Abstract
Anthropogenic disturbances interact with wildfire, altering successional dynamics across North America’s boreal forest. Linear disturbances, including seismic lines used for oil and gas exploration, dissect forests, while wildfire is a fundamental agent of forest succession. However, little is known about early succession dynamics after both seismic line creation and wildfire, especially across transitions from uplands to peatlands. To address this, we characterized and compared regeneration and recruitment after individual and successive disturbances in peatland, transitional, and mesic upland forests across the oil sands region of Alberta, Canada. We used non-metric multidimensional scaling to compare composition and mixed-effects generalized linear models to compare densities of trees and tall shrubs 10 to 24 years after disturbance. Compositionally, regeneration was similar within forest types and between transitional and peatland forests, while patterns in recruitment were more influenced by past disturbances. Overall, we found evidence of dominant, additive, and interactive effects on early successional patterns within linear disturbances in boreal forests. In transitional and peatland forests, disturbances influenced tree and tall shrub regeneration and recruitment in complex ways. Early successional dynamics after disturbance influence forest structure and composition and are vital to understanding recovery in boreal forests, especially across boreal forest transitions.
1. Introduction
Successional dynamics in North American boreal forests depend on the severity and frequency of disturbances and have primarily been characterized after single stand-replacing disturbances [1]. Historically, natural disturbances, especially wildfire, have determined boreal forest structure, composition, productivity, and biodiversity [1,2]. Early succession begins with a regeneration and recruitment phase [3,4]. During regeneration, there is rapid initial growth of grasses, herbaceous vegetation, and woody shrubs, but recruited trees begin to outcompete the initial colonizers during the young forest regrowth phase within 10 years post disturbance [3,4]. While successional dynamics are well known for natural disturbances, such as wildfire, increasingly anthropogenic disturbances are resulting in more frequent interactions with natural disturbances, potentially altering successional dynamics [5].
Forest structure and early successional dynamics are influenced by successive disturbances, such that the rate of spread and severity of subsequent disturbances depend upon previous disturbances [6,7]. Relatively small changes in forest contiguity during initial disturbance can change subsequent disturbance behaviors, such that large, nonlinear, or unexpected effects can occur [5,7]. Under these novel disturbance regimes, unexpected shifts can occur in successional dynamics, especially if forests have not developed ecosystem resistance or resilience to these novel conditions [8]. It is therefore essential to characterize the effects of successive disturbances to determine whether successional dynamics might change under these conditions. Cumulative effects from successive disturbances can be additive, depending on the sum of individual disturbances, dominant where the effect depends primarily on one disturbance, or interactive where cumulative effects between disturbances are greater or smaller than thresholds calculated from individual disturbances [9].
Across western North American boreal forests, anthropogenic disturbances have contributed to the degradation of millions of hectares of forest, as they interact with each other and with natural disturbances [2,10]. Some of the most extensive fragmentation in boreal forests results from linear anthropogenic disturbances related to oil and gas development [10]. In the Athabasca oil sands region in Alberta, Canada, exploration to delineate underground oil and gas deposits has created narrow (3 to 10 m), linear forest clearings called seismic lines [11]. Although the footprint from seismic lines is only ~2% of the region, the functional footprint of seismic lines associated with forest edge effects can be as high as 60%, contributing to forest fragmentation and degradation of habitat for threatened woodland caribou Rangifer tarandus caribou [12,13]. Compared to other linear disturbances, like roads, pipelines, and powerlines, the length of seismic lines in the region is 5 to 11 times greater [14], with seismic line densities as high as 40 km/km2 or an equivalent 50 × 50 m grid spacing [15]. Immediate and persistent impacts of seismic lines depend on initial physical effects on soil and vegetation from equipment used for clearing, but regeneration on seismic lines can be delayed for more than 30 years, especially in peatland and transitional forests where cold and wet soils inhibit tree establishment and growth of slower-growing species like black spruce Picea mariana Mill. [16]. While best management practices have been implemented to reduce the effects of seismic lines, there continues to be variability in regeneration and recruitment within seismic lines [11].
Transitional boreal forests account for nearly 9% of all wetlands in boreal North America and occur at the interface between upland and peatland forests [17]. Transitional boreal forests vary significantly in species composition, organic layer depth, hydrology, topography, carbon dynamics, and function [18]. This is further complicated as margins of transitional boreal forests change over time [18]. Variability within transitional boreal forests is influenced by topographic, nutrient, and soil gradients from uplands to peatlands [18]. As such, classification of transitional boreal forests can be difficult to determine and vary depending on what characteristics are considered [19,20]. Transitional boreal forests are easiest to detect when occurring within steep productivity and topographic gradients, such as between upland and peatland forests margins [18,21]. Despite the complexities associated with classification, transitional boreal forests provide valuable structure and function in boreal landscapes, slowing water flow into adjacent forests during seasonal flooding and providing diverse habitat for wildlife [22]. Although these transitional boreal forest sites do not accumulate significant amounts of peat, they do support carbon accumulation in adjacent peatlands [20].
Across western North American boreal forests, there is a paucity of data related to early succession in transitional boreal forests and, more generally, a gap in knowledge regarding early successional dynamics after multiple disturbances. To address these knowledge gaps, we measured densities and characterized the composition of regenerating (<1.3 m tall or <1 cm diameter at 1.3 m) and recruited (≥1 cm diameter at 1.3 m) trees and tall shrubs (capable of reaching or exceeding 2 m in height) after individual and successive disturbances across boreal forest types. Overall, the objectives of our work were to:
- Characterize regeneration and recruitment of early forest succession in peatland, transitional, and mesic upland boreal forests using tree and tall shrub abundance and composition.
- Identify relative disturbance effects of individual (seismic line or wildfire) and successive disturbances (seismic line followed by wildfire) on regeneration and recruitment densities of trees and tall shrubs.
2. Materials and Methods
2.1. Study Area
The study area (42,400 km2) extends across the oil sands region in Alberta, Canada, within the western boreal forest of North America (Figure 1). Within the study area, wildfires are the primary natural disturbance, but since the early 20th century, anthropogenic disturbances related to oil and gas development have increased in frequency and extent. Within the study area, the estimated total length of seismic lines is 115,577 km, while across Alberta, the estimated total length of seismic lines is 829,838 km [23]. Seismic lines in the study area included avoidance and low-impact seismic lines (57.6% of seismic lines), which are on average less than 3 m wide, and conventional seismic lines (42.4%) that average 5 to 10 m wide [11]. Conventional seismic lines, including legacy lines constructed before 2000 with heavy equipment that cleared lines to mineral soil, are often prioritized for restoration as they have slower recovery compared to low-impact seismic lines, which can regenerate faster [11]. Additionally, due to the spatial scale at which disturbed woodland caribou habitat is assessed (visibility of anthropogenic disturbance on Landsat at a scale of 1:50,000), only conventional seismic lines are identified as disturbed habitat by federal caribou recovery guidelines [24].
