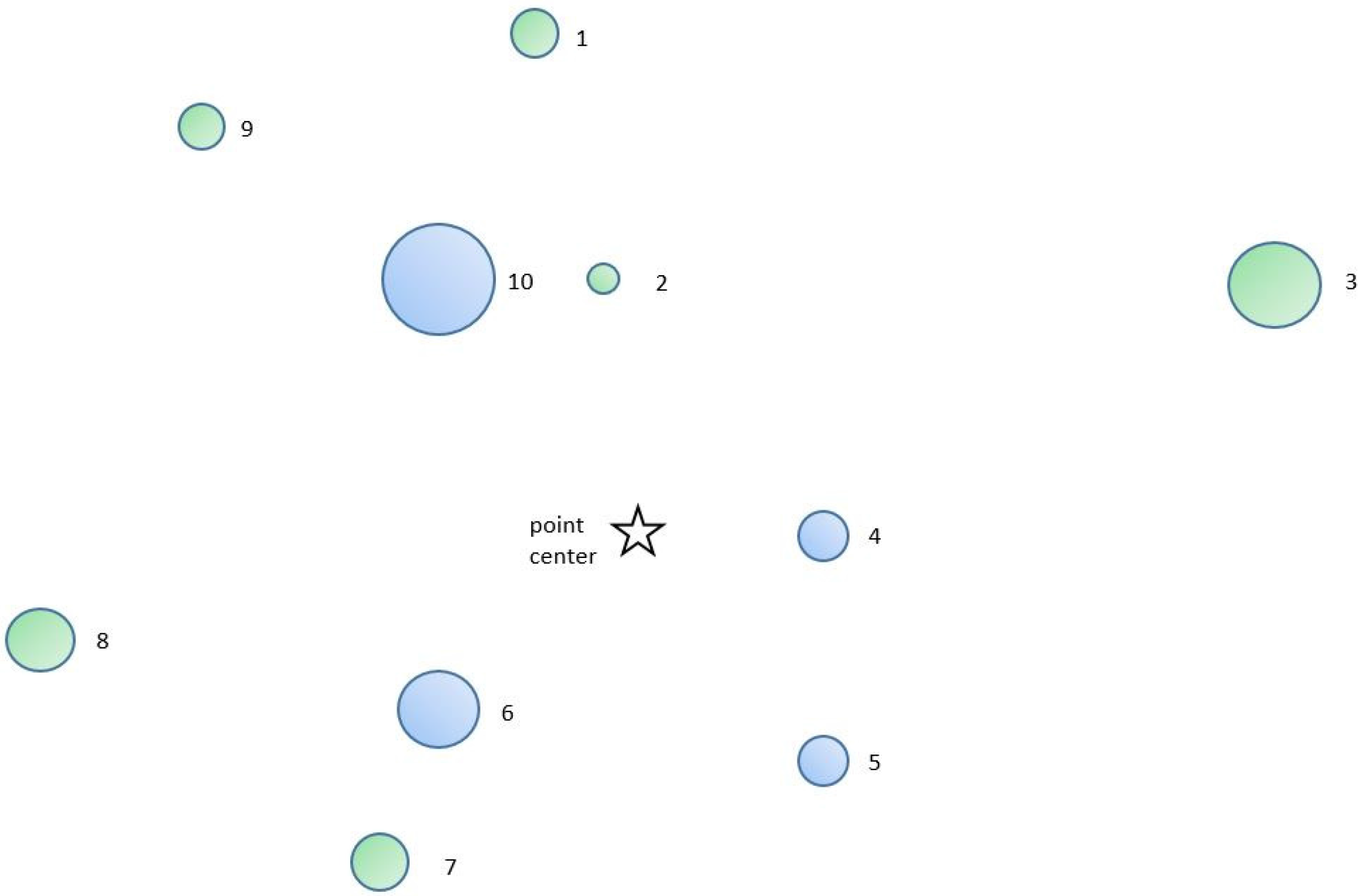
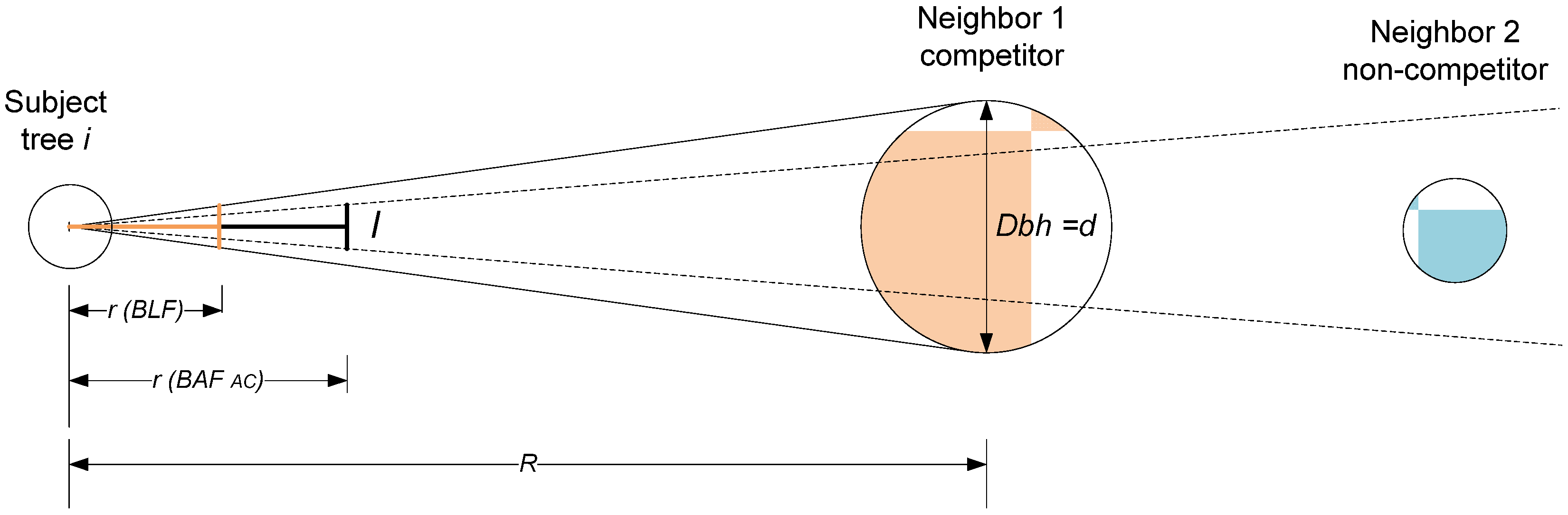
Figure 1, it is possible to use a much simpler stick angle gauge to sight on a sample “in” tree to determine the
BLF in forests of sparse understory. It should be possible to slide the crossarm or sighting intercept back to the point of
r (
BLF) so that the angle is coincident with the angle formed by diameter at breast height (Dbh) and distance
R. It is then easy to devise a scale on the stick that directly furnishes the borderline factor of
BLF or (
d/
R)2/4. Experience in southeastern United States, however, reveals that the Haglöf DME 201 ultrasonic distance measuring instrument is ideally suited for quickly measuring distances to the nearest 0.01 m without walking from the subject tree to the competing “in” tree. In forest conditions with dense understory vegetation consisting of shrubs and arborescents, the distance of many candidate trees that are further from the point center must be verified, since it is difficult to visually verify if the neighboring tree is included “in” the sample. The added time to measure the distance of close-to-point center trees is relatively small.
