


Plant Species Richness and Invasional Meltdown in Different Parts of Acer negundo L. Secondary Range

Abstract
1. Introduction
2. Materials and Methods
2.1. Model Invasive Woody Species—Acer negundo
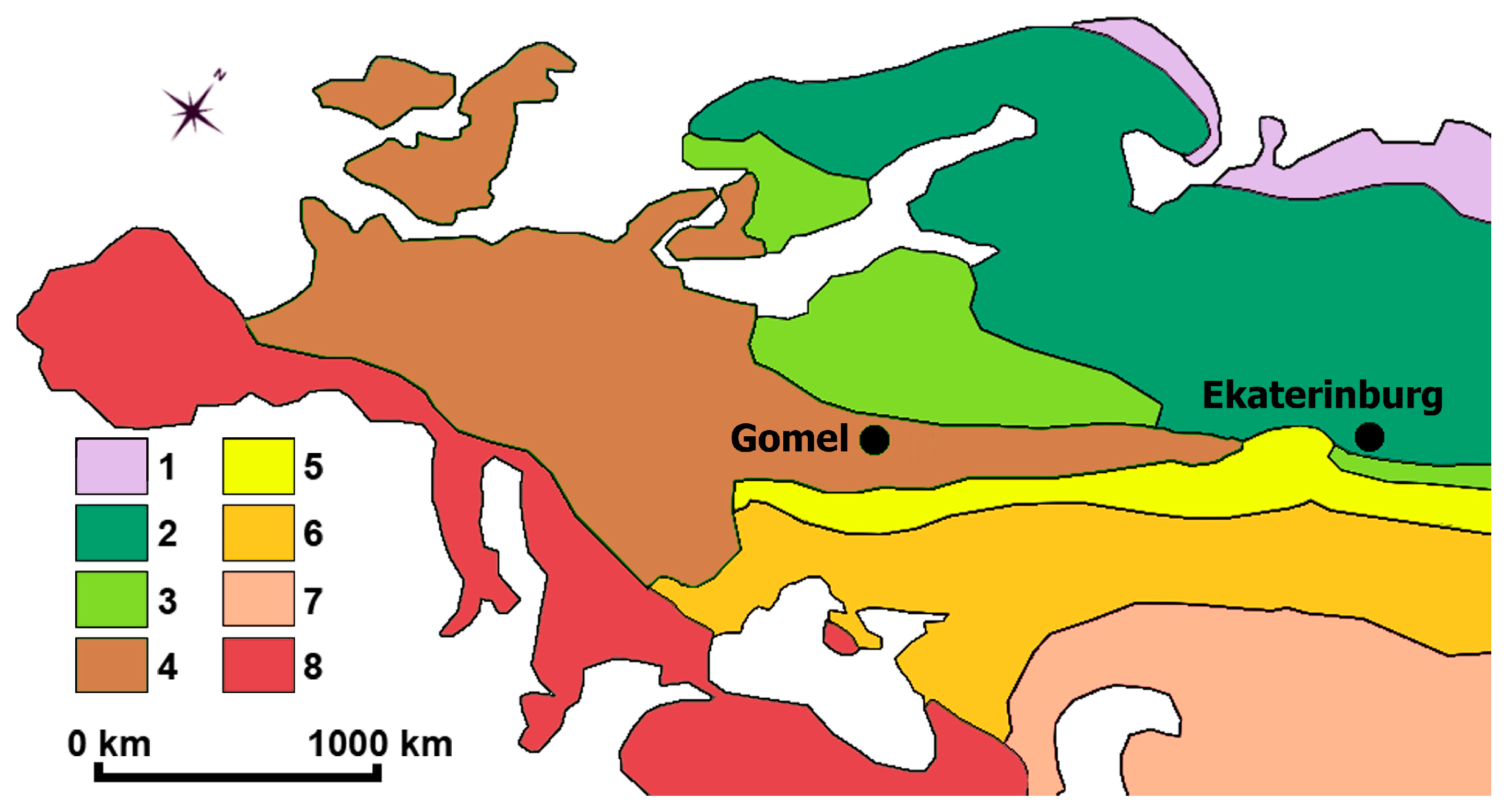
2.2. Study Regions
2.3. Sample Plots
2.4. Vegetation Descriptions
2.5. Data Analysis
3. Results
3.1. Community Composition and Species Occurrence
3.1.1. The Belarusian Polesia
3.1.2. The Middle Urals
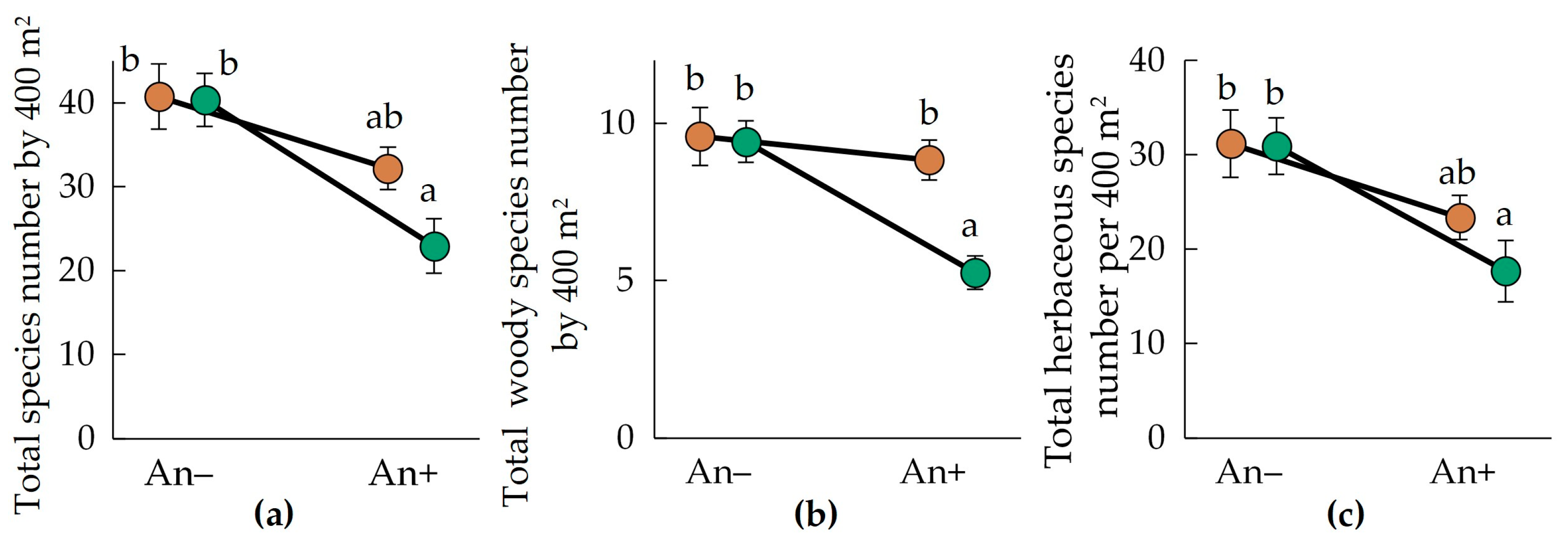
3.2. Species Richness
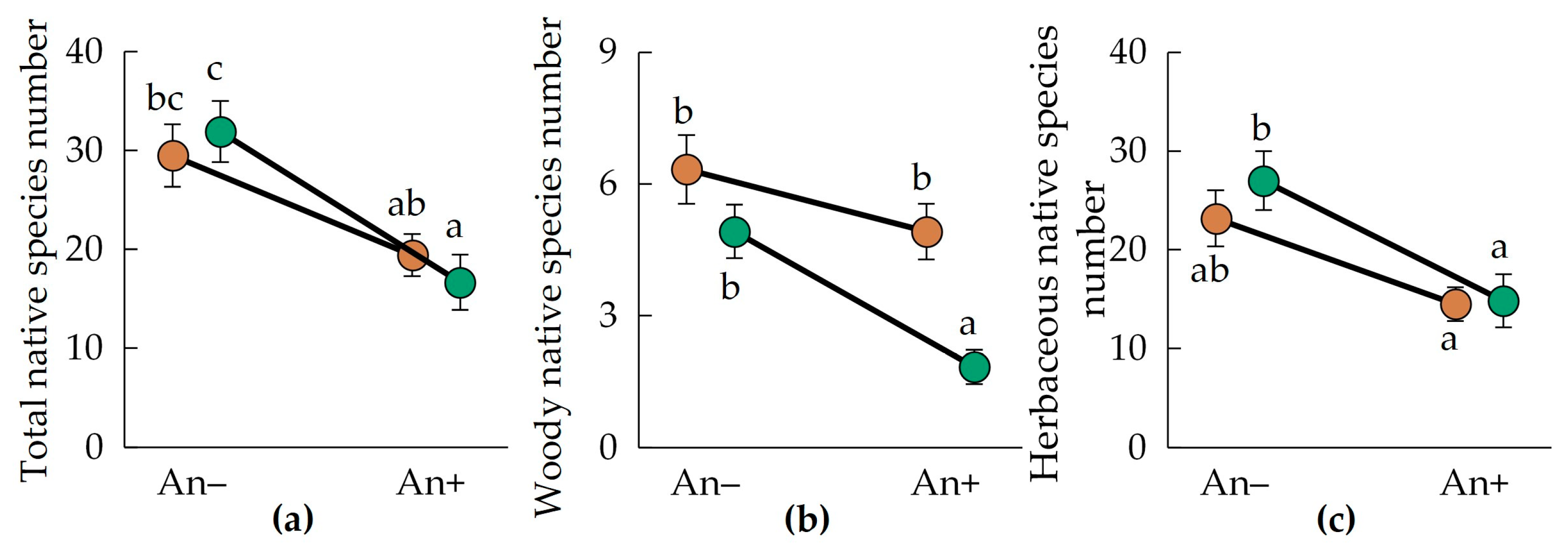
3.3. Native Species Richness
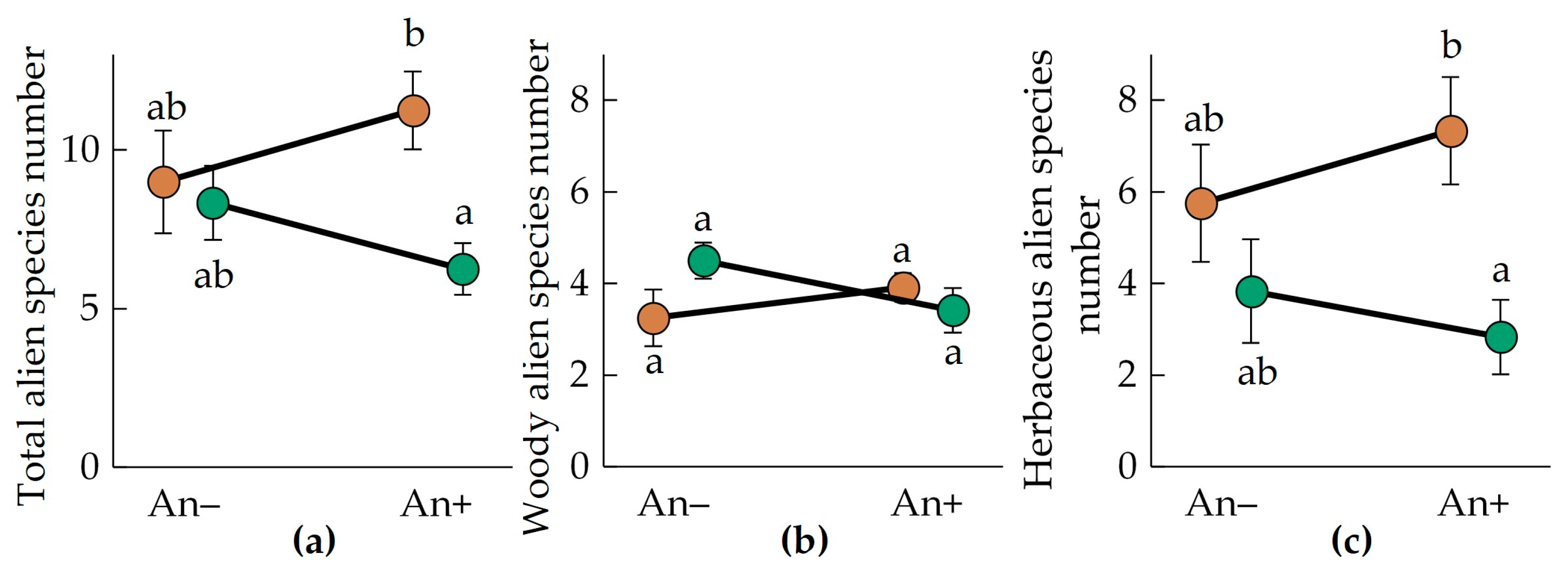
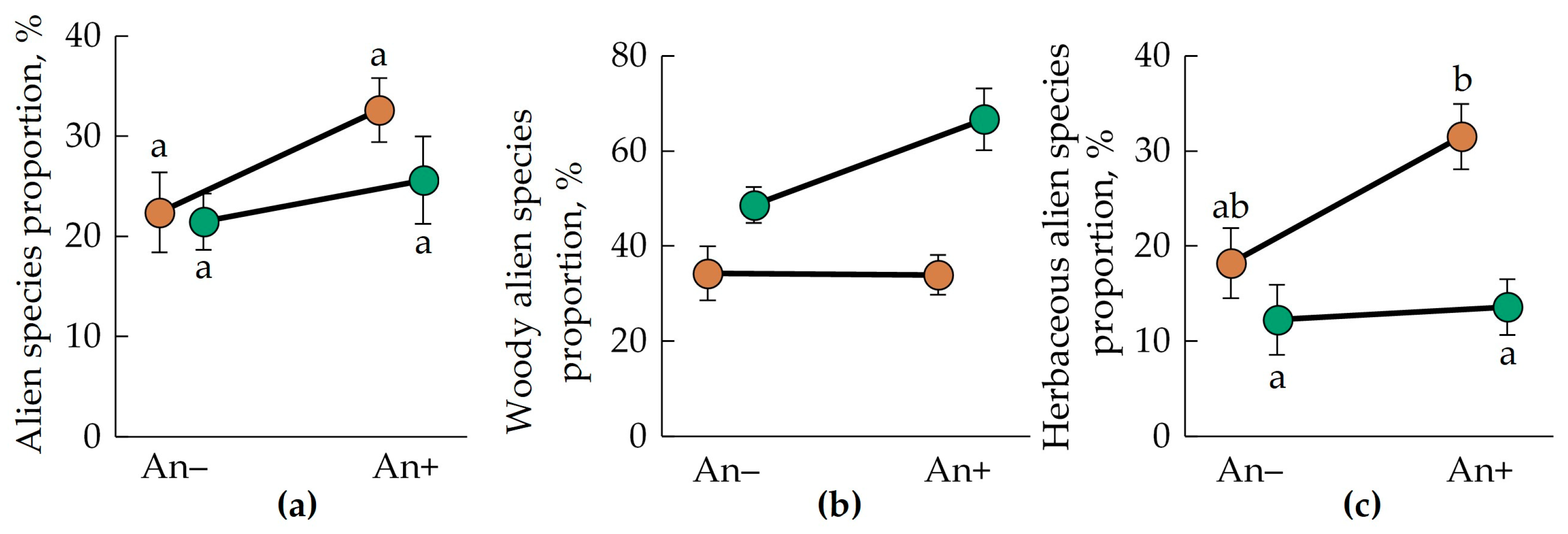
3.4. Alien Species Richness and Proportion
4. Discussion
- The plant species number in the communities invaded by A. negundo in both study regions decreased compared to the non-invaded communities;
- Apparently, this is primarily related to a decrease in the number of native woody and herbaceous species in the A. negundo-invaded communities;
- The alien species proportion is greater in the Belarusian Polesia than in the Middle Urals.
4.1. Acer Negundo Invasion-Induced Species Richness Decrease
4.2. Native-Alien Species Richness Relationship
4.3. Levels of Invasion in Different Regions
5. Conclusions
- In the regions with different geographic and floristic conditions (the Belarusian Polesia and the Middle Urals), plant species richness in the communities invaded by Acer negundo decreased compared to the non-invaded communities. The differences in species richness between the invaded and non-invaded communities in different regions are not equal but have the same directions. This result shows that the negative impact of A. negundo on community species richness is similar on an inter-regional scale, i.e., in different parts of its secondary range.
- In different parts of A. negundo secondary range, its invasion primarily caused a decrease in native plant species richness. This phenomenon may be interpreted as evidence of the presence of weak effects predicted by the invasional meltdown hypothesis in invaded communities. We believe that this result is new in relation to the invasion of A. negundo. This result indicates that A. negundo may have selective impacts on native and alien plants.
