

Geographic Range vs. Occurrence Records in Plant Distribution Mapping: The Case of Arbutus in the Old World

Abstract
1. Introduction
1.1. Geographic Range Maps
1.2. Occurrence Record Maps
1.3. Arbutus in the Western Palearctic Region
2. Materials and Methods
- GBIF: records were selected from countries where Arbutus is known to be native, considering only georeferenced point occurrence data from selected institutions and datasets of specimen collections (Herbaria) and vegetation surveys [110,111,112,113,114,115,116,117,118]. Citizen science datasets were excluded. A complete list of the selected institutions and datasets, along with the DOI download reference, is listed in Table S1.
- Other databases and online repositories that are not donated to GBIF include: Botanical Society of Britain and Ireland [122], Nationwide Vegetation Plot Database–Sapienza University of Rome [123], Flora Ionica [124]; FAO Map Catalog [125]; EU-Forest [126]; Flora Croatica Database [127]; Lebanon Flora [128]; TÜBİVES–Turkish Plants Data Service [129]. Since many occurrences in these databases were only depicted on maps, it was often required to georeference this information. In some cases (e.g., Flora Croatica Database) the mapped occurrences were clearly representing the centroid of a cell of a grid map.
- Literature: an extensive bibliographic survey was carried out on regional floristic checklist, vegetation surveys, conservation reports, scientific papers, and National Floras [94,95,102,105,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168]. When geographic coordinates were not available, records from the literature were georeferenced based on the toponyms and the description of the recorded sites. The literature search was especially devoted to those regions that are underrepresented in the above-listed databases. A complete list of references is available in Table S1.
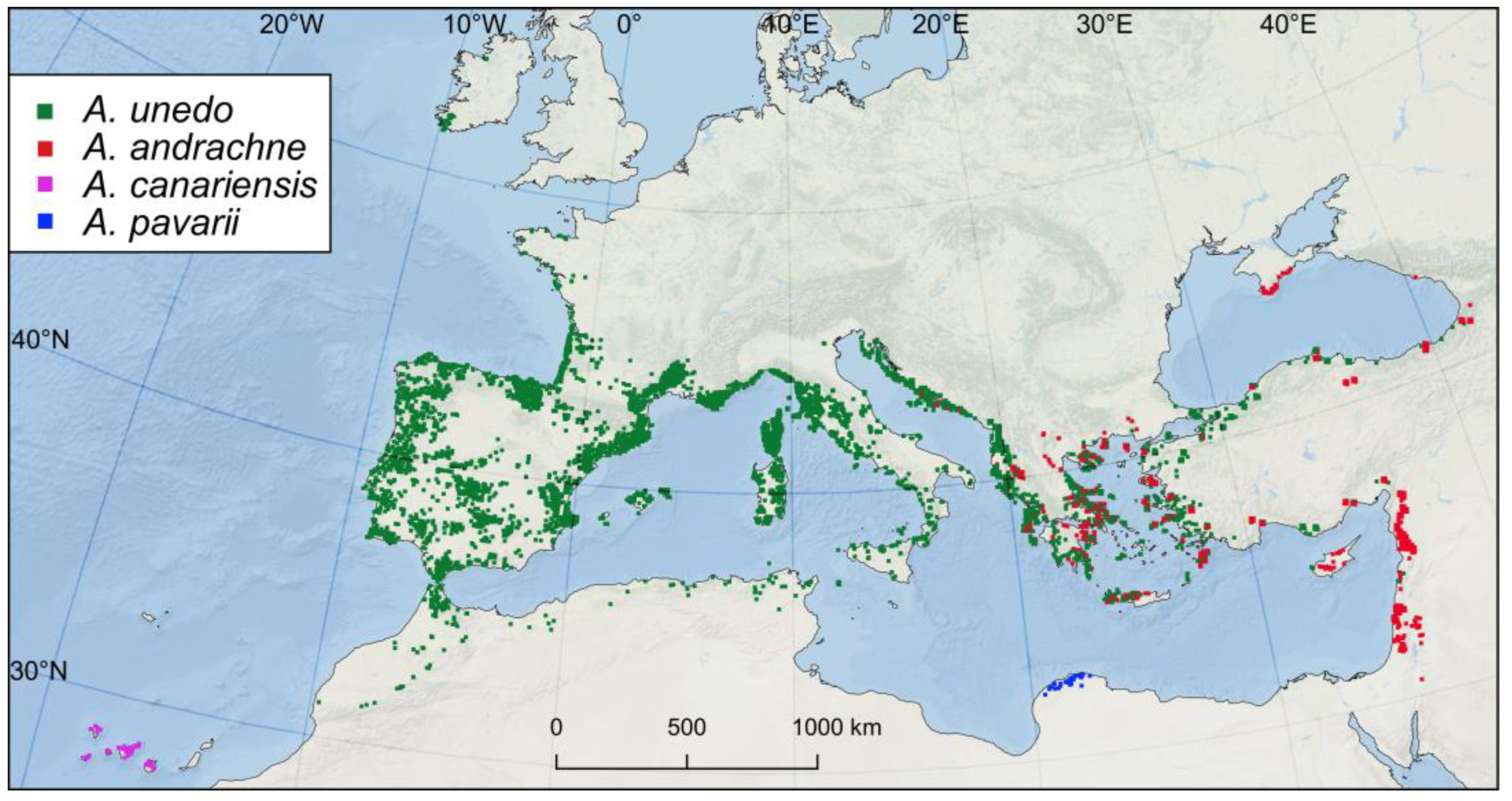
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gleason, H.; Cronquist, A. The Natural Geography of Plants; Columbia University Press: London, UK, 1964; p. 420. ISBN 978-0-231-02668-0. [Google Scholar]
- Meusel, H.; Jäger, E.J.; Weinert, E. Vergleichende Chorologie Der Zentraleuropaischen Flora; Fischer, G., Ed.; Gustav Fischer Verlag: Jena-Stuttgart, Germany, 1965. [Google Scholar]
- Walter, H.; Straka, H. Grundlagen Der Pflanzenverbreitung: 2: Arealkunde; Floristisch-Historische Geobotanik; Eugen Ulmer: Stuttgart, Germany, 1970. [Google Scholar]
- Ellenberg, H.H. Vegetation Ecology of Central Europe; Columbia University Press: London, UK, 1988; ISBN 0-521-23642-8. [Google Scholar]
- Ehrendorfer, F. Woody Plants-Evolution and Distribution Since the Tertiary; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1989; Volume 162, ISBN 3-211-82124-4. [Google Scholar]
- Barkowsky, T.; Freksa, C. Cognitive Requirements on Making and Interpreting Maps; University of Hamburg: Hamburg, Germany, 1997; pp. 347–361. [Google Scholar]
- Bunch, R.L.; Lloyd, R.E. The Cognitive Load of Geographic Information. Prof. Geogr. 2006, 58, 209–220. [Google Scholar] [CrossRef]
- Ooms, K.; De Maeyer, P.; Fack, V.; Van Assche, E.; Witlox, F. Interpreting Maps through the Eyes of Expert and Novice Users. Int. J. Geogr. Inf. Sci. 2012, 26, 1773–1788. [Google Scholar] [CrossRef]
- Keskin, M.; Ooms, K.; Dogru, A.O.; De Maeyer, P. Exploring the Cognitive Load of Expert and Novice Map Users Using EEG and Eye Tracking. ISPRS Int. J. Geo-Inf. 2020, 9, 429. [Google Scholar] [CrossRef]
- Pedrotti, F. Plant and Vegetation Mapping; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-30234-3. [Google Scholar]
- Rapoport, E.H. Areography: Geographical Strategies of Species; Elsevier: Amsterdam, The Netherlands, 2013; Volume 1, ISBN 1-4831-5277-4. [Google Scholar]
- Gaston, K.J. How Large Is a Species’ Geographic Range? Oikos 1991, 61, 434–438. [Google Scholar] [CrossRef]
- Gaston, K.J. Measuring Geographic Range Sizes. Ecography 1994, 17, 198–205. [Google Scholar] [CrossRef]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The Geographic Range: Size, Shape, Boundaries, and Internal Structure. Annu. Rev. Ecol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef]
