
Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest


Abstract
1. Introduction
2. Materials and Methods
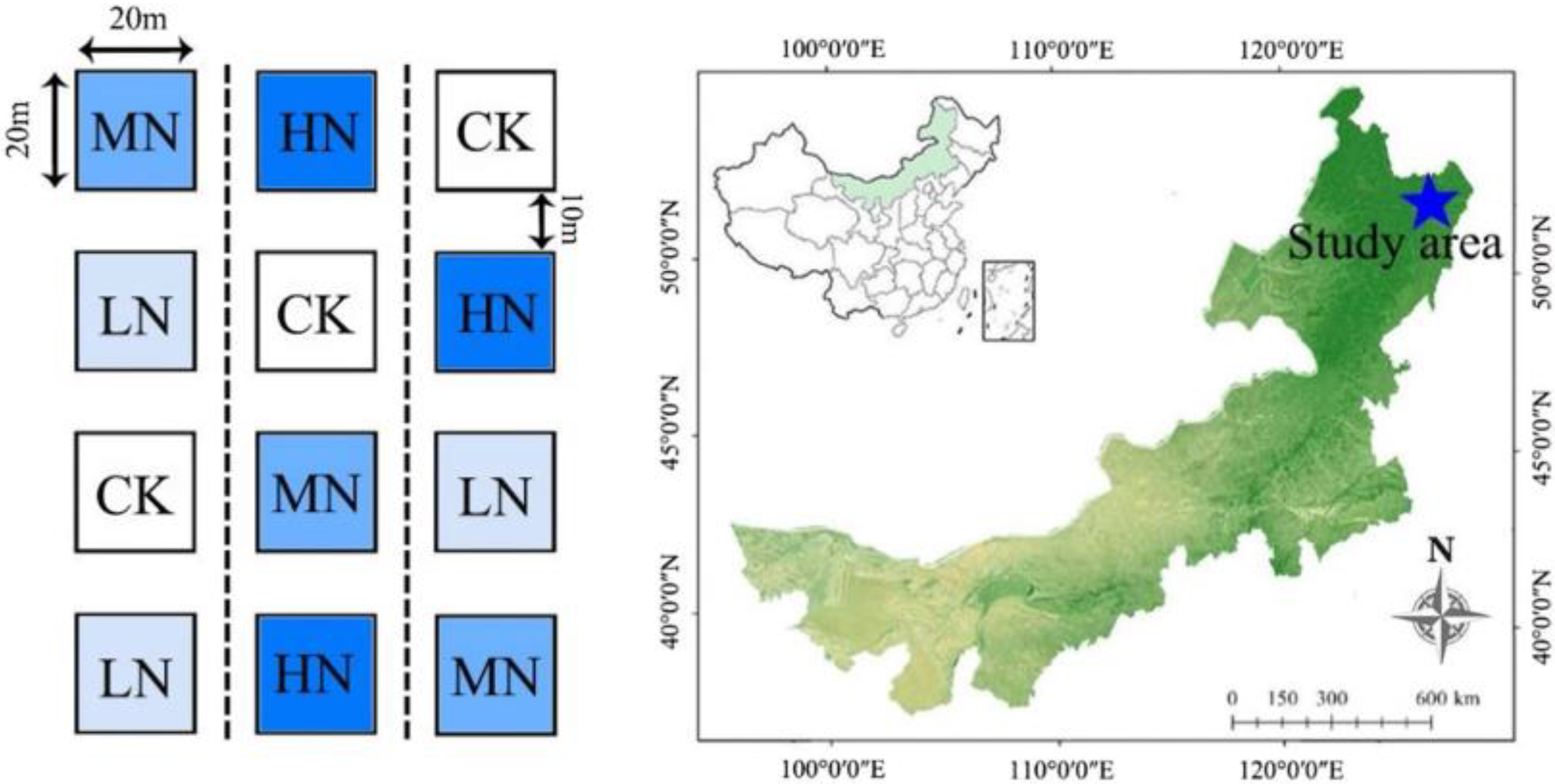
2.1. Site Description and Experimental Design
2.2. Sample Collection and Analysis
2.3. Data Analysis
3. Results
3.1. Soil and Litter Property
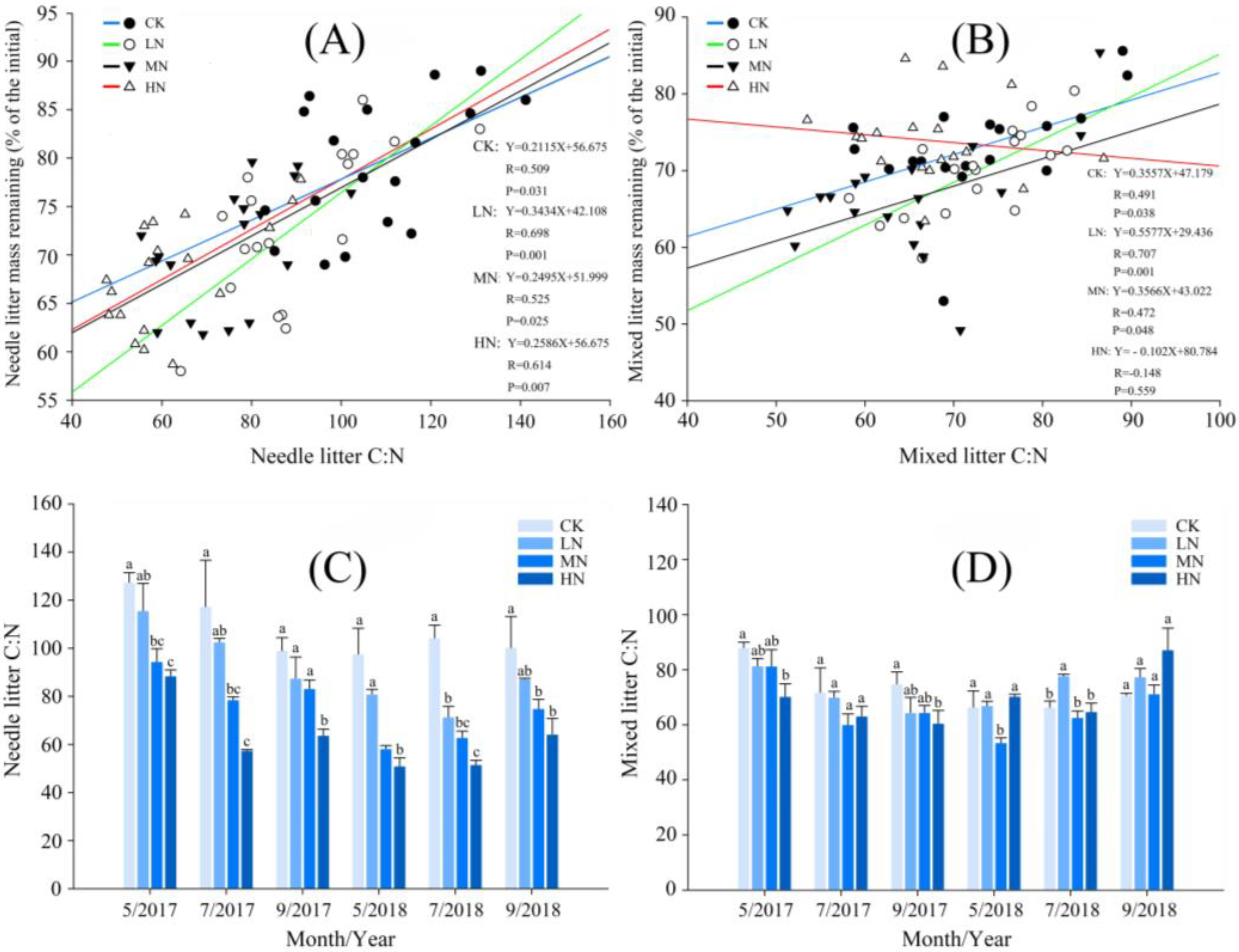
3.2. Effects of N Addition on Decomposition of Needle and Mixed Leaf Litter
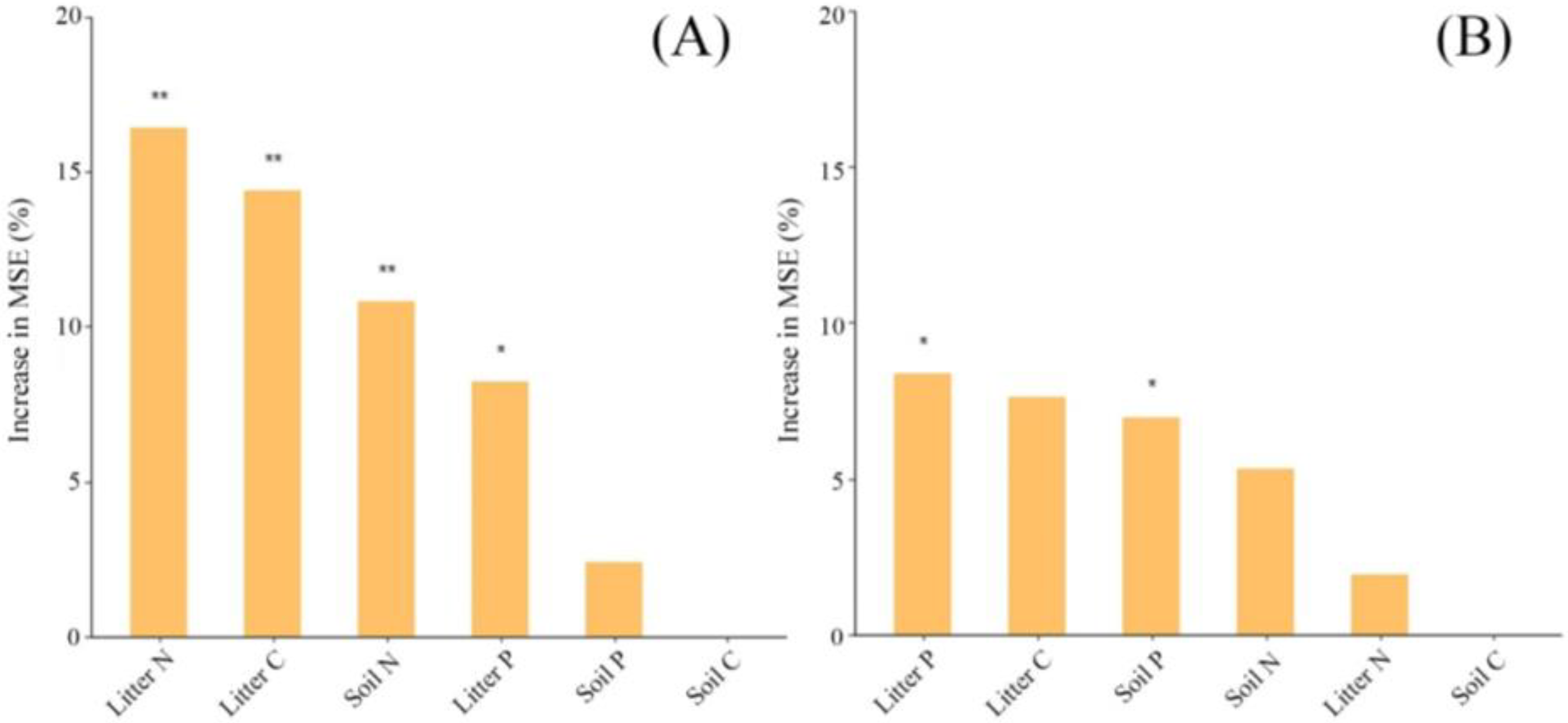
3.3. Relationship of Decomposition between Soil Animals and the Needle or Mixed Leaf Litter under N Addition
4. Discussion
4.1. Effect of N Addition on Decomposition Rate of Needle and Mixed Leaf Litter
4.2. Effects of N Addition and Small Soil Animals on Decomposition of Needle and Mixed Leaf Litter
4.3. Effects of N Addition on Nutrient Release from Needle and Mixed Leaf Litter
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sardans, J.; Alonso, R.; Janssens, I.A.; Carnicer, J.; Vereseglou, S.; Rillig, M.C.; Fernández-Martínez, M.; Sanders, T.G.M.; Peñuelas, J. Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: Relationships with climate, N deposition and tree growth. Funct. Ecol. 2016, 30, 676–689. [Google Scholar] [CrossRef]
- Du, C.; Guo, Q.; Zhang, J. A review on moss nitrogen and isotope signatures evidence for atmospheric nitrogen deposition. Sci. Total Environ. 2022, 806, 150765. [Google Scholar] [CrossRef] [PubMed]
- Buettel, J.C.; Ringwaldt, E.M.; Hovenden, M.J.; Brook, B.W. Importance of the local environment on nutrient cycling and litter decomposition in a tall eucalypt forest. Forests 2019, 10, 340. [Google Scholar] [CrossRef]
- Hernández, E.; Questad, E.J.; Meyer, W.M., III; Suding, K.N. The effects of nitrogen deposition and invasion on litter fuel quality and decomposition in a Stipa pulchra grassland. J. Arid. Environ. 2019, 162, 35–44. [Google Scholar] [CrossRef]
- Lukina, N.V.; Kuznetsova, A.I.; Geraskina, A.P.; Smirnov, V.E.; Ivanova, V.N.; Teben’kova, D.N.; Gornov, A.V.; Shevchenko, N.E.; Tikhonova, E.V. Unaccounted Factors Determining Carbon Stocks in Forest Soils. Russ. Meteorol. Hydrol. 2022, 47, 791–803. [Google Scholar] [CrossRef]
