
Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China



Abstract
:1. Introduction
2. Materials and Methods
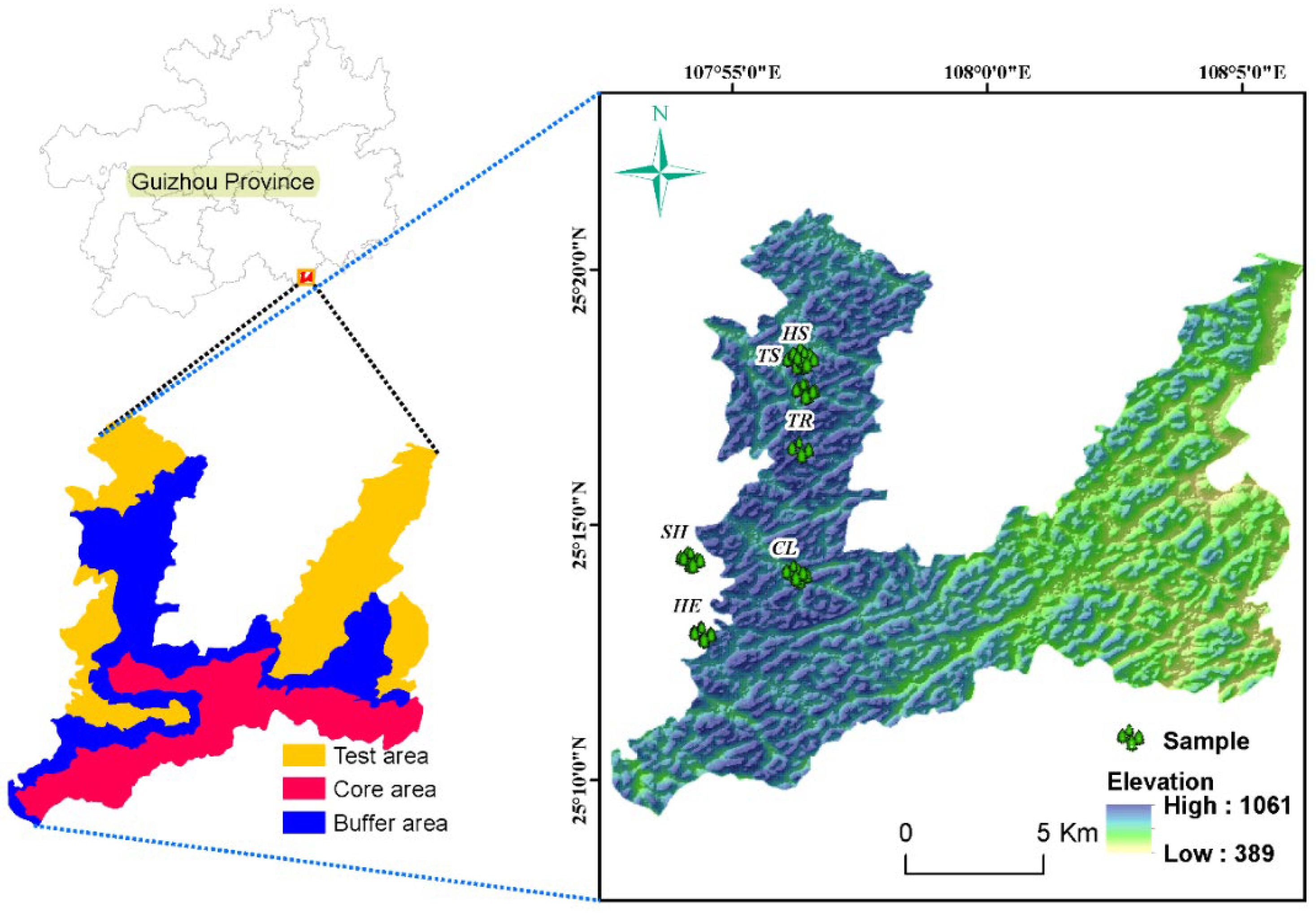
2.1. Overview of the Study Area
2.2. Sample Plot, Sample Square Setting and Plant Community Survey
2.3. Sample Collection and Determination
2.4. Functional Trait Selection and Sample Analysis
2.5. Data Processing and Analysis
2.5.1. Calculation of Community-Weighted Functional Traits
2.5.2. Functional Diversity Calculation
3. Results
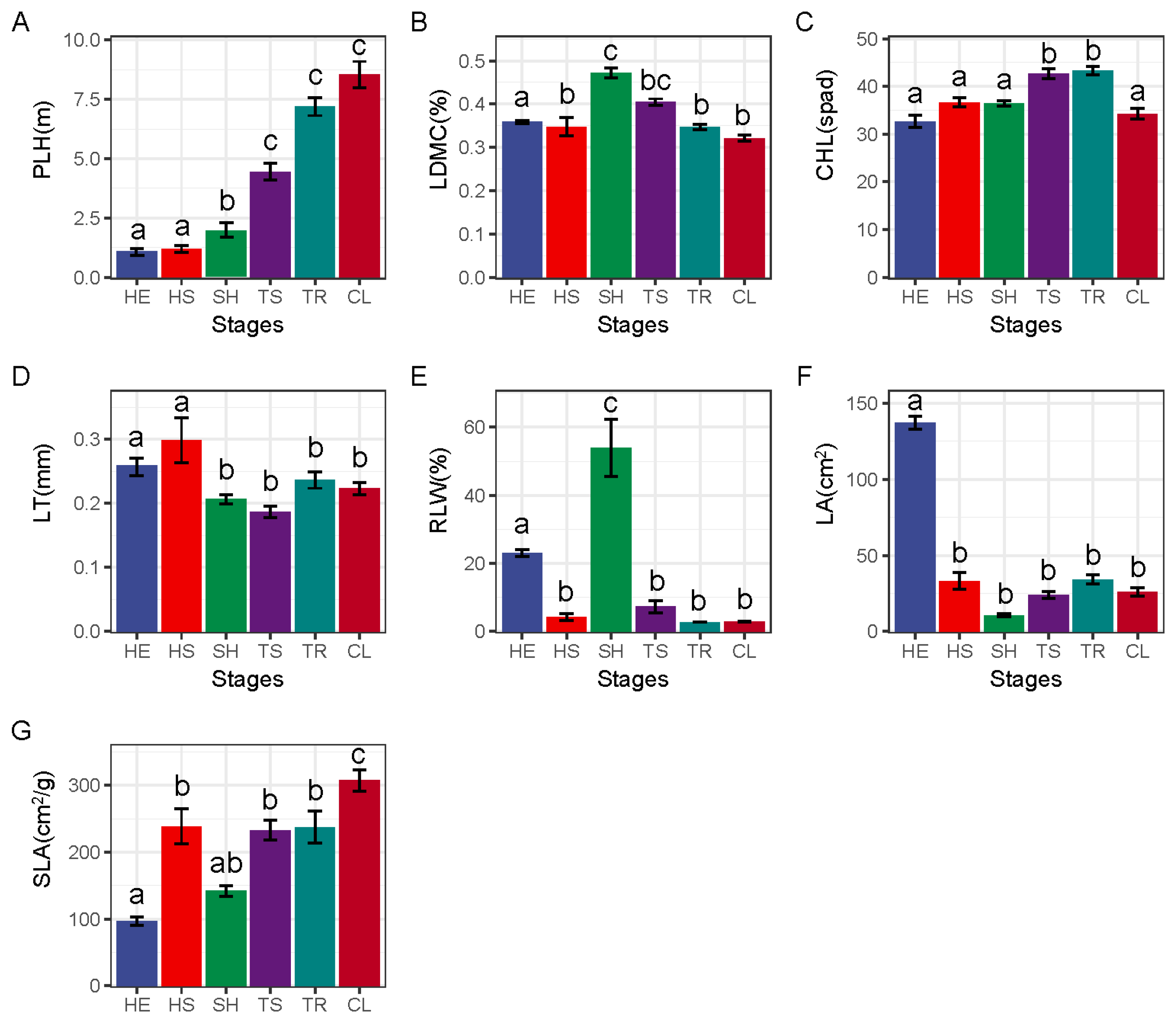
3.1. Weighted Functional Traits of Communities at Different Restoration Stages
3.2. Functional Diversity of Forest Plant Communities at Different Restoration Stages
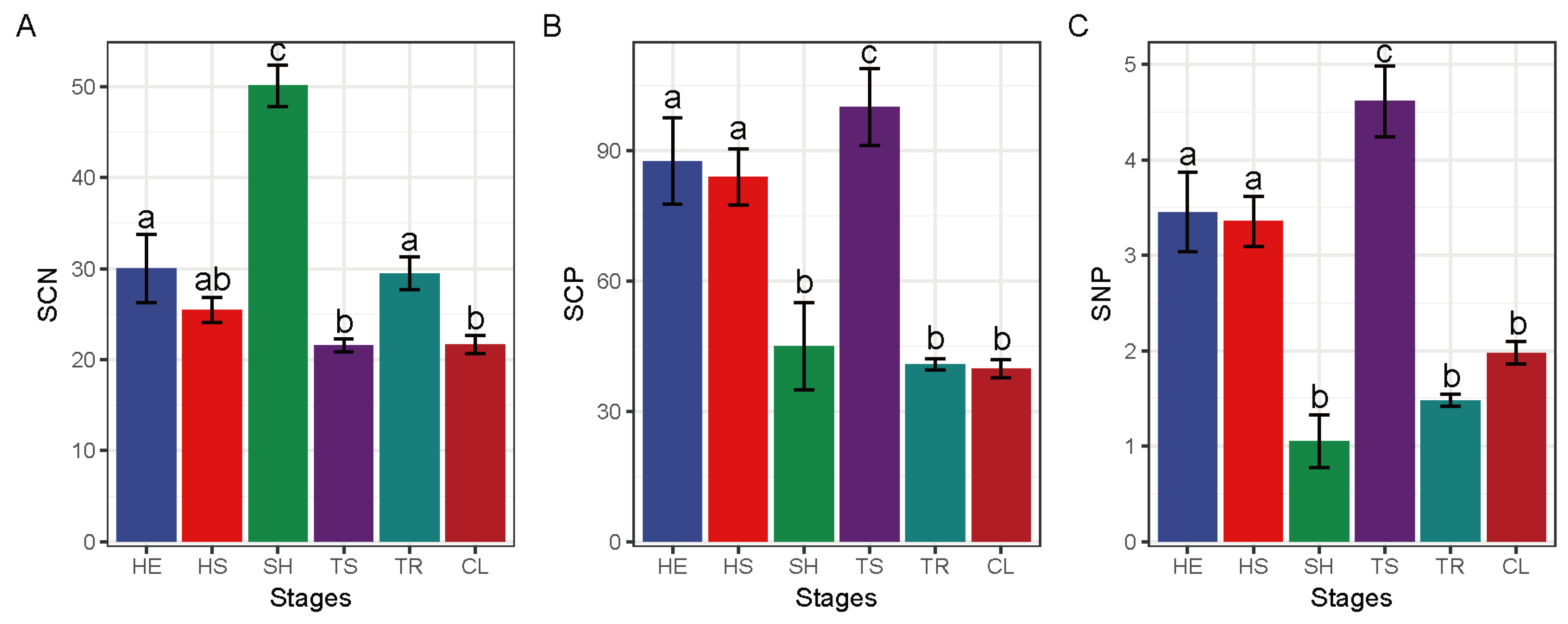
3.3. Changes in Leaf and Soil Nutrient Contents at Different Recovery Stages
3.4. Leaf and Soil Eco-Ecological Stoichiometry Characteristics at Different Restoration Stages
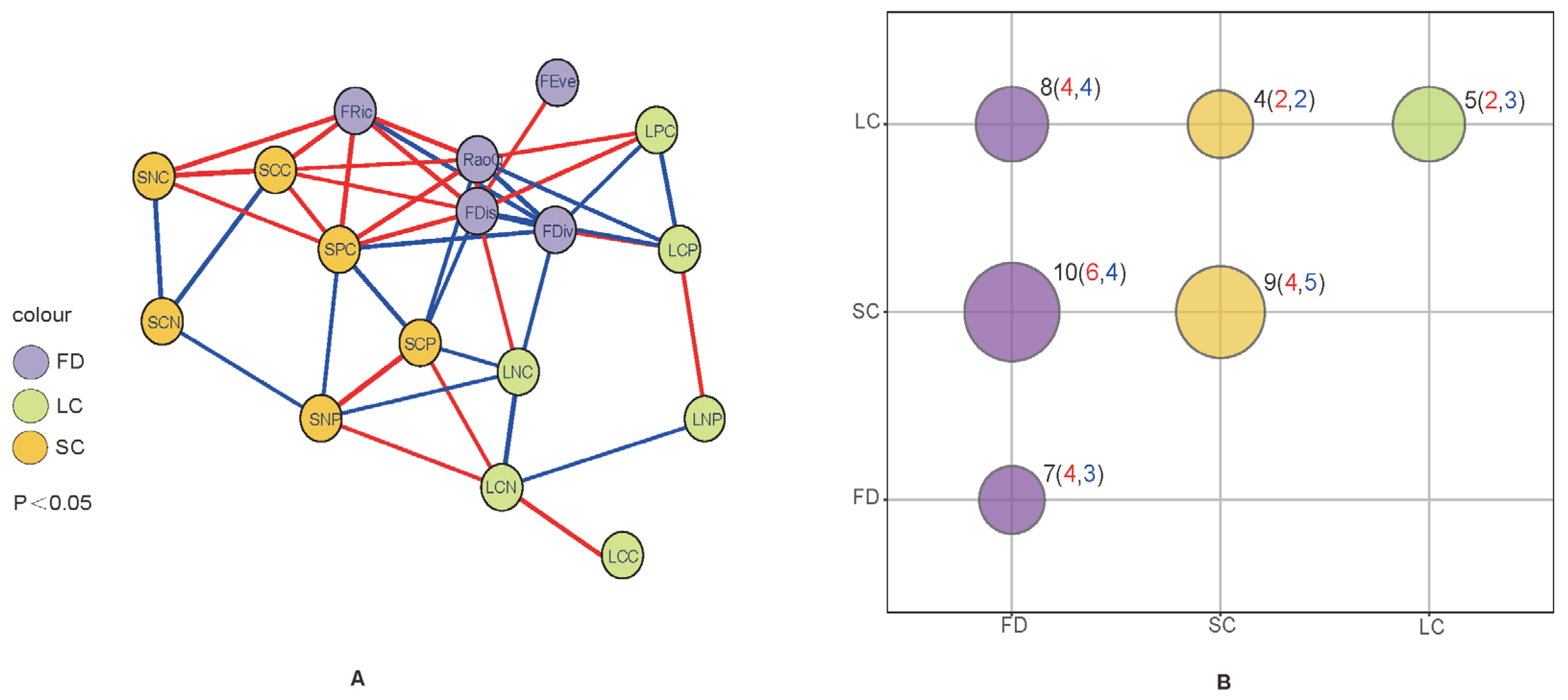
3.5. Network Relationships between the Functional Diversity of Plant Communities and Leaf and Soil Chemical Properties
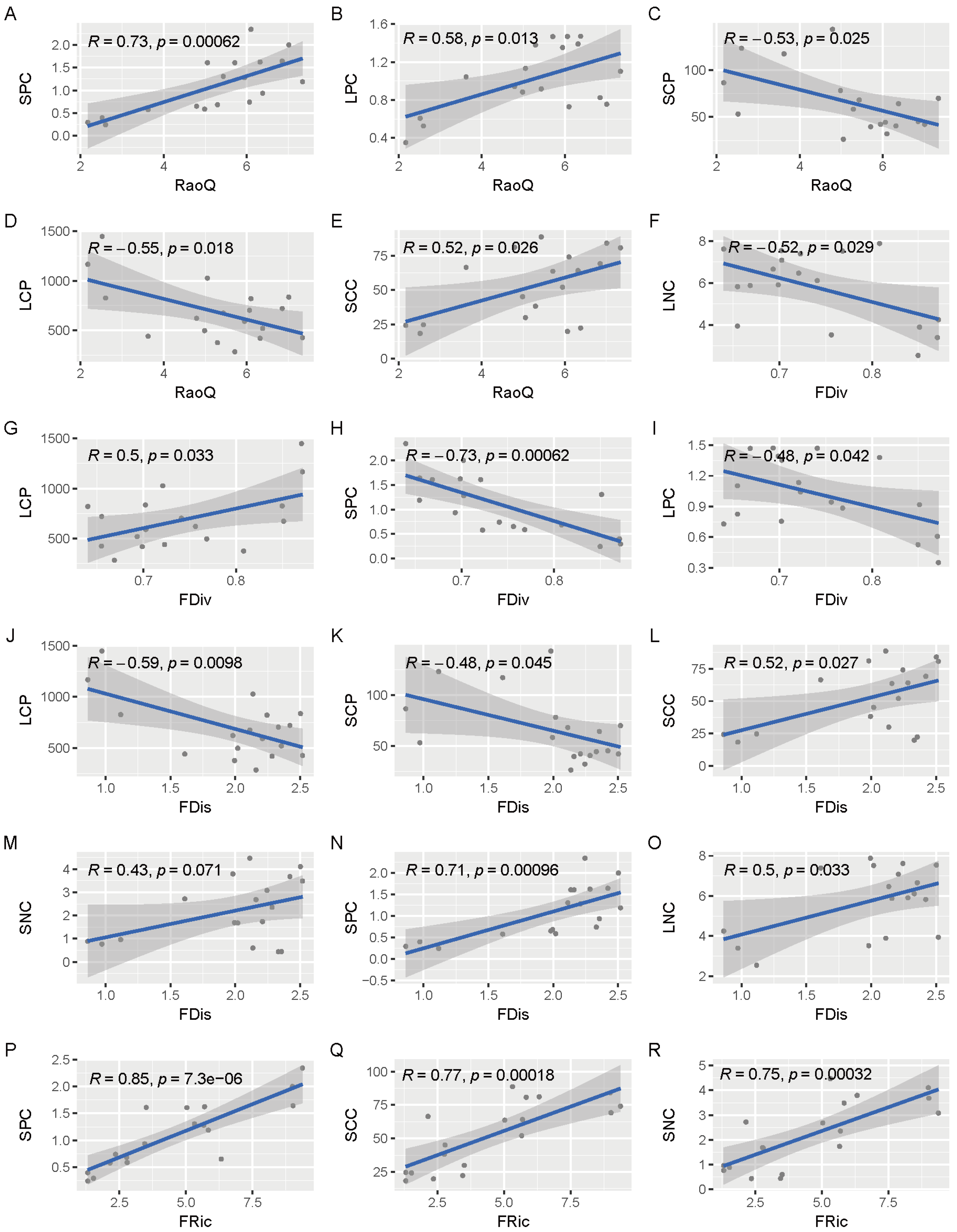
3.6. Correlation Analysis of Functional Diversity of Plant Communities with Leaf and Soil Chemical Properties
4. Discussion
4.1. Response of Community-Weighted Functional Traits to Different Recovery Stages
4.2. Changes in Functional Diversity Patterns at Different Restoration Stages
4.3. Response of Community Functional Diversity to Leaf and Soil Chemical Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, R.X.; Ding, Y.; Ma, W.J.; Niu, J.M.; Zhang, Q. Research advances in plant functional diversity and its relationship with ecosystem functions. Ecol. Environ. Sci. 2016, 25, 1069–1075. [Google Scholar]
