
Physiological Mechanisms of Bretschneidera sinensis Hemsl. Seed Dormancy Release and Germination

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Moisture Content
2.3. Stratification Treatment
2.4. Germination Experiment
2.5. Determination of Soluble Sugar Content
2.6. Determination of Antioxidant Enzyme Activity
2.7. Determination of Endogenous Hormone Contents
2.8. Statistical Analysis
3. Results and Analysis
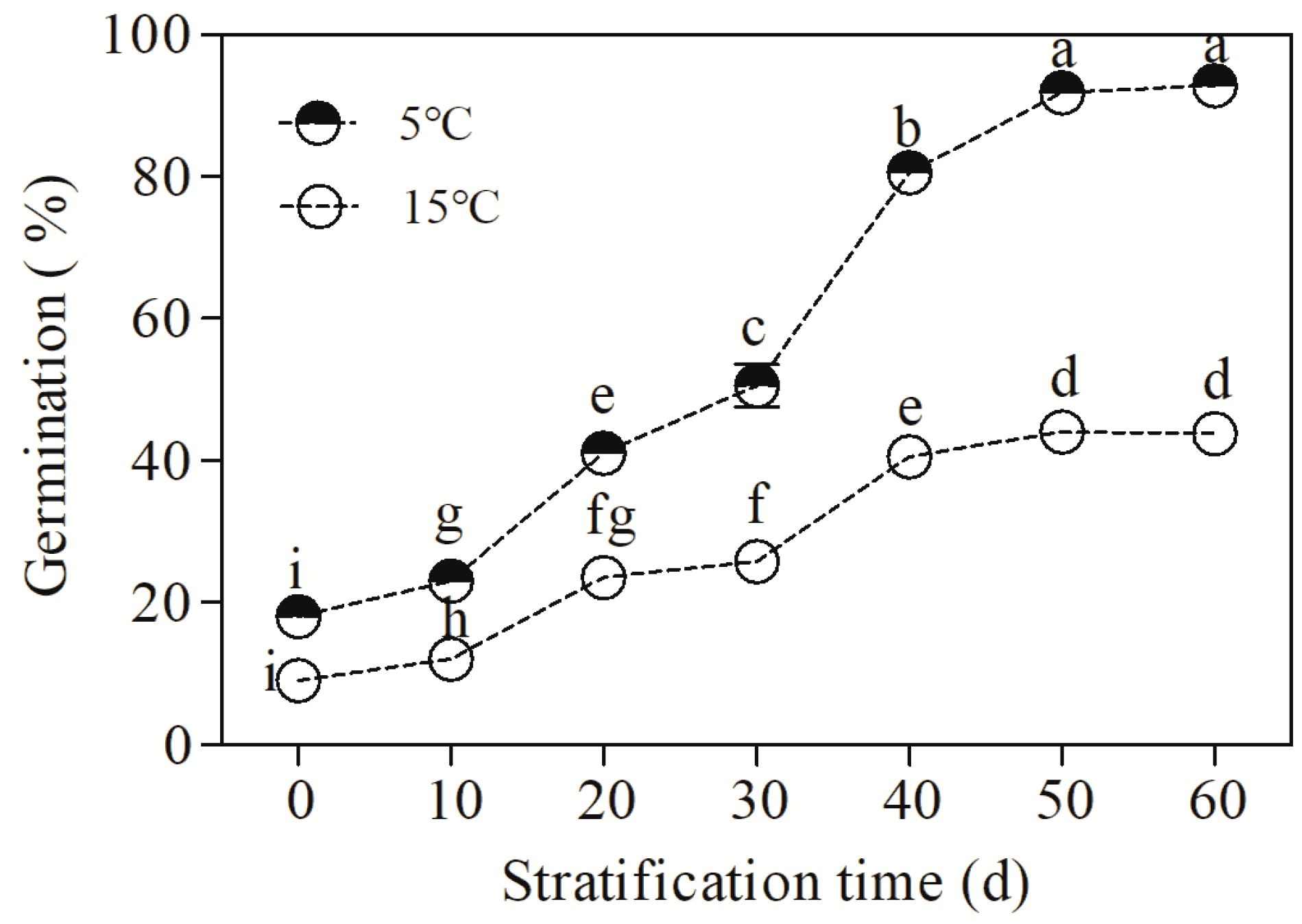
3.1. Effects of Cold Stratification on Seed Dormancy Release and Germination
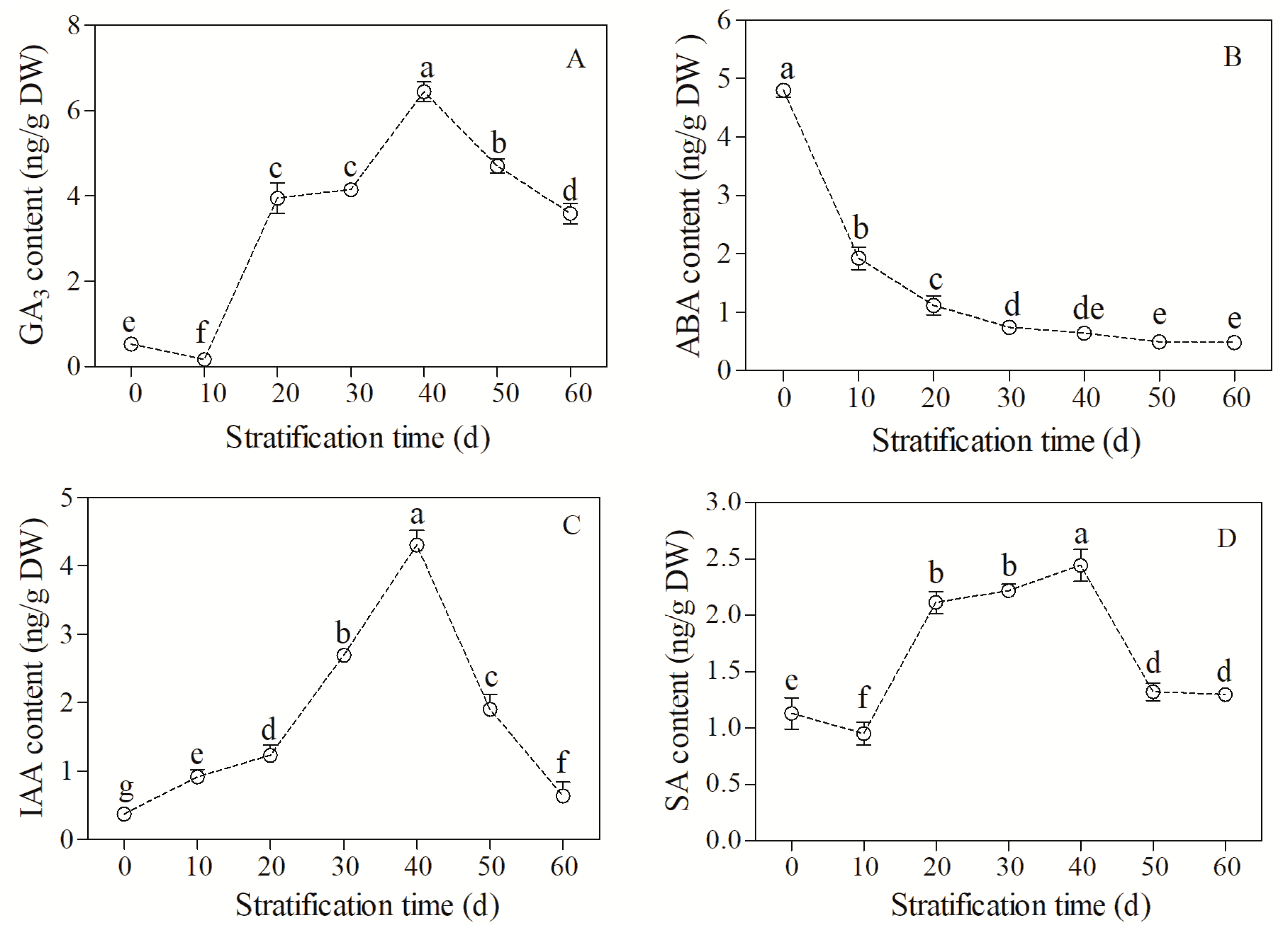
3.2. Changes in Endogenous Hormone Content during Cold Stratification
3.3. Changes in Antioxidant Enzyme Activities during Cold Stratification
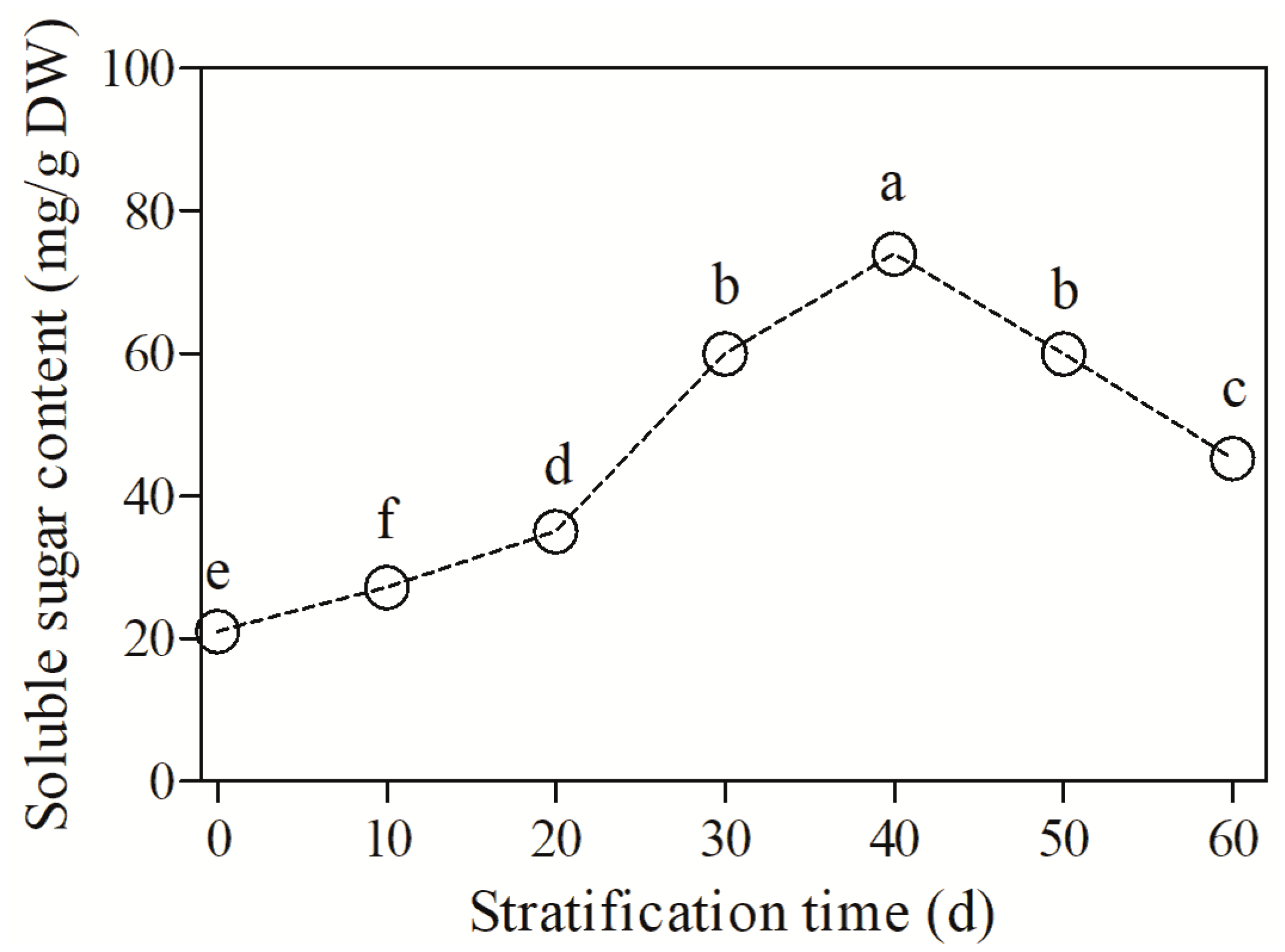
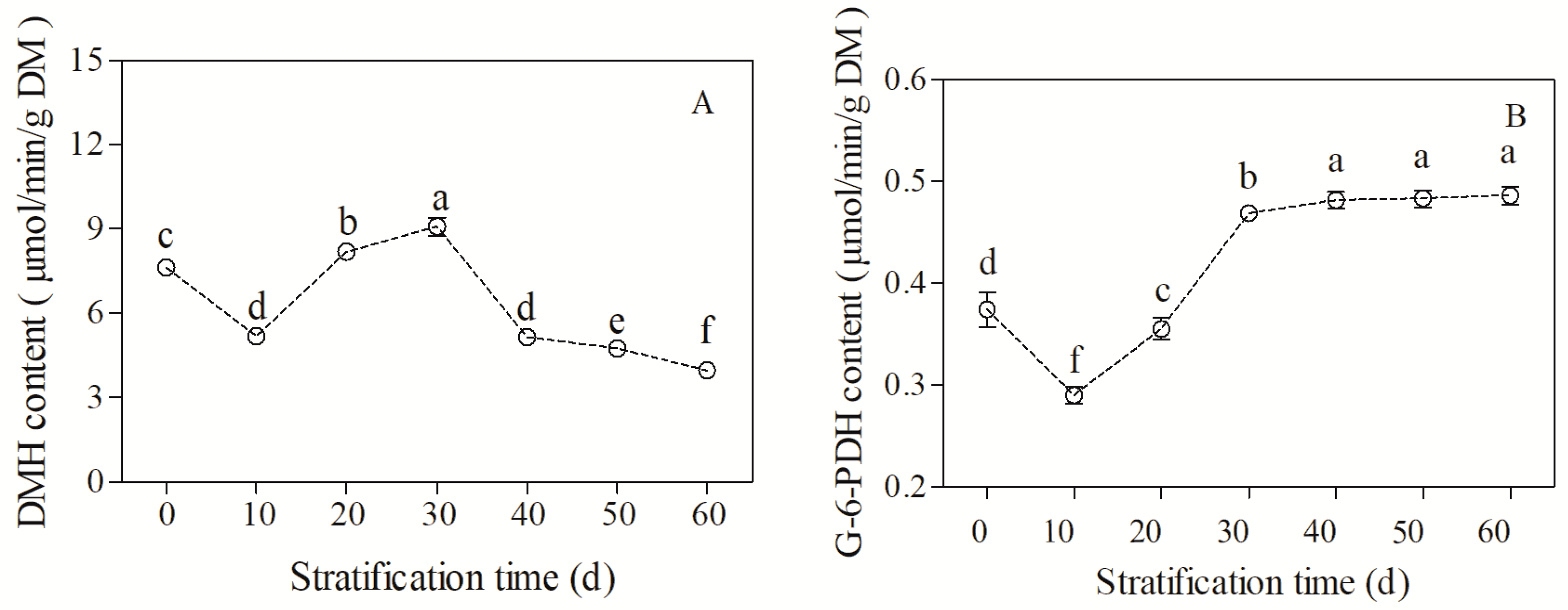
3.4. Changes in Soluble Sugar Content and Two Key Enzymes in Sugar Metabolism during Cold Stratification
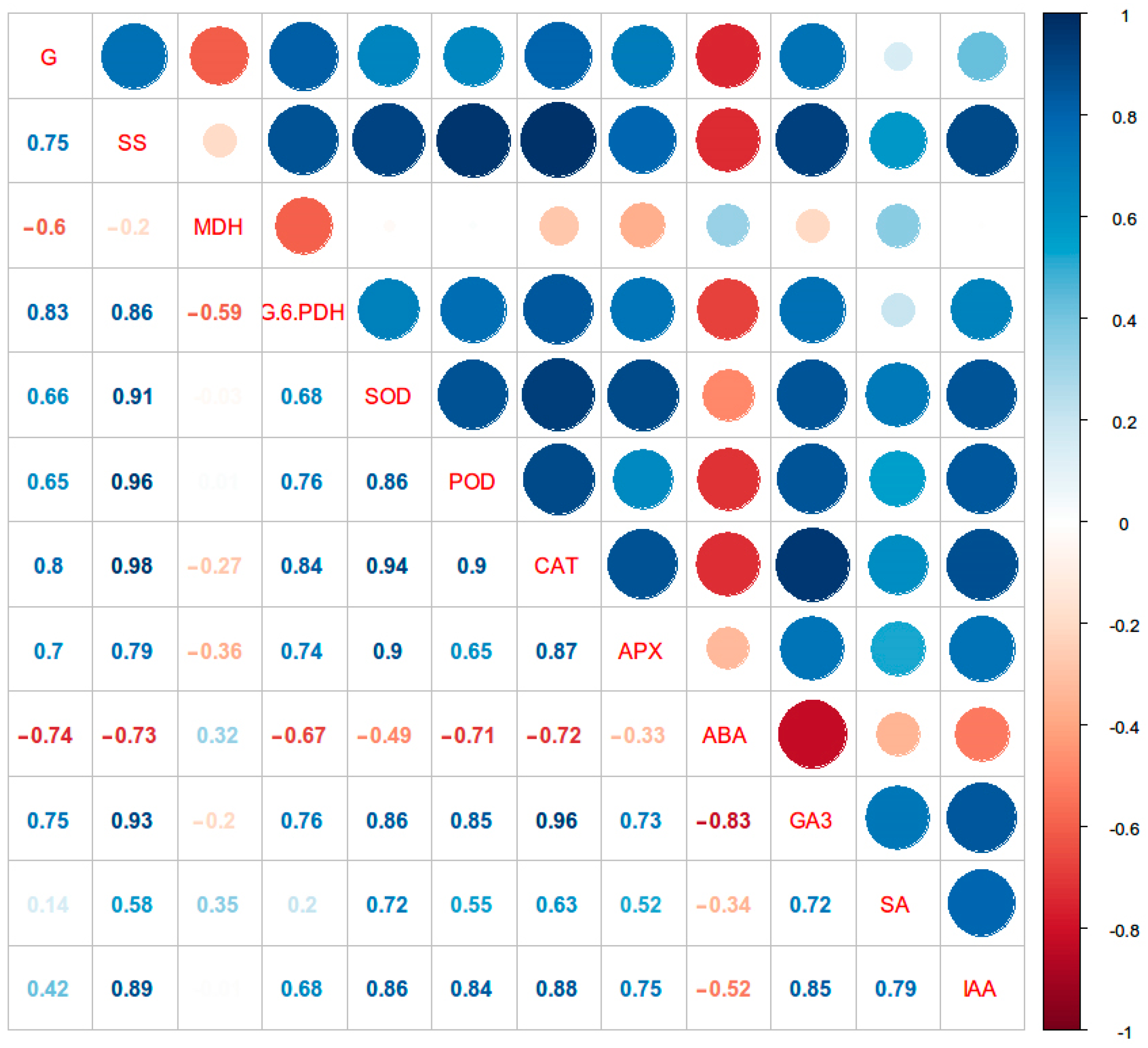
3.5. Correlations of the Germination Rate with Physiological Indicators during Cold Stratification
4. Discussion
4.1. Regulation of Seed Dormancy Release and Germination by Endogenous Hormones
4.2. Antioxidative Stress during Seed Dormancy Release and Germination
4.3. The Role of Energy Metabolism in Seed Dormancy Release and Germination
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier Academic Press Inc.: San Diego, CA, USA, 2014. [Google Scholar]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.V.; Nicotra, A.B.; Godfree, R.C.; Guja, L.K. Polyploidy affects the seed, dormancy and seedling characteristics of a perennial grass, conferring an advantage in stressful climates. Plant Biol. 2020, 22, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Rhie, Y.H.; Kim, K.S. Dormancy breaking and germination requirements of seeds of Thalictrum uchiyamae (Ranuncula ceae) with underdeveloped embryos. Sci. Hortic. 