
Botryosphaeriaceae Species Associated with Stem Canker, Shoot Blight and Dieback of Fraxinus ornus in Italy

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Surveys and Sampling
2.2. Fungal Isolations
2.3. Morphological Identification
2.4. DNA-Based Identification
2.5. Pathogenicity Tests
2.6. Statistical Analyses
3. Results
3.1. Field Surveys
3.2. Isolate Identification
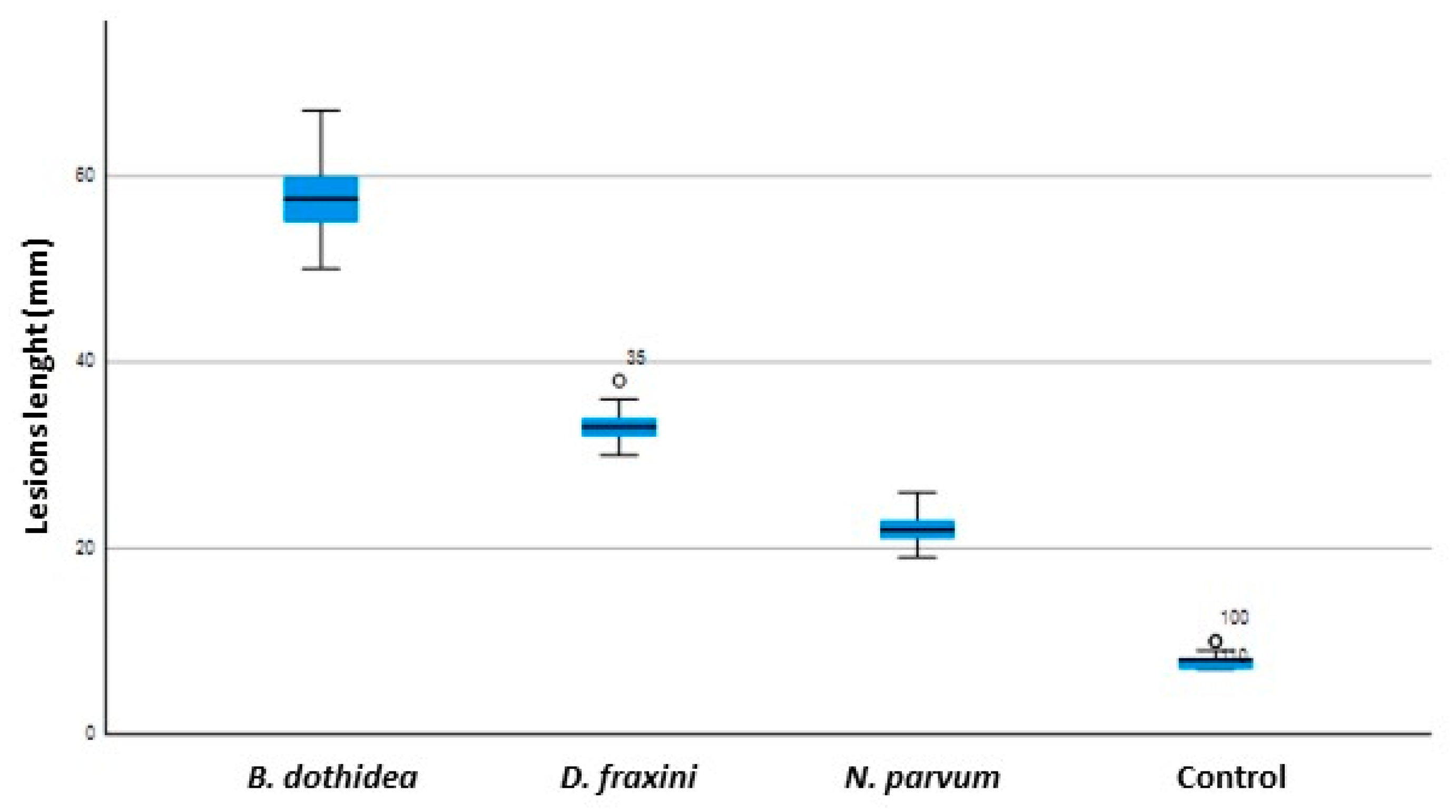
3.3. Pathogenicity Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caudullo, W.; de Rigo, D. Fraxinus ornus in Europe: Distribution, habitat, usage and threats. In The European Atlas of Forest Tree Species: Modelling, Data and Information on Forest Tree Species; De Rigo, D., Caudullo, G., Houston-Durrant, T., San-Miguel-Ayanz, J., Eds.; Publishing Office of the European Union: Luxemburg, 2016; pp. 100–101. [Google Scholar]
- Boshier, D. Ash Species in Europe: Biological Characteristics and Practical Guidelines for Sustainable Use; Oxford Forestry Institute: Oxford, UK, 2005; ISBN 978-0-85074-163-6. [Google Scholar]
- Norris, J.E.; Di Iorio, A.; Stokes, A.; Nicoll, B.C.; Achim, A. Slope Stability and Erosion Control: Ecotechnological Solutions; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; pp. 167–210. [Google Scholar]
- Luchi, N.; Ghelardini, L.; Santini, A.; Migliorini, D.; Capretti, P. First record of ash dieback caused by Hymenoschyphus fraxineus on Fraxinus excelsior in the Apennines (Tuscany, Italy). Plant Dis. 2016, 100, 535. [Google Scholar] [CrossRef]
- Ogris, N.; Hauptman, T.; Jurc, D.; Floreancig, V.; Marsich, F.; Montecchio, L. First report of Chalara fraxinea on common ash in Italy. Plant Dis. 2010, 94, 133. [Google Scholar] [CrossRef]
- Rigling, D.; Hilfiker, S.; Schöbel, C.; Meier, F.; Engesser, R.; Scheidegger, C.; Queloz, V. Il Deperimento del Frassino. Biologia, Sintomi e Raccomandazioni per la Gestione; Notizie per la Pratica 57; Istituto Federale di Ricerca: Birmensdorf, Switzerland, 2018; WSL.; 8p. [Google Scholar]
- Migliorini, D.; Luchi, N.; Nigrone, E.; Pecori, F.; Pepori, A.; Santini, A. Expansion of Ash Dieback towards the scattered Fraxinus excelsior range of the Italian peninsula. Biol. Invasions 2022, 24, 1359–1373. [Google Scholar] [CrossRef]
- Lygis, V.; Vasiliauskas, R.; Larsson, K.-H.; Stenlid, J. Wood-inhabiting fungi in stems of Fraxinus excelsior in declining ash stands of northern Lithuania, with particular reference to Armillaria cepistipes. Scand. J. For. Res. 2005, 20, 337–346. [Google Scholar] [CrossRef]
- Milenković, I.; Keča, N.; Karadžić, D.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Tkaczyk, M. Interaction between Hymenoscyphus fraxineus and Phytophthora species on young Fraxinus excelsior seedlings. For. Chron. 2018, 94, 135–139. [Google Scholar] [CrossRef]
