
Chemical Compositions of Walnut (Juglans Spp.) Oil: Combined Effects of Genetic and Climatic Factors

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Area
2.2. Fatty Acid Composition
2.3. Micronutrients and Secondary Metabolites Determination
2.4. Statistical Analysis
3. Results
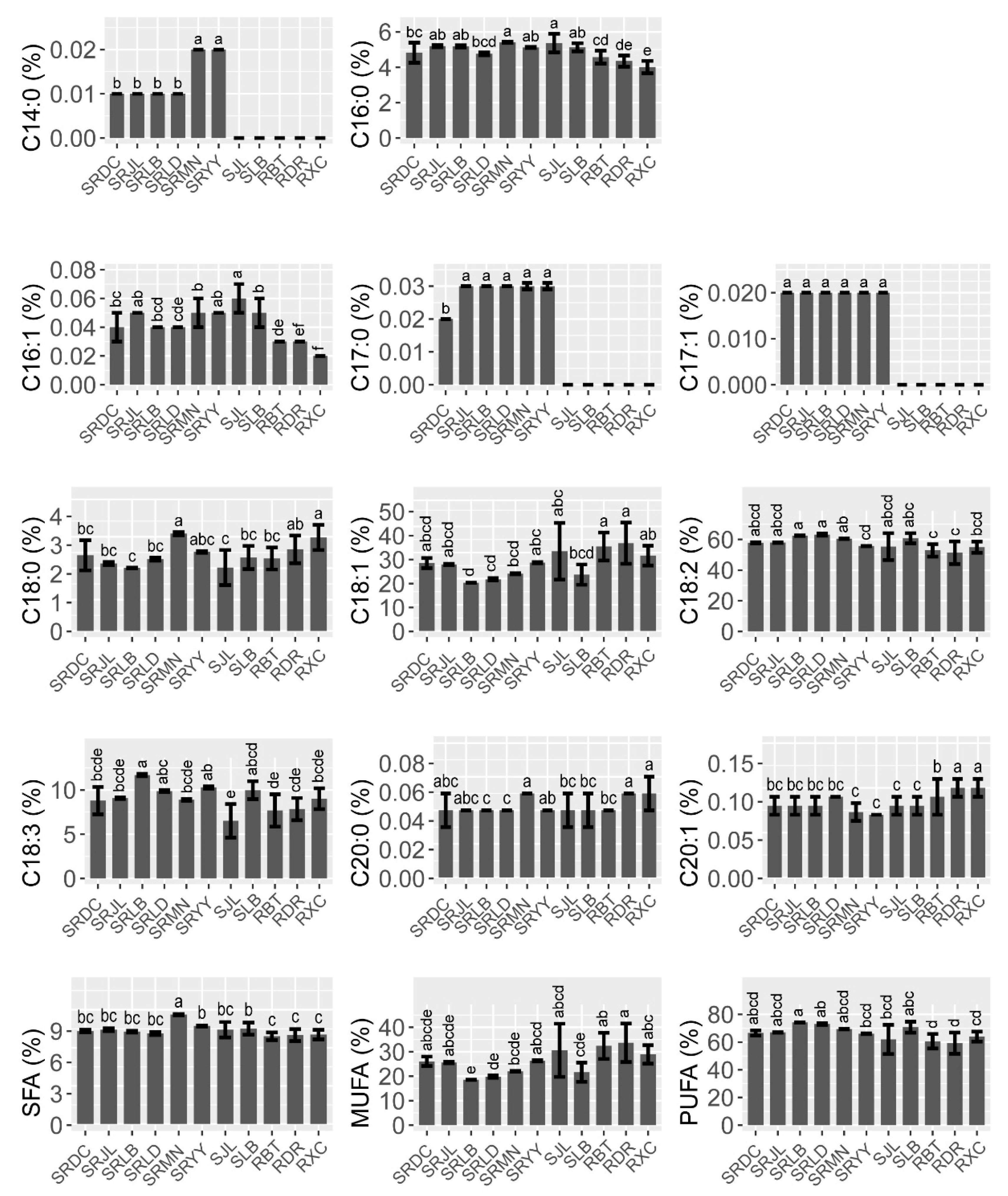
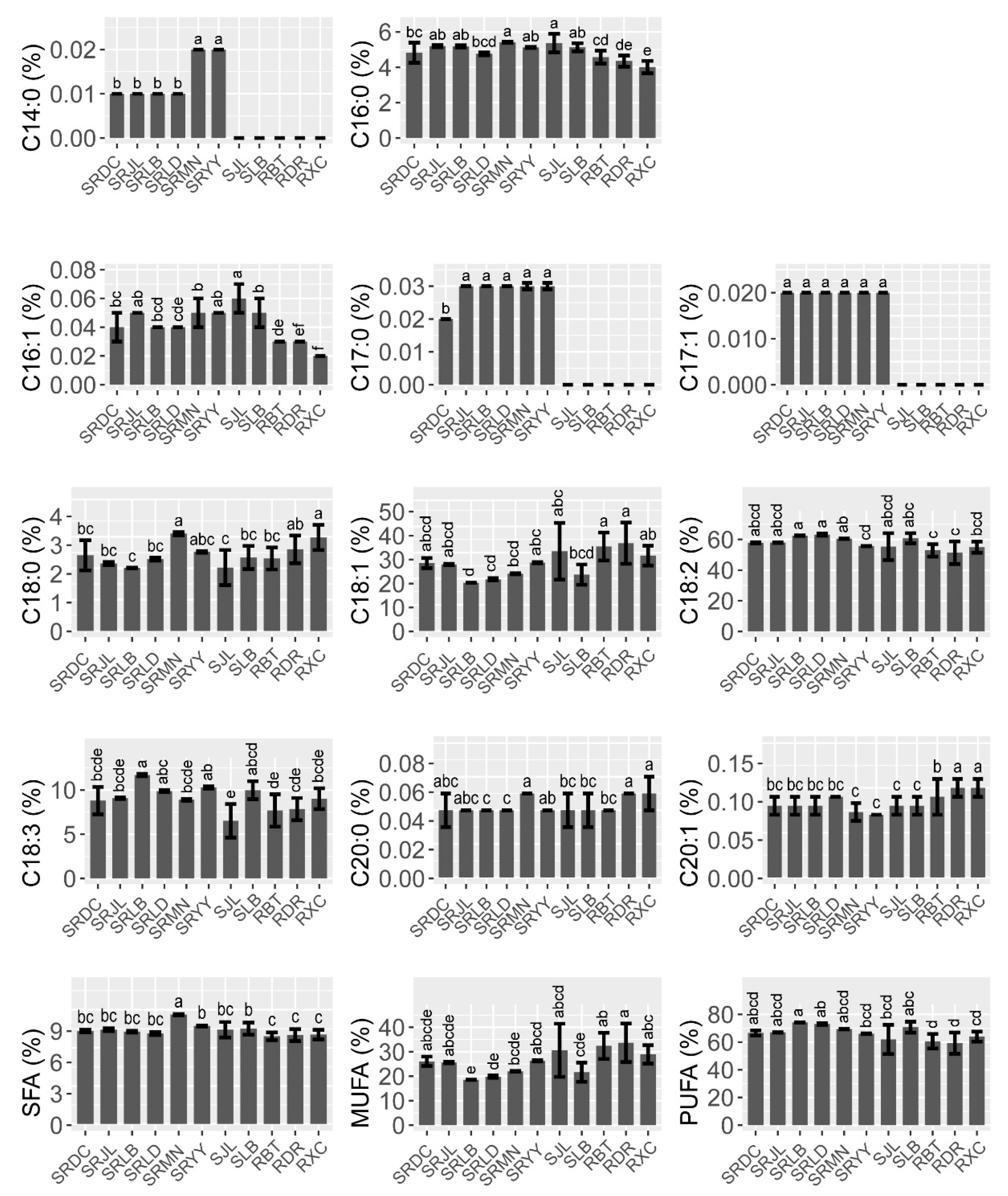
3.1. Fatty Acid Composition
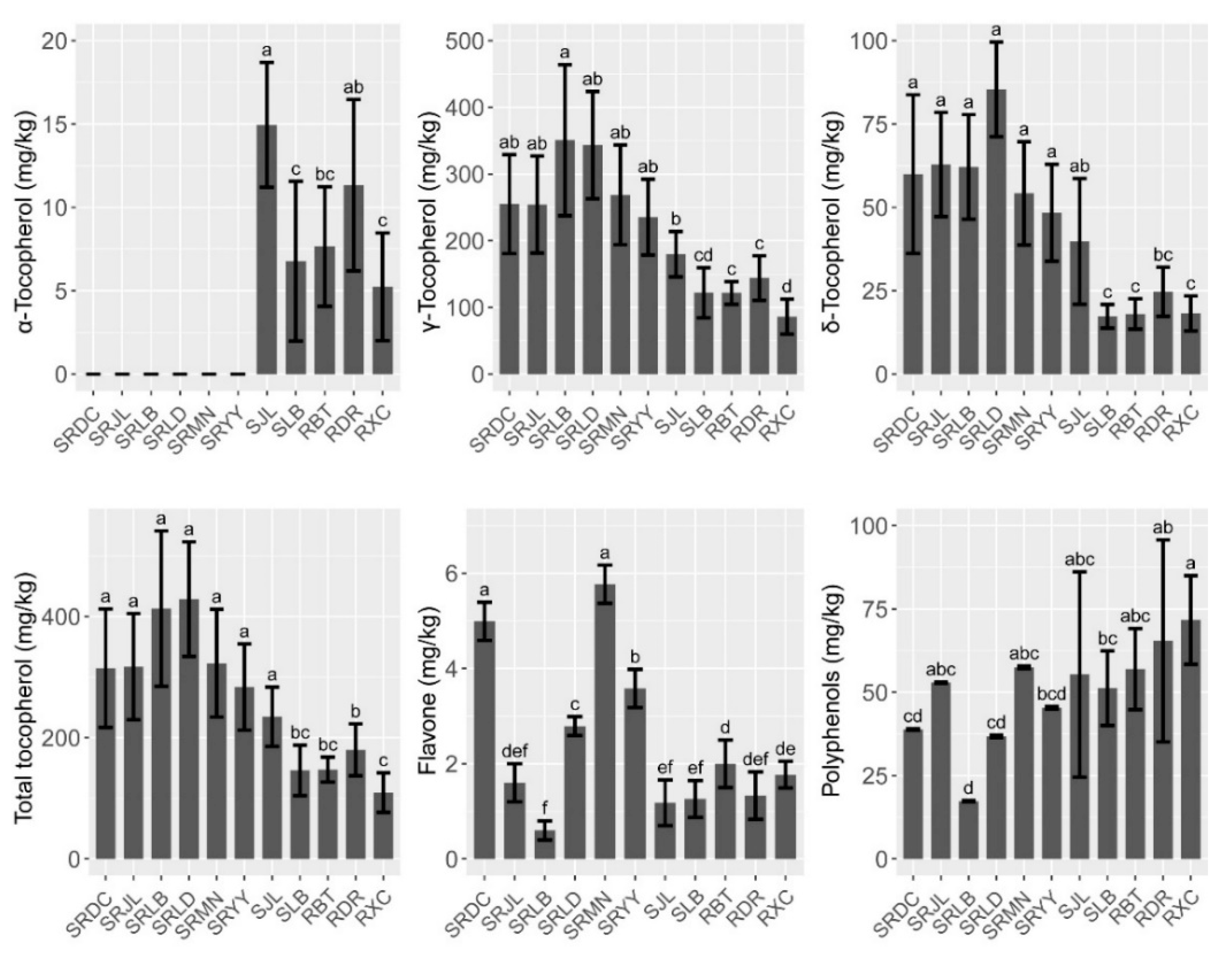
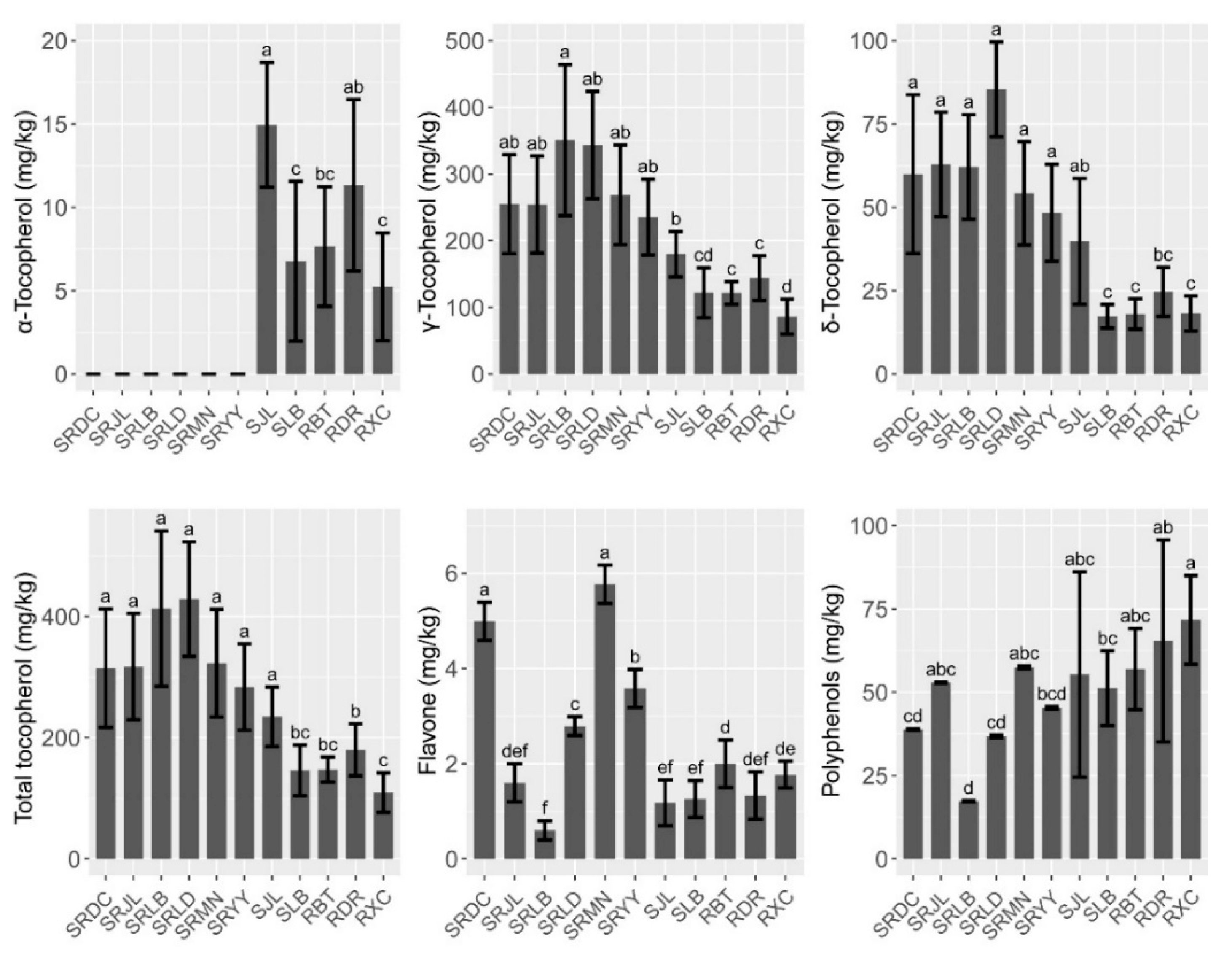
3.2. Micronutrient and Secondary Metabolites Levels
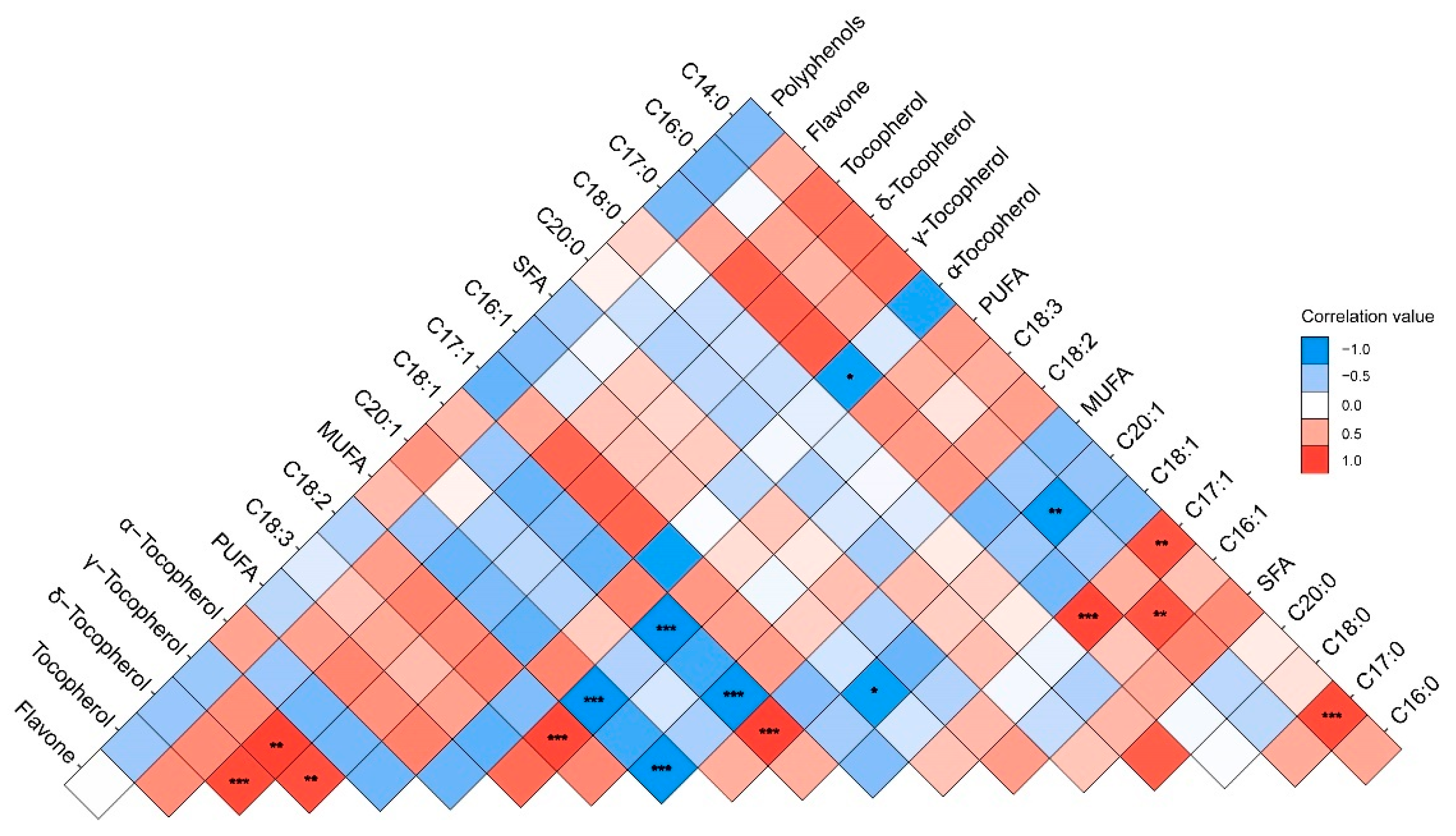
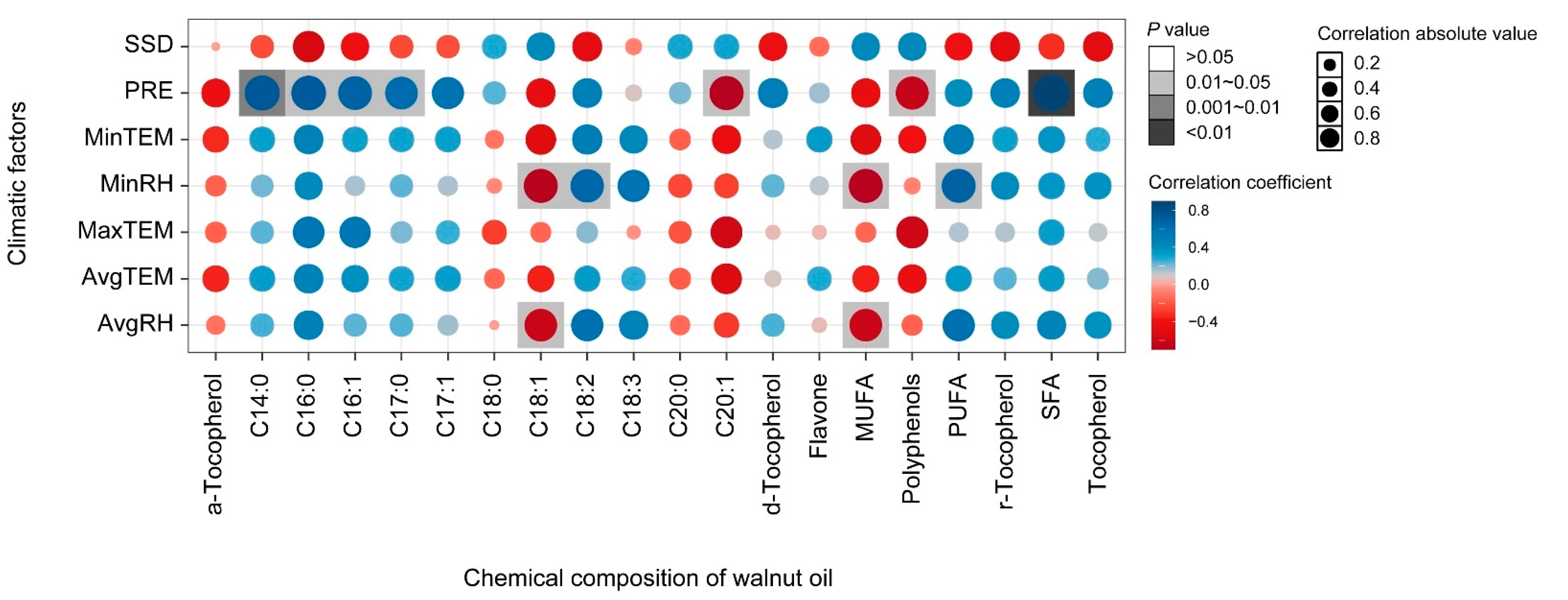
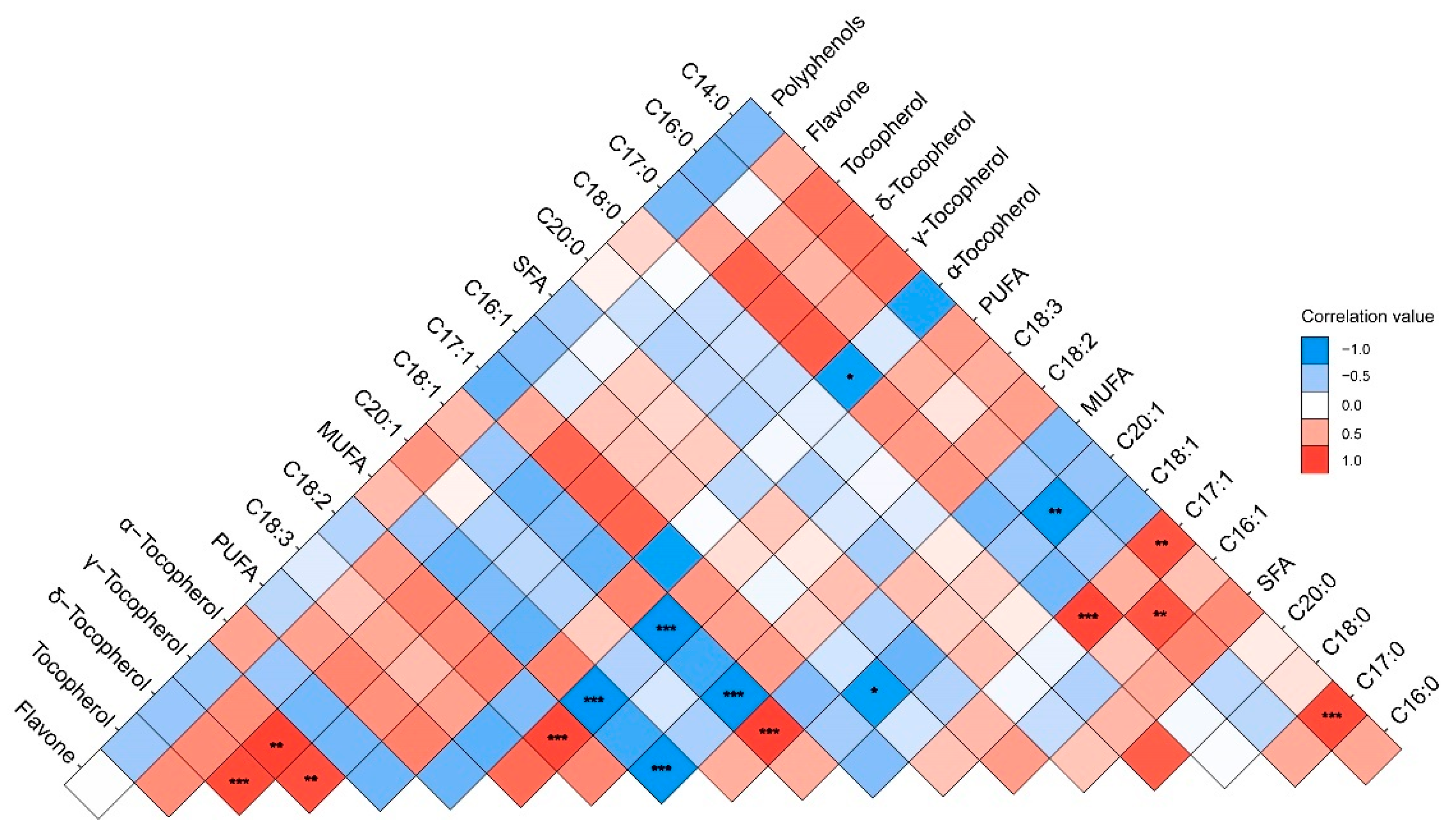
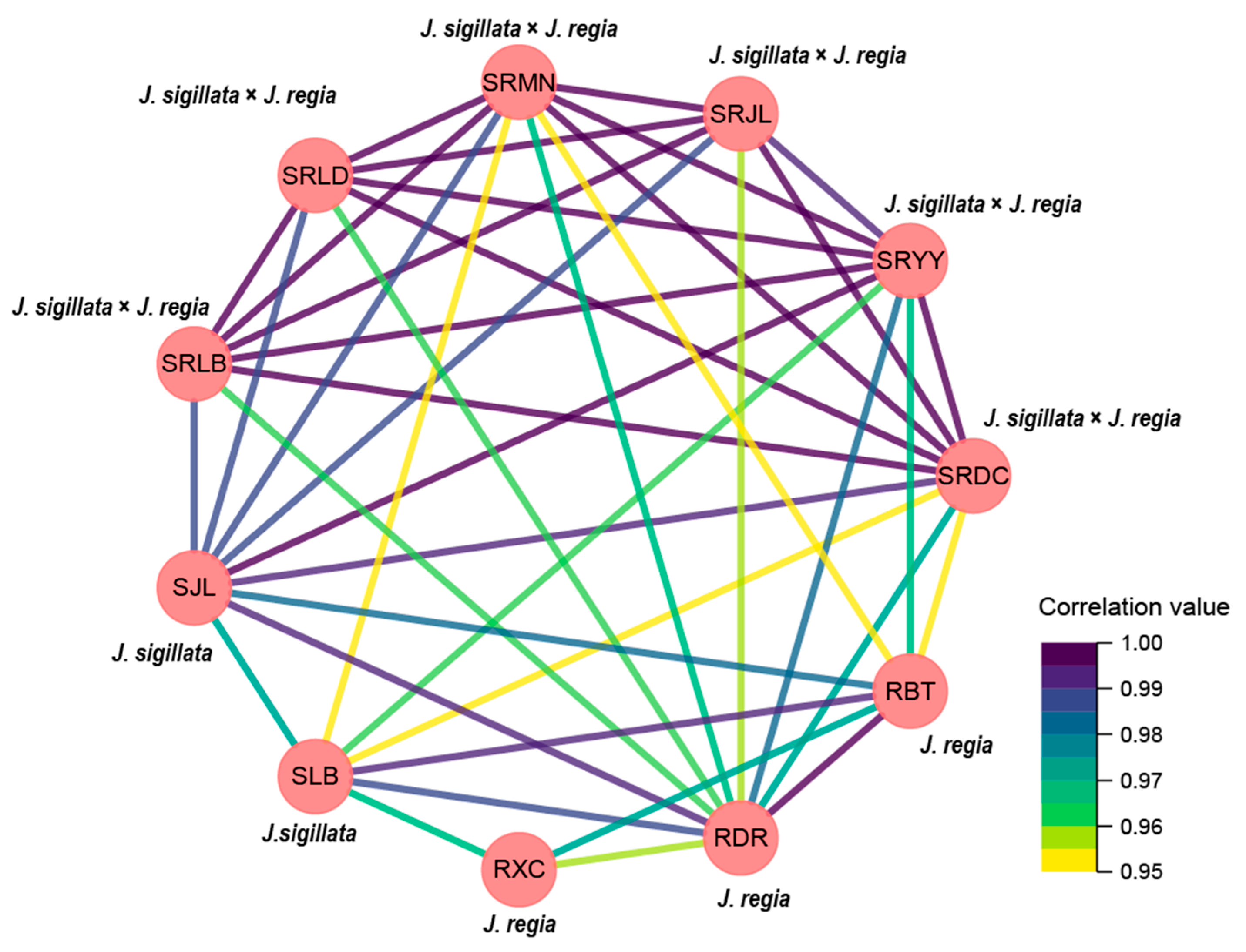
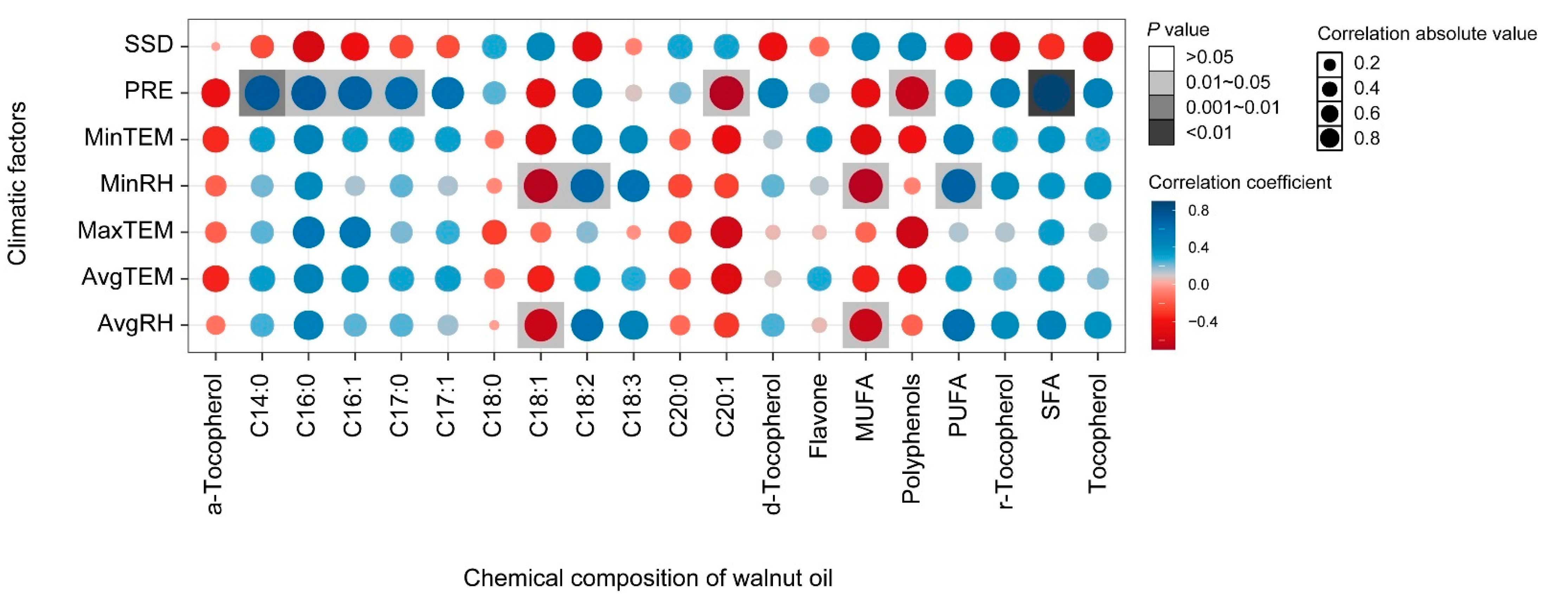
3.3. Correlation Analysis
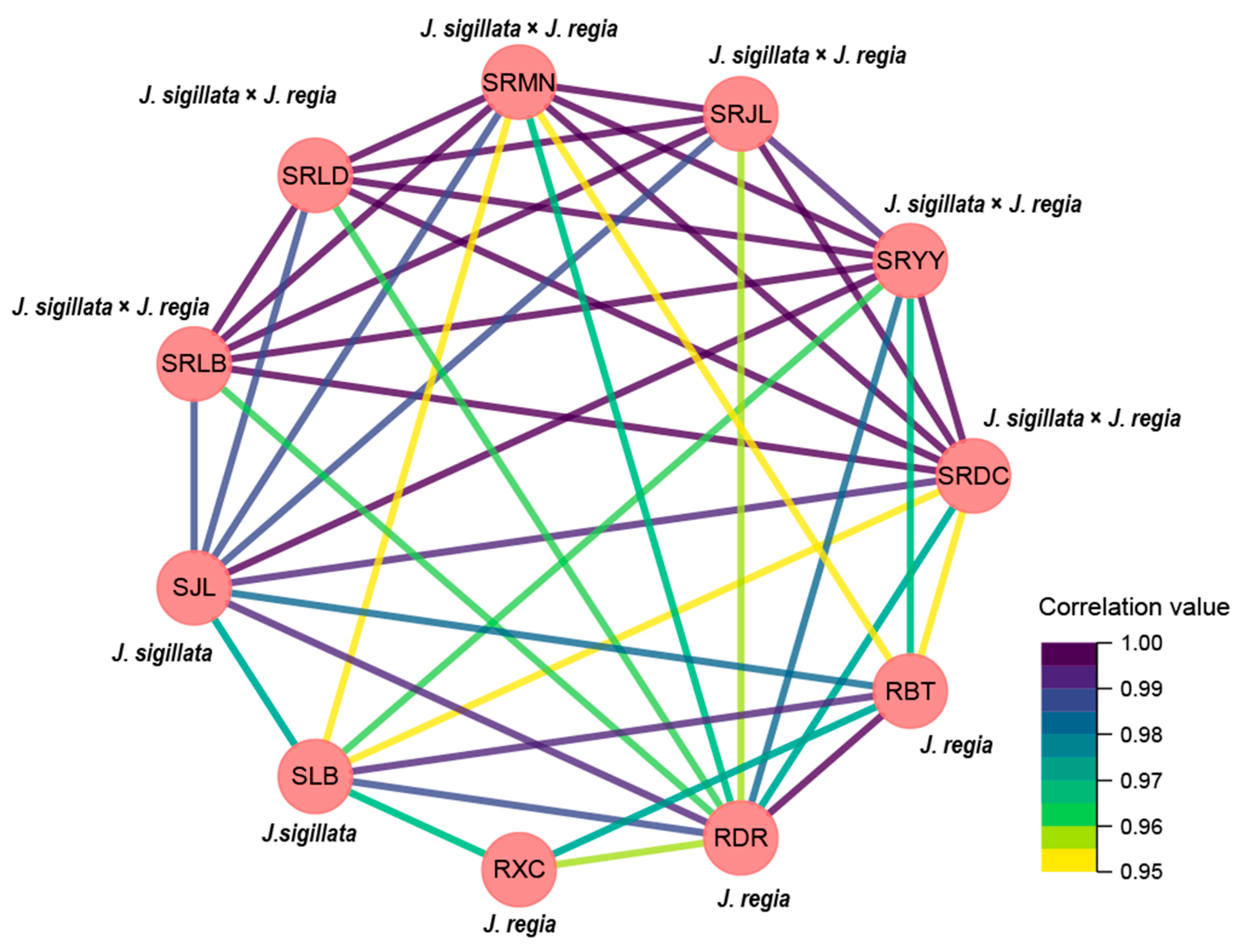
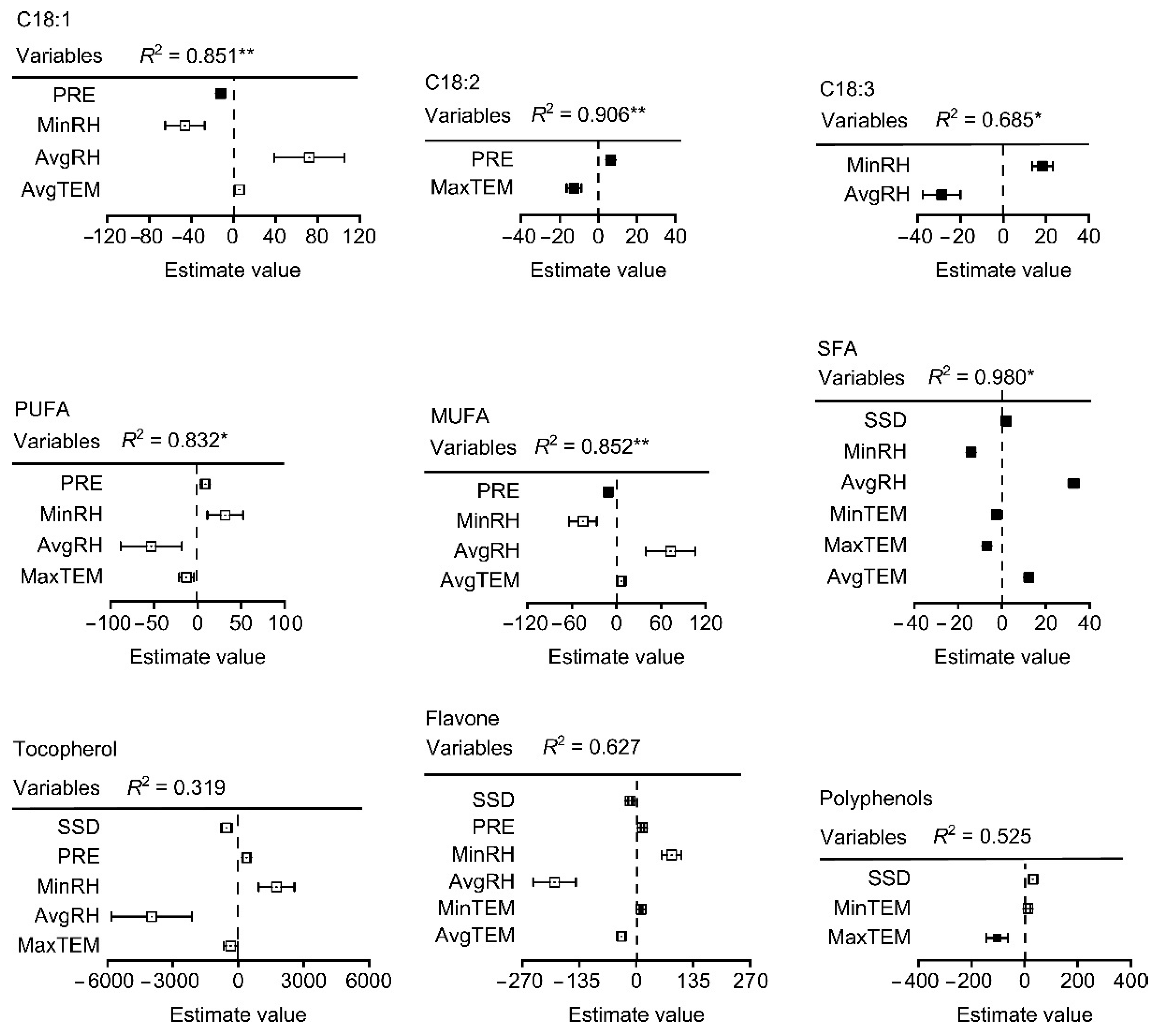
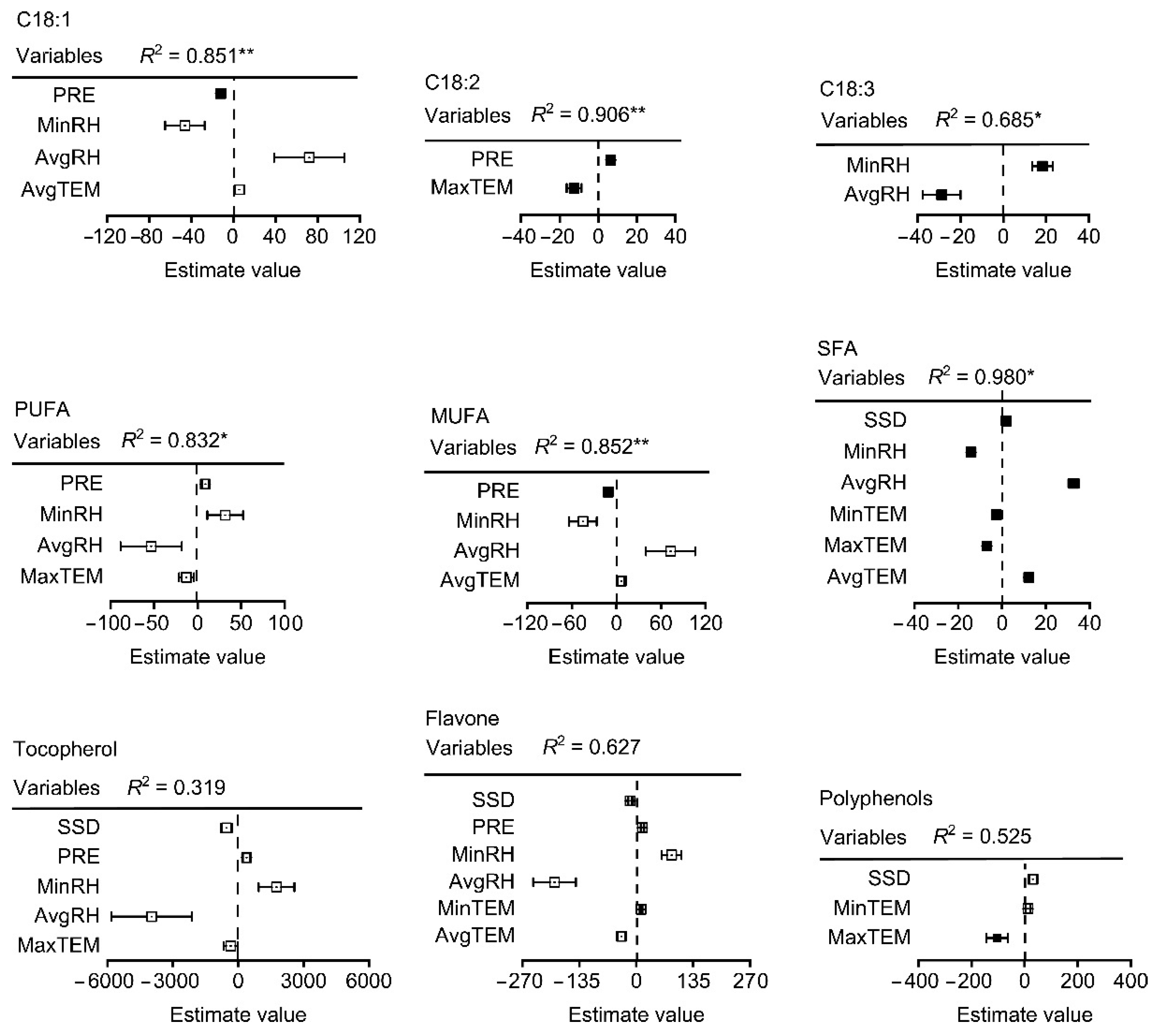
3.4. Climatic Factors and Fatty Acid Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gao, P.; Liu, R.; Jin, Q.; Wang, X. Comparison of different processing methods of Iron walnut oils (Juglans sigillata): Lipid yield, lipid compositions, minor components, and antioxidant capacity. Eur. J. Lipid Sci. Technol. 2018, 120, 1800151. [Google Scholar] [CrossRef]
- Gao, P.; Liu, R.; Jin, Q.; Wang, X. Comparative study of chemical compositions and antioxidant capacities of oils obtained from two species of walnut: Juglans regia and Juglans sigillata. Food Chem. 2019, 279, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Poggetti, L.; Ferfuia, C.; Chiabà, C.; Testolin, R.; Baldini, M. Kernel oil content and oil composition in walnut (Juglans regia L.) accessions from north-eastern Italy. J. Sci. Food Agric. 2018, 98, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.L.; Labuckas, D.O.; Lamarque, A.L.; Maestri, D.M. Walnut (Juglans regia L.): Genetic resources, chemistry, by-products. J. Sci. Food Agric. 2010, 90, 1959–1967. [Google Scholar] [CrossRef]