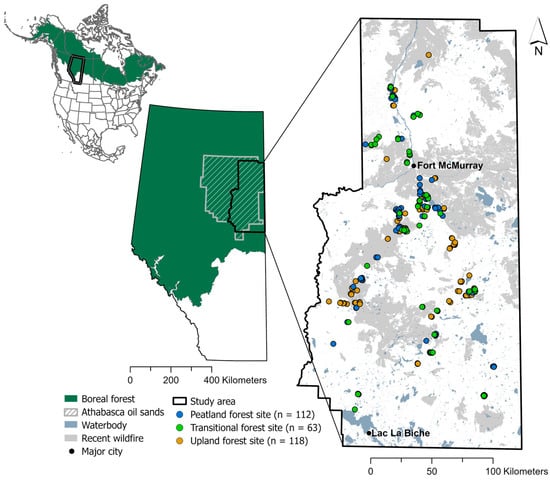
Figure 1.
Sites sampled between 2017 and 2022 across the Athabasca oil sands region of Alberta, Canada, within the North American boreal forest. Study sites symbolized by ecosite and wildfires that recently burned (within 25 years of sampling) are shown.
2.2. Study Design and Field Sampling Methods
In 2022, tree regeneration and recruitment were sampled on passively recovering seismic lines in transitional forests with minimal anthropogenic re-disturbance. This filled a gap in an existing dataset collected in 2017 and 2018 [25,26] (Filicetti and Nielsen 2020; 2022). A total of 293 seismic lines were sampled in this study, including sites in peatland forests (n = 112), transitional forests (n = 63), and mesic upland forests (n = 118). Overstory and understory vegetation were used for ecosite classification at each site following the Field Guide to Ecosites of Northern Alberta [27]. Forest type was then determined based on ecosite classification where peatland forests included bog, poor fen, and rich fen ecosites. Transitional forests included swamp and poor mesic ecosites. While transitional boreal forest communities can range widely, poor mesic ecosites are dominated by black spruce, occasionally jack pine, and feathermoss, with little to no diversity in understories and generally no organic layer on top of mineral soil [27]. Swamp ecosites are temporarily inundated with water tables well within the rooting zone, with little to no bryophyte coverage, organic layer depth less than 40 cm, seasonal water fluctuation, and overstories that include tall shrubs as well as mixed wood, coniferous, and deciduous trees [18,22,27]. Mesic upland forests included medium mesic, medium hygric, and rich hygric ecosites. Sites were sampled proportional to the area of forest types and distributed across the geographic extent of the study area.
All sites had minimal human re-disturbance, as determined by no visual evidence of extensive vehicle trails and no evidence of recent harvest. Re-disturbance was avoided to minimize effects from disturbances other than the initial cutting of the seismic line or recent fire. Sampled seismic lines varied in width from 2 to 16 m, with an average width of 6 m. Burned sites were those that were located within perimeters of recent wildfires that had burned within 25 years of sampling as determined by Government of Alberta historical wildfire spatial data and had between 5% and 100% overstory mortality detected in adjacent forests during sampling, while unburned sites were located outside of wildfire perimeters and had 0% overstory mortality and no evidence of fire in the adjacent forest. In sites where successive disturbances were being sampled seismic line creation occurred first followed by wildfire. Seismic lines sampled were cut between 1958 and 2014, and on average, it had been 24 years since seismic line creation across all sites [23,28]. The average time since the most recent fire across all burned sites was 10 years, with the earliest fire in 1995 and the latest fire in 2016 [29]. The average time between seismic line creation and wildfire was 14 years.
At each site, we used a paired design where we measured densities of regenerating (<1 cm diameter at 1.3 m or <1.3 m tall) and recruited (≥1 cm diameter at 1.3 m) trees and tall shrubs. One plot was in the center of the seismic line, and the paired plot was 20 to 40 m from the edge of the seismic line in the adjacent forest to avoid edge effects but remain in the same forest type. Regeneration density was measured within 1 m to the right side of 30 m belt plots, while recruitment density was measured within 1 m on each side of the 30 m belt plots. Species included trees and tall shrubs capable of reaching or exceeding 2 m. While belt plots centered on seismic lines can underestimate shrub density and height, estimates of tree density and height are similar to plots that extend to the edges of seismic lines, while also being more rapid to survey [30].
2.3. Statistical Analyses
All statistical analyses were performed in R version 4.4.2 [31]. We used multivariate methods to visualize compositional differences among plots where tall shrubs and tree species were present. Additionally, we only included abundant species that occurred in at least 5% of all plots to avoid rare species having disproportionately large influences on dissimilarities [32,33]. Data were stratified by 12 groups, including all combinations of forest type (mesic upland, transitional, or peatland), recent wildfire within 25 years of sampling after seismic line creation (present or absent), and plot type (seismic line or adjacent forest). We then used non-metric multidimensional scaling (NMDS) with a Bray–Curtis dissimilarity index in the ‘vegan’ package version 2.7-1 [34] to accommodate the non-normal distribution of abundance data. NMDS is a distance-based ordination technique that visualizes dissimilarity through relative ordination distances, where greater relative distances among points correspond to greater relative dissimilarity in tree and tall shrub species composition and abundance. We created a distance matrix with relative species abundance, including only plots with at least one species present. We then used the package ‘ggordiplots’ version 0.4.3 [35] to plot the ordinations and assess the accuracy of the ordinations with goodness and stressplot functions in the ‘vegan’ package version 2.7-1 [34]. Accuracy of ordination was determined with non-metric R2 values for stress plots and maximized by selecting the ordination that minimized stress after 1000 permutations.
