
How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes?


Abstract
:1. Introduction
2. Materials and Methods
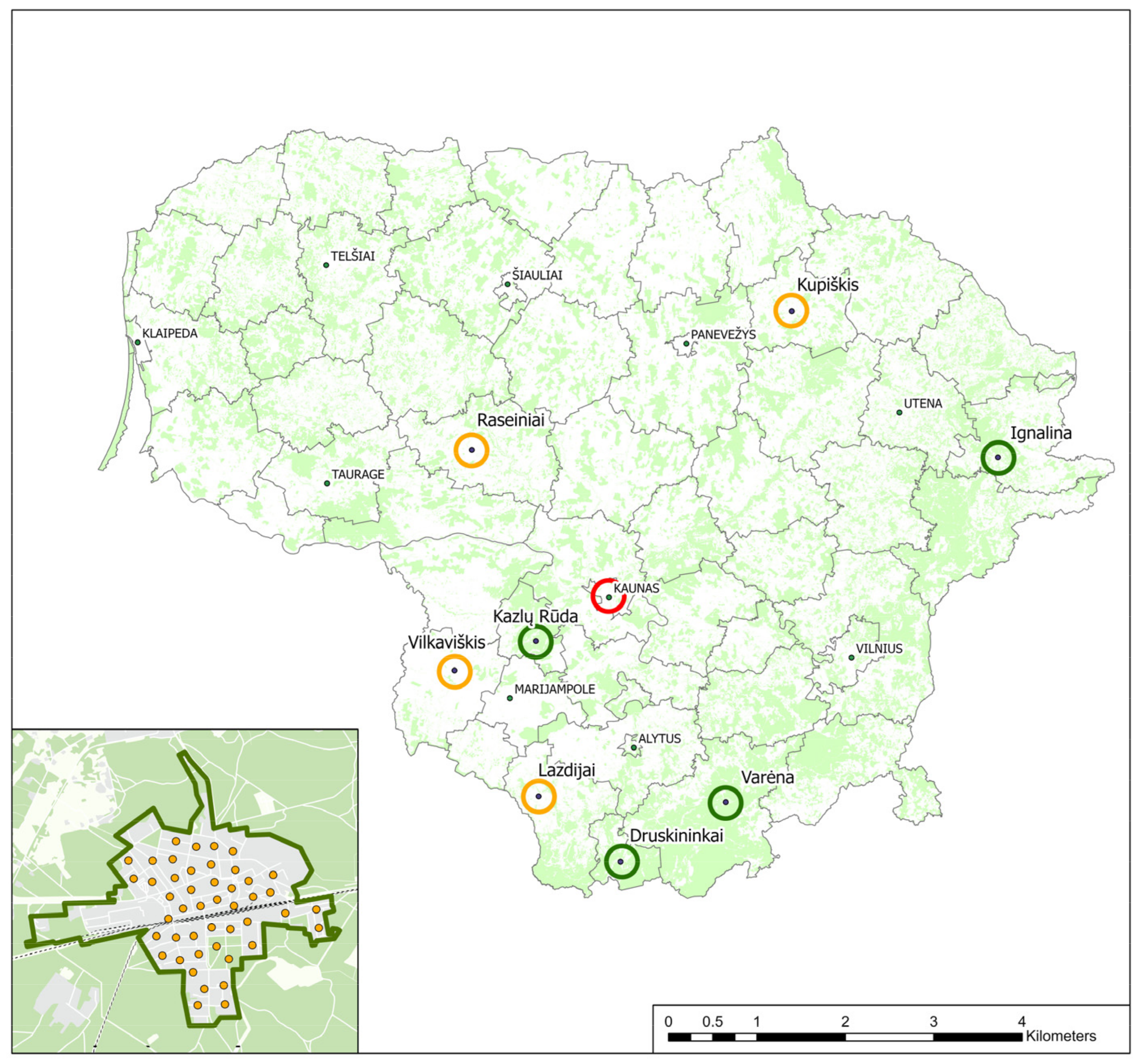
2.1. Study Area
2.2. Bird Census
2.3. Bird Community Indices
2.4. The Ecological Grouping of Bird Species
2.5. Statistical Analysis
3. Results
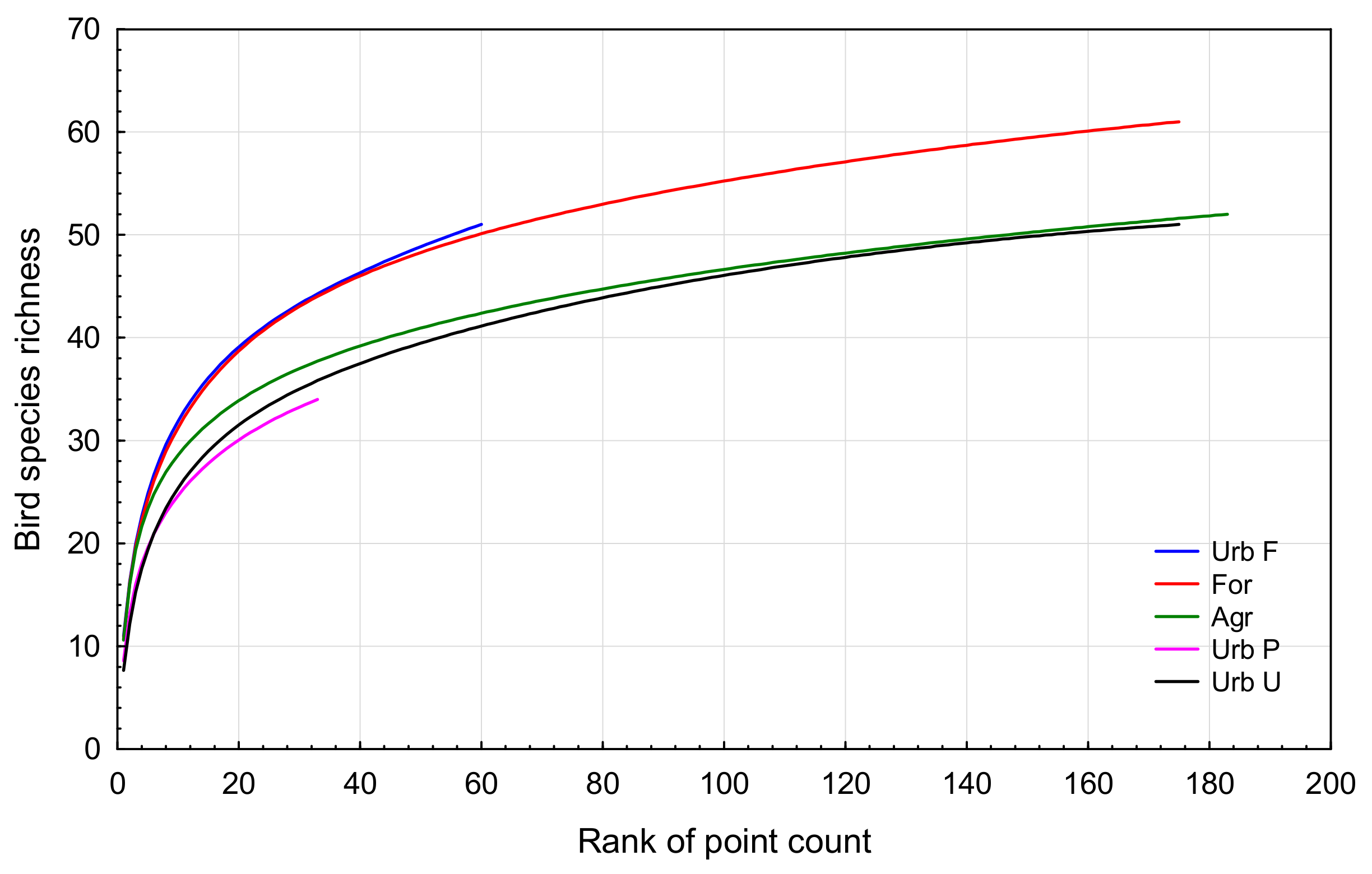
3.1. Bird Community Richness and Relative Abundance
3.2. The Affiliation of Bird Ecological Groups to Urbanized Landscapes
3.2.1. Feeding Areas
3.2.2. Main Food Type
3.2.3. Feeding Behavior
3.2.4. Nest Allocation
3.3. Grouping Bird Species Based on Habitat Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total Observed Individuals | For | Agr | UrbF | UrbP | UrbU | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | ||
| Ciconia ciconia | 16 | 0.029 | 0.013 | 0.060 | 0.018 | 0 | 0 | 0 | 0 | 0 | 0 |
| Columba palumbus | 223 | 0.291 | 0.036 | 0.568 | 0.043 | 0.333 | 0.07 | 0.364 | 0.085 | 0.206 | 0.032 |
| Streptopelia decaocto | 113 | 0.137 | 0.03 | 0.388 | 0.039 | 0.017 | 0.017 | 0.091 | 0.051 | 0.08 | 0.021 |
| Cuculus canorus | 7 | 0 | 0 | 0.005 | 0.005 | 0.083 | 0.036 | 0 | 0 | 0.006 | 0.006 |
| Upupa epops | 4 | 0.023 | 0.011 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Jynx torquilla | 19 | 0.063 | 0.018 | 0.033 | 0.013 | 0 | 0 | 0 | 0 | 0.011 | 0.008 |
| Dryocopus martius | 5 | 0 | 0 | 0 | 0 | 0.083 | 0.036 | 0 | 0 | 0 | 0 |
| Dendrocopos major | 44 | 0.051 | 0.017 | 0 | 0 | 0.5 | 0.073 | 0 | 0 | 0.029 | 0.013 |
| Hirundo rustica | 64 | 0.177 | 0.036 | 0.153 | 0.031 | 0 | 0 | 0 | 0 | 0.029 | 0.013 |
| Motacilla alba | 356 | 0.629 | 0.048 | 0.787 | 0.053 | 0 | 0 | 0.667 | 0.142 | 0.457 | 0.043 |
| Troglodytes troglodytes | 41 | 0.006 | 0.006 | 0.005 | 0.005 | 0.65 | 0.106 | 0 | 0 | 0 | 0 |
| Erithacus rubecula | 75 | 0.069 | 0.019 | 0.022 | 0.011 | 0.817 | 0.102 | 0.061 | 0.042 | 0.046 | 0.016 |
| Luscinia luscinia | 5 | 0.011 | 0.008 | 0.011 | 0.008 | 0.017 | 0.017 | 0 | 0 | 0 | 0 |
| Phoenicurus ochruros | 249 | 0.891 | 0.048 | 0.377 | 0.038 | 0 | 0 | 0 | 0 | 0.137 | 0.028 |
| Phoenicurus phoenicurus | 288 | 0.531 | 0.043 | 0.541 | 0.043 | 0.083 | 0.036 | 0.03 | 0.03 | 0.514 | 0.044 |
| Oenanthe isabellina | 14 | 0.051 | 0.019 | 0.011 | 0.008 | 0 | 0 | 0.03 | 0.03 | 0.011 | 0.008 |
| Turdus merula | 332 | 0.2 | 0.031 | 0.415 | 0.039 | 1.4 | 0.099 | 0.485 | 0.088 | 0.691 | 0.053 |
| Turdus pilaris | 295 | 0.326 | 0.041 | 0.82 | 0.055 | 0.133 | 0.05 | 1.364 | 0.178 | 0.2 | 0.038 |
| Turdus philomelos | 51 | 0.011 | 0.008 | 0.082 | 0.02 | 0.5 | 0.081 | 0 | 0 | 0.023 | 0.011 |
| Turdus viscivorus | 5 | 0 | 0 | 0 | 0 | 0.083 | 0.036 | 0 | 0 | 0 | 0 |