- Pereira, J.A.; Oliveira, I.; Sousa, A.; Ferreira, I.C.F.R.; Bento, A.; Estevinho, L. Bioactive properties and chemical composition of six walnut (Juglans regia L.) cultivars. Food Chem. Toxicol. 2008, 46, 2103–2111. [Google Scholar] [CrossRef]
- Gao, P.; Jin, J.; Liu, R.; Jin, Q.; Wang, X. Chemical compositions of walnut (Juglans regia L.) oils from different cultivated regions in China. J. Am. Oil Chem. Soc. 2018, 95, 825–834. [Google Scholar] [CrossRef]
- Anderson, K.J.; Teuber, S.S.; Gobeille, A.; Cremin, P.; Waterhouse, A.L.; Steinberg, F.M. Walnut polyphenolics inhibit in vitro human plasma and LDL oxidation. J. Nutr. 2001, 131, 2837–2842. [Google Scholar] [CrossRef]
- Park, S.-K.; Page, G.; Kim, K.; Allison, D.; Meydani, M.; Weindruch, R.; Prolla, T. Alpha- and gamma-tocopherol prevent age-related transcriptional alterations in the heart and brain of mice. J. Nutr. 2008, 138, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Kodad, O.; Socias, I.; Company, R. Variability of oil content and of major fatty acid composition in almond (Prunus amygdalus Batsch) and its relationship with kernel quality. J. Agric. Food Chem. 2008, 56, 4096–4101. [Google Scholar] [CrossRef]
- Kodad, O.; Estopañán, G.; Juan, T.; Mamouni, A.; Socias i Company, R. Tocopherol concentration in almond oil: Genetic variation and environmental effects under warm conditions. J. Agric. Food Chem. 2011, 59, 6137–6141. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.; Ning, D.; Ma, T.; Chen, H.; Zhang, Y.; Sun, X. Comprehensive analysis of the components of walnut kernel (Juglans regia L.) in China. J. Food Qual. 2021, 2021, 9302181. [Google Scholar] [CrossRef]