Numerous authors have discussed the demise of the use of permanent sample plots in forests of North America, based on an angle count (point sampling) estimator, for measuring forest stock and growth. Point sampling estimators have been considered appropriate for measuring growth, but this has been debated [
13,
14]. For example, the ubiquitous installation of permanent point sample plots in the 1960s and 1970s in North America was based on the low cost, ease of measurement, and efficiency of basal area and volume estimates. The disfavor of permanent point sample points became evident later when analyzing growth from remeasurements as additive or compatible estimates, such as those reported by Van Deusen et al. [
15]. These estimates typically displayed high variance for the attribute of growth, while the efficient change estimator of Grosenbaugh was non-additive for biomass and/or volume (
V2 ≠ [
V1 + Δ
V], where
∆ represents volume change and
Vi is stand volume at time
i [
16]). Based on this evidence, Green [
17] noted that it is difficult to detect much enthusiasm for the analysis of remeasurements using the angle count methodology.
Given the wide use of point sampling and the need to better quantify tree-level competition, this paper counters the arguments of Green [
17] and explores the use of a new estimator and its properties in the estimation of basal area and volume growth from point sampling [
18]. In addition, the prior estimator by McTague [
18] can be decomposed on an individual tree basis to assign a competition index. Consequently, a comparison is made to the growth estimator using the traditional angle count constant for basal area factor (
BAFAC) created by Bitterlich [
8], and the newer variable basal area factor (
BAFV) [
18]. The primary research objectives of this analysis were to: (1) demonstrate the application of a new estimator for quantifying long-term production; (2) develop a new tree-level competition index based on point sampling; and (3) evaluate both estimators in two contrasting plantations and examine behavior of bias, precision, and additivity.
2. Materials and Methods
2.1. Plantation Locations and Tree Attributes
Two locations of loblolly pine (
Pinus taeda L.) plantations were utilized to test the development of the
BAFV competition index. The first location was a 40 ha forest stand measured with a sampling intensity of one point (plot) per ha. The inclusion of “in” trees was determined with a
BAFAC = 4 using the same point location at ages 12 and 16. The population consists of 500 loblolly pine plantation trees in Santa Catarina, Brazil generated from the growth and yield model described by McTague [
19]. The modeled population is derived from a thinned 12-year old stand, grown to age 16, with a site index, based on dominant height, of 20 m (base age 15). Parameters of the population generated from the growth and yield model are presented in
Table 1. Using prediction equations for the 10th and 63rd diameter percentiles of the growth model [
19], Weibull parameters are recovered for the stand that are consistent with the stand-level estimate of basal area and the user-stipulated 500 trees per ha. Several parameter recovery methods for the Weibull distribution are presented in Burkhart and Tomé [
12], including the use of diameter percentiles. In this case, each diameter class contains a proportion of 1/500 or 0.002 of the total trees, and the diameter class assignment is computed with the following formula by Clutter and Allison [
20]:
where
i = 1–500,
di = diameter breast height of diameter class
i, and
a,
b, and
c, are Weibull distribution parameters.
Individual tree volume and green weight from the stump to the top of the tree were computed with the following equations presented in McTague [
19].
where
v = total stem volume of a tree outside (including) bark in m3
w = total stem green weight outside bark in tons
d = individual tree diameter in cm determined at breast height of 1.3 m
h = total tree height in m
The attributes of crown width and live crown ratio were predicted with equations localized for dominant trees of loblolly pine in Rio Grande do Sul and Santa Catarina, Brazil [
21]. Crown size was used for computing an alternative competition index. The live crown ratio for individual trees was computed by adjusting the predicted live crown ratio by the ratio of individual tree height to Lorey height, while the predicted crown widths of dominant trees were adjusted with the ratio of diameter (
d) to quadratic mean diameter (
dg).
Each of the 40 ha in the simulation at ages 12 and 16 contained the same population of 500 trees. The trees were randomly sorted for each ha and assigned once, at age 12, to grid coordinates. The spacing between rows was maintained at 2.5 m, while the distance between trees in the row was randomly perturbed. Since the simulated stand had been thinned with a combination of row and low thinning, every fourth row was completely removed. The simulation procedure was repeated 500 times using a 40 ha forest stand and a sampling intensity of one point (plot) per ha with multiple random starts within and between iterations. There was, however, no random perturbation associated with tree size. A small random perturbation, associated with the point (plot) location, was also simulated. The edge-effect and the potential bias associated with point locations close to mapped canopy openings or a forest edge were not investigated here as the point locations were at least 12.5 m away from the property boundary in this study.
