Abstract
Floristic composition and syntaxonomy of the boreal oligotrophic pine forests covering vast areas in the boreal, hemiboreal, and temperate zones of continental Eurasia still remain poorly studied in the Southern Ural region. Using the Braun–Blanquet approach and TURBOVEG and JUICE software, the phytocoenotic diversity of boreal oligotrophic pine forests of the Southern Ural region was studied, and their position in the system of ecological and floristic classification of Eurasian vegetation was determined. Geobotanical data on boreal oligotrophic pine forests of Europe, including the European part of Russia; the Southern Urals; and Siberia were compared. A new alliance of oligotrophic boreal pine forests Brachypodio pinnati-Pinion sylvestris all. nov. hoc loco is described. The communities of this new alliance (i.e., five associations from the Southern Ural region) are characterized by a special floristic composition, occupying an intermediate position between the typical European oligotrophic pine forests of the alliance Dicrano-Pinion (Libbert 1933) Matuszkiewicz 1962 and oligotrophic (mainly psammophilous) South Siberian pine forests of the alliance Hieracio-Pinion Anenkhonov et Chytrý 1998. The communities of the alliance Brachypodio pinnati-Pinion sylvestris prefer to grow on poor soils with different moisture conditions. Due to intensive forestry activities, the distribution area of these forests has decreased, and these communities have been replaced by secondary birch forests. We have proposed a set of conservation measures to preserve these communities. A new association of oligotrophic pine forests Psephello sumensis-Pinetum sylvestris ass. nov. hoc loco is also described. These communities from the Kurgan region of Western Siberia were ordered into the alliance Dicrano-Pinion. It confirms the idea that the distribution area of this alliance may reach Siberia. Unlike the Southern Ural pine forests of the alliance Brachypodio pinnati-Pinion sylvestris, the recovery of these West Siberian pine forests after felling is quite high, and these communities do not require special measures for their protection.
1. Introduction
Progress in vegetation survey and classification of widespread Eurasian vegetation types, especially in Russia and adjacent countries, is an important and up-to-date topic of vegetation science [1,2,3]. Phytosociology is the mainstream vegetation classification scheme in Europe, as well as in some countries outside Europe, and has become increasingly popular worldwide from the 1990s onward [4,5,6]. The principal goal of phytosociology is to definition and functional characterization of vegetation types based on the total floristic composition of plant communities. In modern ecology, Braun–Blanquet phytosociology [6,7] represents the most comprehensive and consistent methodology for vegetation classification. Phytosociology distinguishes between concrete vegetation stand (phytocoenosis, plant community), which can be represented by a plot record (relevé), and abstract vegetation type (syntaxon), which represent a group of all stands sharing certain attributes. Braun–Blanquet syntaxonomy, based on the vast body of relevé data, provides a comprehensive but open system of vegetation types. Consisting of abundance data on individual plant species, relevés, and vegetation types organized in large phytosociological databases are an enormous source of biodiversity information. Phytosociological data, coupled with the growing amount of data on abundance in phytocenosis and consistency of plant occurrence and environmental information in geographic information systems (GIS), offer new opportunities to study large-scale ecological patterns and processes, and provide spatially explicit information necessary for environmental management and vegetation protection [6].
Boreal oligotrophic pine forests, dominated by Pinus sylvestris, cover vast areas in Eurasia, but their syntaxonomy and changes in floristic composition along ecological and geographical gradients are still poorly understood in some parts of their global range, such as the Southern Ural region (SUR).
In Eurasia, boreal pine forests cross the borders of several vegetation zones of northern, middle, and southern taiga; hemiboreal and broadleaf forests; and forest–steppe. This indicates that their distribution is not significantly determined by zonal and geographical conditions, but rather by edaphic factors [1,8,9].
There are several concepts of floristic classification of oligotrophic pine forests of Northern Eurasia. A comparative analysis of these forest communities, distributed in northern Europe and Western Siberia [1], revealed their common ecological and floristic features. The authors had sufficiently substantiated the concept of Oberdorfer [10] on the unified Eurasian order of these forests—Pinetalia sylvestris (Oberdorfer 1957). At the same time, they consider that the order Cladonio-Vaccinietalia Keilland-Lund 1967, with the single alliance Phyllodoco-Vaccinion Nordhagen 1936, should be placed into the class Loiseleurio-Vaccinietea Eggler ex Schubert 1960 because this syntaxa includes the non-forest subarctic shrub and dwarf shrub vegetation [1,11].
At present, the order Pinetalia sylvestris Oberdorfer 1957 includes five regional alliances of oligotrophic pine forests [1,2]:
- (1)
- Cladonio stellaris-Pinion sylvestris Keilland-Lund ex Ermakov et Morozova 2011—pine forests of boreal zone of Europe and Western Siberia;
- (2)
- Dicrano-Pinion (Libbert 1933) Matuszkiewicz 1962—pine forests of the temperate zone of Europe;
- (3)
- Hieracio umbellati-Pinion sylvestris Anenkhonov et Chytrý 1998—oligotrophic (mainly psammophilous) pine forests of Southern Siberia;
- (4)
- Saxifrago bronchialis-Pinion sylvestris Ermakov et al., 2002—oligotrophic pine forests developed on sandy deposits on permafrost in the middle boreal subzone of Eastern Siberia (Yakutia);
- (5)
- Daphno glomeratae-Pinion sylvestris Ermakov, Abdurakhmanova, Potapenko 2019—boreal oligotrophic relict pine forests of the North Caucasus, fragmentally distributed at the upper boundary of the forest belt at absolute altitudes of 1600–2350 m.
Despite the fairly detailed studies of the coniferous forests located on the East European Plain [12,13,14,15,16,17] and in Siberia [1,18,19,20,21,22,23,24,25,26], the syntaxonomical position of the boreal oligotrophic pine forest in the SUR remains a subject of controversy. Some authors placed these forest communities in the alliance Dicrano-Pinion sylvestris (Libbert 1933) Matuszkiewicz 1962 [27,28,29], noting that this decision is conditional and requires further clarification. Previously, the position of boreal oligotrophic pine forests of the alliance Dicrano-Pinion in the system of classification of higher units was ambiguous. Some authors included Dicrano-Pinion in the order Piceetalia excelsae Pawłowski, Sokołowski et Wallisch 1928, which combines xeromesophytic and mesophytic dark coniferous boreal forests of the class Vaccinio-Piceetea [4,5,30,31,32,33,34], while the others [12,13,14,34,35] included northern pine forests to the order Cladonio-Vaccinietalia K.-Lund 1967. Here, we follow the concept of Oberdorfer (1992) on the unified Eurasian order of the oligotrophic pine forests of Northern Eurasia, Pinetalia sylvestris, which was confirmed by Ermakov and Morozova [1]. In our study, we assumed that the floristic features of the Southern Ural oligotrophic green moss pine forests may differ significantly from the green moss pine forests of Europe and Siberia, and these differences do not allow to order them into any of the existing vegetation alliances. The use of a standardized classification system based on the Braun–Blanquet approach allow us to conduct a comparative analysis of the similarities and differences between forest types, including oligotrophic pine forests, located in different geographical regions and climatic zones.
This work is aimed to conduct a detailed analysis of the vegetation of the boreal oligotrophic pine forests of the SUR in order to reveal their distribution and regional features, as well as to clarify their position at a high syntaxonomic level.
The tasks of the study included collection of primary data (geobotanical relevés of oligotrophic green moss pine forests of the SUR), the syntaxonomical analysis, cluster analysis of all known associations and subassociations of green moss pine forests in order to identify the peculiarities of the Southern Ural green moss pine forests and to decide whether a new alliance of green moss pine forests should be described.
2. Materials and Methods
2.1. Study Area
The study area (the Southern Ural region, the SUR) includes the Southern Ural mountains and their western and eastern foothills. The main part of the SUR is located on the territory of the Republic of Bashkortostan (N 51°34′–6°31′ and E 53°08′–60°00′). The northern border of the SUR is limited by the Chelyabinsk region on the latitudinal course of the Ufa River, and the southern border runs along the Jewtube and Guberlinskiy ridges in the Orenburg region.
The climate of the study area is mainly continental, with moderately warm, sometimes hot summers and long, moderately cold winters. The average annual temperature ranges from +0.3 to +3.5 °C; the average temperature varies from −14 to −17 °C in January and from +16.5 to +19.5 °C in July. The frost-free period lasts about 107 days (usually from 30 May to 15 September). The sum of temperatures above 10 °C is 1600–2350 °C. The period of active vegetation is about 95 days.
The Ural ridge is a natural barrier to the path of moist and warm Atlantic air masses, having a great influence on the climatic conditions of the western and eastern foothills. The amount of precipitation increases from the western foothills to the central elevated part of the Southern Urals and then decreases in the Bashkir Trans-Urals. The annual precipitation is 450–550 mm in the Bashkir Cis-Urals, 650–700 mm or more on the western slope and in the central elevated part of the Southern Urals, and does not exceed 300–400 mm precipitation in the Bashkir Trans-Urals. About 60%–70% of the annual precipitation falls in the warm season [36,37,38].
In the SUR, the altitude ranges from 60 to 1640 m above sea level, with a maximum in the Southern Urals mountains, where three distinct vegetation belts of mountainous forest, subalpine meadows and alpine tundra are represented [39]. The high vegetation diversity of the SUR depends on a number of regional factors: unique geographic position between Europe and Asia, mountains that—being an important climatic boundary—cause significant differences in vegetation of the western and eastern macroslopes; the complex history of vegetation formation in the Pleistocene and Holocene; the absence of Pleistocene glaciation, which allowed to preserve the ancient elements of the vegetation; etc. [40,41,42,43,44,45,46,47].
In the Bashkir Cis-Urals and on the western slopes of the Southern Urals, the indigenous forest vegetation is represented by nemoral broadleaf forests dominated by Tilia cordata, Acer platanoides, Ulmus glabra, and—less commonly—Quercus robur. Boreal and hemiboreal dark coniferous forests with Picea obovata and Abies sibirica are located mainly in the central elevated part of the Southern Urals. The eastern slopes of the Southern Urals are covered with light-coniferous hemiboreal herb forests dominated by Pinus sylvestris, Larix sukaczewii, and Larix sibirica as well as secondary birch forests. In the forests of the SUR, there is an ecotone effect [48,49] of regional scale, which appears as a combination of species typical for nemoral, boreal, and hemiboreal floristic complexes. Due to this, the boreal forests of the SUR have higher species richness than similar forests in adjacent areas [44,46,50].
In the SUR, boreal oligotrophic pine forests grow near the southern border of their range. In the Bashkir Cis-Urals, these forests are widespread on the Ufa Plateau. In the mountains of the Southern Urals, they are distributed further south, reaching 53 degrees northern latitude (Figure 1). It should be noted that these forests were severely damaged due to logging. Until 1980, forest management in the Republic of Bashkortostan included large-scale deforestation, which was estimated by the number of cubic meters of harvested timber. The planting of forest seedlings on the cutting sites was largely delayed, and as a result, many felled areas were overgrown with birch and aspen during the period of natural reforestation [51]. For this reason, boreal oligotrophic pine forests of the SUR were mainly preserved in protected areas.
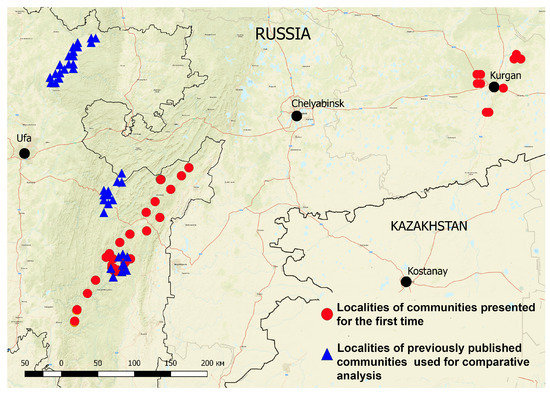
Figure 1.
Localities of relevés of boreal oligotrophic pine forests in the SUR and the Kurgan region.
The oligotrophic pine forests of the Kurgan region (Russia) have also been studied by the authors for syntaxonomic comparison. The scientific literature provides abundant data on oligotrophic pine forests in many regions. However, such data are not sufficient for the Kurgan region. This region is located in the forest–steppe zone of the West Siberian Plain within N53°50′–57°10′ and E61°30′–69°30′ at the altitude of 180–230 m above sea level [52]. The climate of the Kurgan region is continental, with dry winters and hot, dry summers. Average temperature in January ranges from −17 to −12 °C, and the average temperature in July ranges from +17 to +19 °C. Average annual temperature is +1.5 °C, with 100–125 frost-free days. Annual precipitation is 300–350 mm, with 60% occurring in summer [53]. The soil cover of Kurgan region is represented mainly by ashed, leached, and typical chernozems. However, pine forests grow mainly on podzolic sands and sandy loams, which extend along the ancient terraces of the large rivers (Tobol, Iset, Miass, Pyshma, and Ishim) and their tributaries [54].