We used mixed effects generalized linear negative binomial regression models in the ‘glmmTMB’ package version 1.1.11 [36] and estimated marginal means from the ‘ggeffects’ package version 2.2.1 [37] to compare differences in regeneration density (trees per 30 m2) and recruitment density (60 m2) among forest types, recent wildfire within 25 years of sampling after seismic line creation (present or absent), and plot type (seismic line or adjacent forest). Site ID was included as a random effect to account for paired seismic line and adjacent forest plots, and models included a triple interaction among the binary and categorical variables of forest type, fire, and plot type. We used categorical (dummy) variables with the reference condition set as the undisturbed transitional forest in adjacent forest stands. We used the function r.squaredGLMM in package ‘MuMin’ version 1.48.11 [38] to calculate marginal and conditional pseudo-R2 values to assess the variance explained by fixed and random effects. We then used these global models to estimate marginal mean regeneration and recruitment densities to determine differences. Comparisons among disturbance types were made within forest types, and significant differences were defined as those where 95% confidence intervals between two groups did not overlap.
3. Results
3.1. Tree and Tall Shrub Relative Abundances
Regenerating and recruited trees sampled included black spruce, tamarack Larix laricina Du Roi, balsam fir Abies balsamea L., white spruce Picea glauca Moench, jack pine Pinus banksiana Lamb., paper birch Betula papyrifera Marshall, balsam poplar Populus balsamifera L., and trembling aspen Populus tremuloides Michx. Regenerating and recruited tall shrubs sampled included river alder Alnus incana L., mountain alder Alnus viridis Chais, serviceberry Amelanchier alnifolia Nutt., bog birch Betula pumila L., red osier dogwood Cornus sericea L., twinberry honeysuckle Lonicera involucrata Richardson, pin cherry Prunus pensylvanica L., choke cherry Prunus virginiana L., willow species Salix L., russet buffaloberry Shepherdia canadensis L., and squashberry Viburnum edule Michx. Only 1.5% of plots (n = 9) had no regeneration of trees and tall shrubs, although 25% of plots (n = 146) had no recruitment of trees and tall shrubs. Of the plots with no regeneration, most were transitional unburned seismic lines, while the plots with no recruitment were 53% peatland unburned seismic lines and 20% were transitional unburned seismic lines.
Across all forest and disturbance types, tree species accounted for 51% or fewer of regenerating trees and tall shrubs (Figure 2a). Peatland and transitional undisturbed forests had the regenerating trees compared to other disturbances within the same forest type, while undisturbed mesic upland forests had the fewest regenerating trees (Figure 2a). Black spruce was the most abundant regenerating tree in peatland (25%–46%) and transitional (21%–39%) forests, while trembling aspen was the most abundant regenerating tree in mesic upland forests (14%–29%; Figure 2a). Tall shrub regeneration was similar for peatland (50%–69%), transitional (49%–63%) and mesic upland (54%–67%) forests (Figure 2c). Unburned seismic lines in peatland and transitional forests had higher relative abundances of regenerating tall shrubs, 69% and 63%, respectively, when compared to other disturbances within the same forest type while seismic lines in mesic uplands had the lowest relative abundances (54%; Figure 2c). Within peatland forests, willow (22%–36%) and bog birch (23%–32%) were the most abundant regenerating tall shrub across all disturbance types (Figure 2c). Willows (37%–51%) were also the most abundant regenerating tall shrub in transitional forests, and squashberry (23%–45%) was the most abundant regenerating tall shrub in mesic upland forests (Figure 2c). Green alder was also an abundant regenerating tall shrub in mesic upland forests (8%–10%), while willow accounted for 16% of regenerating trees and tall shrubs in burned seismic lines in mesic uplands (Figure 2c).
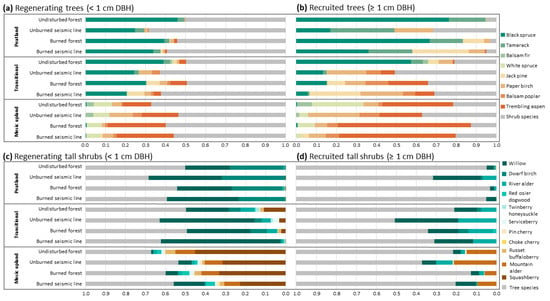
Figure 2.
Relative abundances of all trees and tall shrubs in peatland, transitional, and upland forests. Warmer colors correspond to species more abundant in mesic upland forests, while cooler colors correspond to species more abundant in peatland forests. Relative abundances of regenerating (<1.3 m tall or <1 cm diameter at 1.3 m) trees (a) and recruited (≥1 cm at 1.3 m) trees (b), and relative abundance of regenerating tall shrubs (c) and recruited tall shrubs (d).
Tree species accounted for 68%–97% of recruited trees and tall shrubs in peatland forests, 49%–69% of recruited trees and tall shrubs in transitional forests, and 63%–87% of recruited trees and tall shrubs in mesic upland forests (Figure 2b). Among disturbance types within each forest type, unburned seismic lines had the lowest abundances of recruited trees (49%–68%; Figure 2b). Black spruce (17%–76%) and tamarack (17%–32%) were the most abundant recruited tree species in peatland forests, though disturbed peatlands had higher abundances of recruited jack pine (11%–28%) and recruited paper birch (3%–19%; Figure 2b). In transitional forests, recruited black spruce was most abundant in undisturbed forests (57%), recruited paper birch was most abundant in unburned seismic lines (20%), recruited trembling aspen was most abundant in burned forests (20%), and recruited jack pine was most abundant in burned seismic lines (25%; Figure 2b). In mesic upland forests, trembling aspen was the most abundant recruited tree across all disturbances (21%–66%), except in unburned seismic lines, which had a higher abundance of recruited paper birch (26% compared to 21%; Figure 2b). Undisturbed mesic upland forests had relatively higher abundances of recruited white spruce (25%), although they did not exceed recruited trembling aspen abundance (35%; Figure 2b). Recruited willow was the most abundant tall shrub in peatland forests while in transitional forests recruited willow (11– 32%) and recruited river alder (8%–19%) were both abundant tall shrub species across all disturbance types (Figure 2d). In mesic upland forests, green alder was the most abundant recruited tall shrub in undisturbed forests (15%), unburned seismic lines (21%), burned forests (6%). Both recruited green alder (9%) and willow (10%) were relatively abundant in burned seismic lines in mesic upland forests (Figure 2d).