| Hippolais icterina | 75 | 0.074 | 0.02 | 0.153 | 0.031 | 0 | 0 | 0.576 | 0.123 | 0.086 | 0.024 |
| Acrocephalus palustris | 8 | 0.006 | 0.006 | 0.022 | 0.011 | 0 | 0 | 0.03 | 0.03 | 0.011 | 0.008 |
| Sylvia curruca | 194 | 0.36 | 0.04 | 0.383 | 0.044 | 0 | 0 | 0.182 | 0.081 | 0.314 | 0.039 |
| Sylvia communis | 64 | 0.126 | 0.025 | 0.082 | 0.02 | 0 | 0 | 0.273 | 0.1 | 0.103 | 0.024 |
| Sylvia borin | 34 | 0.023 | 0.011 | 0.027 | 0.012 | 0.133 | 0.05 | 0.121 | 0.072 | 0.074 | 0.02 |
| Sylvia atricapilla | 54 | 0.074 | 0.02 | 0.087 | 0.021 | 0.35 | 0.085 | 0.061 | 0.042 | 0.011 | 0.008 |
| Phylloscopus sibilatrix | 88 | 0.011 | 0.008 | 0 | 0 | 1.383 | 0.152 | 0 | 0 | 0.017 | 0.01 |
| Phylloscopus collybita | 115 | 0.103 | 0.024 | 0.077 | 0.02 | 1.017 | 0.105 | 0.121 | 0.058 | 0.103 | 0.023 |
| Phylloscopus trochilus | 33 | 0.063 | 0.018 | 0.049 | 0.016 | 0.083 | 0.043 | 0.03 | 0.03 | 0.04 | 0.017 |
| Regulus regulus | 64 | 0.011 | 0.008 | 0 | 0 | 0.95 | 0.174 | 0 | 0 | 0.029 | 0.019 |
| Muscicapa striata | 62 | 0.149 | 0.029 | 0.148 | 0.027 | 0.133 | 0.056 | 0 | 0 | 0.006 | 0.006 |
| Ficedula hypoleuca | 61 | 0.177 | 0.03 | 0.022 | 0.011 | 0.133 | 0.044 | 0 | 0 | 0.103 | 0.024 |
| Cyanistes caeruleus | 249 | 0.549 | 0.05 | 0.366 | 0.042 | 0.317 | 0.077 | 0.424 | 0.107 | 0.303 | 0.041 |
| Parus major | 668 | 0.966 | 0.056 | 0.781 | 0.055 | 2.2 | 0.181 | 0.727 | 0.146 | 1.143 | 0.068 |
| Periparus ater | 31 | 0.023 | 0.011 | 0 | 0 | 0.433 | 0.084 | 0 | 0 | 0.006 | 0.006 |
| Lophophanes cristatus | 5 | 0.006 | 0.006 | 0 | 0 | 0.067 | 0.052 | 0 | 0 | 0 | 0 |
| Poecile palustris | 13 | 0.011 | 0.008 | 0.005 | 0.005 | 0.117 | 0.054 | 0.03 | 0.03 | 0.011 | 0.008 |
| Poecile montanus | 7 | 0 | 0 | 0 | 0 | 0.117 | 0.042 | 0 | 0 | 0 | 0 |
| Sitta europaea | 40 | 0.046 | 0.018 | 0.016 | 0.009 | 0.433 | 0.087 | 0 | 0 | 0.017 | 0.01 |
| Certhia familiaris | 24 | 0.006 | 0.006 | 0 | 0 | 0.35 | 0.085 | 0 | 0 | 0.011 | 0.011 |
| Oriolus oriolus | 8 | 0.017 | 0.01 | 0.011 | 0.008 | 0.033 | 0.023 | 0 | 0 | 0.006 | 0.006 |
| Lanius collurio | 4 | 0.006 | 0.006 | 0.005 | 0.005 | 0 | 0 | 0 | 0 | 0.011 | 0.008 |
| Garrulus glandarius | 49 | 0.051 | 0.019 | 0 | 0 | 0.35 | 0.071 | 0.061 | 0.042 | 0.097 | 0.022 |
| Pica pica | 74 | 0.097 | 0.022 | 0.115 | 0.024 | 0 | 0 | 0.121 | 0.058 | 0.183 | 0.029 |
| Coloeus monedula | 40 | 0.046 | 0.02 | 0.126 | 0.027 | 0 | 0 | 0.212 | 0.072 | 0.011 | 0.011 |
| Corvus cornix | 227 | 0.406 | 0.038 | 0.055 | 0.017 | 0.233 | 0.084 | 0.788 | 0.121 | 0.606 | 0.044 |
| Corvus corax | 5 | 0 | 0 | 0 | 0 | 0.083 | 0.036 | 0 | 0 | 0 | 0 |
| Sturnus vulgaris | 950 | 2.011 | 0.097 | 1.918 | 0.089 | 0.067 | 0.047 | 1.424 | 0.204 | 1.12 | 0.08 |