- Wood, T.E.; Lawrence, D.; Clark, D.A. Determinants of leaf litter nutrient cycling in a tropical rain forest: Soil fertility versus topography. Ecosystems 2006, 9, 700–710. [Google Scholar] [CrossRef]
- Chen, F.S.; Feng, X.; Liang, C. Endogenous versus exogenous nutrient affects C, N, and P dynamics in decomposing litters in mid-subtropical forests of China. Ecol. Res. 2012, 27, 923–932. [Google Scholar] [CrossRef]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Penner, J.F.; Frank, D.A. Litter Decomposition in Yellowstone Grasslands: The Roles of Large Herbivores, Litter Quality, and Climate. Ecosystems 2019, 22, 929–937. [Google Scholar] [CrossRef]
- Jabiol, J.; Lecerf, A.; Lamothe, S.; Gessner, M.O.; Chauvet, E. Litter Quality Modulates Effects of Dissolved Nitrogen on Leaf Decomposition by Stream Microbial Communities. Microb. Ecol. 2019, 77, 959–966. [Google Scholar] [CrossRef]
- Jiang, X.; Cao, L.; Zhang, R.; Yan, L.; Mao, Y.; Yang, Y. Effects of nitrogen addition and litter properties on litter decomposition and enzyme activities of individual fungi. Appl. Soil Ecol. 2014, 80, 108–115. [Google Scholar] [CrossRef]
- Zhang, W.; Chao, L.; Yang, Q.; Wang, Q.; Fang, Y.; Wang, S. Litter quality mediated nitrogen effect on plant litter decomposition regardless of soil fauna presence. Ecology 2016, 97, 2834–2843. [Google Scholar] [CrossRef]
- Almagro, M.; Maestre, F.T.; Martínez-López, J.; Valencia, E.; Rey, A. Climate change may reduce litter decomposition while enhancing the contribution of photodegradation in dry perennial Mediterranean grasslands. Soil Biol. Biochem. 2015, 90, 214–223. [Google Scholar] [CrossRef]
- Song, Y.; Song, C.; Ren, J.; Tan, W.; Jin, S.; Jiang, L. Influence of nitrogen additions on litter decomposition, nutrient dynamics, and enzymatic activity of two plant species in a peatland in Northeast China. Sci. Total Environ. 2018, 625, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315. [Google Scholar] [CrossRef]
- Gong, S.; Guo, R.; Zhang, T.; Guo, J. Warming and nitrogen addition increase litter decomposition in a temperate meadow ecosystem. PLoS ONE 2015, 10, e0116013. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Wang, G.G.; Fang, X.M.; Wan, S.Z.; Zhang, Y.; Liang, C. Nitrogen deposition effect on forest litter decomposition is interactively regulated by endogenous litter quality and exogenous resource supply. Plant Soil 2019, 437, 413–426. [Google Scholar] [CrossRef]