2018, 231, 82–88. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Song, S.Q.; Liu, J.; Huang, H.; Wu, J.; Xu, H.; Zhang, Q.; Li, X.; Liang, J. Gibberellin metabolism and signaling and its molecular mechanism in regulating seed germina tion and dormancy. Sci. China Life Sci. 2020, 50, 599–615. [Google Scholar]
- Song, S.Q.; Liu, J.; Xu, H.H.; Liu, X.; Huang, H. ABA metabolism and signaling and their molecular mechanism regulating seed dormancy and germination. Sci. Agric. Sin. 2020, 50, 599–615. [Google Scholar]
- Song, S.Q.; Liu, J.; Tang, C.F.; Zhang, W.H.; Xu, H.H.; Zhang, Q.; Gao, J.D. Metabolism and signaling of auxins and their roles in regulating seed dormancy and germination. Chin. Sci. Bull. 2020, 65, 3924–3943. (In Chinese) [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.; Xiang, Y.; Tong, X.; Wojtyla, Ł.; Wang, Y. Editorial: Molecular basis of seed germination and dormancy. Front. Plant Sci. 2023, 14, 1242428. [Google Scholar] [CrossRef]
- Ali, F.; Qanmber, G.; Li, F.; Wang, Z. Updated role of ABA in seed maturation, dormancy, and germination. J. Adv. Res. 2022, 35, 199–214. [Google Scholar] [CrossRef]
- Sano, N.; Marion-Poll, A. ABA Metabolism and Homeostasis in Seed Dormancy and Germination. Int. J. Mol. Sci. 2021, 22, 5069. [Google Scholar] [CrossRef]
- Longo, C.; Holness, S.; De Angelis, V.; Lepri, A.; Occhigrossi, S.; Ruta, V.; Vittorioso, P. From the Outside to the Inside: New Insights on the Main Factors That Guide Seed Dormancy and Germination. Genes 2020, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Bicalho, E.M.; Pintó-Marijuan, M.; Morales, M.; Müller, M.; Munné-Bosch, S.; Garcia, Q.S. Control of macaw palm seed