
The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
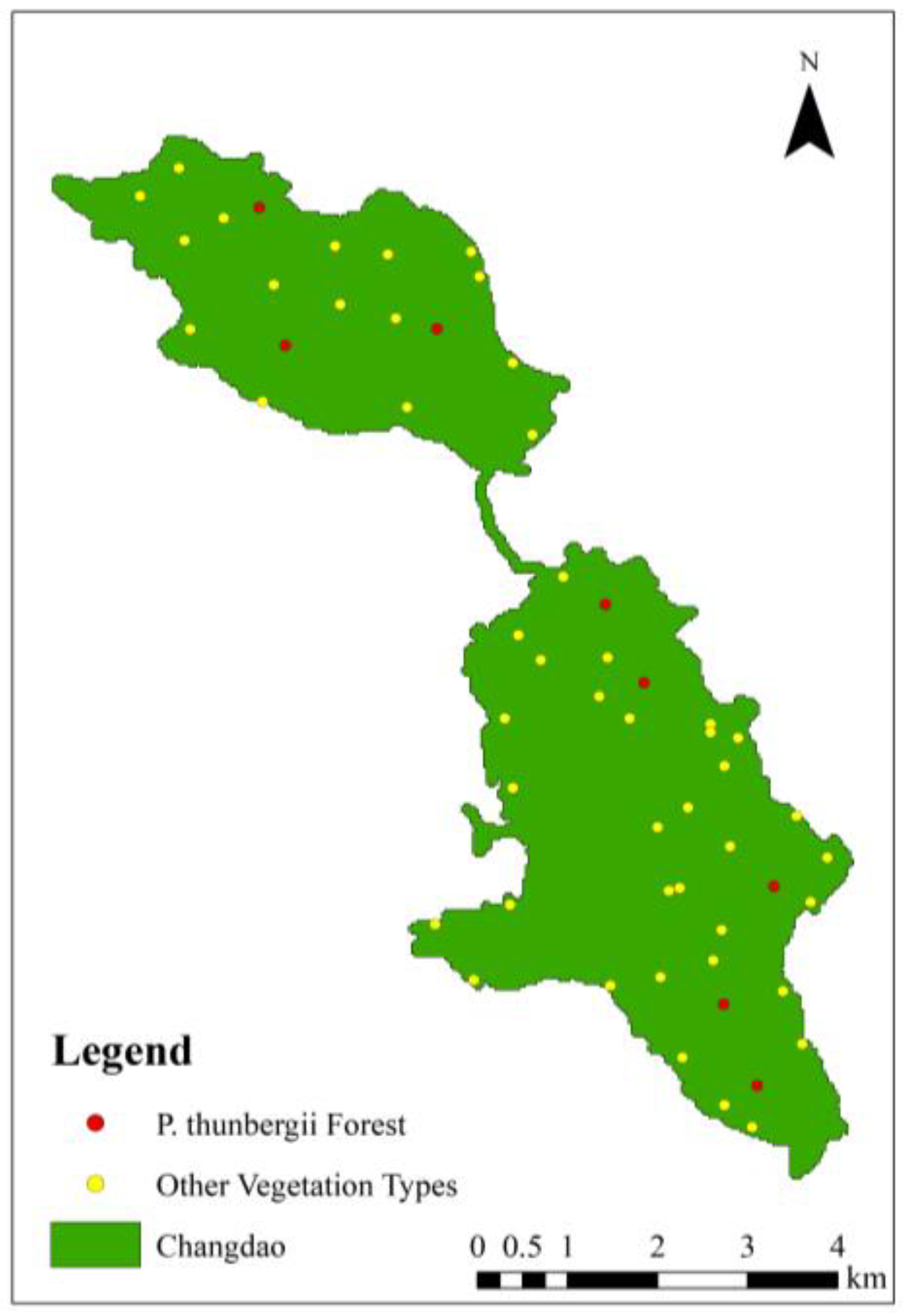
2.1. Research Area
2.2. Classification of Black Pines
2.3. Soil pH Survey
2.4. Regulate Soil pH in Pot
2.5. Regulate Soil pH In Situ Experiment
3. Results
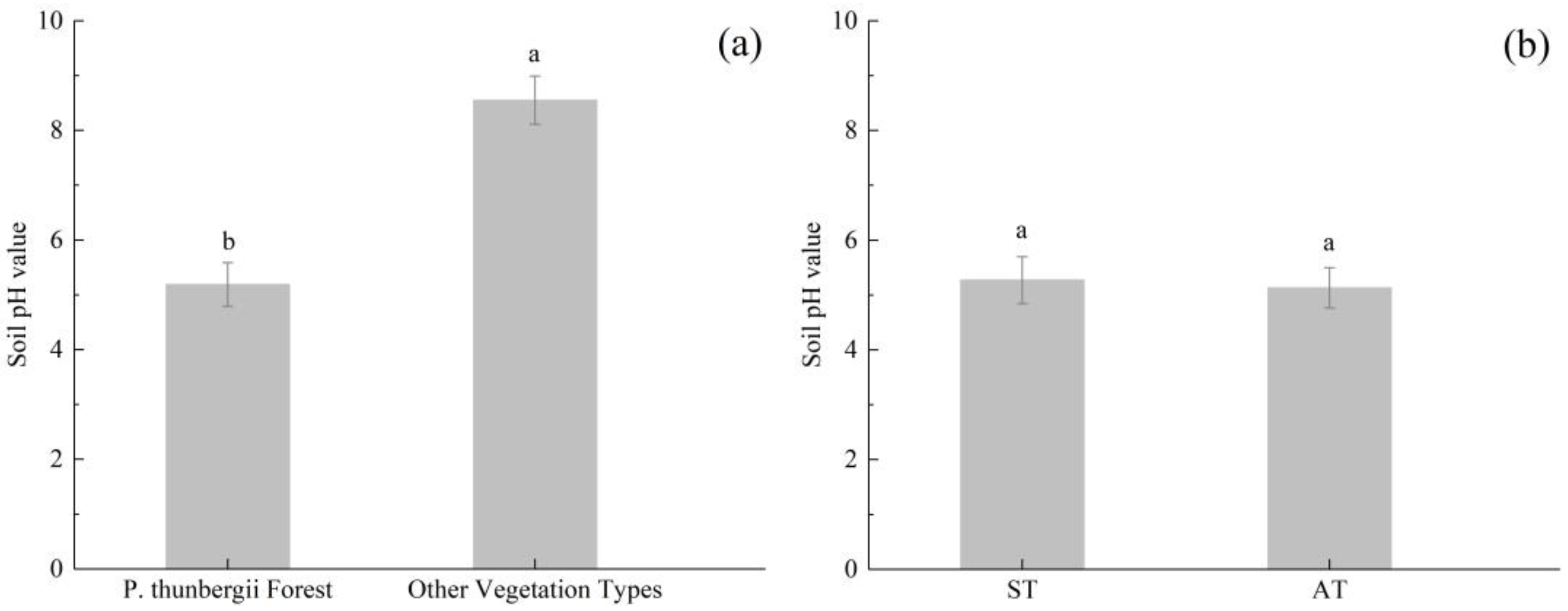
3.1. Soil pH Characteristics
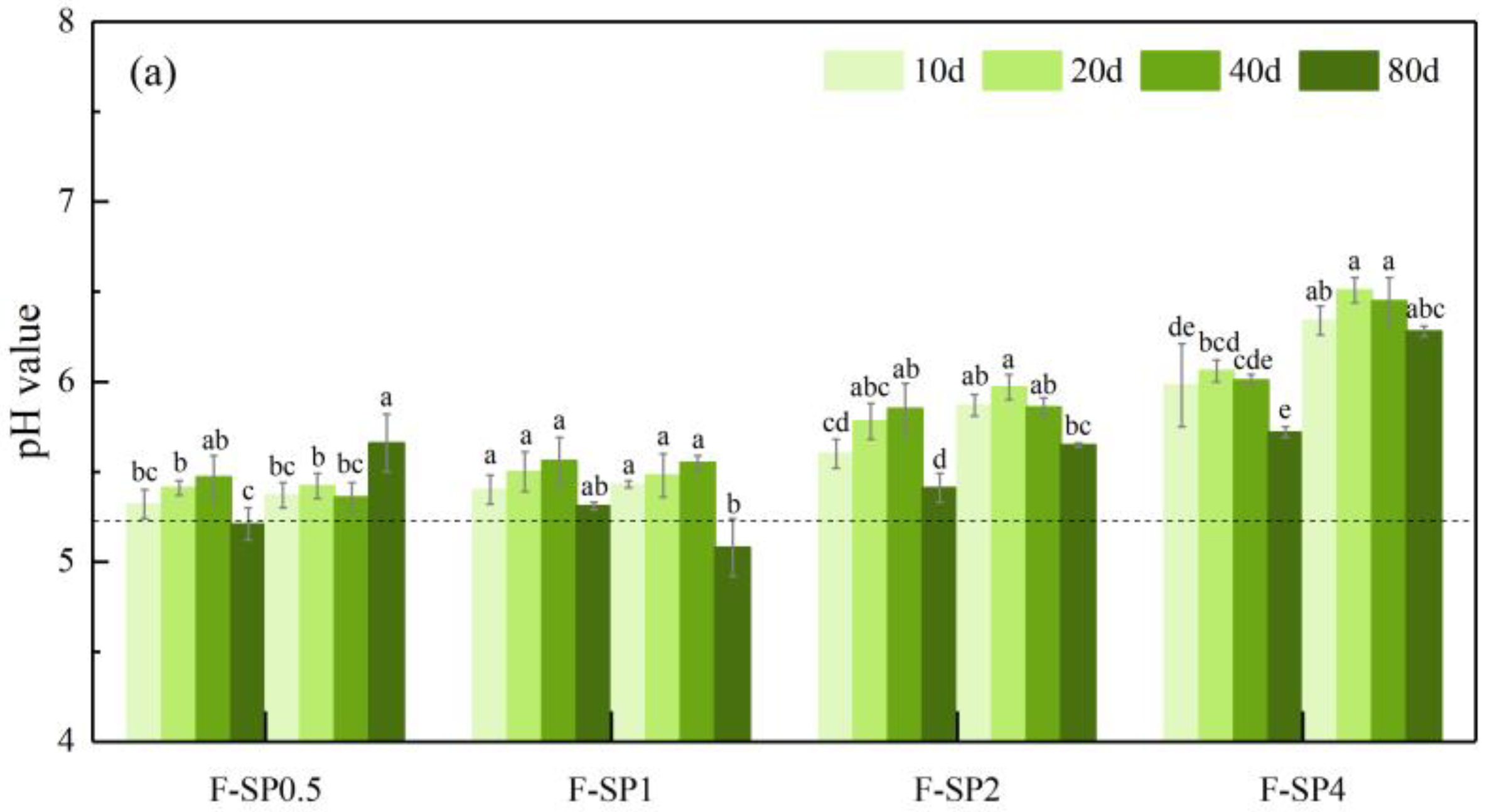
3.2. The Effect of SP with Different Particle Sizes on Regulating Soil pH
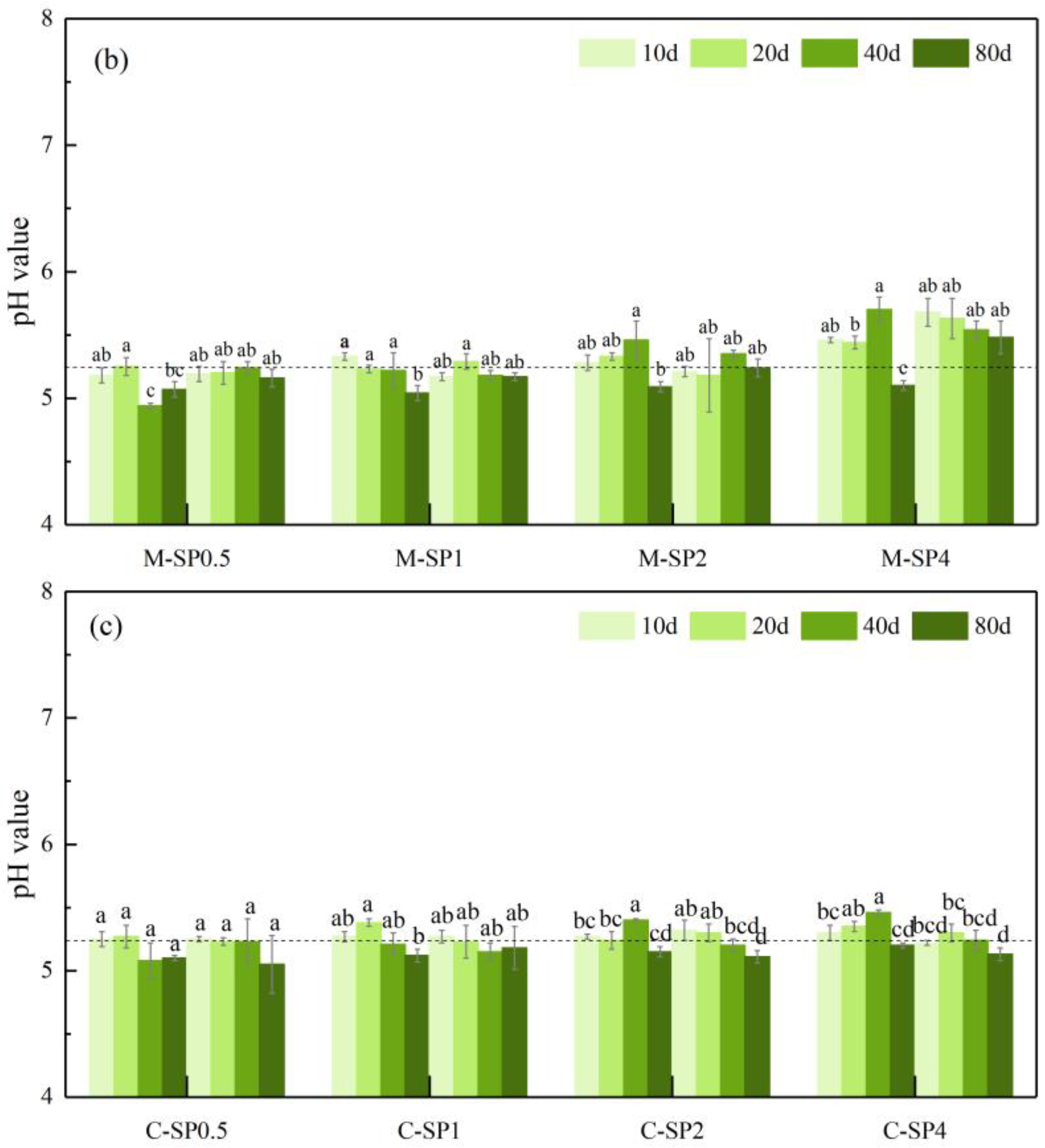
3.3. The Effect of Different Ameliorants on Regulating Soil pH
3.4. The Result of In Situ Experiment
4. Discussion
4.1. Where Does H+ Come from in P. thunbergii Forest?
4.2. The Internal Mechanism between Soil pH and PWD
4.3. The Common Method in Deal with PWD
4.4. Successful Cases of Controlling Plant Diseases by Regulating Soil pH
4.5. Soil pH Regulation with SP and Its Prospects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xu, X.L.; Wu, X.Q.; Ye, J.R.; Huang, L. Molecular characterization and functional analysis of three pathogenesis-related cytochrome P450 genes from Bursaphelenchus xylophilus (Tylenchida: Aphelenchoidoidea). Int. J. Mol. Sci. 2015, 16, 5216–5234. [Google Scholar] [CrossRef]
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Cheng, H.R.; Lin, M.S.; Qian, R.J. A study on morphological diagnosis and pathogenicity of the pine wood nematode. J. Nanjing Agric. Univ. 1986, 2, 55–59. [Google Scholar]
- Li, Z.X.; Tao, J.; Zong, S.X. Cold tolerance in pinewood nematode Bursaphelenchus xylophilus promoted multiple invasion events in mid-temperate zone of China. Forests 2022, 13, 1100. [Google Scholar] [CrossRef]