- Knorr, M.; Frey, S.D.; Curtis, P.S. Nitrogen additions and litter decomposition: A meta-analysis. Ecology 2005, 86, 3252–3257. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, H.L.; Chen, G.; Peng, Y.; Xiao, Y.L.; Hu, T.X.; Zhang, J.; Li, X.W.; Liu, L.; Tang, Y. Nitrogen Addition Significantly Affects Forest Litter Decomposition under High Levels of Ambient Nitrogen Deposition. PLoS ONE 2014, 9, e88752. [Google Scholar] [CrossRef]
- Hobbie, S.E. Nitrogen effects on decomposition: A five-year experiment in eight temperate sites. Ecology 2008, 89, 2633–2644. [Google Scholar] [CrossRef]
- Keiblinger, K.M.; Hall, E.K.; Wanek, W.; Szukics, U.; Hämmerle, I.; Ellersdorfer, G.; Böck, S.; Strauss, J.; Sterflinger, K.; Richter, A.; et al. The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency. FEMS Microbiol. Ecol. 2010, 73, 430–440. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Wang, J.; Bu, W.; Zhao, B.; Zhao, X.; Zhang, C.; Fan, J.; Gadow, K.V. Effects of Nitrogen Addition on Leaf Decomposition of Single-Species and Litter Mixture in Pinus tabulaeformis Forests. Forests 2015, 6, 4462–4476. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Partsch, S.; Parkinson, D.; Scheu, S. Invasion of a deciduous forest by earthworms: Changes in soil chemistry, microflora, microarthropods and vegetation. Soil Biol. Biochem. 2007, 39, 1099–1110. [Google Scholar] [CrossRef]
- Wong, M.K.; Tsukamoto, J.; Yusuyin, Y.; Tanaka, S.; Iwasaki, K.; Tan, N.P. Comparison of soil macro-invertebrate communities in Malaysian oil palm plantations with secondary forest from the viewpoint of litter decomposition. For. Ecol. Manag. 2016, 381, 63–73. [Google Scholar] [CrossRef]
- Yarwood, S.A. The role of wetland microorganisms in plant-litter decomposition and soil organic matter formation: A critical review. FEMS Microbiol. Ecol. 2018, 94, fiy175. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Holmstrup, M.; Schmidt, I.K.; Bachega, L.R.; Schelfhout, S.; Zheng, H.; Heděnec, P.; Yue, K.; Vesterdal, L. Tree species identity is the predominant modulator of the effects of soil fauna on leaf litter decomposition. For. Ecol. Manag. 2022, 520, 120396. [Google Scholar] [CrossRef]