| Passer domesticus | 751 | 1.509 | 0.098 | 2.033 | 0.11 | 0 | 0 | 0.121 | 0.095 | 0.634 | 0.074 |
| Passer montanus | 676 | 1.114 | 0.069 | 0.918 | 0.062 | 0 | 0 | 1.273 | 0.28 | 1.549 | 0.097 |
| Fringilla coelebs | 466 | 0.84 | 0.059 | 0.574 | 0.052 | 1.783 | 0.126 | 1.061 | 0.157 | 0.411 | 0.05 |
| Serinus serinus | 212 | 0.589 | 0.046 | 0.410 | 0.042 | 0 | 0 | 0.303 | 0.102 | 0.137 | 0.03 |
| Chloris chloris | 281 | 0.617 | 0.056 | 0.574 | 0.059 | 0 | 0 | 0.212 | 0.095 | 0.349 | 0.046 |
| Carduelis carduelis | 155 | 0.171 | 0.03 | 0.426 | 0.052 | 0 | 0 | 0.636 | 0.129 | 0.149 | 0.033 |
| Spinus spinus | 19 | 0.040 | 0.015 | 0 | 0 | 0.167 | 0.059 | 0 | 0 | 0.011 | 0.008 |
| Linaria cannabina | 176 | 0.16 | 0.029 | 0.705 | 0.057 | 0 | 0 | 0 | 0 | 0.109 | 0.029 |
| Carpodacus erythrinus | 5 | 0.011 | 0.008 | 0.011 | 0.008 | 0.017 | 0.017 | 0 | 0 | 0 | 0 |
| Pyrrhula pyrrhula | 24 | 0.074 | 0.027 | 0.005 | 0.005 | 0.133 | 0.05 | 0 | 0 | 0.011 | 0.008 |
| Coccothraustes coccothraustes | 78 | 0.097 | 0.022 | 0.115 | 0.024 | 0.35 | 0.085 | 0.121 | 0.072 | 0.086 | 0.025 |
| Emberiza citrinella | 13 | 0.040 | 0.015 | 0.027 | 0.012 | 0.017 | 0.017 | 0 | 0 | 0 | 0 |
Appendix B
| Bird Ecological Group | For | Agr | Urb F | Urb P | Urb U | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | Mean | ±SE | |
| Feeding outside the urban territory (O) | 7.137 | 0.234 | 8.208 | 0.260 | 2.383 | 0.209 | 5.455 | 0.450 | 5.274 | 0.198 |
| Feeding near the nest (N) | 7.109 | 0.232 | 6.393 | 0.212 | 13.467 | 0.501 | 6.545 | 0.535 | 5.034 | 0.204 |
| Herbivorous (V) | 4.163 | 0.130 | 5.184 | 0.162 | 2.200 | 0.135 | 3.133 | 0.302 | 2.898 | 0.112 |
| Feeding on invertebrates (I) | 9.645 | 0.254 | 9.260 | 0.243 | 13.353 | 0.473 | 8.100 | 0.559 | 6.806 | 0.221 |
| Feeding on vertebrates (predators) (P) | 0.438 | 0.037 | 0.157 | 0.026 | 0.297 | 0.086 | 0.767 | 0.113 | 0.605 | 0.041 |
| Foraging on the ground or in the herb layer (IG) | 6.154 | 0.173 | 7.576 | 0.215 | 4.273 | 0.201 | 6.494 | 0.456 | 4.831 | 0.170 |
| Collecting food from leaves. twigs and the air (IL) | 7.983 | 0.214 | 7.020 | 0.191 | 10.277 | 0.460 | 5.506 | 0.471 | 5.421 | 0.173 |
| Foraging on tree—barks (IB) | 0.109 | 0.029 | 0.016 | 0.009 | 1.300 | 0.169 | 0.000 | 0.000 | 0.057 | 0.023 |
| Ground nesting (G) | 1.466 | 0.088 | 2.023 | 0.105 | 4.585 | 0.262 | 1.321 | 0.176 | 1.189 | 0.071 |
| Crown nesting (C) | 4.154 | 0.181 | 4.755 | 0.228 | 6.183 | 0.314 | 5.833 | 0.476 | 3.197 | 0.164 |
| Hole nesting (H) | 8.422 | 0.213 | 7.652 | 0.205 | 4.995 | 0.327 | 4.833 | 0.413 | 5.892 | 0.175 |
References