- Average herbaceous plant invasion levels in the communities of the Belarusian Polesia are greater than in the communities of the Middle Urals. But the average levels of invasion of alien woody plants in the communities of the Belarusian Polesia are lower than in the communities of the Middle Urals. This result highlights the importance of climatic and geographical conditions, land use history, and disturbance regimes in different regions for the distribution and sustainability of alien species in local communities.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Gaertner, M.; Breeyen, D.A.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: A meta-analysis. Prog. Phys. Geogr. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Powell, K.I.; Chase, J.M.; Knight, T.M. A synthesis of plant invasion effects on biodiversity across spatial scales. Am. J. Bot. 2011, 98, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Vila, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarosik, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Hejda, M.; Sádlo, J.; Kutlvašr, J.; Petřík, P.; Vítková, M.; Vojík, M.; Pyšek, P.; Pergl, J. Impact of invasive and native dominants on species richness and diversity of plant communities. Preslia 2021, 93, 181–201. [Google Scholar] [CrossRef]
- Pysek, P.; Jarosik, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vila, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Gilbert, B.; Levine, J.M. Plant invasions and the niche. J. Ecol. 2009, 97, 609–615. [Google Scholar] [CrossRef]
- Perkins, L.B.; Nowak, R.S. Invasion syndromes: Hypotheses on relationships among invasive species attributes and characteristics of invaded sites. J. Arid Land 2013, 5, 275–283. [Google Scholar] [CrossRef][Green Version]
- Gioria, M.; Osborne, B.A. Resource competition in plant invasions: Emerging patterns and research needs. Front. Plant Sci. 2014, 5, 5. [Google Scholar] [CrossRef]