- Xu, G.L.; Mo, J.M.; Fu, S.L.; Gundersen, P.E.R.; Zhou, G.Y.; Jing-Hua, X.U.E. Response of soil fauna to simulated nitrogen deposition: A nursery experiment in subtropical China. J. Environ. Sci. 2007, 19, 603–609. [Google Scholar] [CrossRef]
- Coulis, M.; Hättenschwiler, S.; Fromin, N.; David, J.F. Macroarthropod-microorganism interactions during the decomposition of Mediterranean shrub litter at different moisture levels. Soil Biol. Biochem. 2013, 64, 114–121. [Google Scholar] [CrossRef]
- Manning, P.; Saunders, M.; Bardgett, R.D.; Bonkowski, M.; Bradford, M.A.; Ellis, R.J.; Kandeler, E.; Marhan, S.; Tscherko, D. Direct and indirect effects of nitrogen deposition on litter decomposition. Soil Biol. Biochem. 2008, 40, 688–698. [Google Scholar] [CrossRef]
- Chapin, F.S.; Sala, O.E.; Huber-Sannwald, E. (Eds.) Global Biodiversity in a Changing Environment: Scenarios for the 21st Century (Vol. 152); Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459. [Google Scholar] [CrossRef]
- Yin, W. China Pictorial Keys to Soil Animals; Science Press: Beijing, China, 1998. [Google Scholar]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Eddy, W.C.; Buyarski, C.R.; Adair, E.C.; Ogdahl, M.L.; Weisenhorn, P. Response of decomposing litter and its microbial community to multiple forms of nitrogen enrichment. Ecol. Monogr. 2012, 82, 389–405. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, J.; Ding, W.; Bol, R.; Chen, Z.; Luo, J.; Bolan, N. Stage-specific response of litter decomposition to N and S amendments in a subtropical forest soil. Biol. Fertil. Soils 2016, 52, 711–724. [Google Scholar] [CrossRef]
- Su, Y.; Le, J.; Han, W.; Wang, C.; Li, K.; Liu, X. Long-term nitrogen addition consistently decreased litter decomposition rates in an alpine grassland. Plant Soil 2022, 479, 495–509. [Google Scholar] [CrossRef]
- Pérez-Suárez, M.; Arredondo-Moreno, J.T.; Huber-Sannwald, E. Early stage of single and mixed leaf-litter decomposition in semiarid forest pine-oak: The role of rainfall and microsite. Biogeochemistry 2012, 108, 245–258. [Google Scholar] [CrossRef]