- IUCN Species Survival Commission. IUCN Red List Categories and Criteria; Version 3.1; IUCN Publisher: Gland, Switzerland, 2001. [Google Scholar]
- Jetz, W.; McPherson, J.M.; Guralnick, R.P. Integrating Biodiversity Distribution Knowledge: Toward a Global Map of Life. Trends Ecol. Evol. 2012, 27, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Weigelt, P.; Kreft, H. Multidimensional Biases, Gaps and Uncertainties in Global Plant Occurrence Information. Ecol. Lett. 2016, 19, 992–1006. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the Predictive Performance of Habitat Models Developed Using Logistic Regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting Species Distributions for Conservation Decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Franklin, J.; Davis, F.W.; Ikegami, M.; Syphard, A.D.; Flint, L.E.; Flint, A.L.; Hannah, L. Modeling Plant Species Distributions under Future Climates: How Fine Scale Do Climate Projections Need to Be? Glob. Change Biol. 2013, 19, 473–483. [Google Scholar] [CrossRef]
- Merow, C.; Wilson, A.M.; Jetz, W. Integrating Occurrence Data and Expert Maps for Improved Species Range Predictions. Glob. Ecol. Biogeogr. 2017, 26, 243–258. [Google Scholar] [CrossRef]
- Salvà-Catarineu, M.; Romo, A.; Mazur, M.; Zielińska, M.; Minissale, P.; Dönmez, A.A.; Boratyńska, K.; Boratyński, A. Past, Present, and Future Geographic Range of the Relict Mediterranean and Macaronesian Juniperus phoenicea Complex. Ecol. Evol. 2021, 11, 5075–5095. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; White, E.P. Disparity between Range Map- and Survey-Based Analyses of Species Richness: Patterns, Processes and Implications. Ecol. Lett. 2005, 8, 319–327. [Google Scholar] [CrossRef]
- Graham, C.H.; Hijmans, R.J. A Comparison of Methods for Mapping Species Ranges and Species Richness. Glob. Ecol. Biogeogr. 2006, 15, 578–587. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Jetz, W. Species Richness, Hotspots, and the Scale Dependence of Range Maps in Ecology and Conservation. Proc. Natl. Acad. Sci. USA 2007, 104, 13384–13389. [Google Scholar] [CrossRef]
- Barthlott, W.; Hostert, A.; Kier, G.; Küper, W.; Kreft, H.; Mutke, J.; Rafiqpoor, M.D.; Sommer, J.H. Geographic Patterns of Vascular Plant Diversity at Continental to Global Scales (Geographische Muster Der Gefäßpflanzenvielfalt Im Kontinentalen Und Globalen Maßstab). Erdkunde 2007, 61, 305–315. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Booher, D.B.; Urban, M.C.; Ulrich, W.; Suarez, A.V.; Skelly, D.K.; Russell, D.J.; Rowe, R.J.; Rothendler, M.; Rios, N.; et al. Estimating Species Relative Abundances from Museum Records. Methods Ecol. Evol. 2023, 14, 431–443. [Google Scholar] [CrossRef]
- Araújo, M.B.; Thuiller, W.; Williams, P.H.; Reginster, I. Downscaling European Species Atlas Distributions to a Finer Resolution: Implications for Conservation Planning. Glob. Ecol. Biogeogr. 2005, 14, 17–30. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; El-Kassaby, Y.A.; Yang, X.; Tian, X.; Wang, T. Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana. Forests 2022, 13, 129. [Google Scholar] [CrossRef]
- Graham, C.H.; Ferrier, S.; Huettman, F.; Moritz, C.; Peterson, A.T. New Developments in Museum-Based Informatics and Applications in Biodiversity Analysis. Trends Ecol. Evol. 2004, 19, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Guerin, G.R.; Andersen, A.N.; Rossetto, M.; van Leeuwen, S.; Byrne, M.; Sparrow, B.; Rodrigo, M.; Lowe, A.J. When Macroecological Transitions Are a Fiction of Sampling: Comparing Herbarium Records to Plot-Based Species Inventory Data. Ecography 2018, 41, 1864–1875. [Google Scholar] [CrossRef]
- Rondinini, C.; Stuart, S.; Boitani, L. Habitat Suitability Models and the Shortfall in Conservation Planning for African Vertebrates. Conserv. Biol. 2005, 19, 1488–1497. [Google Scholar] [CrossRef]
- Rocchini, D.; Hortal, J.; Lengyel, S.; Lobo, J.M.; Jiménez-Valverde, A.; Ricotta, C.; Bacaro, G.; Chiarucci, A. Accounting for Uncertainty When Mapping Species Distributions: The Need for Maps of Ignorance. Prog. Phys. Geogr. Earth Environ. 2011, 35, 211–226. [Google Scholar] [CrossRef]