- Peters, S.; Fuchs, S.; Bien, S.; Bußkamp, J.; Langer, G.J.; Langer, E.J. Fungi associated with stem collar necroses of Fraxinus excelsior affected by ash dieback. Mycol. Prog. 2023, 22, 52. [Google Scholar] [CrossRef]
- Przybył, K. Fungi associated with necrotic apical parts of Fraxinus excelsior shoots. For. Pathol. 2002, 32, 387–394. [Google Scholar] [CrossRef]
- Orlikowski, L.B.; Ptaszek, M.; Rodziewicz, A.; Nechwatal, J.; Thinggaard, K.; Jung, T. Phytophthora root and collar rot of mature Fraxinus excelsior in forest stands in Poland and Denmark. For. Pathol. 2011, 41, 510–519. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Thomsen, I.M.; Skovgaard, I.M.; Martinussen, T. Associations among symptoms of dieback in even-aged stands of ash (Fraxinus excelsior L.). For. Pathol. 2010, 40, 7–18. [Google Scholar] [CrossRef]
- Nunes, L.J.; Meireles, C.I.; Gomes, C.J.P.; Ribeiro, N.; Almeida, M.C. The Impact of Climate Change on Forest Development: A Sustainable Approach to Management Models Applied to Mediterranean-Type Climate Regions. Plants 2022, 11, 69. [Google Scholar] [CrossRef]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Serrano, M.S.; Romero, M.A.; Homet, P.; Gómez-Aparicio, L. Climate change impact on the population dynamics of exotic pathogens: The case of the worldwide pathogen Phytophthora cinnamomi. Agric. Meteorol. 2022, 322, 109002. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Ulbrich, U.; Lionello, P.; Belusic, D.; Jacobeit, J.; Knippertz, P.; Kuglitsch, F.G.; Leckebusch, G.C.; Luterbacher, J.; Maugeri, M.; Maheras, P.; et al. Climate of the Mediterranean: Synoptic patterns, temperature, precipitation, winds and their extremes. In Climate of the Mediterranean Region—From the Past to the Future; Elsevier: Sydney, Australia, 2012; pp. 301–346. [Google Scholar]
- Valdes-Abellan, J.; Pardo, M.A.; Tenza-Abril, A.J. Observed precipitation trend changes in the western Mediterranean region. Int. J. Climatol. 2017, 37, 1285–1296. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Pillay, K.; Slippers, B.; Wingfield, M.J.; Gryzenhout, M. Diversity and distribution of co-infecting Botryosphaeriaceae from Eucalyptus grandis and Syzygium cordatum in South Africa. S. Afr. J. Bot. 2013, 84, 38–43. [Google Scholar] [CrossRef]
- Xue, D.S.; Liu, J.; Li, B.H.; Xu, X.M.; Liu, N.; Lian, S.; Dong, X.L.; Wang, C.X. Effect of rainfall and temperature on perithecium production of Botryosphaeria dothidea on cankered apple branches. Phytopathology 2021, 111, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Moricca, S.; Uccello, A.; Turco, E.; Ginetti, B.; Ragazzi, A. Multiple Botryosphaeriaceae infection in forest trees: Synergistic or antagonistic interaction? J. Plant Pathol. 2010, 92, 91. [Google Scholar]