- Britton, A.J.; Mitchell, R.J.; Fisher, J.M.; Riach, D.J.; Taylor, A.F. Nitrogen deposition drives loss of moss cover in alpine moss–sedge heath via lowered C: N ratio and accelerated decomposition. New Phytol. 2018, 218, 470–478. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bonner, K.I.; Nicholson, K.S. Biodiversity and plant litter: Experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos 1997, 79, 247–258. [Google Scholar] [CrossRef]
- Schimel, J.P.; Hättenschwiler, S. Nitrogen transfer between decomposing leaf of different N status. Soil Biol. Biochem. 2007, 39, 1428–1436. [Google Scholar] [CrossRef]
- Kaneko, N.; Salamanca, E. Mixed leaf litter effects on decomposition rates and soil microarthropod communities in an oak–pine stand in Japan. Ecol. Res. 1999, 14, 131–138. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Sinsabaugh, R.L.; Repert, D.A.; Parkhurst, D.F. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology 2000, 81, 2359–2365. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Zhang, W.; Shao, Y.; Duan, H.; Chen, B.; Wei, X.; Fan, H. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Fujii, S.; Mori, A.S.; Kominami, Y.; Tawa, Y.; Inagaki, Y.; Takanashi, S.; Takeda, H. Differential utilization of root-derived carbon among collembolan species. Pedobiologia 2016, 59, 225–227. [Google Scholar] [CrossRef]
- Phillips, H.R.; Bach, E.M.; Bartz, M.L.; Bennett, J.M.; Beugnon, R.; Briones, M.J.; Brown, G.G.; Ferlian, O.; Gongalsky, K.B.; Guerra, C.A.; et al. Global data on earthworm abundance, biomass, diversity and corresponding environmental properties. Sci. Data 2021, 8, 136. [Google Scholar] [CrossRef]
- Aupic-Samain, A.; Baldy, V.; Lecareux, C.; Fernandez, C.; Santonja, M. Tree litter identity and predator density control prey and predator demographic parameters in a Mediterranean litter-based multi-trophic system. Pedobiologia 2019, 73, 1–9. [Google Scholar] [CrossRef]
- Zheng, Z.; Mamuti, M.; Liu, H.; Shu, Y.; Hu, S.; Wang, X.; Li, B.; Lin, L.; Li, X. Effects of nutrient additions on litter decomposition regulated by phosphorus-induced changes in litter chemistry in a subtropical forest, China. For. Ecol. Manag. 2017, 400, 123–128. [Google Scholar] [CrossRef]