2.2. Data Collection
Data to be used were 130 relevés of boreal oligotrophic pine forest communities described by the authors in the SUR and Kurgan regions in 1996–2013 together with unpublished relevés stored on the Vegetation Database Forest of Southern Ural (GIVD ID 00-RU-001) [55] and some published data on the pine forests of the SUR [29]. Sample plots were established in stands that are relatively homogeneous in structure, species composition, and habitat conditions. Sample plots were square or rectangular in shape with side lengths ranging from 10 to 35 m. The areas of sample plots ranged from 200 to 1000 m2 depending on the homogenous of the vegetation cover. The data of each sample plot (relevé) contained data on geographical position, environmental conditions (position in relief, exposure, and slope steepness), and general vegetation structure as well as a complete list of plant species growing on the soil, including mosses, lichens with an estimate of their occurrence and abundance. Species participation in the vegetation cover of the plots was estimated on the Braun–Blanquet scale [7]: r—single species in the sample plot; +—species covers up to 1%; 1—species covers 1% to 5%; a –species covers 5% to 15%; b—species covers 15% to 25%; 3—species covers 25% to 50%; 4—species covers 50% to 75%; 5—species covers more than 75% of the plot. The cover values of moss and lichen species growing on the bark of leaving trees and rotten wood was not specifically evaluated. In Tables S4 and S6, only frequency of occurrence of these species is indicated: r—rare species, +—the species is common within sample plot. These species are included in the tables because they are part of the forest community within the sample plot. To distinguish these species from mosses and lichens growing on the soil, epiphytic and epixylic species are marked with an asterisk. The tree species occurring in different layers are counted separately for each layer (t1, t2, t3) with indication of their projective cover and height. The values of total projective cover of shrub, herb, and moss layers were also indicated in percent.
2.3. Data Analysis
Vegetation plot data were digitized and stored in TURBOVEG (Wageningen, The Netherlands) [56]. The relevés were classified to the levels of class, order, alliance, association and lowest units, according to the Braun–Blanquet approach [7,57,58], using the JUICE software [59]. The major tool for delineation and identification of vegetation units at each syntaxonomical level was diagnostic species combination, determined by calculating fidelity values of species.
To define the position of the boreal oligotrophic pine forests of the SUR and the Kurgan region in the classification system of higher units, we used data on 52 syntaxa of oligotrophic pine forests described in Europe, including the European part of Russia [1,12,13,15,16,34,60,61,62,63,64,65,66,67,68,69], Western Siberia [23,24,70], Eastern Siberia [19,22], and data on nine syntaxa from the SUR [27,28,29]. Data on the boreal pine forests of the alliance Daphno glomeratae-Pinion sylvestris described in the Caucasus and the alliance Saxifrago bronchialis-Pinion from Eastern Siberia were not included in the comparative analysis due to significant floristic and ecological differences with the communities of the SUR.
In the JUICE software, synthetic table was created with the data mentioned above. Each column in Table S1 represented the association or a low-rank unit. The standard scale of constancy was 1 (1%–20%), 2 (21%–40%), 3 (41%–60%), 4 (61%–80%), and 5 (81%–100%) since syntaxonomic units are presented only in the form of synoptic tables in many publications. The synoptic table with the data from the SUR was created using frequency and fidelity index (measure: phi coefficient, presence/absence, standardised to equal group size, Fisher’s exact test for significance with p < 0.01). All species with Φ > 0.25 (with p < 0.01) were selected as diagnostic species. The numbers in Tables S1 and S3 are percentage occurrence frequencies (constancies).
Additionally, hierarchical cluster analysis (Ward’s linkage method and Bray-Curtis distances as a measure of dissimilarity) using the program STATISTICA [71] was additionally carried out to identify the position of the forest community from the SUR in the system of higher classification units.
Syntaxa names follow the International Code of Phytosociological Nomenclature [72,73]. The nomenclature of vascular plant species is mainly given according to “The Plant List” [74], with some exceptions concerning the species distributed only in Russia [75]. Latin names for bryophyte and lichen species are given according to “An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus” [76], and “A checklist of the lichen flora of Russia” [77].
3. Results
3.1. The Diversity of Studied Boreal Oligotrophic Pine Forests
Boreal oligotrophic pine forest communities studied by authors in the SUR and the Kurgan region could be referred to five associations, which include more low-rank units, i.e., three subassociations and six variants (Table A1, Figure 2). The position of the studied communities in the system of higher rank synaxonomic units is presented below:
| Class VACCINIO-PICEETEA Br.-Bl. in Br.-Bl. et al., 1939 Order PINETALIA SYLVESTRIS Oberdorfer 1957 Alliance Dicrano-Pinion sylvestris (Libbert 1933) Matuszkiewicz 1962 Association Psephello sumensis-Pinetum sylvestris ass. nov. hoc loco Alliance Brachypodio pinnati-Pinion sylvestris all. nov. hoc loco Association Violo rupestris-Pinetum sylvestris Martynenko et al., 2003 Variant typica Variant Veronica spicata Association Seseli krylovii-Pinetum sylvestris Martynenko et al., 2008 Association Pleurospermo uralensis-Pinetum sylvestris Martynenko et al., 2003 Subassociation P.u.-P.s. laricetosum sukazhewii subass. nov. hoc loco Subassociation P.u.-P.s. digitalietosum grandiflorae subass. nov. hoc loco Variant typica Variant Cirsium heterophyllum Subassociation P.u.-P.s. anemonastretosum biarmiensis subass. nov. hoc loco Association Zigadeno sibirici-Pinetum sylvestris Martynenko et Zhigunova 2004 Variant Cystopteris fragilis Variant Viburnum opulus |
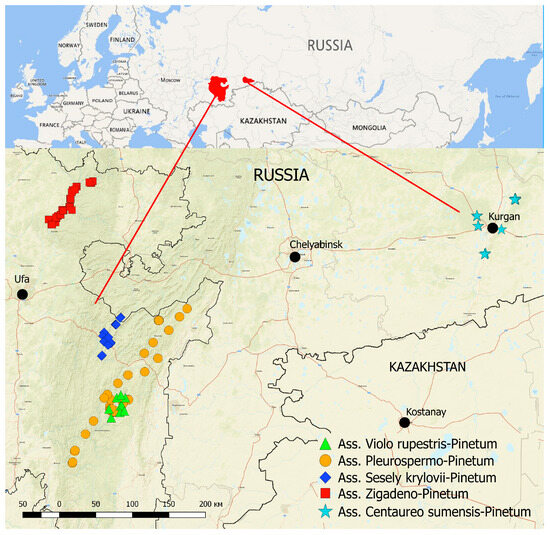
Figure 2.
Distribution of syntaxa of boreal oligotrophic pine forests in the SUR and the Kurgan region.
To results of a comparative analysis of boreal oligotrophic pine forests of the SUR and other regions are presented in Tables S1 and S2.
The hierarchical cluster analysis allowed us to check the integrity of alliances and to identify the most important floristic similarities and differences between higher units of studied boreal oligotrophic pine forests (Figure 3).
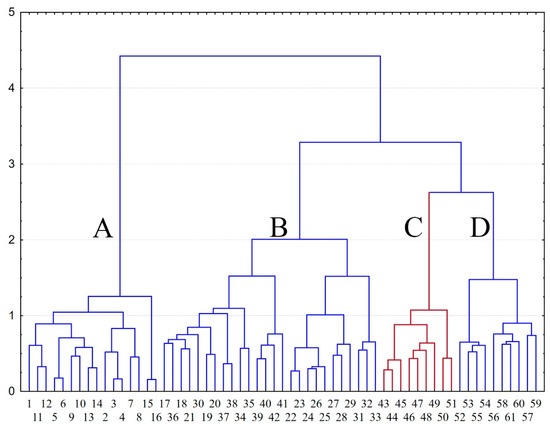
Figure 3.
Tree diagram (Ward’s method, Bray-Curtis distances) of 61 syntaxa of boreal oligotrophic pine forests from alliances Cladonio stellaris-Pinion sylvestris (A: 1–16), Dicrano-Pinion (B: 17–42), Brachypodio-Pinion (C: 43–51, red color), Hieracio-Pinion (D: 52–61). Numbers in the tree diagram correspond to the numbers of the syntaxa in Table S1.
A quantitative treatment of the entire data set (Table S1) revealed a delimitation of all associations into four main groups, corresponding to zonal types of pine forests of the middle and northern taiga zone (Figure 3, cluster A), the temperate zone (Figure 3, cluster B), and to two types of the southern boreal subzone, complicated by an altitudinal belt (Figure 3, clusters C and D). These groups of boreal oligotrophic pine forests represent the communities of four alliances, i.e., Cladonio-Pinion (Figure 3, cluster A), Dicrano-Pinion (Figure 3, cluster B), Brachypodio-Pinion (Figure 3, cluster C) and Hieracio-Pinion (Figure 3, cluster D). The communities of the SUR are more floristically similar to the forests of the alliance Hieracio-Pinion compared to European communities of the alliance Dicrano-Pinion.
3.2. Description of Vegetation Units
All. Brachypodio pinnati-Pinion sylvestris all. nov. hoc loco (Table A1, col. 1–9; Table S1, col. 43–51)
Holotypus: ass. Pleurospermo uralensis-Pinetum sylvestris Martynenko et al., 2003
Diagnostic species: Pleurospermum uralense, Trifolium lupinaster, Geranium pseudosibiricum, Brachypodium pinnatum, Lathyrus pisiformis, Chamaecytisus ruthenicus, Viola collina, Adenophora liliifolia, Seseli krylovii, Poa sibirica, Campanula glomerata, Digitalis grandiflora, Cerastium pauciflorum, Persicaria bistorta, Lilium martagon, Atragene sibirica.
Alliance unites boreal xerophytic and xeromesophytic oligotrophic pine and pine–larch forests of the SUR, which are formed on poorly developed light loamy–sandy podzolized mountain forest soils at the altitude of 150–750 m above sea level. Geographically, these forests are located in the Republic of Bashkortostan and adjacent Urals regions. In the central elevated part of the Southern Urals and on the Ufa Plateau, these forests grow mainly on the southern slopes of mountains or the steep banks of deep mountain river valleys. Near the southern outskirts of the Urals, these forests often grow on mountain slopes with a northern exposure.
In the SUR, these forests are highly fragmented and located in small areas surrounded by dark coniferous hemiboreal forests of the class Asaro-Abietetea Ermakov, Mucina et Zhitlukhina 2016 (in the central elevated part of the Southern Urals), pine–broadleaf forests of the class Carpino-Fagetea Jakucs ex Passarge 1968 (on the western slope of the Southern Urals), or hemiboreal light coniferous and small-leaved herb forests of the class Brachypodio-Betuletea Ermakov, Korolyuk et Lashchinsky 1991 and steppe vegetation of the class Festuco-Brometea Br.-Bl. et Tx. ex Soó (1947) (on the eastern slope and near the southern edge of the Urals).
The tree layer is dominated by Pinus sylvestris and sometimes Larix sukaczewii, with an admixture of Betula pendula, which is more abundantly represented in the second and third tree layers. In the undergrowth, Sorbus aucuparia is usually present. The shrub layer is mostly poorly developed and represented by small specimens of Chamaecytisus ruthenicus, Atragene sibirica, and Rosa majalis.
The herb layer is represented by combination of boreal dwarf shrub species (Vaccinium myrtillus, Vaccinium vitis-idaea), taiga small herb species (Orthilia secunda, Maianthemum bifolium, Trientalis europaea, Cerastium pauciflorum), and hemiboreal species (Brachypodium pinnatum, Viola collina, Adenophora liliifolia, Lathyrus pisiformis, Trifolium lupinaster, etc.). Additionally, species typical for broadleaf and coniferous–broadleaf forests of classes Carpino-Fagetea and Asaro-Abietetea have high constancy. Due to their ecotonic character, the alliance communities have a high species saturation, with up to 64 species of vascular plants on the plot [25]. The moss layer is well developed and represented by typical boreal mosses (Pleurozium schreberi and Hylocomium splendens, with cover ranges from 55 to 95%.
Currently, the alliance includes four associations, with three sub-associations and six variants (Table A1).
Ass. Violo rupestris-Pinetum sylvestris Martynenko et al., 2003 (Table A1, col. 1–2)
Diagnostic species: Pinus sylvestris (dom.), Larix sukaczewii, Persicaria alpina, Dianthus chinensis, Dracocephalum ruyschiana, Festuca rupicola, Filipendula vulgaris, Galium ruthenicum, Lathyrus pratensis, Poa lapponica, Silene amoena, Viola rupestris, Silene viscaria.
The association unites boreal xerophytic larch–pine forests of the SUR, formed on poorly developed stony soils with rock outcrops. The communities were described on the slopes of the Kraka mountain range in the Bashkir State Nature Reserve.
The tree layer is dominated by Pinus sylvestris and Larix sukaczewii. The shrub layer is poorly developed and represented by single specimens of Chamaecytisus ruthenicus and Juniperus communis. The herb layer has a cover value of 15%–35% and is dominated by Calamagrostis arundinacea, Vaccinium myrtillus, Rubus saxatilis, and Brachypodium pinnatum.
The communities are differentiated by the presence with high constancy of xerophytic and some petrophytic species, which are typical for mountain steppes and hemiboreal forests (Festuca rupicola, Viola rupestris, Galium ruthenicum, Dianthus versicolor, Silene viscaria, Filipendula vulgaris).
The moss cover value is high and reaches 95%. Boulders and rock outcrops are usually covered with green mosses, among which Pleurozium schreberi predominates.
The association has two variants:
Var. typica (Table A1, col. 1). This variant represents typical association communities, distributed mainly in the middle parts of the southern mountain slopes or in the upper parts of the northern mountain slopes, where these forests grow on the border with the mountain steppes.