The second study site was located in a 17-year old loblolly pine plantation in the Piedmont physiographic region, Warren County, Georgia, USA. The 13.1 ha forest was completely stem-mapped following a thinning operation. The tract was divided into 135 quadrants with dimensions of 31.15 m × 31.15 m. All trees were tagged and had a measurement of d. Total tree heights were systematically measured on half of the stems in a quadrant. A common localized height-diameter equation was fit to the measured diameters (ds) and heights (hs) of each quadrant of the form: ln(h) = a0 + a1·(1/d), where a0 and a1 are estimated coefficients. Using the quadrant specific values of a0 and a1, trees without observed heights were then assigned a predicted height value.
Total stem green weight outside bark (
w) in kg was estimated with the following equation from Harrison and Borders [
22]:
The attribute of crown width was estimated with coefficients specific for loblolly pine [
23], while live crown ratio was predicted with the equation derived for loblolly pine in southeastern United States [
24].
Remeasurements were not conducted for the southeastern United States plantation. However, the age 17 population parameters are provided in
Table 1. Several salient features of the second study location are demonstrated in
Figure 2, including the appearance of planting rows with considerable curvature and almost random direction. Large gaps in the forest canopy and non-planted openings were also visible, which leads inevitably to high variability and sampling error. Systematic sampling was conducted for the 13.1 ha forest stand using a sample size of 30 points (plots) with a
BAFAC = 4. The procedure was repeated 500 times using multiple random starts within and between iterations. For the sake of avoiding the placement of point (plot) locations too close to the long and narrow tract perimeter, there was no random perturbation of point centers from the multiple systematic grids. Point locations were at least 16 m away from the property boundary in this study location.
2.2. The Application of a New Estimator for Quantifying Long-Term Production
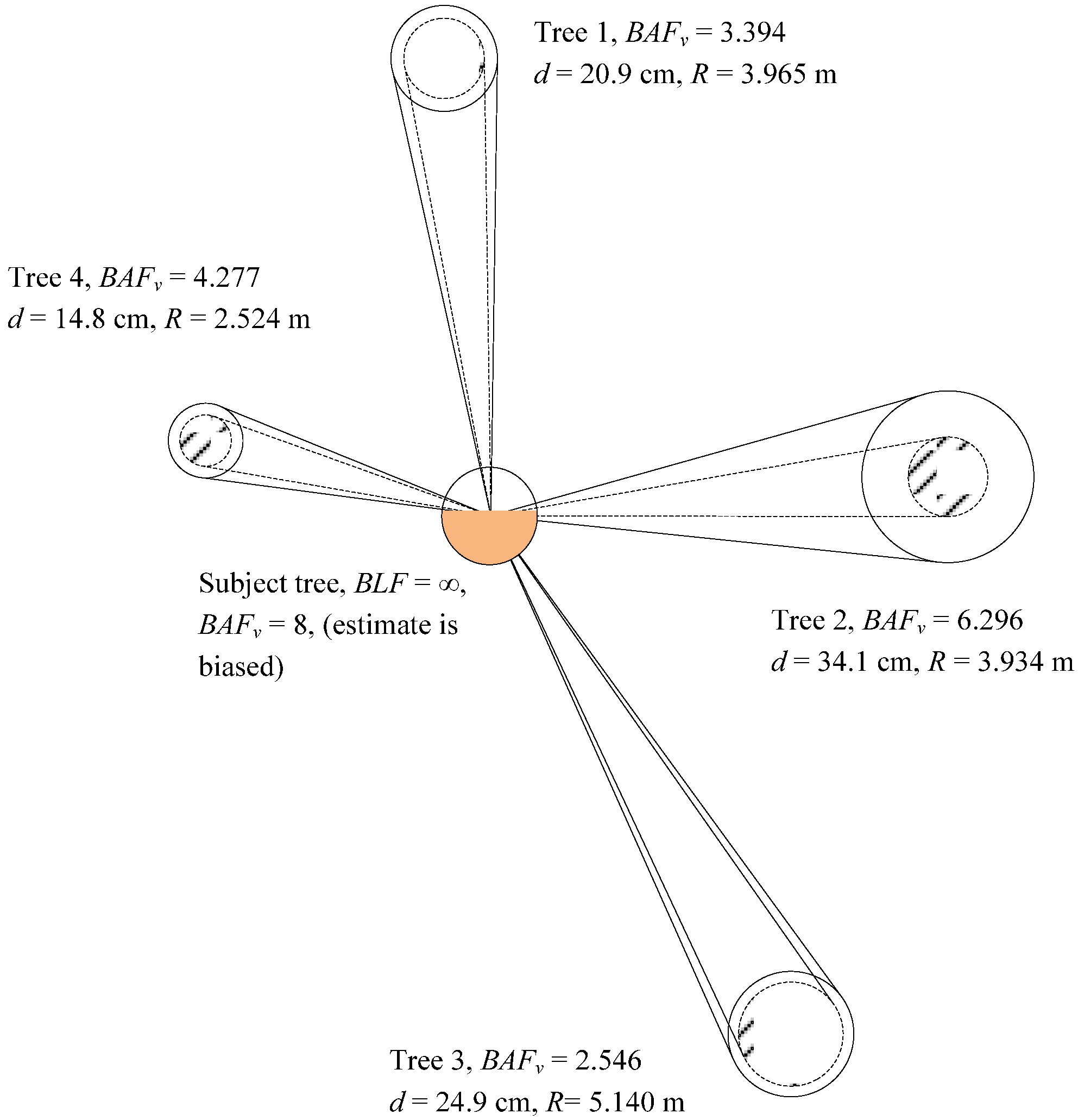
The proposed estimator will be demonstrated to be ideally suited for a system of permanent sample plots (PSPs) using a point sampling methodology for selecting “in” trees at each location. The point (plot) locations themselves are generally based on a probability based design. The change statistics in this study for attributes such basal area, volume, and stem density, were computed using only subject trees of the PSP points. These subject trees were selected by using the Bitterlich angle count basal area factor (BAFAC).