- Ye, J.R. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. [Google Scholar]
- Kiyohara, T.; Togushige, Y. Inoculation experiments of a nematode, Bursaphelenchus sp. onto pine trees. J. Jpn. For. Soc. 1971, 53, 105–114. [Google Scholar]
- Khan, M.A.; Khan, R.; Khan, Y.; Islam, S. A mathematical analysis of Pine Wilt disease with variable population size and optimal control strategies. Chaos Solitons Fractals 2018, 108, 205–217. [Google Scholar] [CrossRef]
- Song, Y.S.; Ye, J.R. The Occurrence and Control of Pine Wood Nematode Disease in China; China Forestry Press: Beijing, China, 2019. [Google Scholar]
- Ghorbani, R.; Wilcockson, S.; Koocheki, A.; Leifert, C. Soil management for sustainable crop disease control: A review. Environ. Chem. Lett. 2008, 6, 149–162. [Google Scholar] [CrossRef]
- Fang, X.L.; You, M.P.; Barbetti, M.J. Reduced severity and impact of Fusarium wilt on strawberry by manipulation of soil pH, soil organic amendments and crop rotation. Eur. J. Plant Pathol. 2012, 134, 619–629. [Google Scholar] [CrossRef]
- Fang, X.L.; Phillips, D.; Li, H.; Sivasithamparam, K.; Barbetti, M.J. Severity of crown and root diseases of strawberry and associated fungal and oomycete pathogens in Western Australia. Australas. Plant Pathol. 2011, 40, 109–119. [Google Scholar] [CrossRef]
- Liu, T.Y.; Liu, M.Y.; Zhao, Z.W. Occurrence and control of Meloidogyne disease in vegetables. China Cucurbits Veg. 2011, 24, 46–48. [Google Scholar]
- Wang, Y.; Song, Y.Q.; Zang, X.Q.; You, J.X. Test of the diseased wood caused by pine wood nematode with two indicators. Acta Phytopathol. Sin. 1996, 26, 371–376. [Google Scholar]