Var. Veronica spicata (Table A1, col. 2). Diagnostic species: Rosa acicularis, Aster alpinus, Carex caryophyllea, Centaurea sibirica, Polygala comosa, Tephroseris integrifolia, Pseudolysimachion spicatum. This variant includes the communities with a less developed moss cover, which occur in drier conditions, mainly on slopes with southern and western exposures.
Ass. Pleurospermo uralensis-Pinetum sylvestris Martynenko et al., 2003 (Table A1, col. 3–6)
Diagnostic species: Pinus sylvestris (dom.), Atragene sibirica, Aconitum lycoctonum, Angelica sylvestris, Persicaria bistorta, Geranium sylvaticum, Hieracium albocostatum, Milium effusum, Pleurospermum uralense, Pulmonaria mollissima, Pyrola minor, Succisa pratensis.
Communities were found on the western and eastern slopes of the Southern Urals (in Burzyansky, Beloretsky, Uchalinsky, and Kugarchinsky districts of the Republic of Bashkortostan). These forests grow on weakly developed and sandy loam mountain forest soils on the northern mountain slopes at the altitude of 200–750 m.
The tree layer has an average cover value of 55% and consists of Pinus sylvestris with an admixture of Betula pendula. The dwarf shrub and herb layer has a cover value of 20–75% and is dominated by Vaccinium myrtillus, Calamagrostis arundinacea, and—rarely—Rubus saxatilis. Communities of the association include a number of mesophytic species with high constancy, such as Angelica sylvestris, Heracleum sibiricum, Geranium sylvaticum, Maianthemum bifolium, Persicaria bistorta, Viola mirabilis, Aegopodium podagraria, or Cirsium heterophyllum. The moss cover is 45%–95% and is dominated by Pleurozium schreberi.
Association Pleurospermo uralensis-Pinetum sylvestris was enlarged by the inclusion at the level of subassociation communities of the former association Digitali grandiflorae-Pinetum sylvestris. Thus, there are currently three subassociations.
Subass. P.u.-P.s. laricetosum sukaczewii subass. nov. hoc loco (Table A1, col. 3; Tables S4 and S5, col. 1–15).
Holotypus: col. 9, Table S4.
Diagnostic species: Larix sukaczewii, Juniperus communis, Persicaria alpina, Lathyrus pratensis.
This subassociation unites communities dominated by Pinus sylvestris and Larix sukaczewii. They are usually distributed in the upper part of the slopes of ridges, where the cenotic position of larch is enhanced. The nomenclature type (holotypus) of this subassociation corresponds to the nomenclature type of the association published in the monograph on the forests of the Bashkir State Nature Reserve.
Subass. P.u.-P.s. digitalietosum grandiflorae Martynenko et al., 2003 stat. nov. (Table A1, col. 4–5, Figure 4b)

Figure 4.
Boreal oligotrophic pine forests in the SUR (authors of photos Shirokikh P.S., Muldashev A.A. ((a) Ass. Zigadeno sibirici-Pinetum sylvestris; (b) Ass. Pleurospermo uralensis-Pinetum sylvestris, subass. digitalietosum grandiflorae; (c) Ass. Pleurospermo uralensis-Pinetum sylvestris, subass. anemonastretosum biarmiensis).
Holotypus: col. 23, Tables S4 and S5.
Diagnostic species: Padus avium, Potentilla erecta, Adonis apennina, Aegopodium podagraria, Bupleurum longifolium, Cerastium pauciflorum, Digitalis grandiflora, Lathyrus gmelinii, Primula macrocalyx, Stachys officinalis, Trollius europeus.
This subassociation unites communities distributed at lower altitudes than the communities of the subassociation P.u.-P.s. laricetosum sukaczewii. In these communities, the coenotic position of birch increases, and this species is often found in the first tree layer. Padus avium occurs in the undergrowth.
The communities of the subassociation border hemiboreal pine–birch forests of the association Bupleuro longifolii-Pinetum sylvestris Fedorov ex Ermakov et al., 2000 (class Brachypodio-Betuletea). Therefore, typical species of hemiboreal forests (Bupleurum longifolium, Pleurospermum uralense, Digitalis grandiflora, Pulmonaria mollissima, Cirsium heterophyllum, Succisa pratensis) are present with high constancy in the floristic composition of these communities.
Currently, the subassociation can be divided into two variants:
Diagnostic species = diagnostic species of subassociation.
This variant represents typical communities of the subassociation, which are widespread in the lower parts of mountain slopes of the Southern Urals in the Bashkortostan Republic.
Diagnostic species: Daphne mezereum, Carex rhizina, Cirsium heterophyllum, Epipactis helleborine, Hypericum maculatum, Pedicularis uralensis, Ranunculus auricomus.
These communities are characterized by high constancy of the nemoral species Epipactis helleborine, Poa nemoralis, and Daphne mezereum.
Subass. P.u.-P.s. anemonastretosum biarmiensis subass. nov. hoc loco (Table A1, col. 6; Tables S4 and S5 col. 43–53, Figure 4c).
Holotypus: col. 45, Tables S4 and S5 in this paper.
Diagnostic species: Anemonastrum biarmiense, Carex montana, Lycopodium annotinum, Molinia caerulea, Hypochaeris maculata.
These communities are distributed mainly in the central elevated part of the Southern Urals on the gentle slopes (1–3°) of the northern exposure. The communities are differentiated by high constancy of the species Vaccinium vitis-idaea, Molinia caerulea, Hypochaeris maculata, and Anemonastrum biarmiense, which are mountainous endemics of the Southern Urals [44]. The moss layer has a cover value of 20%–90% and is dominated by Pleurozium schreberi and Hylocomium splendens.
Ass. Seseli krylovii-Pinetum sylvestris Martynenko et al., 2008 (Table A1, col. 7).
Diagnostic species: Pinus sylvestris (dom.), Abies sibirica, Betula pubescens, Picea obovata, Tilia cordata (t3), Lonicera xylosteum, Carex macroura, C. pilosa, Seseli krylovii, Succisa pratensis, Vaccinium vitis-idaea.
This association unites communities of boreal oligotrophic pine forests of the central elevated part of the Southern Urals, which grow on poorly developed and drained soils with a highly variable moisture regime. Communities were found on the southern slopes along the valleys of mountain rivers and streams. The association was validly published in the monograph on the flora and vegetation of the South Ural State Nature Reserve [29].
The tree layer is dominated by Pinus sylvestris, with occasional Betula pubescens and B. pendula. The cover value of the tree layer is 40%–70%, with an average of 55%. Abies sibirica and Picea obovata are also found in the undergrowth. In the third tree layer, in addition to pine and birch, broadleaf tree species (Tilia cordata, Quercus robur, Acer platanoides, Padus avium) are common. Sorbus aucuparia occurs with high constancy in the undergrowth. The high diversity of tree species is due to the fact that these communities are surrounded by coniferous–broadleaf hemiboreal forests of the order Abietetalia sibiricae (Ermakov in Ermakov et al., 2000) Ermakov 2006 (class Asaro-Abietetea) and dark coniferous boreal forests of the alliance Piceion excelsae Pawłowski, Sokołowski et Wallisch 1928 (class Vaccinio-Piceetea).
The shrub layer is mostly poorly developed. Its cover value usually does not exceed 1% , but can rarely reach 5%. It is composed of single specimens of Rosa majalis, Lonicera xylosteum, Chamaecytisus ruthenicus, Cotoneaster melanocarpus, and Rubus idaeus.
The herb–dwarf shrub layer is dominated by Vaccinium myrtillus, Calamagrostis arundinacea, Rubus saxatilis, and sometimes Vaccinium vitis-idaea and Brachypodium pinnatum. In addition to the species of the alliance Dicrano-Pinion and boreal small herb taiga species of the class Vaccinio-Piceetea, the species of hemiboreal herb forests of the class Brachypodio-Betuletea (Viola collina, Adenophora lilifolia, Carex macroura, Digitalis grandiflora, Hieracium umbellatum, Seseli krylovii, Silene nutans, and Pulmonaria mollissima) are present with high constancy. The cover of the dwarf shrub–herb layer varies from 10% to 65% (35% on average), depending on the development of the soil and moss cover. The moss layer is represented by Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum and Ptilium crista-castrensis and has a cover value of 40%–90%.
Although these communities are enriched in species typical for broadleaf dark coniferous forests, they have the least biodiversity among of all communities of thise alliance (average species richness is about 41 species per sample plot). Therefore, the diagnostic block of the alliance is poorly represented.
Ass. Zigadeno sibirici-Pinetum sylvestris Martynenko et Zhigunova 2004 (Table A1, col. 8–9, Figure 4a).
Diagnostic species: Pinus sylvestris (dom.), Betula pubescens (t1), Caragana frutex, Frangula dodonei, Campanula rotundifolia, Arabidopsis arenosa, Carex alba, Cortusa matthioli, Equisetum scirpoides, Galium album, G. tinctorium, Gymnocarpium robertianum, Primula cortusoides, Valeriana wolgensis, Anticlea sibirica.
This association unites relict communities of pine and birch–pine boreal mesophytic forests with high species diversity. These communities were found on the Ufa Plateau on slopes with southern, southwestern, and southeastern exposures. The forests were described on poorly developed soils with carbonate outcrops and occupy a large area on steep, well-lit slopes along the Ufa River (Karaidel’skiy and Askinskiy districts of the Republic of Bashkortostan).
Communities of the association Zigadeno-Pinetum have an ecotone character and border with hemiboreal coniferous–broadleaf forests of the class Asaro-Abietetea. Additionally, the species typical for hemiboreal birch–pine forests of the class Brachypodio-Betuletea have quite high constancy in these communities. Compared to communities of other associations, the species richness increases and ranges from 50 to 93 species of vascular plants per sample plot (an average of 64 species per plot).
Compared to communities of other associations of this alliance, the tree layer of association Zigadeno sibirici-Pinetum sylvestris has higher cover values, ranging from 60 to 75%. The tree stand is divided into three layers: the first layer is formed by Pinus sylvestris mixed with Betula pendula and Betula pubescens. In the second and third layers, Picea obovata and Abies sibirica are frequent. An undergrowth is formed by broadleaf tree species such as Tilia cordata, Quercus robur, Acer platanoides, and Ulmus glabra.
The shrub layer is poorly developed and rarely reaches 5% but is characterized by high species diversity. In terms of floristic composition, sub-boreal nemoral species and species typical for meadow–steppe communities (Chamaecytisus ruthenicus, Frangula dodonei, Euonymus verrucosa, Atragene sibirica, Lonicera xylosteum, Rosa majalis, Cotoneaster melanocarpus, Cerasus fruticosa, and Caragana frutex) grow together with high constancy.
A characteristic feature of the oligotrophic pine forests of the Ufa plateau is the absence of boreal dwarf shrubs (Vaccinium myrtillus and Vaccinium vitis-idaea) and the absence of pronounced dominants. Rubus saxatilis, Gymnocarpium robertianum, Calamagrostis arundinacea, Carex alba, C. rhizina, Equisetum scirpoides, and Orthilia secunda can reach a relatively high cover. In addition to boreal species, nemoral species (e.g., Aegopodium podagraria, Lathyrus vernus, Pulmonaria obscura, Stellaria holostea, Poa nemoralis), and species typical for hemiboreal herb forests (e.g., Viola collina, Trifolium lupinaster, Adenophora lilifolia, Pleurospermum uralense, Hieracium umbellatum) are presented in the herb–dwarf shrub layer, the cover of which ranges from 15 to 50% (with an average of 30%).
The moss cover varies between 10 and 90% depending on the development of the soils and the herbdwarf shrub layer. Pleurozium schreberi and Hylocomium splendens are the dominant species. Dicranum polysetum and Rhytidiadelphus triquetrus have lower abundancy.
This association was early divided into two variants: typica and Lathyrus vernus [29]. As a result of comprehensive floristic analysis, the composition of their diagnostic blocks was corrected, and the names of the variants were changed.
Var. Cystopteris fragilis (Table A1, col. 8).
Diagnostic species: Cystopteris fragilis, Cerastium uralense, Euphorbia microcarpa.
The variant unites the typical communities of the association. In the first tree layer, in addition to Pinus sylvestris and Picea obovata, Betula pubescens is present. The cover of the herb–dwarf shrub layer is low and ranges from 15% to 30%. The dominant species are Rubus saxatilis, Gymnocarpium robertianum, with occasional Calamagrostis arundinacea and Equisetum scirpoides. The moss cover ranges from 50% to 90% (with average of 80%).
Var. Viburnum opulus (Table A1, col. 9).
Diagnostic species: Viburnum opulus (t3), Tilia cordata (t2), Populus tremula (t3), Ulmus glabra (t3), Astragalus danicus.
These communities were previously described as var. Lathyrus vernus [29,31]. The variant unites communities with a large number of species typical for broadleaf forests. In the floristic composition of these communities, the group of diagnostic species of the class Carpino-Fagetea is better represented than the group of diagnostic species Vaccinio-Piceetea. The cover of the moss layer is low and does not exceed 35% (with an average of 15%).
All. Dicrano-Pinion (Libbert 1933) Matuszkiewicz 1962.