The attributes assigned to subject trees were not computed with the constant
BAFAC. Rather, the estimator is a ratio that employs elements of both the Bitterlich angle count sample and the Spurr borderline factor, specifically (
1−BAFAC/
BLF). The new variable basal area factor estimator (
BAFV) was presented by McTague [
18] as:
where
m = the number of “in” trees at the point (plot)
G = the basal area in m2·ha−1
BLFi = the borderline factor for tree i, or (di/Ri)2/4
di = diameter at 1.3 m (Dbh) in cm for tree i
Ri = distance in m between the sample point and center of tree i
BAFAC = the Bitterlich angle count (point sampling) constant
Stem density, N, was estimated as , where gi is the individual tree basal area calculated as .
Basal area, and weight per ha, W, was predicted as where wi is individual tree green weight outside (including) bark. The basal area value for borderline “in” trees approaches zero, while trees close to the point center have a basal area approaching a value of 2 (BAFAC) with the new estimator. Large trees have more influence on the attributes of basal area, stem density, and volume per ha. Estimates were computed by using the BAFV and BAFAC approaches for both the 16-year old plantation in Brazil and 17-year old plantation in USA in order to compare the results obtained by both methodologies. Yield and variance statistics of repeated sampling for the stand-level attributes of basal area (G), stem density (N), and green weight per ha (W) were computed from analysis-of-variance-type calculations of the between repetitions component.
2.3. Improved Compatible Estimator for Stand-Level Growth
The work of Hradetzky [
25] was extended by McTague [
26] whereby empirical comparisons of bias and precision among growth estimators from permanent point samples were presented. The problem of estimating growth from permanent point samples, as described by [
25], consisted of varying sample composition from one measurement occasion to the next. Excluding the consideration of trees that died, or saplings that grew into merchantable size during the remeasurement interval, the sample plot on each remeasurement occasion would normally contain an augmented list of “in” trees. What differs among various point sampling growth estimators is the computation of the basal area and expansion factors for the new “in” trees at the time of remeasurement.
The original list of “in” trees is generally referred to as survivor trees, since they are measured “in” and are alive on both measurement occasions. Several authors preferred to allocate a basal area and expansion factor of zero to the new “in” trees [
16,
27]. Expanding upon the notation of Hradetzky [
25], unbiased change estimators for green weight per ha of a pure panel design are presented below:
where
ZAC = change in stand green weight (t·ha
−1) using the additive estimator [
15] of angle count sampling
wij = green weight for tree i at time j
gij = individual tree basal area in m2·ha−1 for tree i at time j
mj = number of sample “in” trees at a point (plot) at time j. The number of new “in” trees at the time of remeasurement equals (m2 − m1)
The most widely used estimator for computing change in biomass/volume from point samples is the non-additive Grosenbaugh estimator [
16], which essentially ignores the contribution of the new “in” trees at the time of remeasurement into the calculation of growth:
where
ZG represents the change in green weight (t·ha
−1) using the non-additive Grosenbaugh estimator.
The change estimator by McTague [
26] using
BAFV of Equation (5) is expressed as:
Since the BAFV is very small for a new “in” tree at the time of remeasurement, the ZV change estimator is very efficient, as virtually all the change is attributed to the growth of survivor trees. The equations for ZAC, ZG, and ZV were used to compute the growth of stand-level green weight W. When wij was replaced with gij in Equations (6)–(8), stand-level growth of basal area G was computed.
2.4. Alternative Measures of Individual Tree Competition
Given that the estimator
BAFV was partially comprised of the borderline factor (
BLF), it is only natural to believe that it is highly correlated with the Spurr point density index [
7]. The Spurr point density competition index (
CIPD) requires that the (
n) sample “in” trees be ordered by decreasing values of
BLFj, or (
dj/
Rj)
2/4, and it is expressed as:
The
BAFV also proves to be correlated to other distance-dependent indices. The Hegyi index (
CIH) [
28], as applied by Daniels and Burkhart [
29], was used in a point sampling context and it is a function of size of competing trees and distance:
where
di = Dbh in cm of subject tree i
dj = Dbh in cm of competitor tree j
Rij = distance in m between subject tree i and competing tree j
n = number of trees included “in” the sample for a given BAFAC (excluding the subject tree)
One apparent drawback of both CIPD and CIH is the property that the indices predict values approaching infinity as Rij becomes increasingly smaller. This is particularly problematic for stems that are forked below breast height (1.3 m) as the distance between subject tree i and competing tree j is often less than 0.5 m.