- Ladle, R.; Hortal, J. Mapping Species Distributions: Living with Uncertainty. Front. Biogeogr. 2013, 5, 8–9. [Google Scholar] [CrossRef]
- Fletcher, R.J., Jr.; Hefley, T.J.; Robertson, E.P.; Zuckerberg, B.; McCleery, R.A.; Dorazio, R.M. A Practical Guide for Combining Data to Model Species Distributions. Ecology 2019, 100, e02710. [Google Scholar] [CrossRef]
- Isaac, N.J.B.; Jarzyna, M.A.; Keil, P.; Dambly, L.I.; Boersch-Supan, P.H.; Browning, E.; Freeman, S.N.; Golding, N.; Guillera-Arroita, G.; Henrys, P.A.; et al. Data Integration for Large-Scale Models of Species Distributions. Trends Ecol. Evol. 2020, 35, 56–67. [Google Scholar] [CrossRef]
- Mathias, P.V.C.; Mendonça, C.V.; Rangel, T.F.L.V.B.; Diniz-Filho, J.A.F. Sensitivity of Macroecological Patterns of South American Parrots to Differences in Data Sources. Glob. Ecol. Biogeogr. 2004, 13, 193–198. [Google Scholar] [CrossRef]
- Franklin, J.; Serra-Diaz, J.M.; Syphard, A.D.; Regan, H.M. Big Data for Forecasting the Impacts of Global Change on Plant Communities. Glob. Ecol. Biogeogr. 2017, 26, 6–17. [Google Scholar] [CrossRef]
- Nualart, N.; Ibáñez, N.; Soriano, I.; López-Pujol, J. Assessing the Relevance of Herbarium Collections as Tools for Conservation Biology. Bot. Rev. 2017, 83, 303–325. [Google Scholar] [CrossRef]
- Stropp, J.; Ladle, R.J.; Malhado, A.C.M.; Hortal, J.; Gaffuri, J.; Temperley, W.H.; Olav Skøien, J.; Mayaux, P. Mapping Ignorance: 300 Years of Collecting Flowering Plants in Africa. Glob. Ecol. Biogeogr. 2016, 25, 1085–1096. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Rueda, M.; Rodríguez, M.Á. What Do Range Maps and Surveys Tell Us About Diversity Patterns? Folia Geobot 2008, 43, 345–355. [Google Scholar] [CrossRef]
- Cantú-Salazar, L.; Gaston, K.J. Species Richness and Representation in Protected Areas of the Western Hemisphere: Discrepancies between Checklists and Range Maps. Divers. Distrib. 2013, 19, 782–793. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Besnard, A.G.; Rödder, D.; Secondi, J. Confronting Expert-Based and Modelled Distributions for Species with Uncertain Conservation Status: A Case Study from the Corncrake (Crex crex). Biol. Conserv. 2013, 167, 161–171. [Google Scholar] [CrossRef]
- Nori, J.; Prieto-Torres, D.A.; Villalobos, F.; Loyola, R.; Rojas-Soto, O.; Parra, J.L.; Lira-Noriega, A.; Ortega-Andrade, H.M.; Monjeau, A.; Fuente, S.H.-D.L.; et al. Contrasting Biogeographical Patterns of Threatened Vertebrates on Islands Emerge from Disparities between Expert-Derived Maps and Global Biodiversity Information Facility Data. J. Biogeogr. 2023, 50, 418–427. [Google Scholar] [CrossRef]
- Gaston, K.J. The Structure and Dynamics of Geographic Ranges; Oxford Series in Ecology and Evolution; Oxford University Press: Oxford, UK; New York, NY, USA, 2003; ISBN 978-0-19-852641-4. [Google Scholar]
- Fourcade, Y. Comparing Species Distributions Modelled from Occurrence Data and from Expert-Based Range Maps. Implication for Predicting Range Shifts with Climate Change. Ecol. Inform. 2016, 36, 8–14. [Google Scholar] [CrossRef]
- Alhajeri, B.H.; Fourcade, Y. High Correlation between Species-Level Environmental Data Estimates Extracted from IUCN Expert Range Maps and from GBIF Occurrence Data. J. Biogeogr. 2019, 46, 1329–1341. [Google Scholar] [CrossRef]
- Brown, J.H.; Lomolino, M.V. Biogeography; Sinuer Associates Publishers: Sunderland, MA, USA, 1998; ISBN 0-87893-073-6. [Google Scholar]
- Rondinini, C.; Wilson, K.A.; Boitani, L.; Grantham, H.; Possingham, H.P. Tradeoffs of Different Types of Species Occurrence Data for Use in Systematic Conservation Planning. Ecol. Lett. 2006, 9, 1136–1145. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/en (accessed on 27 September 2022).
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological Maps for the Main European Woody Species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef]
- Perry, C.H.; Finco, M.V.; Wilson, B.T. (Eds.) Forest Atlas of the United States; USDA: Washington, DC, USA, 2022; Volume 1172. [Google Scholar]
- AgroAtlas–Home. Available online: http://www.agroatlas.ru/ (accessed on 27 March 2023).