This alliance unites azonal psammophytic and petrophytic oligotrophic pine forests with a poorly developed shrub layer and species-poor herb layer, which contains acidophilous dwarf shrubs (Calluna vulgaris, Vaccinium myrtillus and V. vitis-idaea) and oligotrophic graminoids and other herbs. The moss layer is well developed. This alliance is distributed throughout the temperate zones from Europe to Southern Urals on nutrient-poor, acidic soils, where deciduous trees are disadvantaged and pine can attain dominance, especially if the stands are occasionally disturbed, e.g., by fires [1,2,3].
Ass. Psephello sumensis-Pinetum sylvestris ass. nov. hoc loco (Tables S1, col. 42, S6 and S7).
Holotypus: col. 9, Tables S6 and S7.
Diagnostic species: Pinus sylvestris (dom.), Betula krylovii, Poa angustifolia, Pulsatilla uralensis, Astragalus danicus, Psephellus sumensis, Eremogone saxatilis, Vincetoxicum hirundinaria, Fragaria vesca, Pseudolysimachion spicatum.
The association unites boreal oligotrophic pine forests with a significantly high moss cover, formed on alluvial sandy soils. They are widespread on the terraces of the Iset and Tobol Rivers on the southernmost border of the southern taiga subzone in Western Siberia [54].
The tree layer is dominated by Pinus sylvestris mixed with Betula krylovii and Betula pubescens. The cover of the tree layer ranges from 35 to 45%. The herb–dwarf shrub layer is dominated by Chimaphila umbellata, Vaccinium vitis-idaea, Orthilia secunda, Rubus saxatilis, and Calamagrostis arundinacea, with cover ranging from 5 to 35%.
The moss layer dominated by Dicranum polysetum, Pleurozium schreberi, and Hylocomium splendens has a cover of 50%–85%.
In the Tyumen region, there are plant communities with a similar floristic composition (comm. Calamagrostis epigeios-Pinus sylvestris, Cnidium dubium-Pinus sylvestris, Calamagrostis epigeios-Pinus sylvestris, Eremogone saxatilis-Pinus sylvestris), representing different stages of post-fire succession [24]. By floral composition, these communities are very similar to the association Peucedano-Pinetum Matuszkiewicz 1962, uniting common xerophytic species with high constancy, such as Viola rupestris, Cnidium dubium, Carex supina, Dianthus chinensis, Trifolium lupinaster, and Silene nutans. At the same time, the association Psephello sumensis-Pinetum sylvestris differs in its more developed moss cover and in the presence of species of high constancy typical of Western Siberia, such as Centaurea sumensis, Pulsatilla uralensis, Eremogone saxatilis, and Vincetoxicum hirundinaria (Table A2).
4. Discussion
4.1. Features of the Floristic Composition of Oligotrophic Green Moss Forests of SUR and Reasons for Need of a New Vegetation Alliance
The cluster analysis of the floristic composition revealed that the boreal oligotrophic pine forests of the SUR form a separate cluster (Figure 3). In the boreal oligotrophic pine forests of the SUR, the group of diagnostic species of Dicrano-Pinion has low phytocoenotic significance, being represented by species which have large ranges and grow in oligotrophic and moderately oligotrophic forests throughout Eurasia (Calamagrostis arundinacea, Polygonatum odoratum, Luzula pilosa, Fragaria vesca). At the same time, oligotrophic pine forests of the SUR differ from the communities of the alliance Hieracio-Pinion growing in the conditions of dry ultra-continental climate of Southern Siberia, by the absence of such species as Carex pediformis, Iris ruthenica, Viola dactyloides, Pyrola asarifolia, Artemisia tanacetifolia, Rhododendron dauricum, Spiraea media, Lathyrus humilis, Aquilegia sibirica, Actaea cimicifuga, etc., which are distributed mainly in the southern part of Northern Asia and do not occur in the SUR.
The boreal oligotrophic pine forests of the SUR combine their high ecological and syntaxonomic diversity, as well as the presence of both a group of mesophytic species typical for European broadleaf, dark coniferous, and mixed forests (e.g., Poa nemoralis, Aegopodium podagraria, Melica nutans, Lathyrus vernus, Stellaria holostea, Rubus idaeus, Viola mirabilis) and species growing in Siberian hemiboreal forests of the class Brachypodio-Betuletea (e.g., Trifolium lupinaster, Geranium pseudosibiricum, Poa sibirica, Lilium martagon, Viola collina, Brachypodium pinnatum, Adenophora liliifolia, Pleurospermum uralense, Seseli krylovii, Lathyrus pisiformis, Bupleurum longifolium). The combination of these species determines the specificity of boreal oligotrophic pine forests of the SUR and distinguishes these communities from the alliances Dicrano-Pinion and Hieracio-Pinion.
In this regard, we decided to separate a new alliance Brachypodio pinnati-Pinion sylvestris, which unites oligotrophic boreal pine forests of the SUR. This alliance is ecologically close to the mountain communities of the South Siberian alliance Hieracio-Pinion, and, apparently, is a vicariant alliance in the Southern Urals part of the range of the boreal oligotrophic pine forests of the order Pinetalia sylvestris.
The communities of alliance Brachypodio pinnati-Pinion sylvestris represent a transition from the alliance Dicrano-Pinion, which unites European oligotrophic pine forests of the temperate zone and the hemiboreal subzone, to the alliance Hieracio umbellati-Pinion sylvestris (boreal oligotrophic xeromesophytic pine forests growing in the southern part of the forest zone and in the forest–steppe zone of Siberia).
4.2. Analysis of the Floristic and Syntaxonomical Position of the West Siberian Dry Oligotrophic Pine Forests of the Association Psephello sumensis-Pinetum sylvestris
The numerical classification of the entire data set using Ward’s method indicated a high degree of floristic similarity of the West Siberian low-ranking syntaxa (e.g., the rangless communities Calamagrostis epigeios-Pinus sylvestris, Cnidium dubium-Pinus sylvestris, Eremogone saxatilis-Pinus sylvestris, and the association Psephello sumensis-Pinetum sylvestris) with the European communities of the alliance Dicrano-Pinion.
At the same time, important similarities were revealed between the floristic compositions of the communities of ass. Psephello sumensis-Pinetum sylvestris and the continental European pine forests of the alliance Dicrano-Pinion (Table S1, Figure 3). Compared to other Siberian boreal oligotrophic pine forests, the communities of ass. Psephello sumensis-Pinetum sylvestris lack many Asian species (e.g., Oxytropis campanulata, Bupleurum scorzonerifolium, Iris ruthenica, Spiraea media, Lathyrus humilis, Pyrola asarifolia, etc.). It can be assumed that the absence of these species is due to the isolation of the Kurgan region from eastern regions of Siberia by the West Siberian lowlands. Therefore, we decided to place this association into the alliance Dicrano-Pinion (Figure 3). Thus, the communities of the alliance Dicrano-Pinion do not occur in the western foothills of the Southern Urals [2], but the eastern boundary of this alliance reaches the terraces of the Ob and Tom rivers in the south of Western Siberia [78]. The latter statement is controversial and requires further research.
4.3. Specific Features and the Need for the Protection of Oligotrophic Boreal Pine Forests in the SUR and Kurgan Region
Boreal oligotrophic pine forests of the alliance Brachypodio pennatae-Pinion sylvestris require special conservation measures due to their stenotopic habitats and limited distribution range. In a certain sense, the communities of the alliance Brachypodio pinnati-Pinion sylvestris can be considered an ecological analogue of the pine forests of Europe in the early Holocene, which were later replaced by the forests of the alliance Dicrano-Pinion, which are protected by UE legislation as the Natura 2000 habitat type code 91T0 [79,80] and needs active protection to preserve [81]. This makes the forests of this alliance a promising object for monitoring climate change.
Due to the limited distribution area and stenotopic habitats, boreal oligotrophic pine forests of the alliance Brachypodio pennatae-Pinion sylvestris require special measures for their conservation. In the SUR, these forests were subjected to intensive deforestation, and the floristic composition of these communities was irreversibly transformed. The felled areas were overgrown with grasses (including Calamagrostis spp.), the turf of which is unfavorable for the development of pine seedlings [82,83,84,85,86,87,88]. In terms of forestry activities, the felled areas dominated by reed are considered as unfavorable sites for the regeneration of coniferous trees in the European part of Russia and in the Urals [89,90].
In the Southern Urals areas, where the boreal oligotrophic forests were felled, secondary long-term birch grass forests of the alliance Trollio-Pinion Fedorov ex Ermakov et al., 2000 (class Brachypodio-Betuletea Ermakov, Koroljuk et Latchinsky 1991) are formed as a result of natural reforestation. This has reduced the floristic and phytocenotic diversity of these forests [91].
As practice shows, for the majority of coniferous forests, including the Southern Urals oines, maximum conservation of biodiversity can be ensured by creating a system of fellings that imitate periodic natural disturbances (windfalls, gap-mosaic, etc.) and creating continuous rows in a certain area trees of different ages and sizes, and rotten wood at different stages of decomposition [92,93,94]. Numerous studies have shown that if more than 30% of trees are retained during logging, this helps to conserve the majority of species typical of old-growth forests [95], including rare species. However, forest users and loggers often ignore sustainable logging methods and measures to promote forest restoration. Success in natural regeneration can only be achieved at the legislative level, as for example in Finland and Sweden, which have documented the development of the forest industry towards greening and conservation of biological diversity [96]. In Russia, there is also an increase in the interest of legislators in forestry problems; responsible forest users are emerging who are ready to introduce new logging technologies.
In contrast to the mountain pine forests of the alliance Brachypodio pinnati-Pinion sylvestris, the oligotrophic green-moss pine forests of ass. Psephello sumensis-Pinetum sylvestris are well regenerated by pine after felling and do not require special measures for their protection. This is probably due to the fact that these forests grow on alluvial sandy soils on the river terraces near the southernmost border of the southern taiga subzone in Western Siberia.
5. Conclusions
A new alliance Brachypodio pinnati-Pinion sylvestris is described based on original and first published data. This alliance unites five associations of oligotrophic boreal pine forests from the Southern Ural region. Additionally, a new association of oligotrophic pine forests Psephello sumensis-Pinetum sylvestris ass. nov. hoc loco is described from the Kurgan region of Western Siberia. We ordered these communities into the alliance Dicrano-Pinion, which confirms the idea that the distribution area of Dicrano-Pinion may reach Siberia.
We conducted a comparative analysis of our data on the oligotrophic pine forest from the Southern Urals and Western Siberia with published works on the pine forest in Europe and other parts of Siberia, which allowed us to establish that pine forests from the Southern Urals are characterized by a special floristic composition, occupying an intermediate position between the typical European oligotrophic pine forests of the alliance Dicrano-Pinion and the oligotrophic (mainly psammophilous) South Siberian pine forests of the alliance Hieracio-Pinion.
Thus, despite the similarity of habitat conditions, boreal oligotrophic forests in Europe, the Southern Urals and Siberia have clearly expressed regional specifics and different histories of nature management. Therefore, these differences should be taken into account when planning measures to protect these forests at the regional level.
In the Southern Urals, due to intensive forestry activities, the distribution area of boreal oligotrophic forests has decreased, and these communities have been replaced by secondary birch forests. To preserve these communities, we proposed a set of regional conservation measures. In the Kurgan region, the recovery of pine forests after felling is quite high, and these communities do not require special measures for their protection.
The results of this work allow us to use syntaxonomical data for long-term monitoring and mapping of forest vegetation in the Southern Ural region. Knowledge about ecological and floristic features of boreal oligotrophic pine forests is useful for predicting the future dynamics of forest vegetation in the study area.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f15081461/s1, Table S1: Shortened synoptic of the associations of alliance Brachypodio-Pinion and their comparison with the alliances Cladonio stellaris-Pinion, Dicrano-Pinion and Hieracio-Pinion; Table S2: References to the entire data set which included 61 low-rank vegetation units represented in the field “Syntaxon number” of Table S1; Table S3: Full synoptic table of the boreal light coniferous forest vegetation in Southern Ural region; Table S4: Species composition of the association Pleurospermo uralensis-Pinetum sylvestris; Table S5: Type relevés of association Pleurospermo uralensis-Pinetum sylvestris: locations and site characteristics; Table S6: Species composition of the association Psephello sumensis-Pinetum sylvestris; Table S7: Type relevés of association Psephello sumensis-Pinetum sylvestris: locations and site characteristics.
Author Contributions
Conceptualization, V.M. and P.S.; methodology, V.M., P.S. and N.F.; software, N.F. and P.S.; validation, V.M., P.S., E.B., A.M. and S.Z.; formal analysis, L.N. and P.S.; investigation, V.M., P.S., A.M., N.F. and E.B.; data curation, V.M. and P.S.; writing—original draft preparation, P.S., E.B. and S.Z.; writing—review and editing, P.S., V.M., E.B., L.N. and N.F; visualization, P.S. and S.Z.; project administration P.S. All authors have read and agreed to the published version of the manuscript.
Funding
The study was was supported by the grant of Russian Science Foundation No. 22-14-00003, https://rscf.ru/en/project/22-14-00003/.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A

Table A1.
Shortened synoptic table of the boreal light coniferous forest vegetation in Southern Ural region.
Table A1.