- Gioria, M.; Hulme, P.E.; Richardson, D.M.; Pyšek, P. Why Are Invasive Plants Successful? Annu. Rev. Plant Biol. 2023, 74, 635–670. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Gurnee, J.; Tirado, R.; Callaway, R.M. Invasion through quantitative effects: Intense shade drives native decline and invasive success. Ecol. Appl. A Publ. Ecol. Soc. Am. 2006, 16, 1821–1831. [Google Scholar] [CrossRef]
- Nilsson, C.; Engelmark, O.; Cory, J.; Forsslund, A.; Carlborg, E. Differences in litter cover and understory flora between stands of introduced lodgepole pine and native Scots pine in Sweden. For. Ecol. Manag. 2008, 255, 1900–1905. [Google Scholar] [CrossRef]
- Cusack, D.F.; McCleery, T.L. Patterns in understory woody diversity and soil nitrogen across native- and non-native-urban tropical forests. For. Ecol. Manag. 2014, 318, 34–43. [Google Scholar] [CrossRef]
- Berg, C.; Drescherl, A.; Essl, F. Using relevé-based metrics to explain invasion patterns of alien trees in temperate forests. Tuexenia 2017, 37, 127–142. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Dubrovin, D.I.; Pustovalova, L.A. High canopy cover of invasive Acer negundo L. affects ground vegetation taxonomic richness. Sci. Rep. 2021, 11, 20758. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Dubrovin, D.I.; Rafikova, O.S.; Lipikhina, Y.A.; Zolotareva, N.V.; Podgaevskaya, E.N.; Pustovalova, L.A.; Yakovleva, A.V. Shading and light interception in thickets of invasive Acer negundo and Sorbaria sorbifolia. Russ. J. Biol. Invasions 2022, 4, 30–42. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Uddin, M.N.; Asaeda, T.; Shampa, S.H.; Robinson, R.W. Allelopathy and its coevolutionary implications between native and non-native neighbors of invasive Cynara cardunculus L. Ecol. Evol. 2020, 10, 7463–7475. [Google Scholar] [CrossRef]
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Simberloff, D. Invasional meltdown 6 years later: Important phenomenon, unfortunate metaphor, or both? Ecol. Lett. 2006, 9, 912–919. [Google Scholar] [CrossRef]
- Rojas-Sandoval, J.; Ackerman, J.D.; Marcano-Vega, H.; Willig, M.R. Alien Species Affect the Abundance and Richness of Native Species in Tropical Forests: The Role of Adaptive Strategies. Ecosphere 2022, 13, e4291. [Google Scholar] [CrossRef]