- Futai, K.; Harashima, S. Effect of simulated acid mist on pine wilt disease. J. Jpn. For. Soc. 1990, 72, 520–523. [Google Scholar]
- Li, M.; Li, H.; Ding, X.L.; Wang, L.C.; Wang, X.Y.; Chen, F.M. The detection of pine wilt disease: A literature review. Int. J. Mol. Sci. 2022, 23, 10797. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.Q. Effect of Oyster Shell Powder on Adjusting Soil pH and Controlling Tobacco Bacterial Wilt. Master’s Thesis, Southwest University, Chongqing, China, 2020. [Google Scholar]
- Asai, E.; Futai, K. The effects of long-term exposure to simulated acid rain on the development of pine wilt disease caused by Bursaphelenchus xylophilus. For. Path. 2001, 31, 241–253. [Google Scholar] [CrossRef]
- Markewitz, D.; Richter, D.D.; Allen, H.L.; Urrego, J.B. Three decades of observed soil acidification in the Calhoun experimental forest: Has acid rain made a difference? Soil Sci. Soc. Am. J. 1998, 62, 1428–1439. [Google Scholar] [CrossRef]
- Nakahara, O.; Takahashi, M.; Sase, H.; Yamada, T.; Matsuda, K.; Ohizumi, T.; Fukuhara, H.; Inoue, T.; Takahashi, A.; Kobayashi, H. Soil and stream water acidification in a forested catchment in central Japan. Biogeochemistry 2020, 97, 141–158. [Google Scholar] [CrossRef]
- Rigueiro-Rodríguez, A.; Mosquera-Losada, M.R.; Fernández-Núñez, E. Afforestation of agricultural land with Pinus radiata D. Don and Betula alba L. in NW Spain: Effects on soil pH, understorey production and floristic diversity eleven years after establishment. Land Degrad. Dev. 2012, 23, 227–241. [Google Scholar] [CrossRef]
- Sollins, P.; McCorison, F.M. Nitrogen and carbon solution chemistry of an old growth coniferous forest watershed before and after cutting. Water Resour. Res. 1981, 17, 1409–1418. [Google Scholar] [CrossRef]