3.2. Similarity of Abundant Tree and Tall Shrub Composition
Minimized stress value was 0.15, and non-metric R2 value was 0.98 for NMDS of regenerating tree and tall shrub composition (Figure S1). Composition of abundant (present in at least 5% of all plots) regenerating trees and tall shrubs in transitional forests was more like peatland forests than mesic upland forests and overall regenerating trees and tall shrubs composition was more similar within the same forest type (Figure 3a). Within peatland forests, the most dissimilar in regenerating trees and tall shrubs composition was between undisturbed peatland forests and unburned seismic lines and the least dissimilarity was between burned peatland forest and burned seismic lines (Figure 3a). In transitional forests, regenerating trees and tall shrubs composition between undisturbed forests and burned seismic lines was most dissimilar, while unburned seismic lines and burned seismic lines were most similar (Figure 3a). Dissimilarities in the composition of regenerating trees and tall shrubs among disturbance types was greater in mesic upland forests compared to peatland and transitional forests (Figure 3a). Composition of regenerating trees and tall shrubs in undisturbed mesic upland forests was most dissimilar with composition of regenerating trees and tall shrubs in burned seismic lines in mesic upland forests, while regenerating trees and tall shrubs composition in unburned seismic lines and burned forests were most similar (Figure 3a).
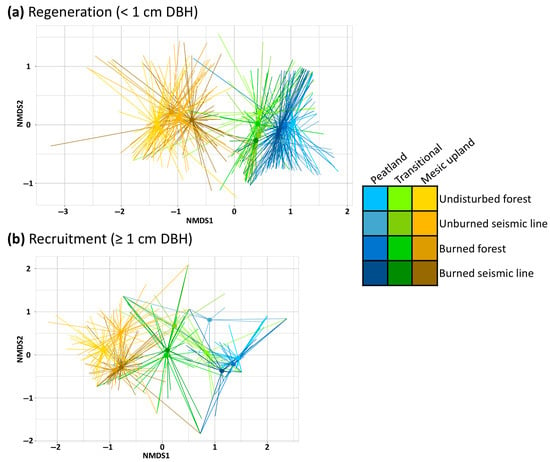
Figure 3.
Non-metric multidimensional scaling (NMDS) ordination of relative differences in composition of abundant (species present in at least 5% of plots) trees and tall shrubs across boreal forest types. Nearer centroids indicate a similar composition. (a) NMDS for regeneration (<1.3 m tall or <1 cm diameter at 1.3 m). (b) NMDS for recruitment (≥1 cm diameter at 1.3 m).
Minimized stress value was 0.12 and non-metric R2 value was 0.99 for NMDS of recruited tree and tall shrub composition (Figure S2). Composition of recruited trees and tall shrubs in transitional forests was more centralized between mesic uplands and peatlands, which were more dissimilar from each other (Figure 3b). Composition of recruited trees and tall shrubs in peatland forests was most similar between burned and undisturbed peatland forests (Figure 3b), while composition of recruited trees and tall shrubs in unburned seismic lines in transitional forests were more dissimilar from all other peatland forests disturbed types and most similar to unburned seismic lines in transitional forests (Figure 3b). Composition of recruited trees and tall shrubs in undisturbed transitional forests was most like burned seismic lines in peatlands and most dissimilar to unburned seismic lines in transitional forests (Figure 3b). In transitional forests, composition of recruited trees and tall shrubs was most similar between burned forests and burned seismic lines and most dissimilar between unburned seismic lines and undisturbed forests (Figure 3b), while composition of recruited tree and tall shrub composition in undisturbed transitional forests was most similar to burned seismic lines in peatland forests (Figure 3b). Composition of recruited trees and tall shrubs was most similar among groups in mesic upland forests compared to the other forest types (Figure 3b) and specifically between burned mesic upland forests and burned seismic lines, while undisturbed mesic upland forests were relatively dissimilar to all other groups (Figure 3b). The greatest dissimilarity in the composition of recruited trees and tall shrubs within mesic upland forests was between unburned seismic lines in mesic upland forests and burned mesic upland forests (Figure 3b).
3.3. Tree and Tall Shrub Observed Densities
Among disturbance types in peatland forests, median regeneration density was highest in burned seismic lines (68,500 stems/ha) and lowest in undisturbed forests (28,667 stems/ha; Figure 4a; Table S1). Seismic lines in peatland forests had a median regeneration density of 46,500 stems/ha, more than the median regeneration density of 33,166 stems/ha in burned peatland forests (Figure 4a; Table S1). In transitional forests, median regeneration density was low in undisturbed forests (7500 stems/ha) and high in burned seismic lines, although burned transitional forests had a lower median regeneration density (27,667 stems/ha) than transitional unburned seismic lines (7666 stems/ha; Figure 4a; Table S1). In mesic upland forests, median regeneration density was low in undisturbed forests (12,333 stems/ha) and high in burned forests (18,000 stems/ha), while seismic lines and burned seismic line median regeneration densities were 15,000 stems/ha and 15,667 stems/ha, respectively (Figure 4a; Table S1).
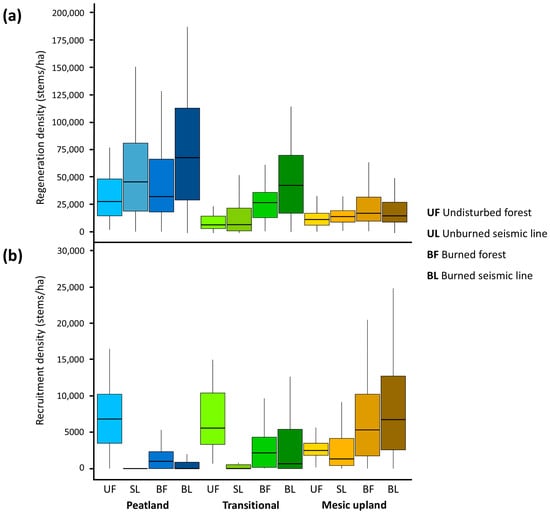
Figure 4.
Observed densities of regenerating ((a) <1.3 m tall or <1 cm diameter at 1.3 m) and recruited ((b) diameters of ≥1 cm at 1.3 m) trees and tall shrubs.
Recruitment patterns in trees and tall shrubs were more similar between peatland and transitional forests than mesic upland forests (Figure 4b; Table S1). Undisturbed forests had the highest median densities of recruited trees and tall shrubs in peatland forests (6833 stems/ha). Although burned peatland forests had a median recruitment density of 1000 stems/ha, both unburned seismic lines and burned seismic lines had median recruitment densities of zero (Figure 4b; Table S1). Similarly, within transitional forests, undisturbed forests had the highest median recruitment density (5583 stems/ha), followed by burned transitional forests (2167 stems/ha; Figure 4b; Table S1). Unburned seismic lines in transitional forests had a median recruitment density of zero, while burned seismic lines in transitional forests had a median recruitment density of 667 stems/ha (Figure 4b; Table S1). In mesic upland forests, burned seismic lines had the highest recruitment density of 6750 stems/ha, while unburned seismic lines had the lowest recruitment density of 1333 stems/ha (Figure 4b; Table S1). Undisturbed mesic upland forests had a median recruitment density of 2500 stems/ha, and burned mesic upland forests had a median recruitment density of 5333 stems/ha.