- Biswas, S.; Mallik, A.U.; Braithwaite, N.T.; Biswas, P.L. Effects of disturbance type and microhabitat on species and functional diversity relationship in stream-bank plant communities. For. Ecol. Manag. 2019, 432, 812–822. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Diaz, S.; Lavorel, S.; de Bello, F.; Quetier, F.; Grigulis, K.; Robson, T.M. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. USA 2007, 104, 20684–20689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillebrand, H.; Matthiessen, B. Biodiversity in a complex world: Consolidation and progress in functional biodiversity research. Ecol. Lett. 2009, 12, 1405–1419. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Petchey, O.L.; Gaston, K.J. Functional Diversity(FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Zhang, J.T.; Fan, L.H. Functional diversity of species and its research methods. J. Mt. Sci. 2011, 29, 513–519. [Google Scholar]
- Lepě, J.; De, B.F.; Lavorel, S. Quantifying and interpreting functional diversity of natural communities: Practical considerrations matter. Preslia 2006, 27, 1254–1261. [Google Scholar]
- Mcgill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Wiegand, T.; Uriate, M.; Kraft, N.J.B.; Shen, G.C.; Wang, X.G.; He, F.L. Spatially explicit metrics of species diversity, functional diversity, and phylogenetic diversity: Insights into plant community assembly processes. Annu. Rev. Ecol. Evol. 2017, 48, 329–351. [Google Scholar] [CrossRef] [Green Version]
- Khalil, M.I.; Gibson, D.J.; Baer, S.G.; Willand, J.E. Functional diversity is more sensitive to biotic filters than phylogenetic diversity during community assembly. Ecosphere 2018, 9, e2164. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. Causes, consequences and ethics of biodiversity. Nature 2000, 405, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcin, K.D.; Natalia, C.; Mateusz, Z.; Jaroslaw, T.; Andrzej, M.J. Functional diversity, succession, and human-mediated disturbances in raised bog vegetation. Sci. Total Environ. 2016, 562, 648–657. [Google Scholar]
- Jesse, E.D.M.; Dai, J.L.; Marina, L.F.; Susan, H. Functional diversity is a passenger but not driver of drought related plant diversity losses in annual grasslands. J. Ecol. 2019, 107, 2033–2039. [Google Scholar]