- Czech, B.; Krausman, P.R.; Devers, P.K. Economic associations among causes of species endangerment in the United States: Associations among causes of species endangerment in the United States reflect the integration of economic sectors, supporting the theory and evidence that economic growth proceeds at the competitive exclusion of nonhuman species in the aggregate. BioScience 2000, 50, 593–601. [Google Scholar]
- Elmqvist, T.; Zipperer, W.C.; Güneralp, B. Urbanization, habitat loss and biodiversity decline: Solution pathways to break the cycle. In The Routledge Handbook of Urbanization and Global Environmental Change; Routledge: London, UK, 2015; pp. 163–175. [Google Scholar]
- Young, H.S.; McCauley, D.J.; Galetti, M.; Dirzo, R. Patterns, causes, and consequences of anthropocene defaunation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 333–358. [Google Scholar] [CrossRef]
- Lepczyk, C.A.; La Sorte, F.A.; Aronson, M.F.; Goddard, M.A.; MacGregor-Fors, I.; Nilon, C.H.; Warren, P.S. Global patterns and drivers of urban bird diversity. In Ecology and Conservation of Birds in Urban Environments; Springer: Cham, 2017; pp. 13–33. [Google Scholar]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Beissinger, S.R.; Osborne, D.R. Effects of urbanization on avian community organization. Condor 1982, 84, 75–83. [Google Scholar] [CrossRef]
- Bezzel, E. Birdlife in intensively used rural and urban environments. Ornis Fenn. 1985, 62, 90–95. [Google Scholar]
- Jonathan, A.F.; Ruth, D.; Gregory, P.A.; Carol, B.; Gordon, B.; Stephen, R.C. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar]
- Dearborn, D.C.; Kark, S. Motivations for conserving urban biodiversity. Conserv. Biol. 2010, 24, 432–440. [Google Scholar] [CrossRef]
- Lawler, J.J.; Lewis, D.J.; Nelson, E.; Plantinga, A.J.; Polasky, S.; Withey, J.C.; Helmers, D.P.; Martinuzzi, S.; Pennington, D.; Radeloff, V.C. Projected land-use change impacts on ecosystem services in the United States. Proc. Natl. Acad. Sci. USA 2014, 111, 7492–7497. [Google Scholar] [CrossRef]
- Liu, Z.; He, C.; Wu, J. The relationship between habitat loss and fragmentation during urbanization: An empirical evaluation from 16 world cities. PLoS ONE 2016, 11, e0154613. [Google Scholar] [CrossRef]
- Goldewijk, K.K. Estimating global land use change over the past 300 years: The HYDE database. Glob. Biogeochem. Cycles 2001, 15, 417–433. [Google Scholar] [CrossRef]
- United Nations. New Urban Agenda (Habitat III); United Nations: Quito, Ecuador, 2017. [Google Scholar]
- European Commission Green Infrastructure InPolicy. 2023. Available online: https://environment.ec.europa.eu/topics/nature-and-biodiversity/green-infrastructure_en#policy (accessed on 18 February 2023).