- Ricciardi, A. Facilitative interactions among aquatic invaders: Is an “invasional meltdown” occurring in the Great Lakes? Can. J. Fish. Aquat. Sci. 2001, 58, 2513–2525. [Google Scholar] [CrossRef]
- Relva, M.A.; Nunez, M.A.; Simberloff, D. Introduced deer reduce native plant cover and facilitate invasion of non-native tree species: Evidence for invasional meltdown. Biol. Invasions 2010, 12, 303–311. [Google Scholar] [CrossRef]
- Montgomery, W.I.; Lundy, M.G.; Reid, N. ‘Invasional meltdown’: Evidence for unexpected consequences and cumulative impacts of multispecies invasions. Biol. Invasions 2012, 14, 1111–1125. [Google Scholar] [CrossRef]
- Von Holle, B. Invasional meltdown. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmánek, M., Eds.; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 2011; pp. 360–364. [Google Scholar]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Stotz, G.C.; Cahill, J.F.; Bennett, J.A.; Carlyle, C.N.; Bork, E.W.; Askarizadeh, D.; Bartha, S.; Beierkuhnlein, C.; Boldgiv, B.; Brown, L.; et al. Not a melting pot: Plant species aggregate in their non-native range. Glob. Ecol. Biogeogr. 2020, 29, 482–490. [Google Scholar] [CrossRef]
- Bennett, J.A.; Riibak, K.; Kook, E.; Reier, Ü.; Tamme, R.; Guillermo-Bueno, C.; Pärtel, M. Species pools, community completeness and invasion: Disentangling diversity effects on the establishment of native and alien species. Ecol. Lett. 2016, 19, 1496–1505. [Google Scholar] [CrossRef]
- Pärtel, M.; Zobel, M.; Zobel, K.; van der Maarel, E. The species pool and its relation to species richness: Evidence from Estonian plant communities. Oikos 1996, 75, 111–117. [Google Scholar] [CrossRef]
- Pärtel, M.; Bennett, J.A.; Zobel, M. Macroecology of biodiversity: Disentangling local and regional effects. New Phytol. 2016, 211, 404–410. [Google Scholar] [CrossRef]
- Cornell, H.V.; Karlson, R.H. Local and Regional Processes as Controls of Species Richness. In Spatial Ecology: The Role of Space in Population Dynamics and Interspecific Interactions (MPB-30); Tilman, D., Karevia, P., Eds.; Princeton University Press: Princeton, NJ, USA, 1997; pp. 250–268. [Google Scholar] [CrossRef]