- Crews, C.; Hough, P.; Godward, J.; Brereton, P.; Lees, M.; Guiet, S.; Winkelmann, W. Study of the main constituents of some authentic hazelnut oils. J. Agric. Food Chem. 2005, 53, 4843–4852. [Google Scholar] [CrossRef] [PubMed]
- Delourme, R.; Falentin, C.; Huteau, V.; Clouet, V.; Horvais, R.; Gandon, B.; Specel, S.; Hanneton, L.; Dheu, J.E.; Deschamps, M.; et al. Genetic control of oil content in oilseed rape (Brassica napus L.). Theor. Appl. Genet. 2006, 113, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Zwarts, G.P.S.D.L.M.L. Fatty acid content of New Zealand-grown walnuts (Juglans regia L.). Int. J. Food Sci. Nutr. 1999, 50, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Canvin, D.T. The effect of temperature on the oil content and fatty acid composition of the oils from several oil seed crops. Can. J. Bot. 1965, 43, 63–69. [Google Scholar] [CrossRef]
- Byfield, G.E.; Upchurch, R.G. Effect of temperature on delta-9 stearoyl-ACP and microsomal omega-6 desaturase gene expression and fatty acid content in developing soybean seeds. Crop Sci. 2007, 47, 1698–1704. [Google Scholar] [CrossRef] [Green Version]
- Kodad, O.; Socias i Company, R.; Prats, M.S.; LÓpez Ortiz, M.C. Variability in tocopherol concentrations in almond oil and its use as a selection criterion in almond breeding. J. Hortic. Sci. Biotechnol. 2006, 81, 501–507. [Google Scholar] [CrossRef]
- Aradhya, M.K.; Potter, D.; Gao, F.; Simon, C.J. Molecular phylogeny of Juglans (Juglandaceae): A biogeographic perspective. Tree Genet. Genomes 2007, 3, 363–378. [Google Scholar] [CrossRef]