- Felson, A.J.; Oldfield, E.E.; Bradford, M.A. Involving ecologists in shaping large-scale green infrastructure projects. BioScience 2013, 63, 882–890. [Google Scholar]
- Lerman, S.B.; Nislow, K.H.; Nowak, D.J.; DeStefano, S.; King, D.I.; Jones-Farrand, D.T. Using urban forest assessment tools to model bird habitat potential. Landsc. Urban Plan. 2014, 122, 29–40. [Google Scholar] [CrossRef]
- Jimenez, M.F.; Pejchar, L.; Reed, S.E.; McHale, M.R. The efficacy of urban habitat enhancement programs for conserving native plants and human-sensitive animals. Landsc. Urban Plan. 2022, 220, 104356. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an ‘urban exploiter’? J. Biogeogr. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.L. Urbanization and species occupancy frequency distribution patterns in core zone areas of European towns. Eur. J. Ecol. 2016, 2, 23–43. [Google Scholar] [CrossRef]
- Battisti, C.; Zullo, F. A recent colonizer bird as indicator of human-induced landscape change: Eurasian collared dove (Streptopelia decaocto) in a small Mediterranean island. Reg. Environ. Chang. 2019, 19, 2113–2121. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Major, R.E.; Wilshire, J.H.; Martin, J.M.; Kingsford, R.T.; Cornwell, W.K. Generalists are the most urban-tolerant of birds: A phylogenetically controlled analysis of ecological and life history traits using a novel continuous measure of bird responses to urbanization. Oikos 2019, 128, 845–858. [Google Scholar] [CrossRef]
- Palacio, F.X. Urban exploiters have broader dietary niches than urban avoiders. Ibis 2020, 162, 42–49. [Google Scholar] [CrossRef]
- Šálek, M.; Grill, S.; Riegert, J. Nest-site selection of an avian urban exploiter, the Eurasian magpie Pica pica, across the urban-rural gradient. J. Vertebr. Biol. 2020, 70, 20086-1. [Google Scholar] [CrossRef]
- Møller, A.P.; Díaz, M. Avian preference for close proximity to human habitation and its ecological consequences. Curr. Zool. 2018, 64, 623–630. [Google Scholar] [CrossRef]
- Adams, L.W. Urban Wildlife Habitats: A Landscape Perspective; U of Minnesota Press: Minneapolis, MN, USA, 1994; Volume 3. [Google Scholar]
- Alberti, M. Advances in Urban Ecology: Integrating Humans and Ecological Processes in Urban Ecosystems; Springer: New York, NY, USA, 2008. [Google Scholar]
- Karimi, J.D.; Corstanje, R.; Harris, J.A. Bundling ecosystem services at a high resolution in the UK: Trade-offs and synergies in urban landscapes. Landsc. Ecol. 2021, 36, 1817–1835. [Google Scholar] [CrossRef]
- Marzluff, J.M. Worldwide urbanization and its effects on birds. In Avian Ecology and Conservation in An Urbanizing World; Springer: Berlin/Heidelberg, Germany, 2001; pp. 19–47. [Google Scholar]
- Jokimäki, J.; Suhonen, J.; Jokimäki-Kaisanlahti, M.L.; Carbó-Ramírez, P. Effects of urbanization on breeding birds in European towns: Impacts of species traits. Urban Ecosyst. 2016, 19, 1565–1577. [Google Scholar] [CrossRef]
- Concepción, E.D.; Moretti, M.; Altermatt, F.; Nobis, M.P.; Obrist, M.K. Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale. Oikos 2015, 124, 1571–1582. [Google Scholar] [CrossRef]
- Donihue, C.M.; Lambert, M.R. Adaptive evolution in urban ecosystems. Ambio 2015, 44, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Lerman, S.B.; Narango, D.L.; Avolio, M.L.; Bratt, A.R.; Engebretson, J.M.; Groffman, P.M.; Hall, S.J.; Heffernan, J.B.; Hobbie, S.E.; Larson, K.L.; et al. Residential yard management and landscape cover affect urban bird community diversity across the continental USA. Ecol. Appl. 2021, 31, e02455. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Marcacci, G.; Westphal, C.; Wenzel, A.; Raj, V.; Nölke, N.; Tscharntke, T.; Grass, I. Taxonomic and functional homogenization of farmland birds along an urbanization gradient in a tropical megacity. Glob. Chang. Biol. 2021, 27, 4980–4994. [Google Scholar] [CrossRef]
- Lancaster, R.K.; Rees, W.E. Bird communities and the structure of urban habitats. Can. J. Zool. 1979, 57, 2358–2368. [Google Scholar] [CrossRef]
- Clergeau, P.; Savard, J.P.L.; Mennechez, G.; Falardeau, G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor 1998, 100, 413–425. [Google Scholar] [CrossRef]
- Shwartz, A.; Muratet, A.; Simon, L.; Julliard, R. Local and management variables outweigh landscape effects in enhancing the diversity of different taxa in a big metropolis. Biol. Conserv. 2013, 157, 285–292. [Google Scholar] [CrossRef]
- Kim, K. Morphological analysis of green infrastructure in the Seoul metropolitan area, South Korea. Landsc. Ecol. Eng. 2015, 11, 259–268. [Google Scholar]
- Morrison, M.L. Bird populations as indicators of environmental change. In Current Ornithology; Springer: Boston, MA, USA, 1986; Volume 3, pp. 429–451. [Google Scholar]
- Canterbury, G.E.; Martin, T.E.; Petit, D.R.; Petit, L.J.; Bradford, D.F. Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv. Biol. 2000, 14, 544–558. [Google Scholar] [CrossRef]