Shortened synoptic table of the boreal light coniferous forest vegetation in Southern Ural region.
| Syntaxon number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| Number of relevés | 14 | 11 | 15 | 13 | 16 | 9 | 18 | 10 | 8 | |
| Cover of tree layer (%) | 55 | 50 | 60 | 50 | 55 | 50 | 55 | 65 | 65 | |
| Cover of shrub layer (%) | 1 | 2 | 2 | 3 | 3 | 1 | 1 | 2 | 5 | |
| Cover of herb layer (%) | 20 | 25 | 35 | 45 | 35 | 50 | 45 | 25 | 35 | |
| Cover of moss layer (%) | 65 | 65 | 40 | 50 | 30 | 65 | 70 | 80 | 20 | |
| Mean number of vascular plant | 49.5 | 54.0 | 51.4 | 62.8 | 67.9 | 50.2 | 41.3 | 61.9 | 70.9 | |
| Mean number of bryophytes | 6.4 | 7.1 | 7.3 | 7.8 | 6.1 | 7.8 | 7.5 | 14.6 | 11.0 | |
| Mean number of lichens | 10.8 | 10.7 | 10.7 | 8.9 | 9.9 | 7.0 | 11.5 | 10.3 | 9.8 | |
| Dominants of tree layer | ||||||||||
| Pinus sylvestris | 1 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Larix sukaczewii | 1 | 86 | 82 | 80 | 17 | 13 | 30 | . | . | . |
| Betula pendula | 1 | 7 | . | . | 58 | 63 | 40 | 33 | 50 | 63 |
| Betula pubescens | 1 | . | . | . | . | . | 60 | 28 | 60 | 38 |
| Picea obovata | 1 | . | . | . | . | . | . | 6 | 50 | 50 |
| Ass. Violo rupestris-Pinetum sylvestris and var. typica | ||||||||||
| Larix sukaczewii | 2 | 93 | 91 | 80 | 17 | 6 | . | . | . | . |
| Larix sukaczewii | 3 | 64 | 91 | 93 | 17 | 13 | 20 | . | . | . |
| Persicaria alpina | 6 | 93 | 100 | 33 | 8 | . | . | . | 20 | . |
| Festuca rupicola | 6 | 43 | 100 | 7 | . | . | . | . | . | . |
| Silene viscaria | 6 | 64 | 73 | 13 | . | . | . | 11 | . | . |
| Filipendula vulgaris | 6 | 50 | 55 | 7 | 25 | 19 | 10 | . | . | . |
| Silene amoena | 6 | 64 | 64 | . | . | . | . | . | 10 | 38 |
| Galium ruthenicum | 6 | 43 | 36 | . | . | . | . | . | . | . |
| Dianthus chinensis | 6 | 36 | 82 | . | 8 | . | . | . | . | . |
| Poa lapponica | 6 | 29 | 45 | 13 | . | . | . | . | 30 | . |
| Dracocephalum ruyschiana | 6 | 21 | 73 | 7 | 8 | . | . | 6 | 10 | . |
| Viola rupestris | 6 | 71 | 91 | 13 | 25 | . | . | 6 | 20 | 25 |
| Lathyrus pratensis | 6 | 64 | 27 | 40 | . | 6 | 10 | . | . | . |
| Var. Veronica spicata | ||||||||||
| Pseudolysimachion spicatum | 6 | 7 | 91 | . | . | . | . | 6 | 10 | 13 |
| Carex caryophyllea | 6 | 14 | 82 | 7 | 25 | 19 | . | . | 10 | . |
| Aster alpinus | 6 | 7 | 64 | . | 8 | . | . | . | . | . |
| Centaurea sibirica | 6 | . | 45 | . | . | . | . | . | 40 | 50 |
| Tephroseris integrifolia | 6 | . | 55 | . | 25 | . | . | . | 40 | 50 |
| Rosa acicularis | 4 | . | 55 | . | 8 | . | . | . | . | . |
| Polygala comosa | 6 | 7 | 45 | . | . | . | . | . | . | 13 |
| Ass. Pleurospermo uralensis-Pinetum sylvestris and subass. P.u.-P.s. laricetosum sukazhewii | ||||||||||
| Pleurospermum uralense | 6 | 21 | 9 | 80 | 67 | 69 | 20 | 28 | 40 | 88 |
| Geranium sylvaticum | 6 | 43 | . | 80 | 58 | 100 | 90 | 67 | 40 | 63 |
| Atragene sibirica | 4 | 14 | 9 | 73 | 67 | 81 | 20 | 11 | 70 | 100 |
| Angelica sylvestris | 6 | 7 | . | 93 | 50 | 88 | 90 | 17 | . | . |
| Hieracium albocostatum | 6 | . | 9 | 40 | 17 | 75 | 20 | 6 | . | . |
| Milium effusum | 6 | . | . | 27 | 50 | 38 | . | . | . | . |
| Pyrola minor | 6 | 7 | . | 80 | 25 | 94 | 70 | 39 | 10 | . |
| Aconitum lycoctonum | 6 | . | . | 13 | 42 | 31 | 30 | . | 10 | 13 |
| Pulmonaria mollissima | 6 | 7 | . | 7 | 75 | 100 | 60 | 39 | 10 | 63 |
| Succisa pratensis | 6 | . | . | 20 | 42 | 100 | 100 | 72 | . | . |
| Subass. P.u.-P.s. digitalietosum grandiflorae and var. typica | ||||||||||
| Potentilla erecta | 6 | . | . | 13 | 33 | 88 | 90 | 28 | . | . |
| Primula macrocalyx | 6 | 7 | 27 | . | 92 | 63 | 30 | 6 | 20 | 75 |
| Digitalis grandiflora | 6 | 7 | 18 | . | 83 | 100 | 70 | 89 | . | 50 |
| Aegopodium podagraria | 6 | . | . | . | 67 | 88 | 30 | 22 | 20 | 63 |
| Bupleurum longifolium | 6 | . | . | . | 67 | 94 | 40 | 11 | 30 | 50 |
| Stachys officinalis | 6 | . | . | . | 33 | 88 | 50 | 39 | . | 13 |
| Padus avium | 3 | 7 | . | . | 58 | 69 | 30 | 28 | 20 | 50 |
| Cerastium pauciflorum | 6 | . | . | . | 50 | 63 | 60 | 22 | 40 | 50 |
| Trollius europaeus | 6 | . | . | . | 58 | 75 | 60 | . | 10 | . |
| Adonis apennina | 6 | . | . | . | 42 | 44 | . | . | 50 | 38 |
| Lathyrus gmelinii | 6 | 7 | . | 7 | 42 | 50 | 40 | 6 | . | . |
| var. Cirsium heterophyllum | ||||||||||
| Cirsium heterophyllum | 6 | . | . | 13 | 17 | 81 | 40 | 6 | . | . |
| Carex rhizina | 6 | . | . | . | 8 | 81 | . | 28 | 70 | 100 |
| Daphne mezereum | 4 | . | . | . | 8 | 63 | 10 | 6 | 20 | 25 |
| Pedicularis uralensis | 6 | . | . | . | 25 | 50 | . | . | 20 | . |
| Epipactis helleborine | 6 | . | . | . | 8 | 50 | . | . | . | . |
| Hypericum maculatum | 6 | . | . | . | . | 50 | 10 | 6 | . | . |
| Ranunculus auricomus | 6 | . | . | . | . | 44 | 10 | . | 10 | . |
| Subass. P.u.-P.s. anemonastretosum biarmiensis | ||||||||||
| Molinia caerulea | 6 | . | . | . | . | . | 100 | . | . | . |
| Anemonastrum biarmiense | 6 | . | . | . | 17 | . | 100 | . | . | . |
| Carex montana | 6 | . | . | . | 8 | . | 100 | 6 | 20 | . |
| Hypochaeris maculata | 6 | . | . | . | 25 | . | 90 | 17 | . | . |
| Lycopodium annotinum | 6 | . | . | . | . | 25 | 80 | . | . | . |
| Ass. Seseli krylovii-Pinetum sylvestris | ||||||||||
| Vaccinium vitis-idaea | 6 | 29 | 18 | 27 | 8 | . | 100 | 83 | . | . |
| Abies sibirica | 3 | . | . | . | . | . | 50 | 83 | 90 | 63 |
| Picea obovata | 3 | . | . | . | 8 | . | 80 | 89 | 100 | 75 |
| Betula pubescens | 2 | . | . | . | . | . | 90 | 44 | 60 | 63 |
| Betula pubescens | 3 | . | . | . | 8 | . | 70 | 56 | 70 | 75 |
| Lonicera xylosteum | 4 | . | . | . | 33 | . | . | 33 | 70 | 75 |
| Carex macroura | 6 | . | . | . | 8 | . | . | 56 | 40 | 25 |
| Tilia cordata | 3 | . | . | . | 17 | . | . | 72 | 70 | 100 |
| Carex pilosa | 6 | . | . | . | . | . | . | 44 | . | . |
| Ass. Zigadeno sibirici-Pinetum sylvestris | ||||||||||
| Anticlea sibirica | 6 | . | . | . | . | . | . | 6 | 80 | 75 |
| Carex alba | 6 | . | . | . | . | . | . | . | 90 | 63 |
| Galium tinctorium | 6 | . | . | . | 8 | . | . | 6 | 40 | 75 |
| Valeriana wolgensis | 6 | . | . | . | 17 | 13 | . | . | 70 | 63 |
| Campanula rotundifolia | 6 | 7 | 9 | . | . | . | . | 17 | 90 | 100 |
| Gymnocarpium robertianum | 6 | . | . | . | . | . | . | . | 90 | 88 |
| Frangula dodonei | 4 | . | . | . | . | . | . | . | 80 | 50 |
| Cortusa matthioli | 6 | . | . | . | . | . | . | . | 70 | 50 |
| Arabidopsis arenosa | 6 | . | . | . | . | . | . | . | 70 | 38 |
| Equisetum scirpoides | 6 | . | . | . | . | . | . | . | 80 | 50 |
| Galium album | 6 | 7 | 9 | . | . | . | . | . | 40 | 38 |
| Primula cortusoides | 6 | . | . | . | . | . | . | . | 70 | 63 |
| Caragana frutex | 4 | . | . | . | . | . | . | 6 | 50 | 25 |
| Var. Cystopteris fragilis | ||||||||||
| Cystopteris fragilis | 6 | . | 9 | . | 17 | . | . | . | 90 | . |
| Cerastium uralense | 6 | . | . | . | . | . | . | . | 50 | . |
| Euphorbia microcarpa | 6 | . | . | . | . | . | . | . | 40 | . |
| Var. Viburnum opulus | ||||||||||
| Viburnum opulus | 3 | . | . | . | 8 | . | . | 6 | 10 | 75 |
| Tilia cordata | 2 | . | . | . | 8 | . | . | . | 10 | 50 |
| Populus tremula | 3 | . | . | . | 17 | 13 | 10 | 6 | . | 63 |
| Astragalus danicus | 6 | . | . | . | 8 | . | . | . | . | 50 |
| Ulmus glabra | 3 | . | . | . | . | . | . | . | . | 50 |
| All. Brachypodio-Pinion | ||||||||||
| Viola collina | 6 | 93 | 91 | 93 | 83 | 100 | 50 | 61 | 100 | 100 |
| Brachypodium pinnatum | 6 | 100 | 100 | 100 | 83 | 100 | 90 | 72 | 30 | 63 |
| Adenophora lilifolia | 6 | 100 | 100 | 100 | 83 | 81 | 40 | 39 | 90 | 88 |
| Seseli krylovii | 6 | 100 | 100 | 100 | 42 | 88 | 10 | 72 | 100 | 100 |
| Lathyrus pisiformis | 6 | 100 | 91 | 100 | 50 | 31 | 20 | 39 | 10 | 50 |
| Persicaria bistorta | 6 | 7 | 9 | 20 | 17 | 56 | 20 | 11 | 70 | 75 |
| Poa sibirica | 6 | 71 | 36 | 87 | 67 | 81 | 10 | . | 10 | 13 |
| Campanula glomerata | 6 | 100 | 91 | 100 | 67 | 100 | 40 | . | 10 | 38 |
| Lilium martagon | 6 | 64 | 45 | 73 | 67 | 88 | 50 | 22 | 80 | 88 |
| Trifolium lupinaster | 6 | 100 | 91 | 100 | 75 | 100 | 80 | 39 | 70 | 50 |