- Yao, X.Y.; Hu, Y.S.; Liu, Y.H. Plant functional traits and functional diversities of different communities in broad-leaved Korean pine forests in the Changbai Mountain. Nat. Sci. Ed. 2014, 3, 77–84. [Google Scholar]
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Hulot, F.D.; Lacroix, G.; Lescher-Moutoué, F.; Loreau, M. Functional diversity governs ecosystem response to nutrient enrichment. Nature 2000, 405, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Heemsbergen, D.A.; Berg, M.P.; Loreau, M.; van Hal, J.R.; Faber, J.H.; Verhoef, H.A. Biodiversity effects on soil processes explained by interspecific functional dissimilarity. Science 2004, 306, 1019–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokany, K.; Ash, J.; Roxburgh, S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. J. Ecol. 2008, 96, 884–893. [Google Scholar] [CrossRef]
- Milcu, A.; Roscher, C.; Gessler, A.; Bachmann, D.; Gockele, A.; Guderle, M.; Landais, D.; Piel, C.; Escape, C.; Devidal, S.; et al. Functional diversity of leaf nitrogen concentrations drives grassland carbon fluxes. Ecol. Lett. 2014, 17, 435–444. [Google Scholar] [CrossRef]
- He, J.S.; Han, X.G. Ecological stoichiometry: Searching for unifying principle from individuals to ecosystems. Chin. J. Plan Ecol. 2010, 34, 2–6. [Google Scholar]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.J. An outline of karst geomorphology zoning in the karst areas of Southern China. Mt. Res. 2003, 33, 120–126. [Google Scholar]
- Wang, L.J.; Wang, P.; Sheng, M.Y. Stoichiometry characteristics of soil nutrient elements and its influencing factors in typical in karst rocky desertification ecosystems, Southwest China. Acta Ecol. Sin. 2018, 38, 6580–6593. [Google Scholar]
- Yang, D.L.; Yu, Y.H.; Qin, S.Y.; Zhong, X.P. Contents and ecological stoichiometry characteristics of soil nutrients under different land utilization types in stony desertification area. Southwest Chin. J. Agric. Sci. 2018, 31, 1875–1881. [Google Scholar]