- Wuf, T.; Xiaof, L.; Chen, S.; Ning, D. Transcriptomics and comparative analysis of three Juglans species; J. regia, J. sigillata and J. cathayensis. Plant OMICS 2015, 8, 361–371. [Google Scholar]
- Li, H.; Sun, Q.; Zhao, S.; Zhang, W. Principle and Technology of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 184–185. [Google Scholar]
- Shi, C.; Chang, M.; Liu, R.; Jin, Q.; Wang, X. Trans-free shortenings through the interesterification of rice bran stearin, fully hydrogenated soybean oil and coconut oil. Int. J. Food Eng. 2015, 11, 467–477. [Google Scholar] [CrossRef]
- Greve, L.C.; McGranahan, G.; Hasey, J.; Snyde, R.; Kelly, K.; Goldhamer, D.; John, M.L. Variation in polyunsaturated fatty acids composition of persian walnut. Am. Soc. Hortic. Sci. 1992, 117, 518–522. [Google Scholar] [CrossRef]
- Ballus, C.A.; Meinhart, A.D.; de Souza Campos, F.A., Jr.; Godoy, H.T. Total phenolics of virgin olive oils highly correlate with the hydrogen atom transfer mechanism of antioxidant capacity. J. Am. Oil Chem. Soc. 2015, 92, 843–851. [Google Scholar] [CrossRef]
- Shi, L.-K.; Mao, J.-H.; Zheng, L.; Zhao, C.-W.; Jin, Q.-Z.; Wang, X.-G. Chemical characterization and free radical scavenging capacity of oils obtained from Torreya grandis Fort. ex. Lindl. and Torreya grandis Fort. var. Merrillii: A comparative study using chemometrics. Ind. Crops Prod. 2018, 115, 250–260. [Google Scholar] [CrossRef]
- Bada, J.C.; León-Camacho, M.; Prieto, M.; Copovi, P.; Alonso, L. Characterization of Walnut Oils (Juglans regia L.) from Asturias, Spain. J. Am. Oil Chem. Soc. 2010, 87, 1469–1474. [Google Scholar] [CrossRef]