3.4. Disturbance Effects on Regeneration and Recruitment of Trees and Tall Shrubs
The conditional and marginal pseudo-R2 values for the negative binomial generalized linear mixed-effect model estimating regeneration density were both 0.38, and for the recruitment density model were both 0.37. Wildfire had a significant effect on regeneration density (p < 0.001), but not seismic lines (p = 0.081), and there was no significant interaction between wildfire and seismic lines (p = 0.850; Table S2). Conversely, both wildfire and seismic lines had significant individual effects (p < 0.001 and p = 0.032, respectively) and a significant interaction effect (p = 0.004) on recruitment density (Table S2).
In peatland forests, mean regeneration density of trees and tall shrubs in burned seismic lines was 101,695 [95% CI 79,504, 130,080] stems/ha and significantly more than undisturbed forests (48,772 [95% CI 38,708, 61,453] stems/ha), suggesting an additive effect between seismic lines and wildfire (Figure 5a; Table S3). Mean regeneration densities were not significantly different among undisturbed forests, unburned seismic lines, and burned forests or among disturbed plots (Figure 5a; Table S3). We detected a dominant effect of wildfire on regeneration density in transitional forests, where burned transitional forests had a mean regeneration density of 43,093 [95% CI 31,329, 59,276] stems/ha and burned seismic lines in transitional forests had a mean regeneration density of 60,169 [95% CI 43,743, 82,763] stems/ha (Figure 5a; Table S3). These both had significantly more regeneration than undisturbed transitional forests and unburned seismic lines (Figure 5a; Table S3). Mean regeneration in undisturbed forests and unburned seismic lines did not differ (Figure 5a; Table S3). Undisturbed transitional forests had a mean regeneration density of 14,173 [95% CI 10,355, 19,398] stems/ha and unburned seismic lines in transitional forests had a mean regeneration density of 21,036 [95% CI 15,370, 28,790] stems/ha (Figure 5a; Table S3). Mesic upland forests had a dominant effect from wildfire, as mean regeneration density in burned forests was 29,613 [95% CI 23,309, 37,622] stems/ha, which was significantly more than undisturbed forests, with a mean regeneration density of 17,650 [95% CI 14,113, 22,074] stems/ha (Figure 5a; Table S3). Seismic lines and burned seismic lines in mesic upland forests did not differ in regeneration density from undisturbed forests, suggesting no individual effect from seismic lines and no interaction effect with wildfire (Figure 5a; Table S3).
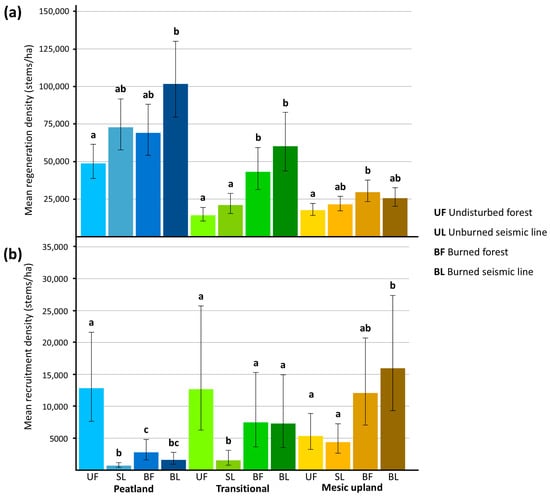
Figure 5.
Estimated marginal mean densities and 95% confidence intervals of trees and tall shrubs on seismic lines and adjacent forests across boreal forest types. Letters indicate significant differences where 95% confidence intervals did not overlap among treatments. (a) Mean density of regenerating trees and tall shrubs (<1.3 m tall or <1 cm diameter at 1.3 m). (b) Mean density of recruited trees and tall shrubs (≥1 cm at 1.3 m).
In peatland forests, seismic line clearing had a dominant negative effect on recruitment of trees and tall shrubs, as mean recruitment densities were lowest in unburned seismic lines (689 [95% CI 409, 1160] stems/ha) and burned seismic lines (1593 [95% CI 915, 2774] stems/ha; Figure 5b; Table S3). These mean recruitment densities were both significantly less than mean recruitment density in undisturbed peatland forests (12,833 [95% CI 7625, 21,598] stems/ha; Figure 5b; Table S3). Mean recruitment density was 2777 [95% CI 1603, 4810] stems/ha in burned peatland forests, which was also significantly less than recruitment density in undisturbed peatlands (Figure 5b; Table S3). Unburned seismic lines in peatland forests also had significantly fewer recruited trees and tall shrubs than burned peatland forests (Figure 5b; Table S3). Recruitment density in burned seismic lines in peatland forests also did not significantly differ from burned peatland forests (Figure 5b). In transitional forests, there was evidence of an interactive effect between subsequent disturbances such that unburned seismic lines had a mean recruitment density of 1527 [95% CI 753, 3096] stems/ha, which was significantly less than undisturbed forests, with a mean recruitment density of 12,688 [95% CI 6257, 25,729] stems/ha, burned transitional forests with a mean recruitment density of 7459 [95% CI 3637, 15,297] stems/ha, and burned seismic lines in transitional forests with a mean recruitment density of 7283 [95% CI 3551, 14,936] stems/ha (Figure 5b; Table S3). Additionally, recruitment in burned transitional forests, burned seismic lines in transitional forests, and undisturbed transitional forests did not significantly differ from one another (Figure 5b; Table S3). In mesic upland forests, we found a dominant effect of wildfire as burned seismic line recruitment (15,967 [95% CI 9312, 27,379] stems/ha) did not significantly differ from recruitment in burned mesic upland forests (12,078 [95% CI 7044, 20,709] stems/ha, but burned plots had more recruitment than unburned and undisturbed plots (Figure 5b; Table S3). Mean recruitment density did not significantly differ between undisturbed mesic upland forests (5356 [95% CI 3236, 8865] stems/ha) and unburned seismic lines (4379 [95% CI 2646, 7247] stems/ha) in mesic upland forests (Figure 5b; Table S3). Burned seismic lines in mesic upland forests had significantly more recruitment than undisturbed mesic upland forests and unburned seismic lines and while burned mesic upland forests had more recruitment mean recruitment density did not differ significantly from undisturbed and unburned mesic upland forests (Figure 5b; Table S3).