- Pi, F.J.; Yuan, C.J.; Yu, L.F.; Yan, L.B.; Wu, L.; Yang, R. Ecological stoichiometry characteristics of plant leaves from the main dominant species of natural secondary forest in the Central of Guizhou. Ecol. Environ. Sci. 2016, 25, 801–807. [Google Scholar]
- Liu, N.; Yu, L.F.; Zhao, Q.; Wu, Y.N.; Yan, L.B. C: N: P stoichiometry of leaf-litter-soil continuum in secondary forests of the rocky desertification regions of the karst plateau. Chin. J. Appl. Environ. Biol. 2020, 26, 681–688. [Google Scholar]
- Vannoppen, W.; Vanmaercke, M.; De Baets, S.; Poesen, J. A review of the mechanical effects of plant roots on concentrated flow erosion rates. Earth Sci. Rev. 2015, 150, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Yan, E.R.; Chang, S.X.; Cheng, J.Y.; Liu, X.Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Sci. Total Environ. 2017, 574, 654–662. [Google Scholar] [CrossRef]
- Yu, Y.H.; Zhong, X.P.; Zheng, W.; Chen, Z.X.; Wang, J.X. Species diversity, functional traits, stoichiometry and correlation of plant communities in different succession stages of karst forest. Acta Ecol. Sin. 2021, 41, 2408–2417. [Google Scholar]
- Zhu, S.Q. Study on Karst Forest Ecology-III; Guizhou Science and Technology Press: Guiyang, China, 2003; pp. 25–122. [Google Scholar]
- Yu, L.F.; Zhu, S.Q.; Ye, J.C.; Wei, L.M.; Chen, Z.R. Evaluation of natural restoration of degraded karst forest. Sci. Silvae Sin. 2000, 36, 12–19. [Google Scholar]
- Zhang, Z.H.; Hu, G.; Ni, J. Effects of Topographical and Edaphic Factors on the Distribution of Plant Communities in two Subtropical Karst Forests, Southwestern China. J. Mt. Sci. 2013, 10, 95–104. [Google Scholar] [CrossRef]
- An, M.T. Studies on Maintenance Mechanism of Plant Species Diversity and Soil Moisture and Nutrient Pattern in Karst Forest. Ph.D. Thesis, Guizhou University, Guiyang, China, 2019. [Google Scholar]