It is possible to demonstrate that the sample tree expansion factor (
BAFV/
g) is also a function of tree size and distance (the expansion factor is used to compute the representative trees per ha of each sample tree of an angle count plot—this is unrelated to the linear expansion factor of Biging and Dobbertin [
30] which is used as an adjustment for plot edge bias). Smaller trees of diameter (
d) located near the angle count limiting distance, namely,
, contribute less to stand weight than larger trees located near the sample point. It was hypothesized that the variable stand weight estimator, computed as the product of individual tree weight (
w) and the expansion factor, might be correlated with the crown volume competition index [
30]. The crown volume unweighted by distance (
CVU) index was formed by projecting a 50° angle from the base of a subject tree and computing the crown volume of competing trees calculated at the point where the height angle intersects the tree bole axis. The crown is assumed to have a profile of a cone. The crown volume index is computed as:
where
CVU = unweighted crown volume index
CVaj = crown volume of competitor tree j above the point where the height angle cuts in height axis
CVi = crown volume of subject tree i
n = number of trees included “in” the sample for a given BAFAC
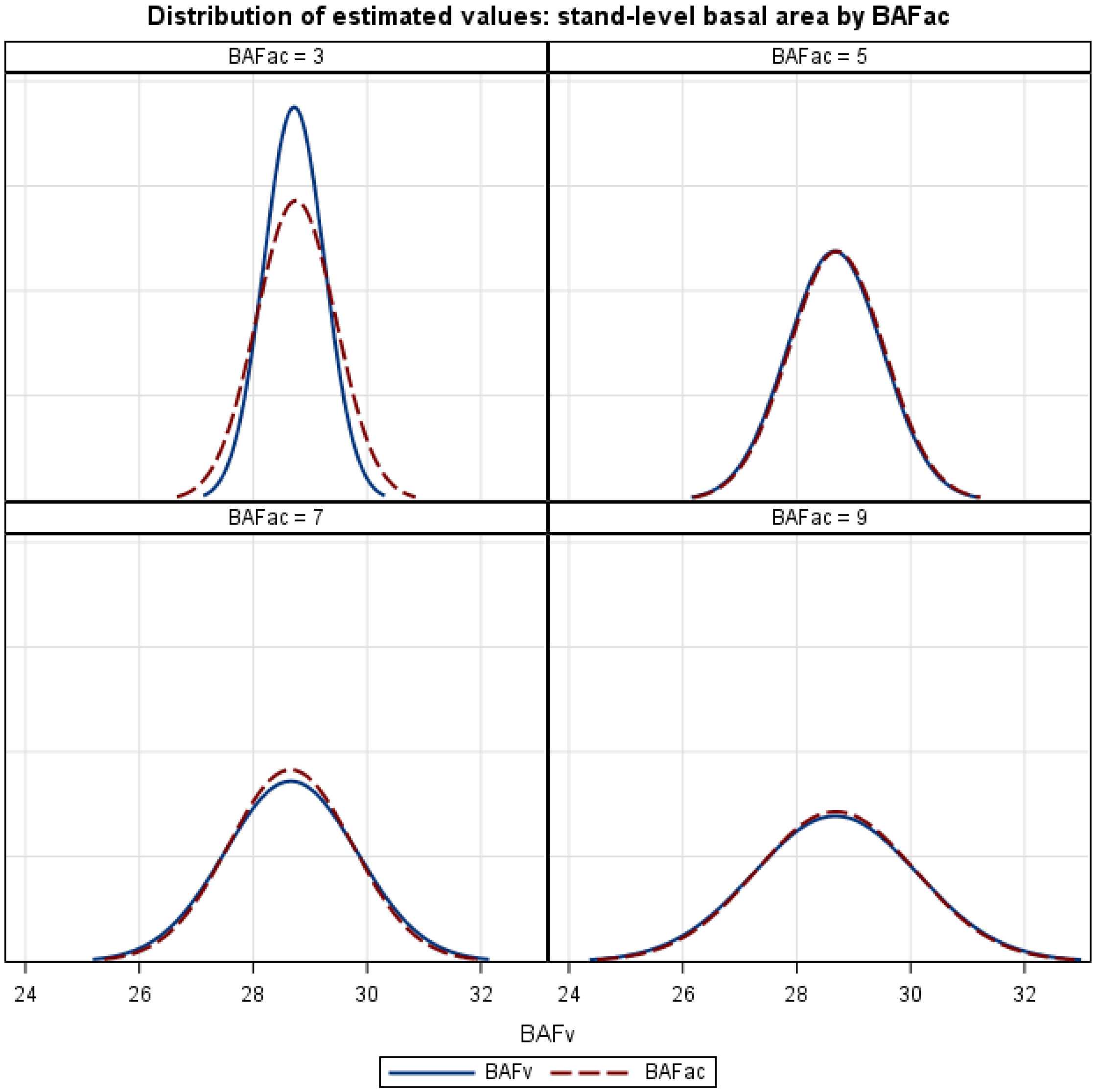
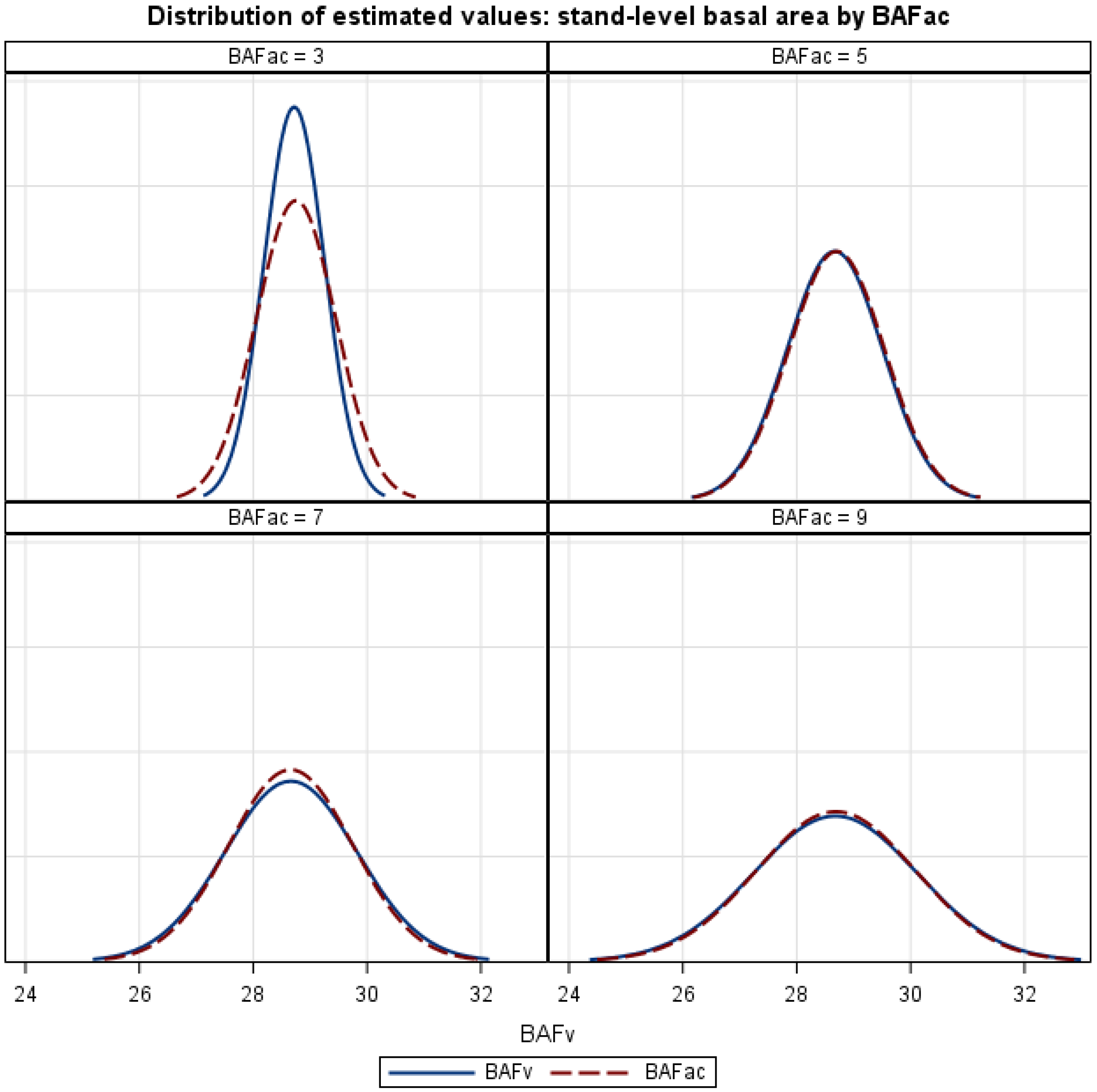
4. Discussion
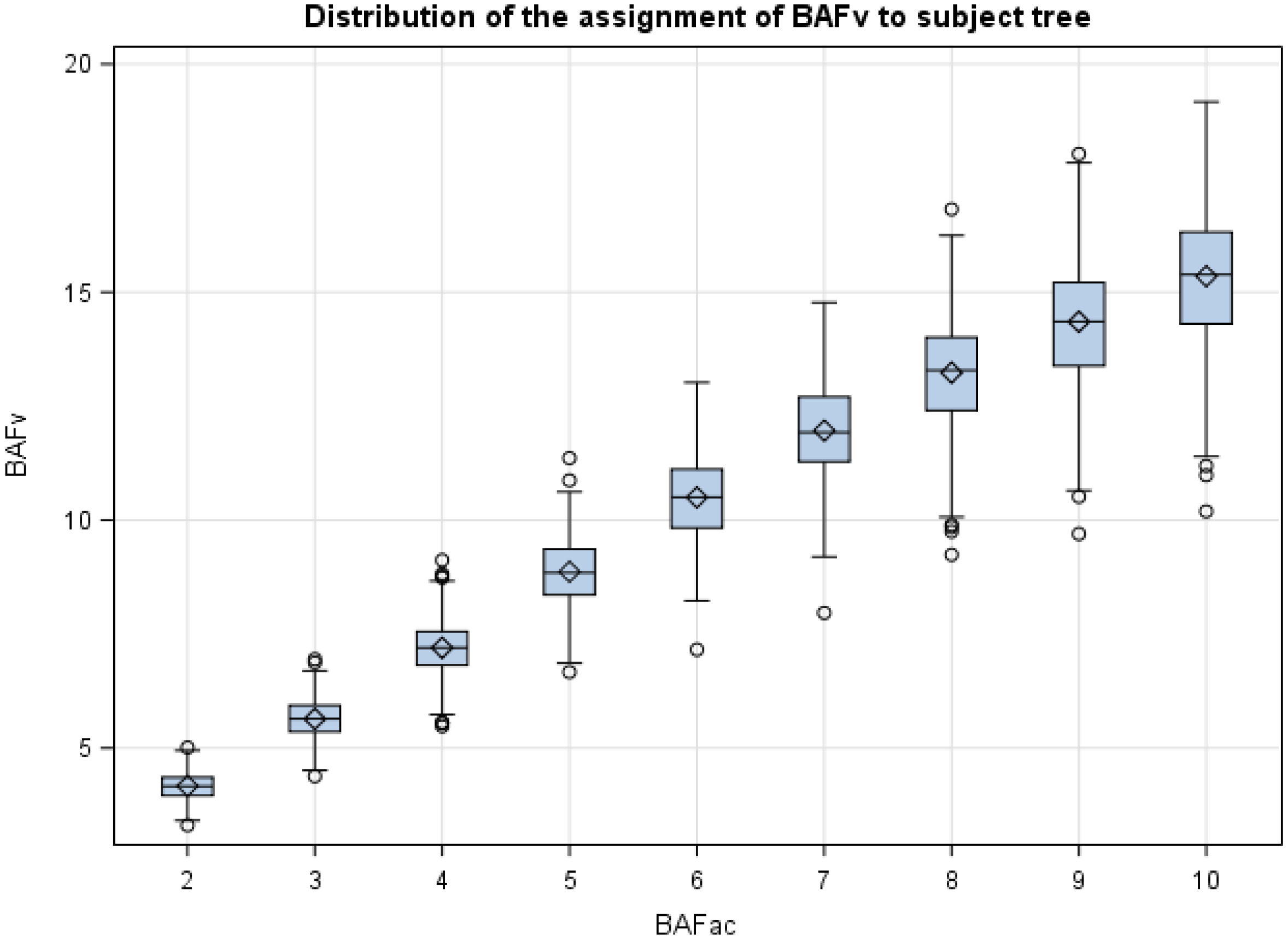