- Moral, J.; Muñoz-Díez, C.; González, N.; Trapero, A.; Michailides, T.J. Characterization and pathogenicity of Botryosphaeriaceae species collected from olive and other hosts in Spain and California. Phytopathology 2010, 100, 1340–1351. [Google Scholar] [CrossRef]
- Manca, D.; Bregant, C.; Maddau, L.; Pinna, C.; Montecchio, L.; Linaldeddu, B.T. First report of canker and dieback caused by Neofusicoccum parvum and Diplodia olivarum on oleaster in Italy. Ital. J. Mycol. 2020, 49, 85–91. [Google Scholar]
- Batista, E.; Lopes, A.; Alves, A. Botryosphaeriaceae species on forest trees in Portugal: Diversity, distribution and pathogenicity. Eur. J. Plant Pathol. 2020, 158, 693–720. [Google Scholar] [CrossRef]
- Benigno, A.; Bregant, C.; Aglietti, C.; Rossetto, G.; Tolio, B.; Moricca, S.; Linaldeddu, B.T. Pathogenic fungi and oomycetes causing dieback on Fraxinus species in the Mediterranean climate change hotspot region. Front. For. Glob. Chang. 2023, 6, 1253022. [Google Scholar] [CrossRef]
- Smith, H.; Wingfield, M.J.; Crous, P.W.; Coutinho, T.A. Sphaeropsis sapinea and Botryosphaeria dothidea endophytic in Pinus spp. and Eucalyptus spp. in South Africa. S. Afr. J. Bot. 1996, 62, 86–88. [Google Scholar] [CrossRef]
- Fiorenza, A.; Gusella, G.; Vecchio, L.; Aiello, D.; Polizzi, G. Diversity of Botryosphaeriaceae species associated with canker and dieback of avocado (Persea americana) in Italy. Phytopathol. Mediterr. 2023, 62, 47–63. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA 502 Genes for Phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Moricca, S.; Bracalini, M.; Benigno, A.; Ginetti, B.; Pelleri, F.; Panzavolta, T. Thousand Cankers Disease caused by Geosmithia morbida and its insect vector Pityophthorus juglandis first reported on Juglans nigra in Tuscany, Central Italy. Plant Dis. 2019, 103, 369. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Moricca, S.; Uccello, A.; Ginetti, B.; Ragazzi, A. First report of Neofusicoccum parvum associated with bark canker and dieback of Acer pseudoplatanus and Quercus robur in Italy. Plant Dis. 2012, 96, 1699. [Google Scholar] [CrossRef] [PubMed]
- Smahi, H.; Belhoucine-Guezouli, L.; Berraf-Tebbal, A.; Chouih, S.; Arkam, M.; Franceschini, A.; Linaldeddu, B.T.; Phillips, A.J.L. Molecular characterization and pathogenicity of Diplodia corticola and other Botryosphaeriaceae species associated with canker and dieback of Quercus suber in Algeria. Mycosphere 2017, 8, 1261–1272. [Google Scholar] [CrossRef]
- Cimmino, A.; Bahmani, Z.; Masi, M.; Di Lecce, R.; Amini, J.; Abdollahzadeh, J.; Tuzi, A.; Evidente, A. Massarilactones D and H, phytotoxins produced by Kalmusia variispora, associated with grapevine trunk diseases (GTDs) in Iran. Nat. Prod. Res. 2020, 35, 5192–5198. [Google Scholar] [CrossRef] [PubMed]