- Li, Y.K. Methods for Conventional Analysis of Soil Agrochemistry; Science Press: Beijing, China, 1989; pp. 1–195. [Google Scholar]
- Pérezharguindeguy, N.; Díaz, S.; Garnier, E. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Lavorel, S.; Grigulis, K.; Mcintyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field-methodology matters. Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Wu, Y.H.; Wang, Q.; Ji, L.B.; Huang, L.J. Effects of environmental factors on stem and leaf functional traits of Island plants. Guihaia 2020, 40, 433–442. [Google Scholar]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 4, 867–876. [Google Scholar] [CrossRef]
- Jiang, X.L.; Zhang, W.G. Functional diversity and its research methods. Acta Ecol. Sin. 2010, 30, 2766–2773. [Google Scholar]
- Prach, K.; Walker, L.R. Four opportunities for studies of ecological succession. Trends Ecol. Evol. 2011, 26, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Buma, B.; Bisbing, S.; Krapek, J.; Wright, G.A. Foundation of ecology rediscovered: 100 years of succession on the William S. Cooper plots in Glacier Bay, Alaska. Ecology 2017, 6, 1513–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keddy, P.A. Assembly and Response Rules: Two Goals for Predictive Community Ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.Q.; Zhao, Y.J.; Liu, Y.G.; Wang, X.; Gao, X.M. Variation and correlation of plant functional traits in karst area of central Guizhou Province. Chin. J. Plant Ecol. 2011, 35, 1000–1008. [Google Scholar] [CrossRef]
- Liu, H.W.; Liu, W.D.; Wang, W.; Chai, J.; Tao, J.P. Leaf traits and nutrient resorption of major woody species in the karst limestone area of Chongqing. Acta Ecol. Sin. 2015, 35, 4071–4080. [Google Scholar]