If tree diameter (
d) and distance from the sample point to the subject “in” tree are measured during forest inventory, it is possible to compute the borderline factor (
BLF) and the variable basal area factor
BAFV of Equation (5). As displayed in
Figure 5, the
BAFV estimator is unbiased, implying that over many points, the expected values for the variable
BAF estimator and the Bitterlich angle count estimator (∑
BAFAC) are identical. The
BAFV estimator also has desirable features, such as additivity and efficiency, when used for estimating change with permanent plots. The general consensus of the literature suggests that the Grosenbaugh growth estimator, Equation (7), is more efficient than the additive estimator, Equation (6), for most forest conditions and remeasurement intervals [
25,
31,
32]. The drawback of the Grosenbaugh estimator is that it is non-additive (incompatible), so that volume (
V2 ≠ [
V1 +
ΔV]). The disadvantage attributed to using the additive (compatible)
ZAC change estimator of Van Deusen et al. [
15] is manifested in terms of high variance, which is normally attributed to the list of new “in” trees at the time of remeasurement [
25]. The other alternatives,
ZG and
ZV, are superior in providing efficient change estimates for basal area and volume per ha. If the goal is to monitor forest growth, the variable and additive
BAF estimator,
ZV, offers an attractive alternative to the common non-additive estimator of
ZG.