- Manetti, G.; Brunetti, A.; Lumia, V.; Sciarroni, L.; Marangi, P.; Cristella, N.; Faggioli, F.; Reverberi, M.; Scortichini, M.; Pilotti, M. Identification and Characterization of Neofusicoccum stellenboschiana in Branch and Twig Dieback-Affected Olive Trees in Italy and Comparative Pathogenicity with N. mediterraneum. J. Fungi 2023, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.R. The status of Botryosphaeriaceae species infecting grapevines. Phytopathol. Mediterr. 2011, 50, 5–45. [Google Scholar]
- van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef]
- Chapman, D.; Purse, B.V.; Roy, H.E.; Bullock, J.M. Global trade networks determine the distribution of invasive non-native species. Glob. Ecol. Biogeogr. 2017, 26, 907–917. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A. Fungal endophytes in Mediterranean oak forests: A lesson from Discula quercina. Phytopathology 2008, 98, 380–386. [Google Scholar] [CrossRef]
- Stavi, I.; Thevs, N.; Welp, M.; Zdruli, P. Provisioning Ecosystem Services Related with Oak (Quercus) Systems: A Review of Challenges and Opportunities. Agrofor. Syst. 2022, 96, 293–313. [Google Scholar] [CrossRef]
- Brown, E.A.; Hendrix, F.F. Pathogenicity and histopathology of Botryosphaeria dothidea on apple stems. Phytopathology 1981, 71, 375–379. [Google Scholar] [CrossRef]
- Ma, Z.; Boehm, E.W.; Luo, Y.; Michailides, T.J. Population structure of Botryosphaeria dothidea from pistachio and other hosts in California. Phytopathology 2001, 91, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Morgan, D.P.; Michailides, T.J. Effects of water stress on Botryosphaeria blight of pistachio caused by Botryosphaeria dothidea. Plant Dis. 2001, 85, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Dakin, N.; White, D.; Hardy, G.E.S.J.; Burgess, T.I. The opportunistic pathogen, Neofusicoccum australe, is responsible for crown dieback of peppermint (Agonis flexuosa) in Western Australia. Australas. Plant Pathol. 2010, 39, 202–206. [Google Scholar] [CrossRef]
- Piskur, B.; Pavlic, D.; Slippers, B.; Ogris, N.; Maresi, G.; Wingfield, M.J.; Jurc, J. Diversity and pathogenicity of Botryosphaeriaceae on declining Ostrya carpinifolia in Slovenia and Italy following extreme weather conditions. Eur. J. Forest Res. 2011, 130, 235–249. [Google Scholar] [CrossRef][Green Version]
- Van Der Linde, J.A.; Six, D.L.; Wingfield, M.J.; Roux, J. Lasiodiplodia species associated with dying Euphorbia ingens in South Africa. South. For. 2011, 73, 165–173. [Google Scholar] [CrossRef]
- Van Der Linde, J.A.; Roux, J.; Wingfield, M.J.; Six, D.L. Die-off of giant Euphorbia trees in South Africa: Symptoms and relationships to climate. S. Afr. J. Bot. 2012, 83, 172–185. [Google Scholar] [CrossRef]
- Elad, Y.; Pertot, I. Climate change impacts on plant pathogens and plant diseases. J. Crop Improv. 2014, 28, 99–139. [Google Scholar] [CrossRef]