| Geranium pseudosibiricum | 6 | 100 | 100 | 27 | 67 | 56 | 10 | . | 30 | 25 |
| All. Dicrano-Pinion | ||||||||||
| Pinus sylvestris | 2 | 93 | 100 | 100 | 92 | 100 | 80 | 100 | 100 | 100 |
| Antennaria dioica | 6 | 93 | 91 | 53 | 42 | 69 | 50 | 56 | 20 | . |
| Pinus sylvestris | 3 | 79 | 82 | 80 | 58 | 44 | 90 | 89 | 90 | 100 |
| Betula pendula | 3 | 100 | 100 | 100 | 83 | 100 | 30 | 33 | 60 | 88 |
| Dicranum polysetum | 9 | 71 | 82 | 67 | 42 | 44 | 20 | 78 | 100 | 88 |
| Juniperus communis | 4 | 50 | 45 | 60 | . | . | 10 | . | . | . |
| Anemone patens | 6 | 100 | 100 | 67 | 67 | 25 | 50 | 22 | 60 | 50 |
| Melampyrum pratense | 6 | . | . | . | 17 | 44 | 70 | 83 | . | . |
| Quercus robur | 3 | . | . | . | 8 | . | . | 44 | 70 | 75 |
| Pteridium aquilinum | 6 | . | . | 13 | 33 | 13 | . | 11 | 20 | 63 |
| Veronica officinalis | 6 | . | . | . | 17 | 19 | . | 11 | . | . |
| Moneses uniflora | 6 | 7 | . | 7 | 25 | 6 | . | 6 | 30 | . |
| Pyrola chlorantha | 6 | 21 | 9 | 20 | . | 31 | . | 28 | . | . |
| Chimaphila umbellata | 6 | . | . | . | . | . | . | 6 | 10 | . |
| Diphasiastrum complanatum | 6 | . | . | . | . | . | 10 | 6 | . | . |
| Calamagrostis epigeios | 6 | . | . | . | 8 | . | . | . | . | . |
| Cl. Vaccinio-Piceetea and Ord. Piceetalia excelsae | ||||||||||
| Pleurozium schreberi | 9 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Hylocomium splendens | 9 | 100 | 100 | 100 | 92 | 81 | 90 | 89 | 80 | 75 |
| Vaccinium myrtillus | 6 | 86 | 100 | 100 | 67 | 75 | 90 | 83 | . | . |
| Dicranum scoparium | 9 | 50 | 73 | 60 | 67 | 31 | 40 | 78 | 70 | 75 |
| Ptilium crista-castrensis | 9 | 100 | 100 | 87 | 67 | 100 | 80 | 89 | 60 | 50 |
| Orthilia secunda | 6 | 100 | 82 | 100 | 75 | 94 | 70 | 78 | 100 | 100 |
| Goodyera repens | 6 | 57 | 55 | 93 | 44 | 44 | 50 | 6 | . | . |
| Pyrola rotundifolia | 6 | . | . | . | . | 19 | 20 | 50 | 20 | 13 |
| Picea obovata | 2 | . | . | . | . | . | . | 33 | 50 | 38 |
| Abies sibirica | 2 | . | . | . | . | . | . | 33 | 10 | 13 |
| Linnaea borealis | 6 | . | . | . | . | . | 40 | 6 | 10 | . |
| Cl. Asaro-Abietetea and Ord. Abietetalia sibiricae | ||||||||||
| Maianthemum bifolium | 6 | 7 | . | 80 | 58 | 75 | 100 | 61 | 50 | 50 |
| Trientalis europaea | 6 | 43 | 9 | 100 | 58 | 100 | 100 | 33 | 10 | 25 |
| Rhytidiadelphus triquetrus | 9 | 86 | 91 | 87 | 83 | 94 | 80 | 6 | 70 | 88 |
| Luzula pilosa | 6 | 100 | 91 | 100 | 83 | 100 | 90 | 94 | 10 | 25 |
| Oxalis acetosella | 6 | . | . | . | 17 | . | . | 6 | . | . |
| Athyrium filix-femina | 6 | . | . | . | . | . | . | . | 20 | . |
| Gymnocarpium dryopteris | 6 | . | . | 33 | . | . | . | 6 | 10 | 13 |
| Cl. Querco-Fagetea | ||||||||||
| Lathyrus vernus | 6 | 100 | 91 | 93 | 92 | 100 | 100 | 72 | 30 | 100 |
| Melica nutans | 6 | 93 | 91 | 100 | 100 | 100 | 70 | 83 | 60 | 88 |
| Sorbus aucuparia | 3 | 100 | 100 | 100 | 92 | 100 | 90 | 94 | 100 | 100 |
| Stellaria holostea | 6 | 21 | 36 | 93 | 83 | 88 | 70 | 33 | 30 | 63 |
| Viola mirabilis | 6 | 50 | 18 | 87 | 75 | 88 | 10 | 6 | 70 | 88 |
| Poa nemoralis | 6 | 7 | . | . | 67 | 69 | 10 | 33 | 50 | 63 |
| Rubus idaeus | 4 | . | . | 13 | 33 | 25 | . | 28 | . | 13 |
| Euonymus verrucosa | 4 | . | . | . | . | . | . | . | 50 | 75 |
| Cl. Brachypodio-Betuletea and Ord. Chamaecytiso-Pinetalia | ||||||||||
| Calamagrostis arundinacea | 6 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Rubus saxatilis | 6 | 100 | 100 | 100 | 100 | 94 | 100 | 100 | 100 | 100 |
| Hieracium umbellatum | 6 | 100 | 100 | 100 | 83 | 88 | 90 | 94 | 40 | 50 |
| Silene nutans | 6 | 100 | 100 | 67 | 67 | 63 | 20 | 50 | 30 | 75 |
| Carex digitata | 6 | 100 | 100 | 100 | 92 | 100 | 10 | 39 | 60 | 88 |
| Viola canina | 6 | 93 | 64 | 87 | 75 | 100 | 100 | 72 | . | . |
| Chamaecytisus ruthenicus | 4 | 100 | 100 | 100 | 75 | 100 | 100 | 44 | 100 | 100 |
| Betula pendula | 2 | 57 | 55 | 73 | 83 | 81 | 40 | 33 | 50 | 75 |
| Vicia sepium | 6 | 43 | 27 | 93 | 50 | 63 | 40 | 22 | 10 | . |
| Thalictrum minus | 6 | 7 | 18 | 33 | 50 | 19 | . | 6 | 30 | 25 |
| Veronica chamaedrys | 6 | 14 | . | 7 | 67 | 81 | 60 | 33 | . | . |
| Trifolium medium | 6 | 7 | . | . | 50 | 44 | . | 6 | . | 25 |
| Cerasus fruticosa | 4 | . | . | . | . | . | . | . | 20 | 63 |
| Other constant species | ||||||||||
| Solidago virgaurea | 6 | 100 | 100 | 100 | 100 | 100 | 90 | 100 | 100 | 100 |
| Polygonatum odoratum | 6 | 86 | 100 | 53 | 58 | 69 | 40 | 72 | 40 | 88 |
| Fragaria vesca | 6 | 93 | 100 | 100 | 92 | 94 | 50 | 83 | 70 | 75 |
| Galium boreale | 6 | 100 | 100 | 100 | 92 | 100 | 90 | 94 | 100 | 100 |
| Euphorbia gmelinii | 6 | 93 | 100 | 93 | 58 | 38 | . | 11 | 30 | 75 |
| Sanguisorba officinaliss | 6 | 100 | 100 | 100 | 58 | 100 | 100 | 17 | 90 | 63 |
| Achillea millefolium | 6 | 100 | 100 | 100 | 50 | 75 | 10 | 28 | 50 | 50 |
| Crepis praemorsa | 6 | 93 | 73 | 87 | 42 | 94 | . | 11 | 10 | 13 |
| Campanula persicifolia | 6 | 71 | 73 | 53 | 67 | 100 | . | . | 30 | 13 |
| Seseli libanotis | 6 | 29 | 45 | 13 | 42 | . | 10 | 6 | . | 25 |
| Rosa majalis | 4 | . | . | 7 | 67 | 63 | 10 | 44 | 60 | 63 |
| Sedum telephium | 6 | 43 | 55 | . | 33 | 31 | . | 22 | 60 | 50 |
| Salix caprea | 3 | 14 | 9 | 27 | 17 | 13 | . | 17 | 20 | . |
| Moehringia lateriflora | 6 | . | . | 20 | 75 | 19 | 20 | 11 | 70 | 75 |
| Platanthera bifolia | 6 | . | . | 13 | 33 | 13 | 60 | 22 | 10 | . |
| Heracleum sibiricum | 6 | 14 | . | 47 | 17 | 25 | . | . | . | 25 |
| Ranunculus monophyllus | 6 | . | . | . | 25 | 31 | 10 | . | 10 | . |
| Origanum vulgare | 6 | 29 | 27 | . | 17 | 25 | . | 22 | 20 | 13 |
| Vicia sylvatica | 6 | . | . | 13 | 58 | 25 | . | 33 | . | 13 |
| Dactylis glomerata | 6 | 7 | . | . | 17 | 56 | 20 | . | . | . |
| Moehringia trinervia | 6 | 29 | 27 | 33 | 8 | . | . | 22 | . | . |
| Vicia cracca | 6 | 14 | . | 20 | 33 | 13 | 20 | . | 70 | 100 |
| Saussurea controversa | 6 | . | . | . | 42 | 25 | . | 6 | 100 | 88 |
| Cotoneaster melanocarpus | 4 | . | . | . | 50 | 13 | 10 | 28 | 50 | 75 |
| Epilobium dodonaei | 6 | . | . | 7 | 17 | 25 | . | . | 30 | 50 |
| Artemisia armeniaca | 6 | 7 | . | 7 | 25 | 13 | . | 6 | 50 | 75 |
| Vincetoxicum albovianum | 6 | . | . | . | . | . | . | . | 30 | 63 |
| Cardamine trifida | 6 | . | . | . | . | . | . | . | 20 | 38 |
| Epipactis atrorubens | 6 | . | . | . | . | . | . | . | 30 | 25 |
| Cypripedium guttatum | 6 | . | . | . | 17 | . | 30 | . | 40 | 25 |
| Poa trivialis | 6 | . | . | . | . | . | . | . | 20 | 38 |
| Salix bebbiana | 4 | . | . | . | . | . | . | . | 30 | 25 |
| Adoxa moschatellina | 6 | . | . | . | 8 | . | . | . | 20 | 25 |
| Paris quadrifolia | 6 | . | . | . | . | 13 | . | . | 20 | 25 |
| Stellaria bungeana | 6 | . | . | . | . | . | . | . | 20 | 38 |
| Equisetum pratense | 6 | . | . | . | . | . | . | . | 30 | 25 |
| Acer platanoides | 3 | . | . | 7 | . | . | . | 17 | 30 | 38 |
Note: The numbers in the table are percentage occurrence frequencies (constancies). Light and dark grey backgrounds indicate fidelity of Φ > 0.25 and Φ > 0.5. Diagnostic species are framed. Syntaxons: 1—var. typica of ass. Violo rupestris-Pinetum sylvestris, 2—var. typica of ass. Violo rupestris-Pinetum sylvestris, 3—subass. P.u.-P.s. laricetosum sukazhewii, 4—var. typica of subass. P.u.-P.s. digitalietosum grandiflorae, 5—var. Cirsium heterophyllum of subass. P.u.-P.s. digitalietosum grandiflorae, 6—subass. P.u.-P.s. anemonastretosum biarmiensis, 7—ass. Seseli krylovii-Pinetum sylvestris, 8—var. Cystopteris fragilis of ass. Zigadeno sibirici-Pinetum sylvestris, 9—var. Viburnum opulus of ass. Zigadeno sibirici-Pinetum sylvestris. Layers: 1—first tree layer, 2—second tree layer, 3—third tree layer, 4—shrub layer, 6—herb layer, 9—moss layer, 0—lichens layer. A full synoptic table of the boreal light-coniferous forest vegetation in South Ural region is provided in the Supplementary Materials (Table S3).
Table A2.
Shortened synoptic table of the light coniferous green moss forest vegetation in the Tyumen and Kurgan regions (Western Siberia).
Table A2.
Shortened synoptic table of the light coniferous green moss forest vegetation in the Tyumen and Kurgan regions (Western Siberia).