germination by the gibberellin/abscisic acid balance. Plant Biol. 2015, 17, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Footitt, S.; Clewes, R.; Feeney, M.; Finch-Savage, W.E.; Frigerio, L. Aquaporins Influence seed dormancy and germination in response to stress. Plant Cell Environ. 2019, 42, 2325–2339. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Q.; Yang, L.; Li, X.; Wang, Z.; Zhang, Y. Changes in carbohydrate metabolism and endogenous hormone regulation during bulblet initiation and development in Lycoris radiata. BMC Plant Biol. 2020, 20, 180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, F.; Li, X.; Cao, H.; Ding, M.; Zhang, C.; Zuo, J.; Xu, C.; Xu, J.; Deng, X.; et al. Arabidopsis seed germination speed is controlled by SNL histone deacetylase-binding factor-mediated regulation of AUX1. Nat. Commun. 2016, 7, 13412. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Kumari, A.; Thakur, M.; Koul, A. Hydrogen peroxide signaling integrates with phytohormones during the germination of magnetoprimed tomato seeds. Sci. Rep. 2019, 9, 8814. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Hao, W.; Liu, D.; Wang, L.; Qiu, S. Effects of SA on the germination of seeds and the stability of cell membrane under cold stress. J. Northwest AF Univ. (Nat. Sci. Ed.) 2010, 38, 183–188. [Google Scholar]
- Wang, Y.M.; Wang, L.J.; Yao, B.; Liu, Z.; Li, F. Changes in ABA, IAA, GA3, and ZR levels during seed dormancy release in Idesia polycarpa Maxim from Jiyuan. Pol. J. Environ. Stud. 2018, 27, 1833–1839. [Google Scholar]