- Cheever, B.M.; Webster, J.R.; Bilger, E.E.; Thomas, S.A. The relative importance of exogenous and substrate-derived nitrogen for microbial growth during leaf decomposition. Ecology 2013, 94, 1614–1625. [Google Scholar] [CrossRef]
- Liu, G.; Xing, Y.; Wang, Q.; Wang, L.; Feng, Y.; Yin, Z.; Wang, X.; Liu, T. Long-term nitrogen addition regulates root nutrient capture and leaf nutrient resorption in Larix gmelinii in a boreal forest. Eur. J. For. Res. 2021, 140, 763–776. [Google Scholar] [CrossRef]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The global stoichiometry of litter nitrogen mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef] [PubMed]
- He, X.T.; Stevenson, F.J.; Mulvaney, R.L.; Kelley, K.R. Incorporation of newly immobilized 15N into stable organic forms in soil. Soil Biol. Biochem. 1988, 20, 75–81. [Google Scholar] [CrossRef]
- Kurokawa, H.; Peltzer, D.A.; Wardle, D.A. Plant traits, leaf palatability and litter decomposability for co-occurring woody. Funct. Ecol. 2010, 24, 513–523. [Google Scholar] [CrossRef]
- Sorensen, P.L.; Michelsen, A. Long-term warming and litter addition affects nitrogen fixation in a subarctic heath. Glob. Change Biol. 2011, 17, 528–537. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CK | LN | MN | HN |
|---|---|---|---|---|
| Needle litter C (g·kg−1) | 493.46 ± 32.83 a | 491.30 ± 16.44 a | 486.46 ± 35.78 a | 488.96 ± 13.29 a |
| Needle litter N (g·kg−1) | 3.22 ± 0.11 b | 3.24 ± 0.11 b | 3.35 ± 0.11 b | 3.71 ± 0.10 a |
| Needle litter P (g·kg−1) | 0.99 ± 0.08 a | 0.90 ± 0.05 ab | 0.83 ± 0.04 ab | 0.80 ± 0.01 b |
| Needle litter C:N | 153.43 ± 15.52 a | 151.54 ± 1.71 a | 145.29 ± 10.80 ab | 131.79 ± 0.23 b |
| Mixed leaf litter C(g·kg−1) | 515.11 ± 42.97 a | 528.92 ± 8.83 a | 532.08 ± 2.67 a | 524.52 ± 9.67 a |
| Mixed leaf litter N (g·kg−1) | 3.64 ± 0.05 c | 4.38 ± 0.03 b | 4.50 ± 0.04 b | 4.80 ± 0.09 a |
| Mixed leaf litter P (g·kg−1) | 1.14 ± 0.01 a | 1.11 ± 0.04 ab | 1.09 ± 0.05 ab | 1.03 ± 0.02 b |
| Mixed leaf litter C:N | 141.58 ± 10.22 a | 120.82 ± 1.25 b | 118.24 ± 0.70 b | 109.36 ± 3.74 b |
| Year | Treatment | T C (g kg−1) | T N (g kg−1) | T P (g kg−1) | C:N | C:P | N:P | pH |
|---|---|---|---|---|---|---|---|---|