- Sekercioglu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Morelli, F.; Reif, J.; Díaz, M.; Tryjanowski, P.; Ibáñez-Álamo, J.D.; Suhonen, J.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Møller, A.P.; Bussière, R.; et al. Top ten birds indicators of high environmental quality in European cities. Ecol. Indic. 2021, 133, 108397. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.R.; Lee, D. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Amaya-Espinel, J.D.; Hostetler, M.; Henriquez, C.; Bonacic, C. The influence of building density on Neotropical bird communities found in small urban parks. Landsc. Urban Plan. 2019, 190, 103578. [Google Scholar] [CrossRef]
- De Groot, M.; Flajšman, K.; Mihelič, T.; Vilhar, U.; Simončič, P.; Verlič, A. Green space area and type affect bird communities in a South-eastern European city. Urban For. Urban Green. 2021, 63, 127212. [Google Scholar] [CrossRef]
- Liordos, V.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Valsamidis, E.; Kontsiotis, V.J. Patch, matrix and disturbance variables negatively influence bird community structure in small-sized managed green spaces located in urban core areas. Sci. Total Environ. 2021, 801, 149617. [Google Scholar] [CrossRef]
- Sander, M.M.; Tietze, D.T. Impacts of Traffic Infrastructure on Urban Bird Communities: A Review. Sustainability 2022, 14, 16805. [Google Scholar] [CrossRef]
- Hennings, L.A.; Edge, W.D. Riparian bird community structure in Portland, Oregon: Habitat, urbanization, and spatial scale patterns. Condor 2003, 105, 288–302. [Google Scholar] [CrossRef]
- Forman, R.F.F.; Godron, M. Landscape Ecology; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Mills, G.S.; Dunning, J.B., Jr.; Bates, J.M. Effects of urbanization on breeding bird community structure in southwestern desert habitats. Condor 1989, 91, 416–428. [Google Scholar] [CrossRef]
- Asoskova, N.I.; Durnev, Y.A.; Moroshenko, N.V. Birds of the Cities of Russia. Association of scientific editions of KMK: St. Petersburg/Moscow, Russia, 2012. [Google Scholar]
- Clergeau, P.; Jokimäki, J.; Savard, J.P.L. Are urban bird communities influenced by the bird diversity of adjacent landscapes? J. Appl. Ecol. 2001, 38, 1122–1134. [Google Scholar] [CrossRef]
- Björklund, M.; Ruiz, I.; Senar, J.C. Genetic differentiation in the urban habitat: The great tits (Parus major) of the parks of Barcelona city. Biol. J. Linn. Soc. 2010, 99, 9–19. [Google Scholar] [CrossRef]
- Aleknavičius, P. Aplinkosaugos ir Aplinkotvarkos teisė: Metodiniai Patarimai; Ardiva: Kaunas, Lietuva, 2008; ISBN 978-9955-760-91-7. [Google Scholar]
- Priednieks, J.; Kuresoo, A.; Kurlavičius, P. Rekomendacii k Ornitologičeskomu Monitoringu v Pribaltike; Zinatne: Ryga, Latvia, 1986. (In Russian) [Google Scholar]
- Bibby, C.J.; Burgess, N.D.; Hillis, D.M.; Hill, D.A.; Mustoe, S. Bird Census Techniques; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Kurlavičius, P. Birds of Forest Islands in South-East Baltic Region; Baltic ECO: Vilnius, Lithuania, 1995; 263p. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Harper Row: New York, NY, USA, 1989; 654p. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Dierschke, F.V. Die Sommervogelbestände nordwestdeutscher Kiefernforsten. Vogelwelt 1973, 94, 201–225. [Google Scholar]
- Kurlavičius, P. Biotopičeskoje raspredelenije ptic v agronasaždenijach; Mokslas: Vilnius, Lietuva, 1986. [Google Scholar]
- Flade, M. Die Brutvogelgemeinschaften Mittel-und Norddeutschlands: Grundlagen für den Gebrauch Vogelkundlicher Daten in der Landschaftsplanung; IHW: Eching, Germany, 1994; ISBN 10: 393016700X/ISBN 13: 9783930167005. [Google Scholar]
- Gil-Tena, A.; Saura, S.; Brotons, L. Effects of forest composition and structure on bird species richness in a Mediterranean context: Implications for forest ecosystem management. For. Ecol. Manag. 2007, 242, 470–476. [Google Scholar] [CrossRef]
- Illera, J.C.; Atienza, J.C. Foraging shifts by the Blue Tit (Parus caeruleus) in relation to arthropod availability in a mixed woodland during the spring-summer period. Ardeola 1995, 42, 39–48. [Google Scholar]
- Hobson, K.A.; Bayne, E. Breeding bird communities in boreal forest of western Canada: Consequences of “unmixing” the mixedwoods. Condor 2000, 102, 759–769. [Google Scholar]
- Díaz, L. Influences of forest type and forest structure on bird communities in oak and pine woodlands in Spain. For. Ecol. Manag. 2006, 223, 54–65. [Google Scholar] [CrossRef]
- Cannon, A. The significance of private gardens for bird conservation. Bird Conserv. Int. 1999, 9, 287–297. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.A.; Ikin, K.; Lerman, S.B. Ecological and social factors determining the diversity of birds in residential yards and gardens. In Ecology and Conservation of Birds in Urban Environments; Springer: Berlin/Heidelberg, Germany, 2017; pp. 371–397. [Google Scholar]
- Havlíček, J.; Riegert, J.; Bandhauerová, J.; Fuchs, R.; Šálek, M. Species-specific breeding habitat association of declining farmland birds within urban environments: Conservation implications. Urban Ecosyst. 2021, 24, 1259–1270. [Google Scholar] [CrossRef]
- Batten, L.A. Breeding bird species diversity in relation to increasing urbanisation. Bird Study 1972, 19, 157–166. [Google Scholar] [CrossRef]