4. Discussion
We sought to characterize regeneration and recruitment of early forest succession in peatland, transitional, and mesic upland boreal forests, using tree and tall shrub abundance and composition, and identify relative disturbance effects of individual (seismic line or wildfire) and successive disturbances (seismic line followed by wildfire) on regeneration and recruitment densities of trees and tall shrubs. Across boreal forest types, all disturbed plots with individual or successive disturbances exhibited early successional dynamics 10 to 24 years after disturbance. Compositionally, tall shrub and tree regeneration was most similar within forest types and between transitional and peatland forests, while tree and tall shrub recruitment was more influenced by disturbance, with unburned seismic lines compositionally dissimilar to other plots within the same forest type. We found evidence of dominant, additive, and interactive effects from seismic lines and wildfire on early succession in boreal forests. Individual and successive disturbances influenced tree and tall shrub regeneration and recruitment in complex ways, particularly in transitional and peatland forests. We have characterized and compared early succession in boreal forests after individual and successive disturbances to further understand early succession dynamics after anthropogenic and natural disturbances. This and future work are vital to assess and predict recovery in boreal forests, especially across boreal forest transitions.
4.1. Compositional Characteristics of Early Succession Across Boreal Forest Transitions
Relative abundances of early successional tree species were higher in disturbed plots compared to undisturbed forests, indicating that disturbed plots were still in the regeneration and recruitment phase, while undisturbed forests were more likely in the young or mature forest phase. Early colonizing tree species in boreal forests are generally shade-intolerant and relatively fast growing after initial colonization, allowing rapid recruitment after disturbance [1,39]. Relative to undisturbed forests, paper birch recruitment increased in unburned seismic lines in all forest types, jack pine recruitment increased after wildfire in peatland and transitional forests, and balsam poplar recruitment increased after all disturbances in transitional and mesic uplands (Figure 2b). In peatland and mesic upland forests, dominant overstory trees were still relatively abundant in both regeneration and recruitment classes suggesting early successional species will be replaced. Notably, disturbed transitional forests had lower relative abundances (6%–15%) of recruited black spruce compared to undisturbed transitional forests (57%; Figure 2b). This may suggest that in transitional forests, early successional species persist longer, resulting in higher species diversity during early forest succession. As such, transitional forests may be more resilient to multiple disturbances as mixed-species forests can often recover more quickly after disturbances due to increased structural diversity [7]. It is important to consider early successional patterns within the first 10 years after disturbance, as differences in growth rates, nutrient and moisture tolerances, and timing of senescence of early colonizers can contribute to shifts in boreal forest communities [39,40].
We compared the composition of abundant tree and tall shrub regeneration and recruitment using ordinations with stress values for the ordinations between 0.1 and 0.2, indicating fair representation of dissimilarities among forest communities [41]. Composition of regenerating trees and tall shrubs was most similar within forest types and more similar between transitional and peatland forests (Figure 3a), supporting that nutrient and moisture gradients strongly influenced regeneration patterns. Conversely, in tree and tall shrub recruitment disturbances seemed to have a stronger influence on composition. However, transitional forests were compositionally centralized between peatland and mesic upland forests (Figure 3b). In peatland and transitional forests, undisturbed forests were most dissimilar to disturbed sites, likely due to undisturbed forests having more mature trees, as we did not use an upper diameter limit for recruitment. Unburned seismic lines in mesic uplands and transitional forests were also relatively dissimilar in recruitment composition to other disturbances and undisturbed forests, perhaps due to higher abundances of tall shrubs (Figure 2d and Figure 3b). Burned forests and burned seismic lines were the most similar in all forest types (Figure 3b), suggesting a dominant effect of wildfire on species composition recruitment.
4.2. Structural Characteristics of Early Succession Across Boreal Forest Transitions
Overall regeneration densities were greater than recruitment densities, supporting that forests, regardless of disturbance history, are following typical successional dynamics where density decreases as tree size increases through density-dependent competition [1,3]. This could also be influenced by decreasing relative abundance of tall shrubs with multiple stems (Figure 2d). However, it is important to acknowledge that median recruitment density was zero for unburned and burned seismic lines in peatland forests and unburned seismic lines in transitional forests (Figure 4b; Table S1). Furthermore, of the 146 plots sampled with no recruitment, 53% were peatland unburned seismic lines and 20% were transitional unburned seismic lines. Regeneration can be delayed in seismic lines in peatland and transitional forests by more than 8 years, such that recruitment is delayed with canopy height nearly 2 m lower on seismic lines in lowland forests compared to adjacent lowland forests [16]. We found evidence that unburned seismic lines in peatland and transitional forests may exhibit arrested succession, thus inhibiting young and mature forest development.
Regeneration and recruitment of trees can be limited by competition with grasses, forbs, and shrubs, and are dependent on light and moisture availability [39]. Relative abundances of regenerating and recruited trees and tall shrubs generally supported that early successional dynamics were proceeding from initial tall shrub colonization towards tree recruitment as there were greater abundances of tree species in the recruitment class than the regenerating class (Figure 2). An exception was in unburned seismic lines where tall shrubs remained relatively abundant (32%–51%) in the recruitment phase of all forest types (Figure 2d). Tall shrubs could persist longer in unburned seismic lines, especially in transitional forests, but given the relative abundance of recruited tree species (49%–68%) it is unlikely tall shrub persistence would result in a shift to shrub dominance (Figure 2b). Additionally, since relative abundance of recruited tall shrubs in burned seismic lines decreased to 5%–31%, it is possible that a subsequent wildfire could inhibit or reset shrub recruitment. Ultimately, a permanent shift in vegetative community from forest to shrub dominance would depend on the extent and severity of disturbance, adjacent stand age and height, and the reproductive capacity of dominant tree species, dependent on distance to disturbance [42].