- Xu, X.; Naqinezhad, A.; Ghazanfar, S.A.; Fragman-Sapir, O.; Oganesian, M.; Dagher Kharrat, M.B.; Taifour, H.; Filimban, F.Z.; Matchutadze, I.; Shavvon, R.S.; et al. Mapping Asia Plants: Current Status on Floristic Information in Southwest Asia. Glob. Ecol. Conserv. 2020, 24, e01257. [Google Scholar] [CrossRef]
- Fang, J.; Wang, Z.; Tang, Z. Atlas of Woody Plants in China: Distribution and Climate; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; ISBN 978-3-642-15017-3. [Google Scholar]
- Debreczy, Z.; Rácz, I.; Musial, K. Conifers around the World; DendroPress: Budapest, Hungary, 2011; ISBN 963-219-063-7. [Google Scholar]
- Fortin, M.-J.; Keitt, T.H.; Maurer, B.A.; Taper, M.L.; Kaufman, D.M.; Blackburn, T.M. Species’ Geographic Ranges and Distributional Limits: Pattern Analysis and Statistical Issues. Oikos 2005, 108, 7–17. [Google Scholar] [CrossRef]
- Rotenberry, J.T.; Balasubramaniam, P. Connecting Species’ Geographical Distributions to Environmental Variables: Range Maps versus Observed Points of Occurrence. Ecography 2020, 43, 897–913. [Google Scholar] [CrossRef]
- Soberón, J.M.; Llorente, J.B.; Oñate, L. The Use of Specimen-Label Databases for Conservation Purposes: An Example Using Mexican Papilionid and Pierid Butterflies. Biodivers. Conserv. 2000, 9, 1441–1466. [Google Scholar] [CrossRef]
- Suarez, A.V.; Tsutsui, N.D. The Value of Museum Collections for Research and Society. BioScience 2004, 54, 66–74. [Google Scholar] [CrossRef]
- Pyke, G.H.; Ehrlich, P.R. Biological Collections and Ecological/Environmental Research: A Review, Some Observations and a Look to the Future. Biol. Rev. 2010, 85, 247–266. [Google Scholar] [CrossRef]
- Hedrick, B.P.; Heberling, J.M.; Meineke, E.K.; Turner, K.G.; Grassa, C.J.; Park, D.S.; Kennedy, J.; Clarke, J.A.; Cook, J.A.; Blackburn, D.C.; et al. Digitization and the Future of Natural History Collections. BioScience 2020, 70, 243–251. [Google Scholar] [CrossRef]
- Roleček, J.; Chytrý, M.; Hájek, M.; Lvončík, S.; Tichý, L. Sampling Design in Large-Scale Vegetation Studies: Do Not Sacrifice Ecological Thinking to Statistical Purism! Folia Geobot 2007, 42, 199–208. [Google Scholar] [CrossRef]
- Chao, A.; Kubota, Y.; Zelený, D.; Chiu, C.-H.; Li, C.-F.; Kusumoto, B.; Yasuhara, M.; Thorn, S.; Wei, C.-L.; Costello, M.J.; et al. Quantifying Sample Completeness and Comparing Diversities among Assemblages. Ecol. Res. 2020, 35, 292–314. [Google Scholar] [CrossRef]
- Guralnick, R.; Van Cleve, J. Strengths and Weaknesses of Museum and National Survey Data Sets for Predicting Regional Species Richness: Comparative and Combined Approaches. Divers. Distrib. 2005, 11, 349–359. [Google Scholar] [CrossRef]
- Meineke, E.K.; Davies, T.J.; Daru, B.H.; Davis, C.C. Biological Collections for Understanding Biodiversity in the Anthropocene. Philos. Trans. R. Soc. B Biol. Sci. 2018, 374, 20170386. [Google Scholar] [CrossRef]
- Nelson, G.; Sweeney, P.; Wallace, L.E.; Rabeler, R.K.; Allard, D.; Brown, H.; Carter, J.R.; Denslow, M.W.; Ellwood, E.R.; Germain-Aubrey, C.C.; et al. Digitization Workflows for Flat Sheets and Packets of Plants, Algae, and Fungi. Appl. Plant Sci. 2015, 3, 1500065. [Google Scholar] [CrossRef]
- Nelson, G.; Ellis, S. The History and Impact of Digitization and Digital Data Mobilization on Biodiversity Research. Philos. Trans. R. Soc. B Biol. Sci. 2018, 374, 20170391. [Google Scholar] [CrossRef]
- GBIF. Available online: https://www.gbif.org/ (accessed on 27 March 2023).
- Edwards, J.L. Research and Societal Benefits of the Global Biodiversity Information Facility. BioScience 2004, 54, 485–486. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.T.; Soberón, J.; Overton, J.M.; Aragón, P.; Lobo, J.M. Use of Niche Models in Invasive Species Risk Assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- García-Roselló, E.; Guisande, C.; Manjarrés-Hernández, A.; González-Dacosta, J.; Heine, J.; Pelayo-Villamil, P.; González-Vilas, L.; Vari, R.P.; Vaamonde, A.; Granado-Lorencio, C.; et al. Can We Derive Macroecological Patterns from Primary Global Biodiversity Information Facility Data? Glob. Ecol. Biogeogr. 2015, 24, 335–347. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, J.; Jiang, M.-C. Global Patterns of Fern Species Diversity: An Evaluation of Fern Data in GBIF. Plant Divers. 2022, 44, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Chytrý, M. Phytosociological Data Give Biased Estimates of Species Richness. J. Veg. Sci. 2001, 12, 441–444. [Google Scholar] [CrossRef]
- Boakes, E.H.; McGowan, P.J.K.; Fuller, R.A.; Chang-qing, D.; Clark, N.E.; O’Connor, K.; Mace, G.M. Distorted Views of Biodiversity: Spatial and Temporal Bias in Species Occurrence Data. PLOS Biol. 2010, 8, e1000385. [Google Scholar] [CrossRef]
- Warren, D.L.; Cardillo, M.; Rosauer, D.F.; Bolnick, D.I. Mistaking Geography for Biology: Inferring Processes from Species Distributions. Trends Ecol. Evol. 2014, 29, 572–580. [Google Scholar] [CrossRef]
- Hughes, A.C.; Orr, M.C.; Ma, K.; Costello, M.J.; Waller, J.; Provoost, P.; Yang, Q.; Zhu, C.; Qiao, H. Sampling Biases Shape Our View of the Natural World. Ecography 2021, 44, 1259–1269. [Google Scholar] [CrossRef]
- Chapman, A.D. Principles of Data Quality; GBIF Secretariat: Copenhagen, Denmark, 2005; ISBN 87-92020-03-8. [Google Scholar]