- Zobel, M. The species pool concept as a framework for studying patterns of plant diversity. J. Veg. Sci. 2016, 27, 8–18. [Google Scholar] [CrossRef]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, I.; Petri, L.; Barnett, D.T.; Beaury, E.M.; Blumenthal, D.M.; Corbin, J.D.; Diez, J.; Dukes, J.S.; Early, R.; Pearse, I.S.; et al. Combining local, landscape, and regional geographies to assess plant community vulnerability to invasion impact. Ecol. Appl. 2023, 33, e2821. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, C.; Ma, L.; Qiang, S.; Silander, J.A.; Qi, L.L. Biotic Homogenization Caused by the Invasion of Solidago canadensis in China. J. Integr. Agric. 2013, 12, 835–845. [Google Scholar] [CrossRef]
- Hejda, M.; Štajerová, K.; Pyšek, P. Dominance has a biogeographical component: Do plants tend to exert stronger impacts in their invaded rather than native range? J. Biogeogr. 2017, 44, 18–27. [Google Scholar] [CrossRef]
- Gazol, A.; Zobel, M.; Cantero, J.J.; Davison, J.; Esler, K.J.; Jairus, T.; Öpik, M.; Vasar, M.; Moora, M. Impact of alien pines on local arbuscular mycorrhizal fungal communities—Evidence from two continents. FEMS Microbiol. Ecol. 2016, 92, fiw073. [Google Scholar] [CrossRef]
- Gusev, A.P.; Shpilevskaya, N.S.; Veselkin, D.V. The influence of Acer negundo L. on progressive successions in Belarusian landscapes. Vestn. Vitebsk. Gos. Univ. 2017, 94, 47–53. (In Russian) [Google Scholar]
- Veselkin, D.V.; Dubrovin, D.I. Diversity of the grass layer of urbanized communities dominated by invasive Acer negundo. Rus. J. Ecol. 2019, 50, 413–421. [Google Scholar] [CrossRef]
- Information Retrieval System of the Central Botanical Garden of the National Academy of Sciences of Belarus. Available online: http://hbc.bas-net.by (accessed on 14 August 2023). (In Russian).
- Knyazev, M.S.; Zolotareva, N.V.; Podgaevskaya, E.N.; Tretyakova, A.S.; Kulikov, P.V. An annotated check list of the flora of Sverdlovsk region. Part I: Spore and gymnosperms plants. Phytodivers. East. Eur. 2016, 10, 11–41. (In Russian) [Google Scholar]