- Vuorinen, A.L.; Markkinen, N.; Kalpio, M.; Linderborg, K.M.; Yang, B.; Kallio, H.P. Effect of growth environment on the gene expression and lipids related to triacylglycerol biosynthesis in sea buckthorn (Hippophaë rhamnoides) berries. Food Res. Int. 2015, 77, 608–619. [Google Scholar] [CrossRef]
- Amaral, J.S.; Alves, M.R.; Seabra, R.M.; Oliveira, B.P.P. Vitamin E composition of walnuts (Juglans regia L.): A 3-year comparative study of different cultivars. J. Agric. Food Chem. 2005, 53, 5467–5472. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Mousavi, S.M.; Hamedi, M.; Khodaiyan, F. Determination and characterization of kernel biochemical composition and functional compounds of persian walnut oil. J. Food Sci. Technol. 2014, 51, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Slatnar, A.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Solar, A. Identification and quantification of phenolic compounds in kernels, oil and bagasse pellets of common walnut (Juglans regia L.). Food Res. Int. 2015, 67, 255–263. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation Site | Species | Longitude/° | Latitude/° | Altitude/m |
|---|---|---|---|---|
| Batang, Ganzi, China (RBT) | Common walnut (J. regia L.) | 99.013 | 29.778 | 2493 |
| Derong, Ganzi, China (RDR) | Common walnut (J. regia L.) | 99.376 | 29.047 | 3196 |
| Jiulong, Ganzi, China (SJL) | Iron walnut (J. sigillata Dode) | 101.723 | 28.528 | 2304 |
| Jiulong, Ganzi, China (SRJL) | Hybrids (J. sigillata Dode × J. regia L.) | |||
| Leibo, Liangshan, China (SLB) | Iron walnut (J. sigillata Dode) | 103.434 | 28.258 | 989 |
| Leibo, Liangshan, China (SRLB) | Hybrids (J. sigillata Dode × J. regia L.) | |||