- Feeley, K.J.; Silman, M.R. Modelling the Responses of Andean and Amazonian Plant Species to Climate Change: The Effects of Georeferencing Errors and the Importance of Data Filtering. J. Biogeogr. 2010, 37, 733–740. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized Cleaning of Occurrence Records from Biological Collection Databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial Bias in the GBIF Database and Its Effect on Modeling Species’ Geographic Distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Daru, B.H.; Park, D.S.; Primack, R.B.; Willis, C.G.; Barrington, D.S.; Whitfeld, T.J.S.; Seidler, T.G.; Sweeney, P.W.; Foster, D.R.; Ellison, A.M.; et al. Widespread Sampling Biases in Herbaria Revealed from Large-Scale Digitization. New Phytol. 2018, 217, 939–955. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, Z.A.; Harris, D.J.; Filer, D.; Wood, J.R.I.; Scotland, R.W. Widespread Mistaken Identity in Tropical Plant Collections. Curr. Biol. 2015, 25, R1066–R1067. [Google Scholar] [CrossRef] [PubMed]
- Kitchener, A.C.; Machado, F.A.; Hayssen, V.; Moehlman, P.D.; Viranta, S. Consequences of the Misidentification of Museum Specimens: The Taxonomic Status of Canis lupaster soudanicus. J. Mammal. 2020, 101, 1148–1150. [Google Scholar] [CrossRef]
- Shirai, M.; Takano, A.; Kurosawa, T.; Inoue, M.; Tagane, S.; Tanimoto, T.; Koganeyama, T.; Sato, H.; Terasawa, T.; Horie, T.; et al. Development of a System for the Automated Identification of Herbarium Specimens with High Accuracy. Sci. Rep. 2022, 12, 8066. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating Species Diversity and Distribution in the Era of Big Data: To What Extent Can We Trust Public Databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M.; Nipperess, D.A.; Gallagher, R.V.; Beaumont, L.J. How Well Documented Is Australia’s Flora? Understanding Spatial Bias in Vouchered Plant Specimens. Austral Ecol. 2017, 42, 690–699. [Google Scholar] [CrossRef]
- Marcer, A.; Haston, E.; Groom, Q.; Ariño, A.H.; Chapman, A.D.; Bakken, T.; Braun, P.; Dillen, M.; Ernst, M.; Escobar, A.; et al. Quality Issues in Georeferencing: From Physical Collections to Digital Data Repositories for Ecological Research. Divers. Distrib. 2021, 27, 564–567. [Google Scholar] [CrossRef]
- Marcer, A.; Chapman, A.D.; Wieczorek, J.R.; Xavier Picó, F.; Uribe, F.; Waller, J.; Ariño, A.H. Uncertainty Matters: Ascertaining Where Specimens in Natural History Collections Come from and Its Implications for Predicting Species Distributions. Ecography 2022, 2022, e06025. [Google Scholar] [CrossRef]
- Rikli, M. Pflanzenkleid der Mittelmeerländer; Hans Huber: Bern, Switzerland, 1943. [Google Scholar]
- Sealy, J.R. Arbutus unedo. J. Ecol. 1949, 37, 365–388. [Google Scholar] [CrossRef]
- Giacomini, V.; Fenaroli, L. La Flora; Touring Club italiano: Milano, Italy, 1958. [Google Scholar]
- Santiso, X.; López, L.; Gilbert, K.J.; Barreiro, R.; Whitlock, M.C.; Retuerto, R. Patterns of Genetic Variation within and among Populations in Arbutus unedo and Its Relation with Selection and Evolvability. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 185–192. [Google Scholar] [CrossRef]
- Wazen, N.; Garavaglia, V.; Picard, N.; Besacier, C.; Fady, B. Distribution Maps of Twenty-Four Mediterranean and European Ecologically and Economically Important Forest Tree Species Compiled from Historical Data Collections. Annals of Silvicultural Research 2020, 44, 95–101. [Google Scholar] [CrossRef]
- Palamarev, E. Paleobotanical Evidences of the Tertiary History and Origin of the Mediterranean Sclerophyll Dendroflora. Pl. Syst. Evol. 1989, 162, 93–107. [Google Scholar] [CrossRef]
- Anderson, C.L.; Channing, A.; Zamuner, A.B. Life, Death and Fossilization on Gran Canaria–Implications for Macaronesian Biogeography and Molecular Dating. J. Biogeogr. 2009, 36, 2189–2201. [Google Scholar] [CrossRef]
- Plants of the World Online|Kew Science. Available online: https://powo.science.kew.org/ (accessed on 27 March 2023).
- International Plant Names Index. Available online: https://www.ipni.org/ (accessed on 27 March 2023).
- Sealy, J.R.; Webb, D.A. Arbutus unedo L. J. Ecol. 1950, 38, 223–236. [Google Scholar] [CrossRef]
- Larcher, W. Temperature Stress and Survival Ability of Mediterranean Sclerophyllous Plants. Plant Biosyst.–Int. J. Deal. All Asp. Plant Biol. 2000, 134, 279–295. [Google Scholar] [CrossRef]
- Mosallam, H.A.E.-A.; Mohamed, A.H.; Sergiwa, S.S.; Gibreel, M.A.-A. Size Distribution of Some Endangered Plant Species, Al-Jabal Al-Akhdar, Libya. Egypt. J. Bot. 2017, 57, 181–197. [Google Scholar] [CrossRef]
- Sealy, J.R. The Swollen Stem-Base in Arbutus unedo. Kew Bull. 1949, 4, 241–251. [Google Scholar] [CrossRef]
- Canadell, J.; López-Soria, L. Lignotuber Reserves Support Regrowth Following Clipping of Two Mediterranean Shrubs. Funct. Ecol. 1998, 12, 31–38. [Google Scholar] [CrossRef]
- Paula, S.; Naulin, P.I.; Arce, C.; Galaz, C.; Pausas, J.G. Lignotubers in Mediterranean Basin Plants. Plant Ecol. 2016, 217, 661–676. [Google Scholar] [CrossRef]
- Bernetti, G. Selvicoltura Speciale; Unione tipografico-editrice Torinese: Torino, Italy, 1995; ISBN 88-02-04867-3. [Google Scholar]
- Braun-Blanquet, J. Plant Sociology. The Study of Plant Communities; McGraw-Hill Book Co.: New York, NY, USA; London, UK, 1932. [Google Scholar]
- Domingo, L.P.; Aedo Pérez, C. CSIC-Real Jardín Botánico-Anthos. Sistema de Información de Las Plantas de España. 2021. Available online: https://www.gbif.org/dataset/4cf3eec1-b902-40c9-b15b-05c5fe5928b6 (accessed on 30 March 2023). [CrossRef]
- Bruno, D.; Solèn, R. INPN–Données Flore Des CBN Agrégées Par La FCBN. 2016. Available online: https://www.gbif.org/dataset/75956ee6-1a2b-4fa3-b3e8-ccda64ce6c2d (accessed on 30 March 2023). [CrossRef]