- Knyazev, M.S.; Tretyakova, A.S.; Podgaevskaya, E.N.; Zolotareva, N.V.; Kulikov, P.V. An annotated checklist of the flora of Sverdlovsk Region. Part II: Monocotyledonous plants. Phytodivers. East. Eur. 2017, 11, 4–108. (In Russian) [Google Scholar]
- Knyazev, M.S.; Tretyakova, A.S.; Podgaevskaya, E.N.; Zolotareva, N.V.; Kulikov, P.V. An annotated checklist of the flora of Sverdlovsk Region. Part III: Dicotyledonous plants (Aristolochiaceae–Monotropaceae). Phytodivers. East. Eur. 2018, 12, 4–95. (In Russian) [Google Scholar] [CrossRef]
- Knyazev, M.S.; Tretyakova, A.S.; Podgaevskaya, E.N.; Zolotareva, N.V.; Kulikov, P.V. Annotated checklist of the flora of Sverdlovsk region. Part IV: Dicotyledonous plants (Empetraceae–Droseraceae). Phytodivers. East. Eur. 2019, 13, 130–196. (In Russian) [Google Scholar] [CrossRef]
- Knyazev, M.S.; Chkalov, A.V.; Tretyakova, A.S.; Zolotareva, N.V.; Podgaevskaya, E.N.; Pakina, D.V.; Kulikov, P.V. Annotated checklist of the flora of Sverdlovsk region. Part V: Dicotyledonous plants (Rosaceae). Phytodivers. East. Eur. 2019, 13, 305–352. (In Russian) [Google Scholar] [CrossRef]
- Knyazev, M.S.; Podgaevskaya, E.N.; Tretyakova, A.S.; Zolotareva, N.V.; Kulikov, P.V. Annotated checklist of the flora of Sverdlovsk Region. Part VI: Dicotyledonous plants (Fabaceae–Lobeliaceae). Phytodivers. East. Eur. 2020, 14, 190–331. (In Russian) [Google Scholar] [CrossRef]
- Knyazev, M.S.; Podgaevskaya, E.N.; Zolotareva, N.V.; Tretyakova, A.S.; Kulikov, P.V. Annotated checklist of the flora of Sverdlovsk Region. Part VII: Dicotyledonous plants (Asteraceae, Cichorioideae). Divers. Plant World 2021, 11, 5–33. (In Russian) [Google Scholar] [CrossRef]
- Knyazev, M.S.; Podgaevskaya, E.N.; Zolotareva, N.V.; Tretyakova, A.S.; Kulikov, P.V. Annotated checklist of the flora of Sverdlovsk Region. Part VIII: Dicotyledonous plants (Asteraceae, Asteroideae). Divers. Plant World 2022, 12, 28–56. (In Russian) [Google Scholar] [CrossRef]
- Fire Effects Information System (FEIS). USDA ForestService, Rocky Mountain Research Station, Fire Sciences laboratory Online. Available online: https://www.feis-crs.org/feis/faces/index.xhtml;jsessionid=643C2E17A6A3897DE7A5164F7B70016B (accessed on 14 August 2023).