| Xiangcheng, Ganzi, China (RXC) | Common walnut (J. regia L.) | 99.467 | 29.087 | 2814 |
| Dechang, Liangshan, China (SRDC) | Hybrids (J. sigillata Dode × J. regia L.) | 102.017 | 27.050 | 1596 |
| Luding, Ganzi, China (SRLD) | Hybrids (J. sigillata Dode × J. regia L.) | 102.017 | 29.050 | 1412 |
| Mianning, Liangshan, China (SRMN) | Hybrids (J. sigillata Dode × J. regia L.) | 102.683 | 28.033 | 2004 |
| Yanyuan, Liangshan, China (SRYY) | Hybrids (J. sigillata Dode × J. regia L.) | 101.050 | 27.033 | 2527 |
| Source | Sum of Square | Mean Square | F Value | Source | Sum of Square | Mean Square | F Value |
|---|---|---|---|---|---|---|---|
| C14:0 | C18:1 | ||||||
| Species | 4.51 × 10−4 | 4.51 × 10−4 | 3464.16 *** | Species | 19.50 | 19.50 | 0.474 |
| Sites | 1.40 × 10−6 | 1.40 × 10−6 | 10.74 * | Sites | 179.00 | 179.00 | 4.348 |
| Species × Sites | 1.40 × 10−6 | 1.40 × 10−6 | 10.74 * | Species × Sites | 1.50 | 1.50 | 0.037 |
| Residuals | 1.00 × 10−6 | 1.00 × 10−7 | Residuals | 330.00 | 41.20 | ||
| C16:0 | C20:1 | ||||||
| Species | 1.55 × 10−1 | 1.55 × 10−1 | 1.551 | Species | 1.96 × 10−4 | 1.96 × 10−4 | 1.69 |
| Sites | 2.64 × 10−1 | 2.64 × 10−1 | 2.654 | Sites | 3.10 × 10−6 | 3.10 × 10−6 | 0.027 |
| Species × Sites | 2.44 × 10−1 | 2.44 × 10−1 | 2.449 | Species × Sites | 2.59 × 10−5 | 2.59 × 10−5 | 0.223 |
| Residuals | 7.97 × 10−1 | 9.96 × 10−2 | Residuals | 9.28 × 10−4 | 1.16 × 10−4 | ||
| C18:0 | MUFA | ||||||
| Species | 5.20 × 10−3 | 5.20 × 10−6 | 0.073 | Species | 19.30 | 19.30 | 0.467 |
| Sites | 3.85 × 10−2 | 3.85 × 10−2 | 0.544 | Sites | 180.00 | 180.00 | 4.36 |
| Species × Sites | 1.82 × 10−1 | 1.82 × 10−1 | 2.569 | Species × Sites | 1.50 | 1.50 | 0.037 |