- Tubby, K.V.; Webber, J.F. Pests and diseases threatening urban trees under a changing climate. Forestry 2010, 83, 451–459. [Google Scholar] [CrossRef]
- Hunjan, M.S.; Lore, J.S. Climate change: Impact on plant pathogens, diseases, and their management. In Crop Protection under Changing Climate; Jabran, K., Florentine, S., Chauhan, B., Eds.; Springer: Cham, Switzerland, 2020; pp. 85–100. [Google Scholar]
- Lamichhane, J.R.; Barzman, M.; Booij, K.; Boonekamp, P.; Desneux, N.; Huber, L.; Kudsk, P.; Langrell, S.R.H.; Ratnadass, A.; Ricci, P.; et al. Robust cropping systems to tackle pests under climate change. A review. Agron. Sustain. Dev. 2015, 35, 443–459. [Google Scholar] [CrossRef]
- Elena, G.; León, M.; Abad-Campos, P.; Armengol, J.; Mateu-Andrés, I.; Güemes-Heras, J. First report of Diplodia fraxini causing dieback of Fraxinus angustifolia in Spain. Plant Dis. 2018, 102, 2645–2646. [Google Scholar] [CrossRef]
- Pavlic-Zupanc, D.; Wingfield, M.J.; Boissin, E.; Slippers, B. The distribution of genetic diversity in the Neofusicoccum parvum/N. ribis complex suggests structure correlated with level of disturbance. Fungal Ecol. 2015, 13, 93–102. [Google Scholar] [CrossRef]
- Mehl, J.W.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Overlap of latent pathogens in the Botryosphaeriaceae on a native and agricultural host. Fungal Biol. 2017, 121, 405–419. [Google Scholar] [CrossRef]
- Hilario, S.; Lopes, A.; Santos, L.; Alves, A. Botryosphaeriaceae species associated with blueberry stem blight and dieback in the Center Region of Portugal. Eur. J. Plant Pathol. 2019, 156, 31–44. [Google Scholar] [CrossRef]
- Golzar, H.; Burgess, T.I. Neofusicoccum Parvum, a causal agent associated with cankers and decline of Norfolk Island pine in Australia. Australas. Plant Pathol. 2011, 40, 484–489. [Google Scholar] [CrossRef]
- Sakalidis, M.L.; Slippers, B.; Wingfield, B.D.; Hardy, G.E.S.J.; Burgess, T.I. The challenge of understanding the origin, pathways and extent of fungal invasions: Global populations of the Neofusicoccum parvum-N. ribis species complex. Divers. Distrib. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Gladieux, P.; Feurtey, A.; Hood, M.E.; Snirc, A.; Clavels, J.; Dutech, C.; Roy, M.; Giraud, T. The population biology of fungal invasions. Mol. Ecol. 2015, 24, 1969–1986. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Brglez, A.; Piškur, B.; Ogris, N. First report of Diplodia fraxini and Diplodia subglobosa causing canker and dieback of Fraxinus excelsior in Slovenia. Plant Dis. 2022, 106, 26–29. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Region | Altitude (m. a.s.l.) | Geographic Coordinates | Number of Samples Collected | |
|---|---|---|---|---|---|
| Latitude (X) | Longitude (Y) | ||||
| 1 | Tuscany | 498 | 43.065824° | 11.070266° | 3 |
| 2 | Tuscany | 448 | 43.055333° | 11.081245° | 6 |