- Jiang, Y.; Chen, X.B.; Ma, J.M.; Liang, S.C.; Huang, J. Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broadleaved mixed forests in karst topography, Guilin, Southwest China. Trop. Conserv. Sci. 2016, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Zheng, J.M.; Wang, G.Z.; Zhou, J.X.; Liu, Y.G.; Ha, W.X. A Study of Functional Traits of Natural Secondary Forests and Their Influencing Factors in Different Restoration Stages in Karst Areas: A Case Study of Dahei Mountain, Yunnan Province. Acta Geol. Sin. 2021, 42, 397–406. [Google Scholar]
- Fu, H.; Zhong, J.; Fang, S.; Hu, J.; Guo, C.; Lou, Q.; Yuan, G.; Dai, T.; Li, Z.; Zhang, M.; et al. Scale-dependent changes in the functional diversity of macrophytes in subtropical freshwater lakes in south China. Sci. Rep. 2017, 7, 8294. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattage, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Bu, W.S.; Zang, R.G.; Ding, Y. Functional diversity increases with species diversity along successional gradient in a secondary tropical lowland rainforest. Trop. Ecol. 2014, 55, 393–401. [Google Scholar]
- Yu, L.F.; Zhu, S.Q.; Ye, J.C.; Wei, L.M.; Chen, Z.R. Dynamics of a degraded Karst forest in the process of natural restoration. Sci. Silvae Sin. 2002, 38, 1–7. [Google Scholar]
- Ren, Y.X. Functional Diversity of Typical Forest Communities in Beijing Mountainous Area. Master’s Thesis, Beijing Forestry University, Beijing, China, 2012. [Google Scholar]
- Li, L.F. Quantitative Ecology of Picea wilsonii Forest in Luya Mountain Nature Reserve. Ph.D. Thesis, Beijing Normal University of China, Beijing, China, 2014; pp. 49–52. [Google Scholar]
- Villeger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; Mccauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Koerselman, W.; Meuleman, A. The vegetation N: P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Zeng, Z.X.; Wang, K.L.; Liu, X.L.; Zeng, F.P.; Song, T.Q.; Peng, W.X.; Zhang, H.; Du, H. Stoichiometric characteristics of live fresh and leaf litter from typical communities in a karst region of northwest Guangxi, China. Acta Ecol. Sin. 2015, 36, 1907–1914. [Google Scholar]