| Residuals | 5.66 × 10−1 | 7.08 × 10−2 | Residuals | 330.00 | 41.30 | ||
| C20:0 | C18:2 | ||||||
| Species | 9.18 × 10−6 | 9.18 × 10−6 | 0.317 | Species | 1.41 | 1.41 | 0.052 |
| Sites | 7.72 × 10−5 | 7.72 × 10−5 | 2.662 | Sites | 73.70 | 73.70 | 2.694 |
| Species × Sites | 2.64 × 10−6 | 2.64 × 10−6 | 0.091 | Species × Sites | 4.90 × 10−1 | 4.90 × 10−1 | 0.018 |
| Residuals | 2.32 × 10−4 | 2.90 × 10−5 | Residuals | 219.00 | 27.30 | ||
| SFA | C18:3 | ||||||
| Species | 6.32 × 10−2 | 6.32 × 10−2 | 2.425 | Species | 12.60 | 12.60 | 8.001 * |
| Sites | 1.07 × 10−1 | 1.07 × 10−1 | 4.12 | Sites | 26.60 | 26.60 | 16.921 ** |
| Species × Sites | 4.20 × 10−3 | 4.20 × 10−3 | 0.161 | Species × Sites | 3.76 × 10−1 | 3.76 × 10−1 | 0.239 |
| Residuals | 2.09 × 10−1 | 2.61 × 10−2 | Residuals | 12.60 | 1.57 | ||
| C16:1 | PUFA | ||||||
| Species | 2.98 × 10−4 | 2.98 × 10−4 | 3.516 | Species | 22.40 | 22.40 | 0.554 |
| Sites | 3.76 × 10−4 | 3.76 × 10−4 | 4.435 | Sites | 189.00 | 189.00 | 4.666 |
| Species × Sites | 3.74 × 10−5 | 3.74 × 10−5 | 0.442 | Species × Sites | 1.70 | 1.70 | 0.043 |
| Residuals | 6.77 × 10−4 | 8.47 × 10−5 | Residuals | 324.00 | 40.50 | ||
| C17:1 | |||||||
| Species | 1.14 × 10−3 | 1.14 × 10−3 | 607.382 *** | ||||
| Sites | 8.20 × 10−6 | 8.20 × 10−6 | 4.375 | ||||
| Species × Sites | 8.20 × 10−6 | 8.20 × 10−6 | 4.375 | ||||
| Residuals | 1.50 × 10−5 | 1.90 × 10−6 |
| Source | Sum of Square | Mean Square | F Value |
|---|---|---|---|
| α-tocopherol | |||
| Species | 47,926 | 47,926 | 9.432 * |
| Sites | 1561 | 1561 | 0.307 |
| Species × Sites | 16,301 | 16,301 | 3.208 |
| Residuals | 40,648 | 5081 | |
| δ-tocopherol | |||
| Species | 3029.9 | 3029.9 | 18.279 ** |
| Sites | 344.5 | 344.5 | 2.078 |