| 2017 | CK | 43.24 ± 5.52 a | 1.76 ± 0.09 b | 0.71 ± 0.06 ab | 24.75 ± 4.17 a | 60.74 ± 2.79 | 2.51 ± 0.34 b | 5.48 ± 0.14 a |
| LN | 49.69 ± 3.47 a | 1.96 ± 0.19 ab | 0.79 ± 0.03 a | 25.77 ± 3.96 a | 62.67 ± 4.09 | 2.48 ± 0.33 b | 5.38 ± 0.13 a | |
| MN | 47.64 ± 5.23 a | 2.30 ± 0.09 a | 0.70 ± 0.01 ab | 20.67 ± 1.69 a | 67.70 ± 7.92 | 3.27 ± 0.14 ab | 5.23 ± 0.05 ab | |
| HN | 49.01 ± 2.79 a | 2.26 ± 0.07 a | 0.65 ± 0.03 b | 21.64 ± 0.72 a | 75.82 ± 6.44 | 3.50 ± 0.18 a | 5.01 ± 0.02 b | |
| 2018 | CK | 45.81 ± 6.89 a | 2.00 ± 0.32 c | 0.83 ± 0.14 a | 22.52 ± 1.08 a | 52.48 ± 2.6 a | 2.41 ± 0.03 b | 5.52 ± 0.12 a |
| LN | 45.25 ± 2.36 a | 2.09 ± 0.17 bc | 0.79 ± 0.19 a | 21.81 ± 0.36 a | 56.96 ± 4.74 a | 2.77 ± 0.31 b | 5.41 ± 0.23 ab | |
| MN | 45.80 ± 3.73 a | 2.54 ± 0.22 ab | 0.81 ± 0.17 a | 18.62 ± 0.31 b | 57.55 ± 5.00 a | 3.30 ± 0.17 a | 5.29 ± 0.06 ab | |
| HN | 48.61 ± 5.72 a | 2.60 ± 0.27 a | 0.79 ± 0.11 a | 18.36 ± 0.87 b | 62.29 ± 3.07 a | 3.32 ± 0.21 a | 4.98 ± 0.05 b |
| Treatments | a | Decomposition Coefficient (k) | Determinant Coefficients (R2) | Time Required to Decompose 50% | Time Required to Decompose 95% |
|---|---|---|---|---|---|
| Needle litter CK | 0.987 | 0.156 ± 0.010 | 0.979 | 4.44 | 19.20 |
| Needle litter LN | 0.967 | 0.190 ± 0.025 | 0.921 | 3.65 | 15.77 |
| Needle litter MN | 0.933 | 0.203 ± 0.029 | 0.908 | 3.41 | 14.76 |
| Needle litter HN | 0.915 | 0.228 ± 0.034 | 0.898 | 3.04 | 13.14 |
| Mixed leaf litter CK | 0.938 | 0.176 ± 0.033 | 0.852 | 3.93 | 17.02 |
| Mixed leaf litter LN | 0.925 | 0.181 ± 0.033 | 0.861 | 3.83 | 16.55 |
| Mixed leaf litter MN | 0.919 | 0.227 ± 0.042 | 0.853 | 3.05 | 13.20 |
| Mixed leaf litter HN | 0.935 | 0.178 ± 0.032 | 0.860 | 3.89 | 16.83 |
| Factors | F(p) Value | ||
|---|---|---|---|
| Time | Treatments | Time × Treatments | |
| Needle litter C | 271.704 (<0.001) | 126.478 (<0.001) | 5.808 (<0.001) |
| Needle litter N | 17.248 (<0.001) | 13.904 (<0.001) | 2.793 (<0.001) |
| Needle litter P | 10.612 (<0.001) | 21.119 (<0.001) | 8.396 (<0.001) |
| Mixed leaf litter C | 9.843 (<0.001) | 3.250 (<0.05) | 1.597 (>0.05) |
| Mixed leaf litter N | 12.396 (<0.001) | 11.158 (<0.001) | 2.949 (<0.05) |