- Katsuya-Gaviria, K.; Caro, E.; Carrillo-Barral, N.; Iglesias-Fernández, R. Reactive Oxygen Species (ROS) and Nucleic Acid Modifications during Seed Dormancy. Plants 2020, 9, 679. [Google Scholar] [CrossRef]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of reactive oxygen species in the regulation of Arabidopsis seed dormancy. Plant Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Dai, Y.; Zheng, C.; Yang, Y.; Chen, W.; Wang, Q.; Chandrasekaran, U.; Du, J.; Liu, W.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Wang, W.B.; Kim, Y.H.; Lee, H.S.; Kim, K.Y.; Deng, X.P.; Kwak, S.S. Analysis of antioxidant enzyme activity during Germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 2009, 47, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; El-Maarouf-Bouteau, H.; Kranner, I.; Bogatek, R.; Corbineau, F.; Bailly, C. The mechanisms involved in seed dormancy alleviation by hydrogen cyanide unravel the role of reactive oxygen species as key factors of cellular signaling during germination. Plant Physiol. 2009, 150, 494–505. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 31, 75–182. [Google Scholar] [CrossRef]
- Zaynab, M.; Pan, D.; Chen, W. Transcriptomic approach to address low germination rate in Cyclobalnopsis gilva seeds. S. Afr. J. Bot. 2018, 119, 286–294. [Google Scholar] [CrossRef]
- Roberts, E. Temperature and seed germination. Symp. Soc. Exp. Biol. 1988, 42, 109–132. [Google Scholar] [PubMed]
- Zhi, L.M.; Zhang, Y.H.; Yu, F.Y. Biochemical and physiological changes of Euscaphis japonica seeds during the period of stratifi. J. Cent. South Univ. For. Technol. 2016, 36, 36–40. [Google Scholar]
- Wu, Z.Y.; Lu, A.M.; Tang, Y.C. The Families and General of Angiosperm in China; Science Press: Beijing, China, 2003; p. 702. [Google Scholar]
- Zhang, J.R.; Cheng, G.F. National level to protect plants-bretschneidera sinensis Hemsl. Bull. Biol. 2009, 44, 7. [Google Scholar]