- Mabuhay, J.A.; Nakagoshi, N. Response of soil microbial communities to changes in a forest ecosystem brought about by pine wilt disease. Landsc. Ecol. Eng. 2012, 8, 189–196. [Google Scholar] [CrossRef]
- Ma, Y.; Qu, Z.L.; Liu, B.; Tan, J.J.; Asiegbu, F.O.; Sun, H. Bacterial community structure of Pinus thunbergii naturally infected by the nematode Bursaphelenchus xylophilus. Microorganisms 2020, 8, 307. [Google Scholar] [CrossRef]
- Li, Z.P.; Li, D.J.; Wang, L.F.; Yang, B.J.; Qu, H.R.; Wu, Z.X.; He, C.Y. Study on occurrence regularity of pine wood nematode in Changdao County. Pract. For. Technol. 2002, 3, 15–16. [Google Scholar]
- Ryss, A.Y.; Kulinich, O.A.; Sutherland, J.R. Pine wilt disease: A short review of worldwide research. For. Sci. Pract. 2011, 13, 132–138. [Google Scholar] [CrossRef]
- Shi, L.N.; Wang, L.; Shi, X.F.; Luo, L.; Ye, J.R. Negative effects of free-living nematodes on the populations of Bursaphelenchus xylophilus in dead pine trees. Biol. Control 2022, 168, 104858. [Google Scholar] [CrossRef]
- Chu, H.L.; Wang, C.Y.; Wang, H.H.; Chen, H.; Tang, M. Pine wilt disease alters soil properties and root-associated fungal communities in Pinus tabulaeformis forest. Plant Soil 2016, 404, 237–249. [Google Scholar] [CrossRef]
- Ge, P.; Da, L.J.; Wang, W.B.; Xu, X.N. Seasonal dynamics of dissolved organic carbon, nitrogen and other nutrients in soil of Pinus massoniana stands after pine wilt disease disturbance. J. Soil Sci. Plant Nutr. 2014, 14, 75–87. [Google Scholar] [CrossRef]