- Nualart, N. Montes Institut Botanic de Barcelona (IBB, CSIC-Ajuntament de Barcelona), BC-Plantae. 2021. Available online: https://www.gbif.org/dataset/6aeebd1a-c3ad-4bc5-bdfe-24de0e2e9052 (accessed on 30 March 2023). [CrossRef]
- Salgueiro González, F.J. Herbario de La Universidad de Sevilla, SEV-Historico. 2020. Available online: https://www.gbif.org/dataset/83771684-f762-11e1-a439-00145eb45e9a (accessed on 30 March 2023). [CrossRef]
- Données de l’inventaire Forestier National de l’IGN–Relevés Floristiques Du Protocole de l’Inventaire National Forestier de l’IGN. 2021. Available online: https://www.gbif.org/dataset/e5f16d86-e225-4822-97be-a64ce17079c7 (accessed on 30 March 2023). [CrossRef]
- Carapeto, A.; Porto, M. Flora-On: Occurrence Data of the Flora of Mainland Portugal. 2016. Available online: https://www.gbif.org/dataset/7fe3eb5c-42bd-49d7-a30b-82c353ef6575 (accessed on 30 March 2023). [CrossRef]
- Horvitz, N. BioGIS–Vascular Plants–Israeli Gene Bank. 2015. Available online: https://www.gbif.org/dataset/25314346-645c-4197-8339-607c3ef4e273 (accessed on 30 March 2023). [CrossRef]
- Horvitz, N.; Danin, A. BioGIS–Vascular Plants–A. Danin’s Grid Database. 2015. Available online: https://www.gbif.org/dataset/863c1269-5f38-4862-a161-ea3a50654d4f (accessed on 30 March 2023). [CrossRef]
- Horvitz, N.; Olsvig-Whittaker, L. BioGIS–Vascular Plants–INPA Database. 2015. Available online: https://www.gbif.org/dataset/5a6b5bd4-cf31-4d5f-9a85-a331b7178a77 (accessed on 30 March 2023). [CrossRef]
- Wilson, B. IUCN Red List of Threatened Species: Arbutus andrachne. 2018. Available online: https://www.iucnredlist.org/species/19181048/119836468 (accessed on 30 March 2023).
- Beech, E. IUCN Red List of Threatened Species: Arbutus canariensis. 2017. Available online: https://www.iucnredlist.org/species/30322/81768520 (accessed on 30 March 2023).
- Harvey-Brown, Y IUCN Red List of Threatened Species: Arbutus pavarii. 2022. Available online: https://www.iucnredlist.org/species/30323/199350094 (accessed on 30 March 2023).
- Stroh, P.A.; Humphrey, T.; Burkmar, R.J.; Pescott, O.L.; Roy, D.B.; Walker, K.J. Strawberry-Tree Arbutus unedo L. In BSBI Online Plant Atlas 2020, Mapping Changes in the Distribution of the British and Irish Flora; Botanical Society of Britain and Ireland, Durham & Princeton University Press: Princeton, NJ, USA; Oxford, UK, 2020. [Google Scholar]
- Agrillo, E.; Alessi, N.; Massimi, M.; Spada, F.; De Sanctis, M.; Francesconi, F.; Attorre, F. Nationwide Vegetation Plot Database–Sapienza University of Rome: State of the Art, Basic Figures and Future Perspectives. Phytocoenologia 2017, 47, 221–229. [Google Scholar] [CrossRef]
- Flora Ionica Working Group Flora Ionica–An Inventory of Ferns and Flowering Plants of the Ionian Islands (Greece). Available online: https://floraionica.univie.ac.at (accessed on 30 March 2023).
- FAO Map Catalog. Available online: https://data.apps.fao.org/map/catalog/srv/eng/catalog.search#/metadata/54b5f7b6-7177-4973-83f1-6984b71aaff1 (accessed on 27 March 2023).
- Mauri, A.; Strona, G.; San-Miguel-Ayanz, J. EU-Forest, a High-Resolution Tree Occurrence Dataset for Europe. Sci. Data 2017, 4, 160123. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, T. Flora Croatica Database (FCD). Available online: https://hirc.botanic.hr/fcd/ (accessed on 30 March 2023).
- Lebanon Flora. Available online: http://lebanon-flora.org/ (accessed on 15 March 2023).
- Babac, M.; Uslu, E.; Bakis, Y. TÜBİVES-Turkish Plants Data Service. Available online: http://www.tubives.com/ (accessed on 30 March 2023).
- Barina, Z. Distribution Atlas of Vascular Plants in Albania; Magyar Természettudományi Múzeum: Budapest, Hungary, 2017; ISBN 963-9877-29-8. [Google Scholar]
- Stevenson, A.C.; Skinner, J.; Hollis, G.E.; Smart, M. The El Kala National Park and Environs, Algeria: An Ecological Evaluation. Environ. Conserv. 1988, 15, 335–348. [Google Scholar] [CrossRef]
- Zeddam, A.; Neff, C.; Meurer, M. Carte Phytoecologique de La Forêt Des Senhadja de Gherbès, Skikda (Algerie). In Proceedings of the Conférence internationale Dynamiques Environnementales et Histoires en domaines méditerranéens/Environmental Dynamics and History in the Mediterranean areas; Université de Paris Sorbonne, Paris, France, 24–26 April 2001. [Google Scholar]
- Boussalah, N.; Boussalah, D.; Cebadera-Miranda, L.; Fernández-Ruiz, V.; Barros, L.; Ferreira, I.C.; Mata, M.C.S.; Madani, K. Nutrient Composition of Algerian Strawberry-Tree Fruits (Arbutus unedo L.). Fruits 2018, 73, 283–297. [Google Scholar] [CrossRef]
- Boutabia, L.; Telailia, S.; Chefrour, A. La Flore Médicinale Du Massif Forestier d’Oum Ali (Zitouna-Wilaya d’El Tarf-Algérie): Inventaire et Étude Ethnobotanique. In Proceedings of the Actes des 15èmes Journées Scientifiques de l’INRGREF: <Valorisation des Produits Forestiers Non Ligneux>, Gammarth, Tunis, 28–29 September 2010. [Google Scholar]
- Kiniouar, S.; Roula, S.E. Caractérisation Des Groupements Végétaux des Falaises Littorales de l’Aire Marine Protégée Du Parc National de Taza, Université de Jijel, 2012, Master’s Thesis. Available online: https://www.theses-algerie.com/3270744763063056/memoire-de-master/universite-mohammed-seddik-ben-yahia---jijel/caract%C3%A9risation-des-groupements-v%C3%A9g%C3%A9taux-des-falaises-littorales-de-l-aire-marine-prot%C3%A9g%C3%A9e-du-parc-national-de-taza (accessed on 30 March 2023).