| Syntaxon number | 1 | 2 | 3 | 4 | |
| Number of relevés | 9 | 15 | 10 | 16 | |
| Cover of tree layer (%) | 27 | 28 | 24 | 40 | |
| Cover of herb layer (%) | 13 | 15 | 40 | 15 | |
| Cover of moss layer (%) | 1 | 54 | 17 | 65 | |
| Mean number of vascular plant | 18.6 | 20.1 | 30.8 | 34.6 | |
| Mean number of bryophytes | 0.8 | 2.8 | 1.5 | 6.4 | |
| Mean number of lichens | 0.3 | 0.9 | 0 | 4.7 | |
| Dominants of tree layer | |||||
| Pinus sylvestris | 1 | 100 | 100 | 100 | 100 |
| Betula pendula | 1 | . | 13 | 40 | . |
| Betula krylovii | 1 | . | . | . | 50 |
| Comm. Cnidium dubium-Pinus sylvestris | |||||
| Viola montana | 6 | . | 13 | 60 | . |
| Fragaria viridis | 6 | . | 7 | 60 | . |
| Lycopodium clavatum | 6 | 11 | 13 | 50 | 6 |
| Pteridium aquilinum | 6 | . | 7 | 40 | . |
| Geranium pratense | 6 | . | . | 40 | . |
| Calamagrostis canescens | 6 | . | 7 | 30 | . |
| Ass. Psephello sumensis-Pinetum sylvestris | |||||
| Betula krylovii | 2 | . | . | . | 63 |
| Betula krylovii | 3 | . | . | . | 81 |
| Pseudolysimachion spicatum | 6 | 67 | 40 | 10 | 88 |
| Eremogone saxatilis | 6 | 56 | 7 | . | 88 |
| Poa angustifolia | 6 | 11 | 7 | . | 75 |
| Astragalus danicus | 6 | . | . | 10 | 63 |
| Psephellus sumensis | 6 | . | . | . | 69 |
| Pulsatilla uralensis | 6 | . | . | . | 94 |
| Fragaria vesca | 6 | . | . | . | 69 |
| Vincetoxicum hirundinaria | 6 | . | . | . | 63 |
| All. Dicrano-Pinion | |||||
| Chimaphila umbellata | 6 | 100 | 93 | 100 | 100 |
| Calamagrostis arundinacea | 6 | 78 | 80 | 100 | 88 |
| Calamagrostis epigeios | 6 | 44 | 87 | 20 | 94 |
| Antennaria dioica | 6 | 89 | 73 | 40 | 88 |
| Dicranum polysetum | 9 | 22 | 100 | 30 | 94 |
| Polygonatum odoratum | 6 | 89 | 47 | 40 | 100 |
| Pinus sylvestris | 2 | 11 | 47 | 70 | 69 |
| Pinus sylvestris | 3 | . | . | . | 100 |
| Melampyrum pratense | 6 | . | 27 | 70 | 13 |
| Pinus sylvestris | 7 | 22 | 47 | 70 | 13 |
| Dryopteris carthusiana | 6 | . | 7 | 20 | 13 |
| Betula pendula | 2 | . | . | 50 | . |
| Frangula dodonei | 4 | . | 7 | 10 | 19 |
| Ord. Pinetalia sylvestris and Cl. Vaccinio-Piceetea | |||||
| Viola rupestris | 6 | 56 | 53 | 40 | 75 |
| Carex ericetorum | 6 | 100 | 87 | 40 | 94 |
| Pyrola chlorantha | 6 | 78 | 60 | 40 | 56 |
| Vaccinium vitis-idaea | 6 | 22 | 40 | 90 | 75 |
| Pleurozium schreberi | 9 | 56 | 100 | 80 | 100 |
| Orthilia secunda | 6 | 56 | 93 | 90 | 100 |
| Goodyera repens | 6 | 22 | 53 | 10 | 6 |
| Vaccinium myrtillus | 6 | 11 | 27 | 70 | 13 |
| Anemone patens | 6 | 78 | 80 | 60 | 6 |
| Dicranum scoparium | 9 | . | . | . | 50 |
| Hylocomium splendens | 9 | . | 80 | 40 | 94 |
| Trientalis europaea | 6 | . | . | 70 | 13 |
| Calluna vulgaris | 4 | . | . | . | 25 |
| Other constant species | |||||
| Equisetum hyemale | 6 | 56 | 73 | 70 | 75 |
| Hieracium umbellatum | 6 | 100 | 87 | 80 | 56 |
| Trifolium lupinaster | 6 | 89 | 93 | 100 | 88 |
| Solidago virgaurea | 6 | 100 | 93 | 90 | 88 |
| Rubus saxatilis | 6 | . | 47 | 100 | 100 |
| Cnidium dubium | 6 | 11 | . | 80 | 63 |
| Maianthemum bifolium | 6 | 11 | 27 | 100 | 44 |
| Carex supina | 6 | 67 | 47 | . | 6 |
| Dianthus chinensis | 6 | 56 | 47 | . | 13 |
| Achillea millefolium | 6 | 11 | 13 | 40 | 63 |
| Vicia cracca | 6 | . | 13 | 40 | 63 |
| Vicia sepium | 6 | 22 | 27 | 70 | 31 |
| Lathyrus vernus | 6 | 11 | 7 | 60 | 38 |
| Lathyrus pisiformis | 6 | . | 13 | 20 | 44 |
| Galium boreale | 6 | 11 | 7 | 40 | 50 |
| Silene nutans | 6 | 22 | 20 | 40 | 25 |
| Dracocephalum ruyschiana | 6 | . | . | 10 | 38 |
| Sorbus aucuparia | 3 | . | . | . | 56 |
| Viola canina | 6 | . | . | . | 38 |
| Rosa majalis | 4 | . | . | 10 | 44 |
| Sorbus aucuparia | 4 | . | 20 | 60 | . |
| Populus tremula | 4 | 11 | 7 | 70 | . |
| Pinus sylvestris | 4 | . | 7 | 40 | . |
| Brachypodium pinnatum | 6 | . | . | . | 31 |
| Pyrola rotundifolia | 6 | . | 20 | 40 | 6 |
| Diphasiastrum complanatum | 6 | 11 | 33 | 20 | 31 |
| Sorbus aucuparia | 7 | . | 13 | 40 | 6 |
| Melica nutans | 6 | . | . | 30 | 25 |
| Platanthera bifolia | 6 | . | 7 | 30 | 19 |
| Pimpinella saxifraga | 6 | . | 13 | 30 | . |
| Cladonia stellaris | 0 | 11 | 27 | . | . |
| Potentilla erecta | 6 | . | . | 30 | 13 |
| Equisetum sylvaticum | 6 | . | 7 | 30 | . |
| Populus tremula | 3 | . | . | . | 31 |
Note: The numbers in the table are percentage occurrence frequencies (constancies). Light and dark grey backgrounds indicate fidelity of Φ > 0.25 and Φ > 0.5, respectively. Diagnostic species are framed. Syntaxons: 1—comm. Eremogone saxatilis-Pinus sylvestris, 2—comm. Calamagrostis epigeios-Pinus sylvestris, 3—comm. Cnidium dubium-Pinus sylvestris, 4—ass. Psephello sumensis-Pinetum sylvestris. Layers: 1—first tree layer, 2—second tree layer, 3—third tree layer, 4—shrub layer, 6—herb layer, 9—moss layer, 0—lichens layer.
References
- Ermakov, N.; Morozova, O. Syntaxonomical survey of boreal oligotrophic pine forests in northern Europe and Western Siberia. Appl. Veg. Sci. 2011, 14, 524–536. [Google Scholar] [CrossRef]
- Ermakov, N.B. The higher units of pine forests of Russia in connection with the general concept of vegetation classification of Northern Eurasia. Plant Biol. Hortic. Theory Innov. 2020, 157, 94–113. [Google Scholar] [CrossRef]
- Willner, W. How to classify forests? A case study from Central Europe. Veg. Classif. Surv. 2024, 5, 17–26. [Google Scholar] [CrossRef]
- Mucina, L.; Grabherr, G.; Wallnöfer, S. Die Pflanzengesellschaften Ősterreichs. Teil 3. Wälder und Gebüsche; Gustav Fischer: Jena, Germany, 1993; 578p. [Google Scholar]
- Rodwell, J.S.; Schaminee, J.H.J.; Mucina, L.; Pignatii, S.; Dring, J.; Moss, D. The Diversity of European Vegetation—An Overview of Phytosociological Alliances and Their Relationships to EUNIS Habitats; National Reference Centre for Agriculture, Nature and Fisheries: Wageningen, UK, 2002; 168p. [Google Scholar]
- Dengler, J.; Chytry, M.; Ewald, J. Phytosociology. In General Ecology. Vol. 4 of Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; Volume 5, pp. 2767–2779. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer: Wien, Austria; New York, NY, USA, 1964; 631p. [Google Scholar] [CrossRef]
- Rysin, L.P. Pine Forests of the European Part of the USSR; Nauka: Moscow, Russia, 1975; 212p. [Google Scholar]
- Gribova, S.A.; Isachenko, T.I.; Lavrenko, E.M. Vegetation of the European Part of the USSR; Nauka: Leningrad, Russia, 1980; 429p. [Google Scholar]
- Oberdorfer, E. (Ed.) Süddeutsche Pflanzengesellshaften. Teil IV, 2. Auflage, B. Tabellenband; Springer Nature: Stuttgart, Germany, 1992. [Google Scholar]
- Daniëls, F.J.A. Vegetation classification in Greenland. J. Veg. Sci. 1994, 5, 781–790. [Google Scholar] [CrossRef]
- Korotkov, K.O. Valday Forests; Nauka: Moscow, Russia, 1991; 157p. [Google Scholar]
- Korotkov, K.O.; Morozova, O.V.; Belonovskaya, E.A. The USSR Vegetation Syntaxa Prodromus; Vilchek, G.E., Ed.; G.E.Vilchek: Moscow, Russia, 1991. [Google Scholar]
- Morozova, O.V.; Zaugol’nova, L.B.; Isaeva, L.G.; Kostina, V.A. Classification of boreal forests in the north of European Russia. I. Oligotrophic coniferous forests. Veg. Russ. 2008, 13, 61–81. [Google Scholar] [CrossRef]
- Bulokhov, A.D.; Solomeshch, A.I. Ecological and Floristic Classification of the Forest of the Southern Non-Black Earth Region; Izdatel’stvo BGU: Bryansk, Russia, 2003; 359p. [Google Scholar]
- Zaugol’nova, L.B.; Morozova, O.V. Forests of the boreal (taiga) strip. Distribution and classification of boreal forests. In Eastern European Forests; Smirnova, O.V., Ed.; Nauka: Moscow, Russia, 2004. [Google Scholar]
- Zaugol’nova, L.B.; Smirnova, O.V.; Braslavskaya, T.Y.; Degteva, S.V.; Prokazina, T.S.; Lugovaya, D.L. Tall herb boreal forests of the eastern part of European Russia. Veg. Russ. 2009, 15, 3–26. [Google Scholar] [CrossRef]
- Faliński, J.B.; Adamowski, W.; Lapshina, E.; Luczaj, L.; Venanzoni, R. Vegetation of the Kudrjašovskij Bor. In South Western Siberian Taiga Project. Report of Geobotanical Researches 2 Phytocoenosis, N.S.: Archivum Geobotanicum 2; Faliński, J.B., Pedrotti, F., Eds.; Bialowieska Stacja GeobotanicznaUW: Bialowieza, Poland, 1995; Volume 3, pp. 19–24. [Google Scholar]
- Anenkhonov, O.; Chytrý, M. Syntaxonomy of vegetation of the Svyatoi nos Peninsula, lake Baikal. 2. Forests and krummholz in comparison with other regions of northern Buratia. Folia Geobot. 1998, 33, 31–75. [Google Scholar] [CrossRef]
- Ermakov, N.; Cherosov, M.; Gogoleva, P. Classification of ultracontinental boreal forests in Central Yakutia. Folia Geobot. 2002, 37, 419–440. [Google Scholar] [CrossRef]
- Ermakov, N.; Cherosov, M. Differentiation of the Vaccinio-Piceetea and Loiseleurio-Vaccinietea in mountains of Yakutia. Ann. Bot. Nuova Ser. 2005, 5, 15–27. [Google Scholar]
- Polyakova, M.A.; Ermakov, N.B. Classification of pine forests on sandy deposits in the Minusinskaya intermountain basin (Southern Siberia). Veg. Russ. 2008, 13, 82–105. [Google Scholar] [CrossRef]
- Makhatkov, I.D.; Ermakov, N.B. Association of oligotrophic pine forest (Pineta sibiricae-silvestris) subzones of the northern taiga of the West Siberian Plain. Bull. Novosib. State Univ. Ser. Biol. Clin. Med. 2010, 8, 152–159. [Google Scholar]
- Freitag, M.; Mathar, W.; Yurtaev, A.; Hölzel, N. Floristic composition and environmental determinants of pine forests in the hemiboreal zone of Western Siberia. Tuexenia 2015, 35, 31–51. [Google Scholar] [CrossRef]
- Lashchinsky, N.N.; Korolyuk, A.Y.; Lashchinskaya, N.V. Pyrolo-Pinetea and Vaccinio-Piceetea classes in the Burla ribbon pine forests vegetation cover. Plant Life Asian Russ. 2017, 26, 63–84. [Google Scholar] [CrossRef]
- Lashchinsky, N.N.; Korolyuk, A.Y.; Tischenko, M.P.; Lashchinskaya, N.V. Syntaxonomy and spatial structure of the Burla ribbon pine forests. Plant Life Asian Russ. 2018, 29, 57–81. [Google Scholar] [CrossRef]
- Solomeshch, A.I.; Grigoriev, I.N.; Alimbekova, L.M. Syntaxonomy of the Southern Ural forests. VI. Coniferous forests. In Dep. VINITI 11.12.92. No. 3494-B 92; Redkollegiya zhurnala “Biologicheskiye nauki”: Ufa, Russia, 1992. [Google Scholar]
- Martynenko, V.B. Syntaxonomy of native forests of the South Ural region and their mountain analogues. In Diversity and Dynamics of Forest Ecosystems of Russia; Isaev, A.S., Ed.; KMK: Moscow, Russia, 2013; Volume 2, pp. 67–129. [Google Scholar]
- Shirokikh, P.S.; Baisheva, E.Z.; Martynenko, V.B.; Muldashev, A.A.; Alibaev, F.H.; Gorichev, Y.P.; Davydychev, A.N. Flora and Vegetation of the South Ural State Nature Reserve; Mirkin, B.M., Ed.; Gilem: Ufa, Russia, 2008; 528p. [Google Scholar]
- Libbert, W. Die Vegetationseinheiten der Neumärkischen Staubeckenlandschaft; Verhandlungen des Botanischen Vereins der Provinz Brandenburg: Brandenburg, Germany, 1933; pp. 229–348. [Google Scholar]
- Matuszkiewicz, W.; Matuszkiewicz, J. Phytosociological overview of forest communities in Poland. Vol. 2. Pine forests. Phytocoenosis. Biul. Fitosocjologiczny 1973, 2, 273–356. [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas Mit den Alpen, 4th ed.; Ulmer: Stuttgart, Germany, 1986. [Google Scholar]
- Borhidi, A. Social behaviour types, their naturalness and relative ecological indicator values of the higher plants of the Hungarian flora. Acta Bot. Hung. 1995, 39, 97–182. [Google Scholar]