- Li, T.H.; Zhou, Y.X.; Duan, X.P. A preliminary study of physiology dormancy character bretschneidera sinensis Hemsl Seeds. J. Cent. South For. Univ. 1997, 17, 41–44. [Google Scholar]
- Li, X.Z.; Simpson, W.R.; Song, M.L.; Bao, G.S.; Niu, X.L.; Zhang, Z.H.; Xu, H.F.; Liu, X.; Li, Y.L.; Li, C.J. Effects of seed moisture content and Epichloe endophyte on germination and physiology of Achnatherum inebrians. S. Afr. J. Bot. 2022, 134, 407–414. [Google Scholar] [CrossRef]
- Ma, L.Y.; Cheng, N.L.; Han, G.J.; Li, L. Effects of exogenous salicylic acid on seed germination and physiological characteristics of Coronilla varia under drought stress. Chin. J. Appl. Ecol. 2017, 28, 3274–3280. [Google Scholar]
- Wu, M.J.; Wu, J.Y.; Gan, Y.B. The new insight of auxin functions: Transition from seed dormancy to germination and floral opening in plants. Plant Growth Regul. 2020, 91, 169–174. [Google Scholar] [CrossRef]
- Boter, M.; Calleja-Cabrera, J.; Carrera-Castaño, G.; Wagner, G.; Hatzig, S.V.; Snowdon, R.J.; Legoahec, L.; Bianchetti, G.; Bouchereau, A.; Nesi, N.; et al. An integrative approach to analyze seed germination in Brassica napus. Front. Plant Sci. 2019, 10, 1342. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Su, J.; Liu, W.; Li, S. Dormancy release and germination of Taxus yunnanensis seeds during wet sand storage. Sci. Rep. 2018, 8, 3205. [Google Scholar] [CrossRef]
- Lee, S.; Kim, S.G.; Park, C.M. Salicylic acid promotes seed germination under high salinity by modulating antioxidant activity in Arabidopsis. New Phytol. 2010, 188, 626–637. [Google Scholar] [CrossRef]
- Zhang, J.J.; Wei, X.; Chai, S.F.; Wu, S.H.; Zou, R.; Qin, X.M.; Fu, R. Dormancy mechanism of the seeds of a rare and endangered plant, Garcinia paucinervis. Chin. J. Ecol. 2018, 37, 1371–1381. [Google Scholar]