- Pyšek, P.; Lambdon, P.W.; Arianoutsou, M.; Kühn, I.; Pino, J.; Winter, M. Alien Vascular Plants of Europe. In Handbook of Alien Species in Europe; Springer: Dordrecht, The Netherlands, 2009; pp. 43–61. [Google Scholar] [CrossRef]
- Dubovik, D.V.; Dmitrieva, S.A.; Laman, N.A.; Lebedko, V.N.; Levkovich, A.V.; Maslovskiy, O.M.; Parfenov, V.I.; Prokhorov, V.N.; Pugachevskiy, A.V. Black Book of the Flora of Belarus: Alien Harmful Plants; Parfenov, V.I., Pugachevskii, A.V., Eds.; Belaruskaja navuka: Minsk, Belarus, 2020; ISBN 978-985-08-2549-0. (In Russian) [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Khorun, L.V. The Black Book of Flora of Middle Russia; Dgebuadze, Y.Y., Demidov, A.S., Eds.; GEOS: Moscow, Russia, 2010; ISBN 978-88-911-9487-9. (In Russian) [Google Scholar]
- Dgebuadze, Y.Y.; Petrosyan, V.G.; Khlyap, L.A. The Most Dangerous Invasive Species of Russia (TOP-100); Dgebuadze, Y.Y., Petrosyan, V.G., Khlyap, L.A., Eds.; KMK Scientific Press: Moscow, Russia, 2018; ISBN 978-5-907099-53-1. (In Russian) [Google Scholar]
- Tretyakova, A.S.; Kulikov, P.V. Adventitious component of the flora Sverdlovsk region: Bioecological characteristics. Vestn. Udmurt. Universiteta. Ser. “Biologija. Nauk. O Zemle” 2014, 1, 57–67. (In Russian) [Google Scholar]
- Mamaev, S.A.; Dorofeyeva, L.M. Introduction of Maple in the Ural; Mamaev, S.A., Ed.; UB RAS: Ekaterinburg, Russia, 2005; ISBN 5-7691-1604-8. (In Russian) [Google Scholar]
- Veselkin, D.V.; Korzhinevskaya, A.A.; Podgayevskaya, E.N. Tree species composition and abundance of alien and invasive understory shrubs and trees in urban forests of Yekaterinburg. Vestn. Tomsk. Gos. Universitrta. Ser. “Biol.” 2018, 42, 102–118. [Google Scholar] [CrossRef]
- Kulikov, P.V.; Zolotareva, N.V.; Podgaevskaya, E.N. Endemic Plants of the Urals in the Flora of the Sverdlovsk Region; Mukhin, V.A., Ed.; Goshchitskii: Ekaterinburg, Russia, 2013; ISBN 978-5-98829-041-4. (In Russian) [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Kulikov, P.V. Key to Vascular Plants of the Chelyabinsk Region; Mukhin, V.A., Ed.; UB RAS: Ekaterinburg, Russia, 2010; ISBN 978-5-7691-2101-2. (In Russian) [Google Scholar]
- Majevski, P.F. Flora of the Middle Zone of the European Part of Russia, 11th ed.; Novikov, V.S., Pavlov, V.N., Sokolov, D.D., Timonin, A.K., Alekseev, Y.E., Mayorov, S.R., Eds.; KMK Scientific Press: Moscow, Russia, 2014; ISBN 978-5-87317-958-9. (In Russian) [Google Scholar]
- Tretyakov, D.I.; Savchuk, S.S. Flora of vascular plants biosphere reserve «Pribuzhskoye Polesie». Phytodivers. East. Eur. 2011, 9, 83–130. [Google Scholar]
- Atrasheuski, O.A. Flora of the Yukhnovskiy Biological Sanctuary (Belarus). Phytodivers. East. Eur. 2019, 1, 30–69. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org (accessed on 14 August 2023).