- Yu, L.; Nicolaisen, M.; Larsen, J.; Ravnskov, S. Succession of root-associated fungi in Pisum sativum during a plant growth cycle as examined by 454 pyrosequencing. Plant soil 2012, 358, 225–233. [Google Scholar] [CrossRef]
- Treu, R.; Karst, J.; Randall, M.; Pec, G.J.; Cigan, P.W.; Simard, S.W.; Cooke, J.E.K.; Erbilgin, N.; Cahill, J.F. Decline of ectomycorrhizal fungi following a mountain pine beetle epidemic. Ecology 2014, 95, 1096–1103. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Li, Y.X.; Liu, Z.K.; Li, D.Z.; Wen, X.J.; Feng, Y.Q.; Zhang, X.Y. Pinewood nematode alters the endophytic and rhizospheric microbial communities of Pinus massoniana. Microb. Ecol. 2021, 81, 807–817. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Li, Y.X.; Wei, P.F.; Sun, N.N.; Wen, X.J.; Liu, Z.K.; Li, D.Z.; Feng, Y.Q.; Zhang, X.Y. Differences between microbial communities of Pinus species having differing level of resistance to the pine wood nematode. Microb. Ecol. 2021, 84, 1245–1255. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008; pp. 573–611. [Google Scholar]
- Kanda, S.; Aimi, T.; Kano, S.; Ishihara, S.; Kitamoto, Y.; Morinaga, T. Ambient pH signaling regulates expression of the serine protease gene (spr1) in pine wilt nematode-trapping fungus, Monacrosporium megalosporum. Microbiol. Res. 2008, 163, 63–72. [Google Scholar] [CrossRef]
- Negi, S.; Ye, J.R. Occurrence of nematode species associated with pine wood in Jiangsu, Zhejiang and Gansu provinces in China. J. Nanjing For. Univ. 2011, 54, 149–150. [Google Scholar]
- Räty, M.; Huhta, V. Earthworms and pH affect communities of nematodes and enchytraeids in forest soil. Biol. Fertil. Soils 2003, 38, 52–58. [Google Scholar] [CrossRef]