We found evidence of dominant, additive, and interactive effects from seismic lines and wildfire on early succession in boreal forests. Wildfire promotes the regeneration of early successional species, including trembling aspen and paper birch, by increasing light availability [42]. Wildfire has also been shown to increase the rate of regeneration after fire by stimulating sucker production in trembling aspen within the first year after disturbance, as long as wildfires have not killed belowground roots [43]. While in mesic upland forests the dominant effect of wildfire promoted regeneration and recruitment, in transitional and peatland forests, responses were more complex. An interactive effect on recruitment in transitional forests and a dominant seismic line effect on recruitment in peatland forests suggest that seismic lines in these forests are exhibiting arrested succession. In transitional forests, wildfire’s interactive and dominant effect suggests that wildfire will promote regeneration and recruitment even in arrested seismic lines. In peatland forests, while subsequent wildfires had an additive effect on seismic line regeneration, it was not sufficient to increase seismic line recruitment. This may suggest that seismic lines in peatland forests exhibit arrested succession for longer. Increased regeneration after fire in transitional and peatland forests was likely related to increased abundance of early successional species, including deciduous hardwoods. Persistence of early successional species like trembling aspen and jack pine, with limited recruitment of late successional species like black spruce, could result in forest community shifts in transitional and peatland forests. Similar shifts have been observed in black spruce forests of interior Alaska, where conifer forests have shifted to deciduous forests after successive severe wildfires [44].
5. Conclusions
Compositional and structural forest development depends on early successional dynamics, species life history, biotic processes including competition, site conditions, and disturbances, such that there is much complexity and many possible pathways for forest development [1,39,42]. Here, we characterized and compared early succession in boreal forests after individual and successive disturbances to better understand structural and compositional shifts occurring in boreal forests. Future work should consider longer time spans (>50 years) after disturbance and quantify disturbance characteristics, landscape structure, and climate to predict whether these patterns persist, particularly across transitional ecotones in boreal forests. Understanding the complexities and intricacies of early succession after anthropogenic and natural disturbance will continue to be vital, especially when assessing or predicting recovery in boreal forests.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f16081333/s1.
Author Contributions
Conceptualization, C.M.S. and S.E.N.; methodology, C.M.S. and S.E.N.; formal analysis, C.M.S. with assistance from S.E.N.; writing—original draft preparation, C.M.S. with assistance from S.E.N.; writing—review and editing, C.M.S. and S.E.N.; visualization, C.M.S.; supervision, S.E.N.; funding acquisition, S.E.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research is part of the Boreal Ecosystem Recovery and Assessment (BERA) project (www.bera-project.org), and was supported by a Natural Sciences and Engineering Research Council of Canada (ALLRP 548285-19) in partnership with Alberta-Pacific Forest Industries Inc., Canadian Natural Resources Ltd., Cenovus Energy, ConocoPhillips Canada Resources Corp., Imperial Oil Resources Ltd., Canadian Forest Service’s Northern Forestry Centre, and the Alberta Biodiversity Monitoring Institute. Additional funding provided by Northern Scientific Training Program (NSTP) and the Alberta Conservation Association through the ACA Grants in Biodiversity Program.
Data Availability Statement
Dataset available on request from the authors.
Acknowledgments
We immensely appreciate Angelo Filicetti and the Applied Conservation Ecology lab for sharing their extensive peatland and mesic upland dataset and assistance in the field by Thomas Sperling.
Conflicts of Interest
The authors declare no conflicts of interest and note that the project funders had no input on the study’s conceptualization, design, analysis, and interpretation of the data, and no input on the decision to publish these research findings.
References
- Brassard, B.W.; Chen, H.Y.H. Stand Structural Dynamics of North American Boreal Forests. Crit. Rev. Plant Sci. 2006, 25, 115–137. [Google Scholar] [CrossRef]
- Brandt, J.P.; Flannigan, M.D.; Maynard, D.G.; Thompson, I.D.; Volney, W.J.A. An Introduction to Canada’s Boreal Zone: Ecosystem Processes, Health, Sustainability, and Environmental Issues. Environ. Rev. 2013, 21, 207–226. [Google Scholar] [CrossRef]
- Chen, H.Y.; Popadiouk, R.V. Dynamics of North American Boreal Mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y.H.; Wulder, M.A.; White, J.C. Trends in Post-Disturbance Recovery Rates of Canada’s Forests Following Wildfire and Harvest. For. Ecol. Manag. 2016, 361, 194–207. [Google Scholar] [CrossRef]
- Buma, B. Disturbance Interactions: Characterization, Prediction, and the Potential for Cascading Effects. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Burton, P.J.; Jentsch, A.; Walker, L.R. The Ecology of Disturbance Interactions. BioScience 2020, 70, 854–870. [Google Scholar] [CrossRef]
- Mitchell, J.C.; Kashian, D.M.; Chen, X.; Cousins, S.; Flaspohler, D.; Gruner, D.S.; Johnson, J.S.; Surasinghe, T.D.; Zambrano, J.; Buma, B. Forest Ecosystem Properties Emerge from Interactions of Structure and Disturbance. Front. Ecol. Environ. 2023, 21, 14–23. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing Disturbance Regimes, Ecological Memory, and Forest Resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Côté, I.M.; Darling, E.S.; Brown, C.J. Interactions among Ecosystem Stressors and Their Importance in Conservation. Proc. R. Soc. B 2016, 283, 20152592. [Google Scholar] [CrossRef]
- Webster, K.L.; Beall, F.D.; Creed, I.F.; Kreutzweiser, D.P. Impacts and Prognosis of Natural Resource Development on Water and Wetlands in Canada’s Boreal Zone. Environ. Rev. 2015, 23, 78–131. [Google Scholar] [CrossRef]