- Qureshi, H.; Anwar, T.; Mohibullah, M.; Fatima, S.; Younas, R.; Habiba, U.; Malik, L.; Hanif, A.; Iqbal, M. Paired plot experiments to assess impact of invasive species on native floral diversity in Pakistan. Front. Environ. Sci. 2023, 10, 1037319. [Google Scholar] [CrossRef]
- Kiełtyk, P.; Delimat, A. Impact of the alien plant Impatiens glandulifera on species diversity of invaded vegetation in the northern foothills of the Tatra Mountains, Central Europe. Plant Ecol. 2019, 220, 1–12. [Google Scholar] [CrossRef]
- Dubrovin, D.I.; Rafikova, O.S.; Veselkin, D.V. Soil moisture in urbanized habitats invaded by alien Acer negundo. Rus. J. Ecol. 2022, 5, 334–344. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Rafikova, O.S.; Ekshibarov, E.D. Soil from thickets of invasive Acer negundo is unfavorable for mycorrhiza formation in native grasses. J. Gen. Biol. 2019, 3, 214–225. [Google Scholar] [CrossRef]
- Rafikova, O.S.; Veselkin, D.V. Leaf water extracts from invasive Acer negundo do not inhibit seed germination more than leaf extracts from native species. Manag. Biol. Invasions 2022, 4, 705–723. [Google Scholar] [CrossRef]
- Veselkin, D.V.; Betekhtina, A.A.; Gusev, A.P. The intensity of formation of arbuscles is decreased in mycorrhizal alien plants of Belarusian Polesia. Rus. J. Ecol. 2022, 3, 143–151. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Belarusian Polesia, Gomel | Middle Urals, Ekaterinburg |
|---|---|---|
| Coordinates | ||
| Latitude | N 52°20′40″–52°30′20″ | N 56°39′20″–56°57′50″ |
| Longitude | E 30°48.5′30″–31°5′30″ | E 60°22′50″–60°50′30″ |
| Climate 1 | Cold with warm summer (Dfb) | Cold with warm summer (Dfb) |
| Average annual temperature, °C | +4.2 | +3.0 |
| Average temperature in July, °C | +20.4 | +19.0 |
| Average temperature in January, °C | –3.3 | –12.6 |
| Average annual precipitation, mm | 631 | 601 |
| The number of days with temperature above 10 °C | 167 | 132 |
| Soil | Sod-podzolic, sod-podzolic water-logged, peat-bog | Sod-podzolic, burozems |
| Landscape conformation | Plain | Low-mountain |
| Zonal community types | Broad-leaved forests | Coniferous forests |
| Main types of anthropogenic disturbance of vegetation | Deforestation for development and agriculture, fires, recreational use of native forests | |
| Proportion of human-disturbed landscapes, % | 50–60 | |
| Regional plant species richness 2 | ||
| Total | 1964 | 1696 |
| Herbaceous plants | 1758 | 1590 |
| Woody plants | 206 | 106 |
| Native plants | 1390 | 1401 |
| Native herbaceous plants | 1231 | 1312 |
| Native woody plants | 159 | 89 |
| Alien plants | 574 | 295 |
| Alien herbaceous plants | 527 | 278 |
| Alien woody plants | 47 | 17 |
| Species Number or Proportion | Variability Source | ||
|---|---|---|---|
| Plot Type (An+ or An−) (df = 1) | Region (Urals or Polesia) (df = 1) | Plot Type × Region (df = 1) | |
| Species number per 400 m2: | |||
| Total | 0.0001 | 0.0714 | 0.0335 |
| Woody | 0.0008 | 0.0063 | 0.0058 |
| Herbaceous | 0.0009 | 0.1438 | 0.0834 |
| Native species number per 400 m2: | |||
| Total | 0.0001 | 0.4096 | 0.1507 |
| Woody | 0.0001 | 0.0001 | 0.0198 |
| Herbaceous | 0.0002 | 0.7175 | 0.2574 |
| Alien species number per 400 m2: | |||
| Total | 0.5236 | 0.1777 | 0.0438 |
| Woody | 0.9069 | 0.4948 | 0.0295 |
| Herbaceous | 0.3988 | 0.0118 | 0.2958 |
| Alien species proportion, %: | |||
| Total | 0.0335 | 0.5718 | 0.2610 |
| Woody | 0.5929 | 0.1800 | 0.1890 |
| Herbaceous | 0.0349 | 0.0158 | 0.3073 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubrovin, D.I.; Veselkin, D.V.; Gusev, A.P. Plant Species Richness and Invasional Meltdown in Different Parts of Acer negundo L. Secondary Range. Forests 2023, 14, 2118. https://doi.org/10.3390/f14112118
Dubrovin DI, Veselkin DV, Gusev AP. Plant Species Richness and Invasional Meltdown in Different Parts of Acer negundo L. Secondary Range. Forests. 2023; 14(11):2118. https://doi.org/10.3390/f14112118
Chicago/Turabian StyleDubrovin, Denis I., Denis V. Veselkin, and Andrei P. Gusev. 2023. "Plant Species Richness and Invasional Meltdown in Different Parts of Acer negundo L. Secondary Range" Forests 14, no. 11: 2118. https://doi.org/10.3390/f14112118
APA StyleDubrovin, D. I., Veselkin, D. V., & Gusev, A. P. (2023). Plant Species Richness and Invasional Meltdown in Different Parts of Acer negundo L. Secondary Range. Forests, 14(11), 2118. https://doi.org/10.3390/f14112118