- Su, H.L.; Zhou, X.Z.; Li, X. Physicochemical changes of Paris polyphylla var. Chinensis seed during different stages of germination. Chin. Tradit. Herb. Drugs 2017, 48, 4755–4763. [Google Scholar]
- Pluskota, W.E.; Pupel, P.; Głowacka, K.; Okorska, S.B.; Jerzmanowski, A.; Nonogaki, H.; Górecki, R.J. Jasmonic acid and ethylene are involved in the accumulation of osmotin in germinating tomato seeds. J. Plant Physiol. 2018, 232, 74–81. [Google Scholar] [CrossRef]
- Xu, L.; Wang, P.; Ali, B.; Yang, N.; Chen, Y.; Wu, F.; Xu, X. Changes of the phenolic compounds and antioxidant activities in germinated adlay seeds. J. Sci. Food Agric. 2017, 97, 4227–4234. [Google Scholar] [CrossRef]
- Oracz, K.; Bouteau, H.E.; Farrant, J.M.; Cooper, K.; Belghazi, M.; Job, C.; Job, D.; Corbineau, F.; Bailly, C. ROS production and protein oxidation as a novel mechanism for seed dormancy alleviation. Plant J. 2007, 50, 452–465. [Google Scholar] [CrossRef]
- Huang, W.; Mayton, H.S.; Amirkhani, M.; Wang, D.; Taylor, A.G. Seed dormancy,germination and fungal infestation of eastern gamagrass seed. Ind. Crops Prod. 2017, 99, 109–116. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, B.; Xu, Z.; Shi, Z.; Chen, S.; Huang, X.; Chen, J.; Wang, X. Involvement of reactive oxygen species in endosperm cap weakening and embryo elongation growth during lettuce seed germination. J. Exp. Bot. 2014, 65, 3189–3200. [Google Scholar] [CrossRef]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef]
- Amooaghaie, R. Triangular interplay between ROS, ABA and GA in dormancy alleviation of Bunium persicum seeds by cold stratification. Russ. J. Plant Physiol. 2017, 64, 588–599. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.X.; Xu, X. The activity of Principal antioxidant enzymes and the content of metabolites in dormancy breaking and germination of Davidia involucrate seeds. Plant Divers. Resour. 2015, 37, 779–787. [Google Scholar]