| 3 | Tuscany | 439 | 43.050120° | 11.082984° | 4 |
| 4 | Tuscany | 722 | 43.033821° | 11.094413° | 3 |
| 5 | Tuscany | 554 | 43.024153° | 11.101669° | 1 |
| 6 | Tuscany | 553 | 43.023907° | 11.101867° | 3 |
| 7 | Tuscany | 545 | 43.021426° | 11.093438° | 3 |
| 8 | Tuscany | 312 | 43.014474° | 11.091555° | 3 |
| 9 | Tuscany | 306 | 43.002083° | 11.111248° | 4 |
| 10 | Tuscany | 323 | 43.091973° | 11.170991° | 4 |
| 11 | Tuscany | 334 | 43.090548° | 11.171011° | 4 |
| 12 | Tuscany | 347 | 43.142534° | 11.115229° | 5 |
| 13 | Tuscany | 333 | 43.140372° | 11.112651° | 5 |
| 14 | Tuscany | 431 | 43.172679° | 11.045179° | 2 |
| 15 | Tuscany | 438 | 43.165697° | 11.032799° | 2 |
| 16 | Tuscany | 609 | 43.153484° | 11.031911° | 4 |
| 17 | Tuscany | 556 | 43.154064° | 11.035591° | 4 |
| 18 | Veneto | 240 | 45.309950° | 11.773347° | 6 |
| 19 | Veneto | 216 | 45.289807° | 11.703996° | 5 |
| 20 | Veneto | 184 | 45.300556° | 11.767728° | 4 |
| 21 | Veneto | 259 | 45.318134° | 11.708450° | 2 |
| 22 | Veneto | 50 | 45.264882° | 11.717811° | 5 |
| 23 | Friuli Venezia Giulia | 567 | 45.988357° | 13.634403° | 3 |
| 24 | Friuli Venezia Giulia | 64 | 45.945518° | 13.552939° | 4 |
| 25 | Friuli Venezia Giulia | 187 | 45.8334681° | 13.580679° | 2 |
| Species | No. of Isolates | Sites | ITS GenBank Code |
|---|---|---|---|
| Botryosphaeria dothidea | 53 a + 21 b | 1–20; 22–24 | OR119872 |
| Diplodia fraxini | 42 a + 15 b | 1–5; 9–22; 25 | OR177960 |
| Diplodia subglobosa | 3 b | 19; 24 | OR805517 |
| Dothiorella iberica | 2 b | 18 | OR805518 |
| Dothiorella omnivora | 2 b | 21 | OR805519 |
| Neofusicoccum parvum | 34 a + 3 b | 2; 4; 7–9; 11–15; 19; 23 | OR835588 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benigno, A.; Aglietti, C.; Rossetto, G.; Bregant, C.; Linaldeddu, B.T.; Moricca, S. Botryosphaeriaceae Species Associated with Stem Canker, Shoot Blight and Dieback of Fraxinus ornus in Italy. Forests 2024, 15, 51. https://doi.org/10.3390/f15010051
Benigno A, Aglietti C, Rossetto G, Bregant C, Linaldeddu BT, Moricca S. Botryosphaeriaceae Species Associated with Stem Canker, Shoot Blight and Dieback of Fraxinus ornus in Italy. Forests. 2024; 15(1):51. https://doi.org/10.3390/f15010051
Chicago/Turabian StyleBenigno, Alessandra, Chiara Aglietti, Giovanni Rossetto, Carlo Bregant, Benedetto Teodoro Linaldeddu, and Salvatore Moricca. 2024. "Botryosphaeriaceae Species Associated with Stem Canker, Shoot Blight and Dieback of Fraxinus ornus in Italy" Forests 15, no. 1: 51. https://doi.org/10.3390/f15010051
APA StyleBenigno, A., Aglietti, C., Rossetto, G., Bregant, C., Linaldeddu, B. T., & Moricca, S. (2024). Botryosphaeriaceae Species Associated with Stem Canker, Shoot Blight and Dieback of Fraxinus ornus in Italy. Forests, 15(1), 51. https://doi.org/10.3390/f15010051