- Xie, L.Q.; Zhao, B.G. Post-inoculation population dynamics of Bursaphelenchus xylophilus and associated bacteria in pine wilt disease on Pinus thunbergii. J. Phytopathol. 2008, 156, 385–389. [Google Scholar] [CrossRef]
- dos Anjos Gonçalves, L.D.; Piccoli, R.H.; de Paula Peres, A.; Saúde, A.V. Predictive modeling of Pseudomonas fluorescens growth under different temperature and pH values. Braz. J. Microbiol. 2017, 48, 352–358. [Google Scholar] [CrossRef]
- Guo, Y.J.; Lin, Q.N.; Chen, L.; Carballar-Lejarazú, R.; Zhang, A.S.; Shao, E.S.; Liang, G.H.; Hu, X.; Wang, R.; Xu, L.; et al. Characterization of bacterial communities associated with the pinewood nematode insect vector Monochamus alternatus Hope and the host tree Pinus massoniana. BMC Genom. 2020, 21, 337. [Google Scholar] [CrossRef]
- Tantyama, T. Terrible Acid Rain; Godoshuppan: Tokyo, Japan, 1989. [Google Scholar]
- Asai, E.; Futai, K. Effects of inoculum density of pinewood nematode on the development of pine wilt disease in Japanese black pine seedlings pretreated with simulated acid rain. For. Path. 2005, 35, 135–144. [Google Scholar] [CrossRef]
- Asai, E.; Hata, K.; Futai, K. Effect of simulated acid rain on the occurrence of Lophodermium on Japanese black pine needles. Mycol. Res. 1998, 102, 1316–1318. [Google Scholar] [CrossRef]
- Hao, X.; Wang, B.W.; Chen, J.; Wang, B.Y.; Xu, J.Y.; Pan, J.L.; Ma, L. Molecular characterization and functional analysis of multidrug resistance-associated genes of Pinewood nematode (Bursaphelenchus xylophilus) for nematicides. Pestic. Biochem. Physiol. 2021, 177, 104902. [Google Scholar] [CrossRef]