- Marta, B.; Szafrańska, K.; Posmyk, M. Exogenous melatonin improves antioxidant defense in cucumber seeds (Cucumis sativus L.) germinated under chilling stress. Front. Plant Sci. 2016, 7, 575. [Google Scholar] [CrossRef]
- Kazmi, R.H.; Willems, L.A.J.; Joosen, R.V.L.; Khan, N.; Ligterink, W.; Hilhorst, H.W.M. Metabolomic analysis of tomato seed germination. Metabolomics 2017, 13, 145. [Google Scholar] [CrossRef]
- Alencar, N.L.; Innecco, R.; Gomes-Filho, E.; Gallão, M.I.; Alvarez-Pizarro, J.C.; Prisco, J.T.; De Oliveira, A.B. Seed reserve composition and mobilization during germination and early seedling establishment of Cereus jamacaru D.C. ssp. jamacaru (Cactaceae). An. Acad. Bras. Ciênc. 2012, 84, 823–832. [Google Scholar] [CrossRef]
- Han, C.; Zhen, S.; Zhu, G.; Bian, Y.; Yan, Y. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination. Plant Physiol. Biochem. 2017, 115, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Sang, S.; Chen, Y.; Wei, Z.; Wang, P. The role of Arabidopsis inositol polyphosphate kinase AtIPK2β in glucose suppression of seed germination and seedling development. Plant Cell Physiol. 2017, 59, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, H.; Yan, H.; Qiu, L.; Baskin, C.C. Mobilization and role of starch,protein, and fat reserves during seed germination of six wild grassland species. Front. Plant Sci. 2018, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sakshi, G.; Munshi, S. Changes in lipid and carbohydrate composition of germinating soybean seeds under different storage conditions. Asian J. Plant Sci. 2007, 6, 596. [Google Scholar]
- Roberts, E.H.; Major, W. Dormancy in cereal seeds: II. The nature of gaseous exchange in imbibed barley and rice seeds. J. Exp. Bot. 1968, 19, 90–101. [Google Scholar]