- Idzelis, R.L.; Jankaitė, A.; Oškinis, V.; Pakalnis, R.; Venckus, Z.; Virbickas, T. Assessment of the state of natural environment in Vilnius city. In Proceedings of the 7th International Conference Environmental Engineering, Vilnius, Lithuania, 22–23 May 2008; pp. 154–162. [Google Scholar]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Bisikirskienė, L.; Brazaitis, G. The influence of urbanization on diversity and abundance of breeding bird species in Kaunas city. In Human and Nature Safety 2020: Selected Papers; Vytautas Magnus University Agriculture Academy: Akademija, Lithuania, 2020; pp. 123–125. [Google Scholar]
- Shochat, E.; Lerman, S.B.; Anderies, J.M.; Warren, P.S.; Faeth, S.H.; Nilon, C.H. Invasion, competition, and biodiversity loss in urban ecosystems. BioScience 2010, 60, 199–208. [Google Scholar] [CrossRef]
- Boal, C.W.; Mannan, R.W. Nest-site selection by Cooper’s Hawks in an urban environment. J. Wildl. Manag. 1998, 62, 864–871. [Google Scholar] [CrossRef]
- DeStefano, S.; DeGraaf, R.M. Exploring the ecology of suburban wildlife. Front. Ecol. Environ. 2003, 1, 95–101. [Google Scholar] [CrossRef]
- Wilson, D.S.; Yoshimura, J. On the coexistence of specialists and generalists. Am. Nat. 1994, 144, 692–707. [Google Scholar] [CrossRef]
- Julliard, R.; Clavel, J.; Devictor, V.; Jiguet, F.; Couvet, D. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 2006, 9, 1237–1244. [Google Scholar] [CrossRef]
- Staude, I.R.; Overbeck, G.E.; Fontana, C.S.; Bencke, G.A.; Silva, T.W.D.; Mimet, A.; Pereira, H.M. Specialist birds replace generalists in grassland remnants as land use change intensifies. Front. Ecol. Evol. 2021, 8, 597542. [Google Scholar] [CrossRef]
- Logminas, V.; Nedzinskas, V.; Drobelis, E. Lietuvos fauna. Paukščiai 2; Mokslas: Vilnius, Lietuva, 1991; ISBN 5-420-00398-8. [Google Scholar]
- IUCN 2023. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org (accessed on 19 April 2023).
- Kareklas, K.; Wilson, J.; Kunc, H.P.; Arnott, G. Signal complexity communicates aggressive intent during contests, but the process is disrupted by noise. Biol. Lett. 2019, 15, 20180841. [Google Scholar] [CrossRef] [PubMed]
- Juškaitis, R. Is nesting in closed nestboxes advantageous for the European Robin (Erithacus rubecula)? Avian Res. 2021, 12, 21. [Google Scholar] [CrossRef]
- Önsal, Ç.; Yelimlieş, A.; Akçay, Ç. Aggression and multimodal signalling in noise in the European robin. Biorxiv 2022. [Google Scholar] [CrossRef]
- Logminas, V.; Nedzinskas, V.; Drobelis, E. Lietuvos fauna. Paukščiai 1; Mokslas: Vilnius, Lietuva, 1990; ISBN 5-420-00397-X. [Google Scholar]
- Bea, A.; Svazas, S.; Grishanov, G.; Kozulin, A.; Stanevicius, V.; Astafieva, T.; Olano, I.; Raudonikis, L.; Butkauskas, D.; Sruoga, A. Woodland and urban populations of the woodpigeon Columba palumbus in the Eastern Baltic region. Ardeola 2011, 58, 315–321. [Google Scholar] [CrossRef]
- Jusys, V.; Karalius, S.; Raudonikis, L. Lietuvos Paukščių Pažinimo Vadovas; Lutute: Kaunas, Lietuva, 2012; ISBN 978-9955-37-144-1. [Google Scholar]
- Tomiałojć, L. Impact of Nest Predators on Migratory Woodpigeons Columba palumbus in Central Europe—Breeding Densities and Nesting Success in Urban Versus Natural Habitats. Acta Ornithol. 2021, 55, 139–154. [Google Scholar] [CrossRef]
- Svazas, S. Population status of pigeons and doves in the eastern Baltic region. Naturzale 2001, 16, 71–81. [Google Scholar]
- Bochio, G.M.; Anjos, L. The importance of considering bird detectability for assessing biological integrity. Nat. Conserv. 2012, 10, 72–76. [Google Scholar] [CrossRef]
- van Heezik, Y.; Seddon, P.J. Counting birds in urban areas: A review of methods for the estimation of abundance. In Ecology and Conservation of Birds in Urban Environments; Springer: Berlin/Heidelberg, Germany, 2017; pp. 185–207. [Google Scholar]
- Morelli, F.; Brlík, V.; Benedetti, Y.; Bussière, R.; Moudrá, L.; Reif, J.; Svitok, M. Detection rate of bird species and what it depends on: Tips for field surveys. Front. Ecol. Evol. 2022, 9, 671492. [Google Scholar] [CrossRef]
- Conway, C.J.; Gibbs, J.P. Summary of intrinsic and extrinsic factors affecting detection probability of marsh birds. Wetlands 2011, 31, 403–411. [Google Scholar] [CrossRef]
- Farnsworth, G.L.; Pollock, K.H.; Nichols, J.D.; Simons, T.R.; Hines, J.E.; Sauer, J.R. A removal model for estimating detection probabilities from point-count surveys. Auk 2002, 119, 414–425. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Virkkala, R.; Rainio, K. Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic. J. Appl. Ecol. 2004, 41, 824–835. [Google Scholar] [CrossRef]
- Catterall, C.P.; Freeman, A.N.; Kanowski, J.; Freebody, K. Can active restoration of tropical rainforest rescue biodiversity? A case with bird community indicators. Biol. Conserv. 2012, 146, 53–61. [Google Scholar] [CrossRef]
- Thompson, R.; Tamayo, M.; Sigurðsson, S. Urban bird diversity: Does abundance and richness vary unexpectedly with green space attributes? J. Urban Ecol. 2022, 8, juac017. [Google Scholar] [CrossRef]
- Pan-European Common Bird Monitoring Scheme. 2002. Available online: https://pecbms.info/ (accessed on 13 October 2023).