- Kielland-Lund, J. Die Waldgesellschaften SO Norwegens. Phytocoenologia 1981, 9, 53–250. [Google Scholar] [CrossRef]
- Dierßen, K. Vegetation Nordeuropas; Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Kadilnikova, E.I.; Tsvetaev, A.A.; Smirnova, E.S. Physico-Geographical Zoning of the Bashkir ASSR; Kadilnikov, I.P., Ed.; Polygrafcombinat: Ufa, Russia, 1964; 210p. [Google Scholar]
- Khalevitskaya, G.S.; Babkina, M.I.; Kuznetsova, V.V. Agroclimatic Resources of the Bashkir ASSR; Gidrometeoizdat: Leningrad, Russia, 1976; 235p. [Google Scholar]
- Yaparov, I.M. (Ed.) Atlas of the Republic of Bashkortostan; Bashkortostan: Ufa, Russia, 2005. [Google Scholar]
- Gorchakovskiy, P.L. Flora of the High Mountainous Urals; Nauka: Moskow, Russia, 1975; 285p. [Google Scholar]
- Krasheninnikov, I.M.; Kucherovskaya-Rozhanets, S. Natural Resources of the Bashkir SSR; V.1. Izdatel’stvo AN SSSR: Moscow/Leningrad, Russia, 1941; 156p. [Google Scholar]
- Il’in, A. On the relict flora of the Southern Urals. Izv. Gl. Bot. Sada 1922, 21, 1–11. [Google Scholar]
- Krasheninnikov, I.M. On the History of the Landscape Development of the Southern Urals; A.B.S.S.R. Bashkirskij nar. kom. zem.: Leningrad, Russia, 1927; 28p. [Google Scholar]
- Kleopov, Y.D. Analysis of the Flora of Deciduous Forests of the European Part of USSR; Naukova Dumka: Kiev, Ukraine, 1990; 350p. [Google Scholar]
- Gorchakovskiy, P.L. The main problems of historical phytogeography of Ural. In Trudy Instituta Ekologii Rasteniy i Zhivotnykh Ural’Skogo Filiala AN SSSR; Redaktsionno-izdatel'skiy sovet Ural'skogo filiala AN SSSR: Sverdlovsk, Russia, 1969; Volume 66, 286p. [Google Scholar]
- Makovskiy, V.I.; Panova, N.K. Formation of vegetation of the upper mountain belt of the Southern Urals in the Holocene. Tr. Instituta Ekol. Rasteniy Zhivotnykh 1977, 105, 3–17. [Google Scholar]
- Popov, G.V. Geography and ecology of some rare flora species of Bashkiria. In Problems of Complex Study, Development and Protection of Landscapes of the Urals; Maksyutov, F.A., Ed.; B.i.: Ufa, Russia, 1980; 159p. [Google Scholar]
- Andreeva, N.M.; Bartalev, S.A.; Basova, E.V.; Bugaeva, K.S.; Butusov, O.B.; Golovanov, A.S.; Gromtsev, A.N.; Danilina, D.M.; Drobushevskaya, O.V.; Ershov, D.V.; et al. Diversity and Dynamics of Russian Forest Ecosystems; Isaev, A.S., Ed.; Tovarishchestvo nauchnykh izdaniy KMK: Moscow, Russia, 2013; 477p. [Google Scholar]
- Clements, F.E. Plant Succession and Indicators; Hafner Press: New York, NY, USA, 1973. [Google Scholar]
- Reymers, N.F. Nature Management; Mysl’: Moskva, Russia, 1980. [Google Scholar]
- Mart’yanov, N.A.; Batalov, A.A.; Kulagin, A.Y. Broad-Coniferous Forests of the Ufa Plateau: Phytocenotic Characteristics and Renewal; Gilem: Ufa, Russia, 2002; 222p. [Google Scholar]
- Shirokikh, P.S.; Kunafin, A.M.; Martynenko, V.B. Syntaxonomy of mid-successional Stages of secondary forests of the central elevated part of the Southern Urals. Veg. Russ. 2012, 20, 109–134. [Google Scholar] [CrossRef]
- Shumilova, L.V. Botanical Geography of Siberia; TSU: Tomsk, Russia, 1962. [Google Scholar]
- Khalevitskaya, G.S. Main climatic features. In Agroclimatic Resources of the Kurgan Oblast; Gidrometeoizdat: Leningrad, Russia, 1977. [Google Scholar]
- Naumenko, H.I. Flora and Vegetation of the Southern Trans-Urals; Izdatel’stvo KSU: Kurgan, Russia, 2008; 511p. [Google Scholar]
- Martynenko, V.; Shirokikh, P.; Solomeshch, A.; Muldashev, A. Vegetation Database Forest of Southern Ural. Biodivers. Ecol. 2012, 4, 289. [Google Scholar] [CrossRef]
- Hennekens, S.M.; Schamineé, J.H.J. TURBOVEG, a comprehensive data base management system for vegetation data. J. Veg. Sci. 2001, 12, 589–591. [Google Scholar] [CrossRef]
- Westhoff, V.; van der Maarel, E. The Braun-Blanquet approach. In Classification of Plant Communities; Whittaker, R.H., Ed.; Dr. W. Junk Publishers: Hague, The Netherlands, 1978. [Google Scholar]
- Mirkin, B.M.; Solomeshch, A.I.; Zhuravleva, S.E. Vegetation of Russia in the area of the syntaxonomy of Braun-Blanquet: Development and results. Zhurnal Obs. Biol. 2000, 1, 5–21. [Google Scholar]
- Tichý, L.; Holt, J.; Nejezchlebová, M. JUICE Program for Management, Analysis and Classification of Ecological Data, 2nd ed.; Masaryk University: Brno, Czech Republic, 2011. [Google Scholar]
- Lakari, O.J. Studies on forest types in northern Finland (Referat: Investigations on forest types in Northern Finland). Acta For. Fenn. 1920, 14, 1–85. [Google Scholar]
- Kielland-Lund, J. Zur Systematik der Kiefernwälder Fennoscandiens. Mitteilungen der Floristisch-soziologischen Arbeitsgemeinschaft. Mitteilungen FlorSoz Heft 1967, 11–12, 127–141. [Google Scholar]
- Matuszkiewicz, W. Zur Systematik der natürlichen Kiefernwälder des mittel- und osteuropäischen Flachlands. Mitteilungen Florist.-Soziologischen Arbeitsgemeinschaft 1962, 9, 145–186. [Google Scholar]
- Matuszkiewicz, J.M.; Solon, J.; Orzechowski, M.; Kozłowska, A.; Różański, W.; Szczygielski, M.; Mateiczuk, G.; Lorens, B.; Kowalska, A. Geobotanical Observe of Trends in the Development of Forest Communities in Some Regions of Poland; PAN IGiPZ: Warszawa, Poland, 2007. [Google Scholar]
- Oksanen, J.; Ahti, T. Lichen-rich pine forest vegetation in Finland. Ann. Bot. Fenn. 1982, 19, 275–301. [Google Scholar]
- Chytrý, M.; Douda, J.; Roleček, J.; Sádlo, J.; Boublík, K.; Hédl, R.; Vítková, M.; Zelený, D.; Navrátilová, J.; Hrivnák, R. Vegetation of Czech Republic 4. Forest and Scrub Vegetation; Academia: Praha, Czech Republic, 2013. [Google Scholar]
- Sokolov, S.Y. Types of forests in the eastern part of the Bakovo-Varnavino educational forestry enterprise. In Nature and Economy of the Training Forestry Farms of the Forestry Technical Academy; Gossel’khozgiz: Moscow/Leningrad, Russia, 1931; pp. 115–251. [Google Scholar]
- Saburov, D.N. Pinega Forests; Nauka: Leningrad, Russia, 1972; 173p. [Google Scholar]
- Morozova, O.V.; Korotkov, V.N. Classification of forest vegetation of Kostomukshsky reserve. Zapovednoe Delo 1999, 5, 56–78. [Google Scholar]
- Fedorchuk, V.N.; Neshataev, V.Y.; Kuznetsova, M.L. Forest Ecosystems of North-Western Regions of Russia: Typology, Dynamics, Forest Management Features; Nauka: St-Petersburg, Russia, 2005; 382p. [Google Scholar]
- Ermakov, N.; Makhatkov, I. Classification and ordination of north boreal light-coniferous forests of the West Siberian Plain. Plant Biosyst. 2011, 145, 199–207. [Google Scholar] [CrossRef]
- Hill, T.; Lewicki, P. Statistics Methods and Applications; StatSoft: Tulsa, OK, USA, 2007. [Google Scholar]
- Theurillat, J.-P.; Willner, W.; Fernández-González, F.; Bueltmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. 4th edition. Appl. Veg. Sci. 2020, 24, e12491. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- The Plant List. Available online: http://theplantlist.com/1/ (accessed on 3 July 2024).
- Plantarium. Plants and Lichens of Russia and Neighboring Countries: Open Online Galleries and Plant Identification Guide. Available online: https://www.plantarium.ru/lang/en.html (accessed on 3 July 2024).
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Urbanavichus, G.P. A Checklist of the Lichen Flora of Russia; Nauka: St-Petersburg, Russia, 2010; 198p. [Google Scholar]
- Heinken, T. Vaccinio-Piceetea (H7). Beerstrauch-Nadelwälder. Teil 1: Dicrano-Pinion. Sand- und Silikat-Kiefernwälder. In Synopsis Pflanzenges. Dtschl.; Dierschke, H., Ed.; Druckerei Goltze GmbH & Co. KG: Göttingen, Germany, 2008; Volume 10, 88p. [Google Scholar]
- Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Commun. 1992, 206, 1–9.
- Kolbek, J.; Chytrý, M.; Kučera, T.; Kočí, M.; Grulich, V.; Lustyk Suché, P. Bory–Dry Pine Forests. In Katalog Biotopu Cēské Republiky; Chytry, M., Kucera, T., Koci, M., Grulich, V., Lustyk, P., Eds.; Agentura Ochrany Prirody a Krajiny CR: Praga, Czech Republic, 2010; pp. 331–334. [Google Scholar]
- Wegrzyn, M.H.; Kolodziejczyk, J.; Falowska, P.; Wezyk, P.; Zieba-Kulawik, K.; Szostak, M.; Turowska, A.; Grzesiak, B.; Wietrzyk-Pelka, P. Influence of the environmental factors on the species composition of lichen Scots pine forests as a guide to maintain the community (Bory Tucholskie National Park, Poland). Glob. Ecol. Conserv. 2020, 22, e01017. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Macdonald, S.E.; Hogg, E.H. Ecology of and control strategies for Calamagrostis canadensis in boreal forest sites. Can. J. For. Res. 1993, 23, 2070–2077. [Google Scholar] [CrossRef]
- Ulanova, N.G.; Belova, I.N.; Logofet, D.O. On the competition among discrete-structured populations: A matrix model for population dynamics of woodreed and birch growing together. Biol. Bull. Rev. 2008, 69, 478–494. [Google Scholar]
- Logofet, D.O.; Ulanova, N.G.; Klochkova, I.N.; Demidova, A.N. Structure and dynamics of a clonal plant population: Classical model results in a non-classic formulation. Ecol. Model. 2006, 192, 95–106. [Google Scholar] [CrossRef]
- Prach, K.; Pysek, P. How do species dominating in succession differ from others? J. Veg. Sci. 1999, 10, 383–392. [Google Scholar] [CrossRef]
- Prach, K.; Wade, P.M. Population characteristics of expansive perennial herbs. Preslia. Praha 1992, 64, 45–51. [Google Scholar]
- Pysek, P. Pattern of species dominance and factors affecting community composition in areas deforested due to air pollution. Vegetatio 1994, 112, 45–56. [Google Scholar] [CrossRef]
- Rebele, F.; Lehmann, C. Biological flora of Central Europe: Calamagrostis epigejos (L.) Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Korkonosova, L.I. To the question of the formation of reedgrass clearings in the European North. In Issues of Taiga Forestry in the European North; Korkonosova, L.I., Ed.; Nauka: Moscow, Russia, 1967; pp. 101–113. [Google Scholar]
- Obydennikov, V.I. Calamagrostis cuttings in the taiga zone of the USSR. Sci. Work. Mosc. For. Tech. Inst. 1976, 88, 38–42. [Google Scholar]
- Martynenko, V.B.; Shirokikh, P.S.; Mirkin, B.M.; Naumova, L.G. Syntaxonomic analysis of restorative succession after cutting down light coniferous forests of South Ural region. Biol. Bull. Rev. 2014, 75, 478–490. [Google Scholar]
- Smirnova, O.V. (Ed.) Forests of Eastern Europe: Holocene History and Modern Times. Book 2; Russian Academy of Sciences, Centre for Problems of Forest Ecology and Productivity: Moskow, Russia, 2004; 575p. [Google Scholar]
- Galetto, L.; Torres, C.; Martínez Pastur, G.J. Variable retention harvesting: Conceptual analysis according to different environmental ethics and forest valuation. Ecol. Process. 2019, 8, 40. [Google Scholar] [CrossRef]
- Kryshen, A.M.; Sinkevich, S.M.; Shorokhova, E.V. Variable retention forestry is targeted to preserve the temporal and spatial continuity of forest habitats and ecosystem functions. Plant Resour. 2020, 56, 195–201. [Google Scholar] [CrossRef]
- Gustafsson, L.; Hannerz, M.; Koivula, M.; Shorohova, E.; Vanha-Majamaa, I.; Weslien, J. Research on retention forestry in Northern Europe. Ecol. Process. 2020, 9, 3. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Lindberg, H.; Vanha-Majamaa, I.; Keto-Tokoi, P.; Punttila, P. Low-level retention forestry, certification, and biodiversity: Case Finland. Ecol. Process. 2019, 8, 47. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).