| Mixed leaf litter P | 9.171 (<0.001) | 11.459 (<0.001) | 5.706 (<0.001) |
| Needle litter C:N | 25.949 (<0.001) | 90.288 (<0.001) | 1.505 (>0.05) |
| Mixed leaf litter C:N | 15.628 (<0.001) | 8.848 (<0.001) | 3.126 (<0.001) |
| Month | Treatment | Isotomidae | Onychiuridae | Entomobryidae | Hypogastruridae | Oribatida | Mesostigmata |
|---|---|---|---|---|---|---|---|
| 5 | CK | 35 ± 5.31 b | 6 ± 1.25 a | 20 ± 5.72 a | 13 ± 6.13 a | 42 ± 3.26 b | 9 ± 1.25 b |
| LN | 67 ± 13.47 a | 4 ± 1.25 a | 10 ± 2.45 ab | 10 ± 2.45 a | 57 ± 2.87 a | 25 ± 0.47 a | |
| MN | 21 ± 2.05 bc | 4 ± 1.63 a | 15 ± 4.90 ab | 14 ± 7.35 a | 33 ± 3.68 c | 8 ± 2.87 b | |
| HN | 10 ± 4.08 c | 5 ± 2.05 a | 6 ± 0.47 b | 17 ± 0.47 a | 32 ± 3.27 c | 9 ± 1.63 b | |
| 7 | CK | 105 ± 4.08 b | 12 ± 0.47 b | 13 ± 4.50 c | 9 ± 0.00 ab | 75 ± 5.72 b | 19 ± 3.67 a |
| LN | 201 ± 9.39 a | 26 ± 0.82 a | 61 ± 3.27 a | 16 ± 5.31 a | 174 ± 5.31 a | 12 ± 2.05 ab | |
| MN | 223 ± 18.78 a | 17 ± 2.45 b | 38 ± 8.98 b | 15 ± 0.00 a | 57 ± 10.61 ab | 7 ± 0.82 bc | |
| HN | 57 ± 3.27 c | 7 ± 0.82 c | 15 ± 0.47 c | 6 ± 0.82 b | 44 ± 8.98 c | 4 ± 0.00 c | |
| 9 | CK | 37 ± 6.53 a | 4 ± 0.00 a | 7 ± 1.63 b | 6 ± 2.05 a | 39 ± 15.11 a | 9 ± 0.47 a |
| LN | 17 ± 3.27 b | 5 ± 0.47 a | 12 ± 1.25 a | 5 ± 0.82 a | 19 ± 1.63 ab | 6 ± 2.05 a | |
| MN | 3 ± 0.82 c | 3 ± 0.00 a | 3 ± 0.00 c | 3 ± 0.82 a | 25 ± 11.8 ab | 6 ± 2.05 a | |
| HN | 12 ± 2.87 bc | 5 ± 1.63 a | 7 ± 0.00 b | 6 ± 1.63 a | 9 ± 0.81 b | 5 ± 1.63 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Liu, G.; Xing, Y.; Yan, G.; Wang, Q. Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest. Forests 2024, 15, 372. https://doi.org/10.3390/f15020372
Wang M, Liu G, Xing Y, Yan G, Wang Q. Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest. Forests. 2024; 15(2):372. https://doi.org/10.3390/f15020372
Chicago/Turabian StyleWang, Miao, Guancheng Liu, Yajuan Xing, Guoyong Yan, and Qinggui Wang. 2024. "Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest" Forests 15, no. 2: 372. https://doi.org/10.3390/f15020372
APA StyleWang, M., Liu, G., Xing, Y., Yan, G., & Wang, Q. (2024). Long-Term Nitrogen Addition Accelerates Litter Decomposition in a Larix gmelinii Forest. Forests, 15(2), 372. https://doi.org/10.3390/f15020372