- Dabros, A.; Pyper, M.; Castilla, G. Seismic Lines in the Boreal and Arctic Ecosystems of North America: Environmental Impacts, Challenges, and Opportunities. Environ. Rev. 2018, 26, 214–229. [Google Scholar] [CrossRef]
- Nagy-Reis, M.; Dickie, M.; Calvert, A.M.; Hebblewhite, M.; Hervieux, D.; Seip, D.R.; Gilbert, S.L.; Venter, O.; DeMars, C.; Boutin, S.; et al. Habitat Loss Accelerates for the Endangered Woodland Caribou in Western Canada. Conserv. Sci. Pract. 2021, 3, e437. [Google Scholar] [CrossRef]
- Viliani, L.; Sutheimer, C.M.; Nielsen, S.E. Identifying Restoration Priorities for Habitat Defragmentation: A Case Study in Alberta’s Oil Sands. Landsc. Ecol. 2024, 39, 178. [Google Scholar] [CrossRef]
- Pattison, C.A.; Quinn, M.S.; Dale, P.; Catterall, C.P. The Landscape Impact of Linear Seismic Clearings for Oil and Gas Development in Boreal Forest. Northwest Sci. 2016, 90, 340. [Google Scholar] [CrossRef]
- Filicetti, A.; Cody, M.; Nielsen, S. Caribou Conservation: Restoring Trees on Seismic Lines in Alberta, Canada. Forests 2019, 10, 185. [Google Scholar] [CrossRef]
- Sutheimer, C.M.; Filicetti, A.T.; Viliani, L.; Nielsen, S.E. Regeneration Lags and Growth Trajectories Influence Passive Seismic Line Recovery in Western North American Boreal Forests. Restor. Ecol. 2024, 33, e14353. [Google Scholar] [CrossRef]
- Amani, M.; Mahdavi, S.; Afshar, M.; Brisco, B.; Huang, W.; Mohammad Javad Mirzadeh, S.; White, L.; Banks, S.; Montgomery, J.; Hopkinson, C. Canadian Wetland Inventory Using Google Earth Engine: The First Map and Preliminary Results. Remote Sens. 2019, 11, 842. [Google Scholar] [CrossRef]
- Bauer, I.E.; Bhatti, J.S.; Swanston, C.; Wieder, R.K.; Preston, C.M. Organic Matter Accumulation and Community Change at the Peatland–Upland Interface: Inferences from 14C and 210Pb Dated Profiles. Ecosystems 2009, 12, 636–653. [Google Scholar] [CrossRef]
- Locky, D.A.; Bayley, S.E.; Vitt, D.H. The Vegetational Ecology of Black Spruce Swamps, Fens, and Bogs in Southern Boreal Manitoba, Canada. Wetlands 2005, 25, 564–582. [Google Scholar] [CrossRef]
- Davidson, S.J.; Dazé, E.; Byun, E.; Hiler, D.; Kangur, M.; Talbot, J.; Finkelstein, S.A.; Strack, M. The Unrecognized Importance of Carbon Stocks and Fluxes from Swamps in Canada and the USA. Environ. Res. Lett. 2022, 17, 053003. [Google Scholar] [CrossRef]
- Dimitrov, D.D.; Bhatti, J.S.; Grant, R.F. The Transition Zones (Ecotone) between Boreal Forests and Peatlands: Modelling Water Table along a Transition Zone between Upland Black Spruce Forest and Poor Forested Fen in Central Saskatchewan. Ecol. Model. 2014, 274, 57–70. [Google Scholar] [CrossRef]
- Government of Alberta. Alberta Wetland Classification System; Alberta Environment and Sustainable Resource Development: Edmonton, Alberta, 2015; ISBN 978-1-4601-2258-7. [Google Scholar]
- Alberta Biodiversity Monitoring Institute. The Human Footprint Inventory (HFI) for Alberta 2022. 2025. Available online: https://abmi.ca/data-portal/80.html (accessed on 7 June 2025).
- Environment Canada. Recovery Strategy for the Woodland Caribou (Rangifer Tarandus Caribou), Boreal Population, in Canada; Species at Risk Act Recovery Strategy Series; Environment Canada: Ottawa, ON, Canada, 2012; pp. xi + 138. ISBN 978-1-100-20769-8. [Google Scholar]
- Filicetti, A.T.; Nielsen, S.E. Tree Regeneration on Industrial Linear Disturbances in Treed Peatlands Is Hastened by Wildfire and Delayed by Loss of Microtopography. Can. J. For. Res. 2020, 50, 936–945. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Nielsen, S.E. Effects of Wildfire and Soil Compaction on Recovery of Narrow Linear Disturbances in Upland Mesic Boreal Forests. For. Ecol. Manag. 2022, 510, 120073. [Google Scholar] [CrossRef]
- Beckingham, J.D.; Archibald, J.H. Field Guide to Ecosites of Northern Alberta; Special Report; Canadian Forest Service: Ottawa, ON, Canada; Northern Forestry Centre: Edmonton, AB, Canada, 1996; ISBN 978-0-660-16369-7. [Google Scholar]
- Environment and Protected Areas Cutline and Trail Attribution Project; Government of Alberta: Edmonton, AB, Canada, 2025; Unpublished work.
- Government of Alberta Historical Wildfire Perimeter Data: 1931 to 2024. 2025. Available online: https://www.alberta.ca/system/files/fp-historical-wildfire-perimeter-data.zip (accessed on 2 February 2025).
- Jones, C.; Van Dongen, A.; Harvey, J.; Degenhardt, D. Field Sampling Methods on Seismic Lines: A Comparison between Circular Plots and Belt Transects. Restor. Ecol. 2021, 30, e13619. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically Meaningful Transformations for Ordination of Species Data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; ISBN 978-1-4419-7975-9. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2025. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 June 2025).
- Quensen, J.; Simpson, G.; Oksanen, J. Ggordiplots: Make “ggplot2” Versions of Vegan’s Ordiplots. 2024. Available online: https://CRAN.R-project.org/package=ggordiplots (accessed on 5 June 2024).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-Inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378. [Google Scholar] [CrossRef]
- Lüdecke, D. Ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. 2025. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 5 June 2025).
- Bergeron, Y.; Chen, H.Y.H.; Kenkel, N.C.; Leduc, A.L.; Macdonald, S.E. Boreal Mixedwood Stand Dynamics: Ecological Processes Underlying Multiple Pathways. For. Chron. 2014, 90, 202–213. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Celis, G.; Chapin, F.S., III; Hollingsworth, T.N.; Jean, M.; Mack, M.C. Factors Shaping Alternate Successional Trajectories in Burned Black Spruce Forests of Alaska. Ecosphere 2020, 11, e03129. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Statistics for Biology and Health; Springer: New York, NY, USA; London, UK, 2007; ISBN 978-0-387-45967-7. [Google Scholar]
- Anyomi, K.A.; Neary, B.; Chen, J.; Mayor, S.J. A Critical Review of Successional Dynamics in Boreal Forests of North America. Environ. Rev. 2022, 30, 563–594. [Google Scholar] [CrossRef]
- Brown, J.K.; DeByle, N.V. Fire Damage, Mortality, and Suckering in Aspen. Can. J. For. Res. 1987, 17, 1100–1109. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Rupp, T.S.; Olson, M.; Verbyla, D. Modeling Impacts of Fire Severity on Successional Trajectories and Future Fire Behavior in Alaskan Boreal Forests. Landsc. Ecol. 2011, 26, 487–500. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).