- Tian, H.Q.; Chen, G.S.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C: N: P ratios in China’s soil: A synthsis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Gao, S.P.; Li, J.X.; Xu, M.C.; Chen, X.; Dai, J. Leaf N and P stoichiometry of common species in succession stages of the evergreen broad-leaved forest in Tiantong National Forest Park, Zhejiang Province, China. Acta Ecol. Sin. 2007, 27, 947–952. [Google Scholar]
- Chen, L.L.; Deng, Q.; Yuan, Z.Y.; Mu, X.M.; Kallenbach, R.L. Age-related C: N: P stoichiometry in two plantation forests in the Loess Plateau of China. Ecol. Eng. 2018, 120, 857–866. [Google Scholar] [CrossRef]
- Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 1982, 119, 553–572. [Google Scholar] [CrossRef]
- Bowman, W.D. Accumulation and use of nitrogen and phosphorus following fertilization in two alpine tundra communities. Oikos 1994, 70, 261–270. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sample Size (m×m) | Longitude (°) | Latitude (°) | Elevation (m) | Soil Type | Parent Material |
|---|---|---|---|---|---|---|
| HE1 | 30 × 30 | 107.90682 | 25.214433 | 821.58 | Calcareous soil | Dolomite and limestone |
| HE2 | 30 × 30 | 107.90686 | 25.214346 | 824.78 | - | - |
| HE3 | 30 × 30 | 107.90699 | 25.214055 | 845.2 | - | - |
| HS1 | 30 × 30 | 107.93928 | 25.303444 | 697.16 | - | - |
| HS2 | 30 × 30 | 107.93734 | 25.303432 | 702.69 | - | - |
| HS3 | 30 × 30 | 107.93891 | 25.304911 | 693.62 | - | - |
| SH1 | 30 × 30 | 107.90229 | 25.23859 | 865.16 | - | - |
| SH2 | 30 × 30 | 107.90249 | 25.238667 | 857.85 | - | - |
| SH3 | 30 × 30 | 107.90341 | 25.238372 | 863.41 | - | - |
| TS1 | 30 × 30 | 107.94053 | 25.303972 | 693.55 | - | - |
| TS2 | 30 × 30 | 107.93925 | 25.3054 | 732.76 | - | - |
| TS3 | 30 × 30 | 107.9406 | 25.303961 | 777.35 | - | - |
| TR1 | 30 × 30 | 107.94091 | 25.293638 | 710.35 | - | - |
| TR2 | 30 × 30 | 107.93885 | 25.27445 | 716.87 | - | - |