- Li, Z.L.; Tong, K.; Yan, S.; Yang, H.; Wang, Q.; Tang, Y.B.; Deng, M.S.; Tian, M.L. Physiological and biochemical change of Paris seed in after-ripening during variable temperature stratifications. China J. Chin. Mater. Med. 2015, 40, 629–633. [Google Scholar]
- Smolikova, G.; Leonova, T.; Vashurina, N.; Frolov, A.; Medvedev, S. Desiccation Tolerance as the Basis of Long-Term Seed Viability. Int. J. Mol. Sci. 2021, 22, 101. [Google Scholar] [CrossRef]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef]
- Couée, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of soluble sugars in reactive oxygen species balance and responses to oxide tive stress in plants. J. Exp. Bot. 2006, 57, 449–459. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, L.; Dong, H.; Deng, Z.; Li, J.; Xu, L.; Mou, J.; Deng, S. Physiological Mechanisms of Bretschneidera sinensis Hemsl. Seed Dormancy Release and Germination. Forests 2023, 14, 2430. https://doi.org/10.3390/f14122430
Zhong L, Dong H, Deng Z, Li J, Xu L, Mou J, Deng S. Physiological Mechanisms of Bretschneidera sinensis Hemsl. Seed Dormancy Release and Germination. Forests. 2023; 14(12):2430. https://doi.org/10.3390/f14122430
Chicago/Turabian StyleZhong, Lijun, Hongxing Dong, Zhijun Deng, Jitao Li, Li Xu, Jiaolin Mou, and Shiming Deng. 2023. "Physiological Mechanisms of Bretschneidera sinensis Hemsl. Seed Dormancy Release and Germination" Forests 14, no. 12: 2430. https://doi.org/10.3390/f14122430
APA StyleZhong, L., Dong, H., Deng, Z., Li, J., Xu, L., Mou, J., & Deng, S. (2023). Physiological Mechanisms of Bretschneidera sinensis Hemsl. Seed Dormancy Release and Germination. Forests, 14(12), 2430. https://doi.org/10.3390/f14122430