- Laouicha, S.; Senator, A.; Kherbache, A.; Bouriche, H. Total Phenolic Contents and Antioxidant Properties of Algerian Arbutus unedo L. Extracts. J. Drug Deliv. Ther. 2020, 10, 159–168. [Google Scholar] [CrossRef]
- Rebbas, K.; Vela, E.; Gharzouli, R.; Djellouli, Y.; Alatou, D.; Gachet, S. Phytosociological Characterization of the Vegetation of Gouraya National Park (Bejaia, Algeria). Rev. D Ecol.-La Terre Et La Vie 2011, 66, 267–289. [Google Scholar]
- Messaoudene, M.; Laribi, M.; Derridj, A. Etude de La Diversité Floristique de La Forêt de l’Akfadou (Algérie). Bois. Forets Des. Trop. 2007, 291, 75–81. [Google Scholar]
- Bessah, R.; Benyoussef, E.-H. Essential Oil Composition of Arbutus unedo L. Leaves from Algeria. J. Essent. Oil Bear. Plants 2012, 15, 678–681. [Google Scholar] [CrossRef]
- Sarmoum, M.; Feddag, F.; Masloub, A.; Belkaid, B. Diagnostic de l’état Actuel de La Suberaie Du Parc National de Theniet El Had (Wilaya Tissemsilt). In Proceedings of the Journées d’étude sur la réhabilitation des subéraies incendiées et reboisements, Recueil des résumes, Tlemcen, Algeria, 16–17 January 2013; pp. 17–18. [Google Scholar]
- Bouzid, A.; Chadli, R.; Bouzid, K. Ethnobotanical Study of the Medicinal Plant Arbutus unedo L. in the Region of Sidi Bel Abbes in Western Algeria. Phytothérapie 2017, 15, 373–378. [Google Scholar] [CrossRef]
- Letreuch-Belarouci, A.; Medjahdi, B.; Letreuch-Belarouci, N.; Benabdeli, K. Diversité Floristique des Subéraies Du Parc National de Tlemcen (Algérie). Acta Bot. Malacit. 2009, 34, 77–89. [Google Scholar] [CrossRef]
- Melia, N.; Gabedava, L.; Barblishvili, T.; Jgenti, L. Reproductive Biology Studies towards the Conservation of Two Rare Species of Colchic Flora, Arbutus andrachne and Osmanthus decorus. Turk. J. Bot. 2012, 36, 55–62. [Google Scholar] [CrossRef]
- Cordova, C.E. The Mediterraneanization of Crimea. Méditerranée. Rev. Géographique Des Pays Méditerranéens/J. Mediterr. Geogr. 2016, 126, 25–36. [Google Scholar] [CrossRef]
- Strid, A. Atlas of the Aegean Flora, Part 1: Text & Plates; Part 2: Maps; Englera; Botanic Garden and Botanical Museum Berlin, Freie Universität Berlin: Berlin, Germany, 2016; Volume 33, ISBN 978-3-921800-97-3/978-3-921800-98-0. [Google Scholar]
- Horvatić, S. Biljnogeografski Položaj i Raščlanjenje Našeg Primorja u Svjetlu Suvremenih Fitocenoloških Istraživanja. Acta Bot. Croat. 1963, 22, 27–81. [Google Scholar]
- Taifour, H.; El-Oqlah, A. Jordan Plant Red List; Royal Botanic Garden: Ammann, Jordan, 2014. [Google Scholar]
- Hegazy, A.; Boulos, L.; Kabiel, H.; Sharashy, O. Vegetation and Species Altitudinal Distribution in Al-Jabal Al-Akhdar Landscape, Libya. Pak. J. Bot 2011, 43, 1885–1898. [Google Scholar]
- Ait Lhaj, Z.; Bchitou, R.; Gaboun, F.; Abdelwahd, R.; Benabdelouahab, T.; Kabbour, M.R.; Pare, P.; Diria, G.; Bakhy, K. Moroccan Strawberry Tree (Arbutus unedo L.) Fruits: Nutritional Value and Mineral Composition. Foods 2021, 10, 2263. [Google Scholar] [CrossRef]
- Zitouni, H.; Hssaini, L.; Messaoudi, Z.; Ourradi, H.; Viuda-Martos, M.; Hernández, F.; Ercisli, S.; Hanine, H. Phytochemical Components and Bioactivity Assessment among Twelve Strawberry (Arbutus unedo L.) Genotypes Growing in Morocco Using Chemometrics. Foods 2020, 9, 1345. [Google Scholar] [CrossRef] [PubMed]
- Wahid, N.; Faida, R.; Aabdousse, J.; Boulli, A.; Bouda, S. Ethnobotanical Uses and Distribution Status of Arbutus (Arbutus unedo L.) in Morocco. Ethnobot. Res. Appl. 2019, 18, 1–12. [Google Scholar]
- Ajbilou, R.; Marañón, T.; Arroyo, J. Ecological and Biogeographical Analyses of Mediterranean Forests of Northern Morocco. Acta Oecologica 2006, 29, 104–113. [Google Scholar] [CrossRef]
- Mehdiqui, R.; Chattou, Z. Interrelation between Socio-Economic Factors of the Nearby Populations and Plant Resources of the Amsittene Forest (Essaouira-Morocco). Arx. De Sociol. 2009, 20, 71–83. [Google Scholar]
- Pavlović, D.R.; Branković, S.; Kovačević, N.; Kitić, D.; Veljković, S. Comparative Study of Spasmolytic Properties, Antioxidant Activity and Phenolic Content of Arbutus unedo from Montenegro and Greece. Phytother. Res. 2011, 25, 749–754. [Google Scholar] [CrossRef]
- Žnidaršič, A. Razširjenost in Ogroženost Navadne Jagodičnice (Arbutus unedo L.) v Sloveniji: Diplomsko Delo-Univerzitetni Študij. Master’s Thesis, University of Ljubljana, Ljubljana, Slovenia, 2014. [Google Scholar]
- Ghazal, A. Landscape Ecological, Phytosociological and Geobotanical Study of Eumediterranean in West of Syria. PhD Thesis, Faculty of Agricultural Science, University of Hohenheim, Stuttgart, Germany, 2008. [Google Scholar]
- Takrouni, M.M.; Boussaid, M. Genetic Diversity and Population’s Structure in Tunisian Strawberry Tree (Arbutus unedo L.). Sci. Hortic. 2010, 126, 330–337. [Google Scholar] [CrossRef]