- Domínguez, J.; Negrín, M.A.; Rodríguez, C.M. Soil chemical characteristics in relation to Fusarium wilts in banana crops of Gran Canaria Island (Spain). Commun. Soil Sci. Plant Anal. 1996, 27, 2649–2662. [Google Scholar] [CrossRef]
- Robson, A. Soil Acidity and Plant Growth; Elsevier: Amsterdam, The Netherlands, 1989; pp. 139–165. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pine Status | AT | ST | DT | IR | |||||
|---|---|---|---|---|---|---|---|---|---|
| Ameliorant | Before% | After% | Before% | After% | Before% | After% | Before% | After% | |
| F-SP | 50.8 | 68.9 | 33.9 | 18.0 | 15.3 | 13.1 | 49.2 | 31.1 | |
| QL | 45.6 | 50.0 | 43.5 | 38.1 | 10.9 | 11.9 | 54.4 | 50.0 | |
| CG | 61.8 | 50.7 | 32.4 | 42.5 | 5.8 | 6.8 | 38.2 | 49.3 | |
| p (Kruskal–Wallis) | 0.051 | 0.066 | 0.066 | 0.039 | 0.027 | 0.058 | 0.051 | 0.066 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, Y.; Liu, L.; Wu, C.; Yu, G.; Wang, Z.; Fan, J.; Tu, C. The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest. Forests 2023, 14, 1583. https://doi.org/10.3390/f14081583
Fan Y, Liu L, Wu C, Yu G, Wang Z, Fan J, Tu C. The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest. Forests. 2023; 14(8):1583. https://doi.org/10.3390/f14081583
Chicago/Turabian StyleFan, Ying, Lin Liu, Chuchen Wu, Guoxu Yu, Zhengyu Wang, Jingjing Fan, and Chengyi Tu. 2023. "The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest" Forests 14, no. 8: 1583. https://doi.org/10.3390/f14081583
APA StyleFan, Y., Liu, L., Wu, C., Yu, G., Wang, Z., Fan, J., & Tu, C. (2023). The Effect of Regulating Soil pH on the Control of Pine Wilt Disease in a Black Pine Forest. Forests, 14(8), 1583. https://doi.org/10.3390/f14081583