Quantifying the competition surrounding the subject tree with the BAFV estimator was advantageous because the effect of size and distance of neighboring trees is reported in absolute units of m2·ha−1 (or t ha−1 in the case of green weight). Most competition indices, such as Hegyi, Equation (10), or the CVU, Equation (11), do not include the contribution of the subject tree in the competition computation, since the value of the subject tree equals infinity. Of course, this drawback is not true for the BAFV estimator. In contrast, the contribution of the subject tree to the competition index based on BAFV must be included if the sum of the individual competition values are to be additive with the stand level estimate of basal area or volume.
Following the establishment of the permanent sample plots (points), neighboring competitor trees that are centered around each subject tree can also be measured. As a practical matter, a
BAFAC < 3 m
2·ha
−1 is not recommended. It was desirable to keep the total sample size of subject and competing trees manageable by using a
BAFAC ≥ 3. One recent investigation arrived at the conclusion that the Bitterlich angle count method was not the best method for selecting competitor trees [
6]. Here, we contend that Bitterlich method should not be used exclusively for just selecting competitor trees, but rather the point sampling expansion factors should also be employed to scaling the competition indices up to a per ha level. If the expansion factor of (
BAFV/
g) is used, the competition indices are both spatially explicit and can express the competition in absolute values at a per ha level.
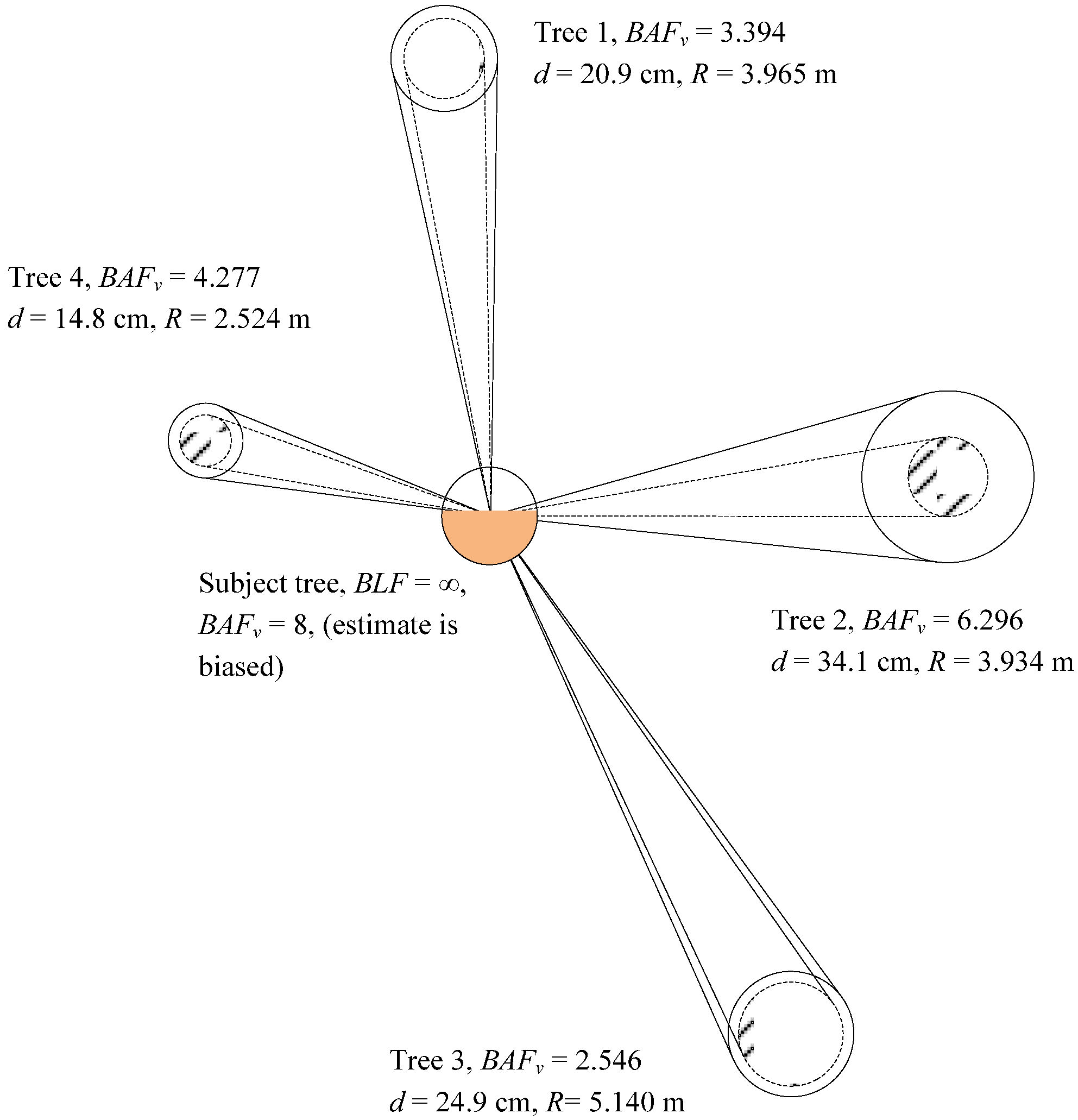
A drawback of the BAFV estimator for measuring competition is that it is biased for measuring the competition contribution of the subject tree. This paper presents a methodology for arriving at a stand level value for BAFV for the subject tree that removed the bias. The proposed method then assigns the same BAFV value to all subject trees over all points (plots) in the stand. Compatibility or additivity is achieved at the stand level, but not at the point or plot level. Compatibility is mathematically possible at the point (plot) level, however, it may result in either the negative assignment of BAFV for the subject tree, or the need to adjust the values of both subject and competing trees. Clearly, the drawbacks of achieving compatibility at the point (plot) level outweigh the possible benefits.