- Dallali, S.; Bouraoui, H.; Mouhbi, R.; Zaouaoui, I.; Hasnaoui, F.; Toumi, L. Impact of Degradation on the Above-Ground Biomass of Plants Composing the Shrub Layer of Djebel Zaghouan. Int. J. Innov. Appl. Stud. 2020, 31, 336–350. [Google Scholar]
- Bachet, F.; Benhaj, S.; Bernard, F.; Delauge, J.; Harmelin, J.; Mante, A.; Pascal, M.; Tillmann, M.; Vela, E.; Vidal, P. Réserve Des Iles Habibas. Notes Naturalistes. Petites Îles de Méditerranée; Conservatoire de l’espace littoral et des rivages lacustres; Initiative PIM pour les Petites Iles de Méditerranée: Marseille, France, 2007. [Google Scholar]
- Davis, P.H. Flora of Turkey; Edinburgh University Press: Edinburgh, UK, 1965. [Google Scholar]
- Post, G.E. Flora of Syria, Palestine, and Sinai; Pипoл Kлассик: Beirut, Syria, 1932; ISBN 978-5-87410-965-3. [Google Scholar]
- Zohary, M. Geobotanical Foundations of the Middle East; Fischer: Amsterdam, The Netherlands, 1973; ISBN 90-265-0157-9. [Google Scholar]
- Fournier, P. Quatre Flores de La France; P. Lechevalier: Paris, France, 1946. [Google Scholar]
- Gimingham, C.H.; Walton, K. Environment and the Structure of Scrub Communities on the Limestone Plateaux of Northern Cyrenaica. J. Ecol. 1954, 42, 505–520. [Google Scholar] [CrossRef]
- Castroviejo, S. Flora Ibérica: Plantas Vasculares de La Península Ibérica e Islas Baleares; CSIC, Consejo Superior de Investigaciones Científicas: Madrid, Spain, 2020. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia Vol. 3, Seconda Edizione ed; Edagricole Milano: Milano, Italy, 2018. [Google Scholar]
- Meikle, R. Flora of Cyprus; Volume II; Bentham-Moxon Trust: Kew: London, UK, 1985. [Google Scholar]
- Kabiel, H.F.; Hegazy, A.K.; Lovett-Doust, L.; Al-Rowaily, S.L.; Borki, A.E.-N.E. Demography of the Threatened Endemic Shrub, Arbutus pavarii, in the Al-Akhdar Mountainous Landscape of Libya. J. For. Res. 2016, 27, 1295–1303. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.; Valentine, D.; Moore, D.; Ball, P.; Chater, A.; Walters, S.; DeFilipps, R.; Webb, D. Flora Europaea: Diapensiaceae to Myoporaceae; Cambridge University Press: Cambridge, UK, 1972; Volume 3. [Google Scholar]
- Euro+Med PlantBase Home. Available online: https://www.emplantbase.org/home.html (accessed on 15 March 2023).
- Dahl, E. The Phytogeography of Northern Europe (British Isles, Fennoscandia and Adjacent Areas).; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Santiso, X.; Lopez, L.; Retuerto, R.; Barreiro, R. Population Structure of a Widespread Species under Balancing Selection: The Case of Arbutus Unedo L. Front. Plant Sci. 2016, 6, 1264. [Google Scholar] [CrossRef]
- Skeffington, M.S.; Scott, N. Is the Strawberry Tree, Arbutus unedo (Ericaceae), Native to Ireland, or Was It Brought by the First Copper Miners? Br. Ir. Bot. 2021, 3, 385–418. [Google Scholar] [CrossRef]
- Di Domenico, F.; Lucchese, F.; Magri, D. Buxus in Europe: Late Quaternary Dynamics and Modern Vulnerability. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 354–362. [Google Scholar] [CrossRef]
- Khela, S.; Malin Rivers, D.H. IUCN Red List of Threatened Species: Arbutus unedo. Available online: https://www.iucnredlist.org/species/202930/68076133; (accessed on 30 March 2023).
- Magri, D.; Parra, I.; Di Rita, F.; Ni, J.; Shichi, K.; Worth, J.R.P. Linking Worldwide Past and Present Conifer Vulnerability. Quat. Sci. Rev. 2020, 250, 106640. [Google Scholar] [CrossRef]
- Almeida, A.M.; Martins, M.J.; Campagnolo, M.L.; Fernandez, P.; Albuquerque, T.; Gerassis, S.; Gonçalves, J.C.; Ribeiro, M.M. Prediction Scenarios of Past, Present, and Future Environmental Suitability for the Mediterranean Species Arbutus unedo L. Sci. Rep. 2022, 12, 84. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Santis, S.; Spada, F.; Magri, D. Geographic Range vs. Occurrence Records in Plant Distribution Mapping: The Case of Arbutus in the Old World. Forests 2023, 14, 1010. https://doi.org/10.3390/f14051010
De Santis S, Spada F, Magri D. Geographic Range vs. Occurrence Records in Plant Distribution Mapping: The Case of Arbutus in the Old World. Forests. 2023; 14(5):1010. https://doi.org/10.3390/f14051010
Chicago/Turabian StyleDe Santis, Simone, Francesco Spada, and Donatella Magri. 2023. "Geographic Range vs. Occurrence Records in Plant Distribution Mapping: The Case of Arbutus in the Old World" Forests 14, no. 5: 1010. https://doi.org/10.3390/f14051010
APA StyleDe Santis, S., Spada, F., & Magri, D. (2023). Geographic Range vs. Occurrence Records in Plant Distribution Mapping: The Case of Arbutus in the Old World. Forests, 14(5), 1010. https://doi.org/10.3390/f14051010