- Blair, R.B. Land use and avian species diversity along an urban gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Batáry, P.; Kurucz, K.; Suarez-Rubio, M.; Chamberlain, D.E. Non-linearities in bird responses across urbanization gradients: A meta-analysis. Glob. Chang. Biol. 2018, 24, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Heggie-Gracie, S.D.; Krull, C.R.; Stanley, M.C. Urban divide: Predictors of bird communities in forest fragments and the surrounding urban matrix. Emu-Austral Ornithol. 2020, 120, 333–342. [Google Scholar] [CrossRef]
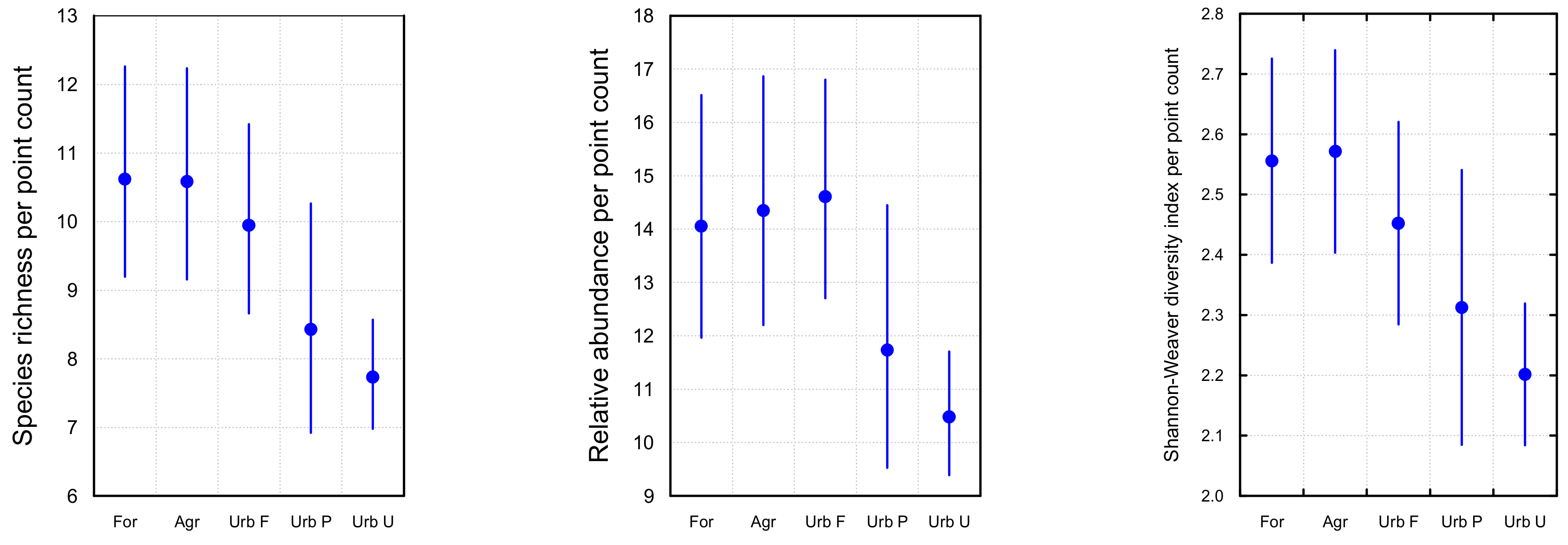

| Fixed Effect | Estimate | ±SE | p |
|---|---|---|---|
| Species richness per point | |||
| Agr | 10.583 | 0.781 | 0.001 |
| For | 10.62 | 0.778 | 0.001 |
| Urb F | 9.944 | 0.7 | 0.001 |
| Urb P | 8.428 | 0.845 | 0.446 |
| Urb U | 7.732 | 0.404 | 0.001 |
| Bird relative abundance per point | |||
| Agr | 14.347 | 1.182 | 0.002 |
| For | 14.056 | 1.152 | 0.003 |
| Urb F | 14.61 | 1.04 | 0.001 |
| Urb P | 11.735 | 1.244 | 0.347 |
| Urb U | 10.483 | 0.589 | 0.001 |
| Shannon–Weaver diversity index | |||
| Agr | 2.556 | 0.086 | 0.001 |
| For | 2.572 | 0.086 | 0.001 |
| Urb F | 2.453 | 0.086 | 0.002 |
| Urb P | 2.313 | 0.116 | 0.395 |
| Urb U | 2.202 | 0.06 | 0.001 |
| Landscape Type | For | Agr | UrbF | Urb U | Urb P |
|---|---|---|---|---|---|
| For | - | A | R A D | R A D | |
| Agr | - | A | R A D | R A D | |
| UrbF | A | A | - | R A D | R A D |
| Urb U | R A D | R A D | R A D | - | R A D |
| Urb P | R A D | R A D | R A D | R A D | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisikirskienė, L.; Brazaitis, G.; Šimkevičius, K.; Brazaitytė, G. How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes? Forests 2023, 14, 2119. https://doi.org/10.3390/f14112119
Bisikirskienė L, Brazaitis G, Šimkevičius K, Brazaitytė G. How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes? Forests. 2023; 14(11):2119. https://doi.org/10.3390/f14112119
Chicago/Turabian StyleBisikirskienė, Loreta, Gediminas Brazaitis, Kastytis Šimkevičius, and Gailenė Brazaitytė. 2023. "How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes?" Forests 14, no. 11: 2119. https://doi.org/10.3390/f14112119
APA StyleBisikirskienė, L., Brazaitis, G., Šimkevičius, K., & Brazaitytė, G. (2023). How Are Urban Birds Affected by Surrounding Forests and Agricultural Landscapes? Forests, 14(11), 2119. https://doi.org/10.3390/f14112119