| Species × Sites | 300.8 | 300.8 | 1.815 |
| Residuals | 1326.1 | 165.7625 | |
| Tocopherol | |||
| Species | 61,329 | 61,329 | 8.943 * |
| Sites | 219 | 219 | 0.032 |
| Species × Sites | 22,853 | 22,853 | 3.332 |
| Residuals | 54,864 | 6858 | |
| Flavone | |||
| Species | 0.0312 | 0.0312 | 0.419 |
| Sites | 0.2115 | 0.2115 | 2.842 |
| Species × Sites | 1.6337 | 1.6337 | 21.950 ** |
| Residuals | 0.5958 | 0.0744 | |
| Polyphenols | |||
| Species | 182.1 | 182.1 | 2.823 |
| Sites | 745.1 | 745.1 | 11.550 ** |
| Species × Sites | 1176.7 | 1176.7 | 18.239 ** |
| Residuals | 516.1 | 64.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Xiao, X.; Li, J.; Wang, F.; Mi, J.; Shi, Y.; He, F.; Chen, L.; Zhang, F.; Wan, X. Chemical Compositions of Walnut (Juglans Spp.) Oil: Combined Effects of Genetic and Climatic Factors. Forests 2022, 13, 962. https://doi.org/10.3390/f13060962
Yang H, Xiao X, Li J, Wang F, Mi J, Shi Y, He F, Chen L, Zhang F, Wan X. Chemical Compositions of Walnut (Juglans Spp.) Oil: Combined Effects of Genetic and Climatic Factors. Forests. 2022; 13(6):962. https://doi.org/10.3390/f13060962
Chicago/Turabian StyleYang, Hanbo, Xu Xiao, Jingjing Li, Fang Wang, Jiaxuan Mi, Yujie Shi, Fang He, Lianghua Chen, Fan Zhang, and Xueqin Wan. 2022. "Chemical Compositions of Walnut (Juglans Spp.) Oil: Combined Effects of Genetic and Climatic Factors" Forests 13, no. 6: 962. https://doi.org/10.3390/f13060962
APA StyleYang, H., Xiao, X., Li, J., Wang, F., Mi, J., Shi, Y., He, F., Chen, L., Zhang, F., & Wan, X. (2022). Chemical Compositions of Walnut (Juglans Spp.) Oil: Combined Effects of Genetic and Climatic Factors. Forests, 13(6), 962. https://doi.org/10.3390/f13060962