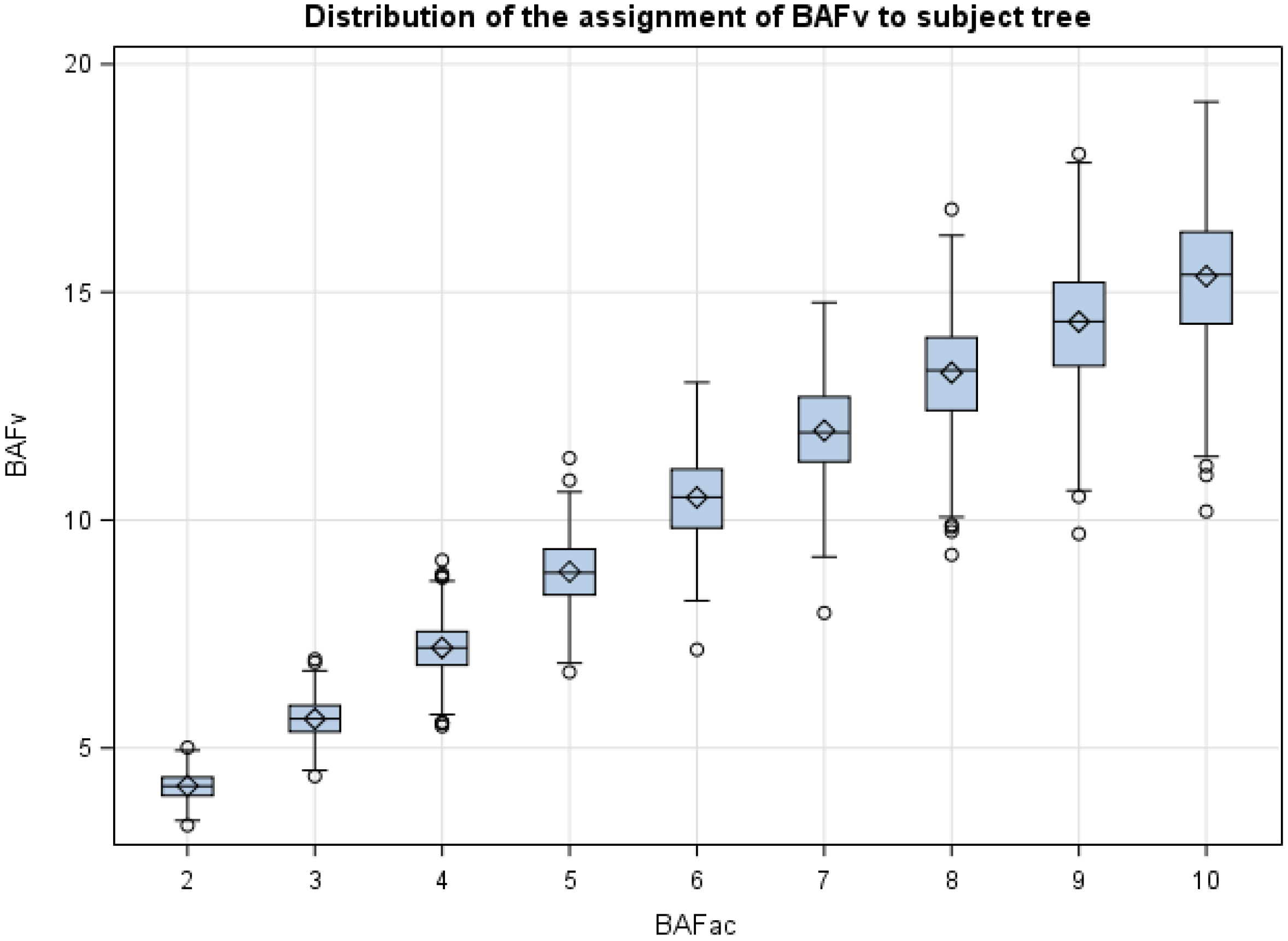
One puzzling outcome of this study was the difference of 2.83 m2·ha−1 (7.19 m2·ha−1 vs. 4.36 m2·ha−1) between the assignment of the average unbiased BAFV value for the subject tree, using a BAFAC = 4, when comparing southeastern United States and Brazil. Clearly the variability in the simulated Brazilian plantation was less pronounced than the observed conditions in southeastern United States, with large holes in the forest canopy and frequent null tallies at some points (plots) in the latter. In hindsight, the study would have benefited from a larger stem-mapped forest in the United States, or with a more equal distribution of forest canopy holes between the center and periphery of the stand. Without question, the assignment of 2(BAFAC) to each subject tree results in a positive bias, irrespective of the forest structure and conditions.
The additive competition indices proposed here are well suited for growth and yield studies that rely on PSPs, where it is imperative that change statistics are estimated efficiently. In this case, the BAFV estimator is perfectly suited for measuring growth. In addition, it is well known that angle count samples are inexpensive to install and maintain. With a selection of a BAFAC ≥ 3, it should be physically and economically feasible to measure all competitor trees and obviate the requirement for edge correction. A difficulty can arise, however, when using a BAFAC ≥ 3, if the forest is subjected to intermediate harvests that involve strips or mechanical row thinning. Depending on the width of the harvest cut, a large proportion of the subject and competing trees of the point can be eliminated, compromising the utility of the remeasured plot over the next measurement interval. This is particularly true for forest stands where the quadratic mean diameter (dg) is less than 15 cm and the equivalent plot size is rather small. As trees grow larger in size and the effective plot size increases, the abrupt change in sample size associated with cutting diminishes.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}