| TR3 | 30 × 30 | 107.94022 | 25.293434 | 962.5 | - | - |
| CL1 | 30 × 30 | 107.93777 | 25.233763 | 668.87 | - | - |
| CL2 | 30 × 30 | 107.93838 | 25.233347 | 693.13 | - | - |
| CL3 | 30 × 30 | 107.93704 | 25.234201 | 729.35 | - | - |
| Restoration Stage | Forest Stand Type | Coverage (%) | Height (m) | Average Diameter at Breast (cm) | Mean Ground Diameter (cm) |
|---|---|---|---|---|---|
| HE | Miscanthus sinensis Anderss, Cyclosorus interruptus Community | 88 | 0.61 | null | 0.67 |
| HS | Nandina domestica Community | 85 | 1.16 | null | 1.25 |
| SH | Loropetalum chinense Community | 78 | 1.31 | null | 1.88 |
| TS | Lindera communis Hemsl. Nandina domestica Community | 87 | 6.85 | 11.76 | null |
| TR | Swida wilsoniana (Wanger.) Sojak Community | 90 | 9.9 | 15.46 | null |
| CL | Swida wilsoniana (Wanger.) Sojak Community | 92 | 11.77 | 19.18 | null |
| Constitute | Succession | CC (g/kg) | CV (cc) | NC (g/kg) | CV (nc) | PC (g/kg) | CV (pc) |
|---|---|---|---|---|---|---|---|
| Soil | HE | 22.44 ± 6.92 a | 30.89% | 0.87 ± 0.32 b | 36.78% | 0.31 ± 0.15 a | 48.39% |
| HS | 49.45 ± 17.38 b | 35.15% | 1.99 ± 0.70 c | 35.18% | 0.61 ± 0.10 b | 16.39% | |
| SH | 24.01 ± 7.63 a | 31.78% | 0.49 ± 0.17 a | 34.69% | 1.10 ± 0.74 c | 67.27% | |
| TS | 82.88 ± 20.46 d | 24.69% | 3.86 ± 0.64 d | 16.16% | 0.99 ± 0.39 c | 39.39% | |
| TR | 60.30 ± 17.14 c | 28.42% | 2.25 ± 0.91 c | 40.44% | 1.51 ± 0.48 d | 31.79% | |
| CL | 76.14 ± 23.92 d | 31.42% | 3.63 ± 0.78 d | 21.49% | 2.00 ± 0.69 d | 34.50% | |
| Leaf | HE | 412.65 ± 27.72 b | 6.72% | 3.23 ± 0.84 a | 26.01% | 0.53 ± 0.27 a | 50.94% |
| HS | 368.76 ± 53.49 a | 14.51% | 7.86 ± 2.14 c | 27.23% | 1.16 ± 0.76 bc | 47.50% | |
| SH | 409.45 ± 45.54 b | 11.12% | 6.66 ± 4.05 bc | 60.81% | 1.30 ± 1.30 bc | 100.00% | |
| TS | 413.59 ± 76.43 b | 18.48% | 4.03 ± 1.80 a | 44.67% | 1.00 ± 0.51 b | 51.00% | |
| TR | 405.91 ± 69.53 ab | 17.13% | 6.30 ± 3.25 b | 51.59% | 1.42 ± 0.48 c | 33.80% | |
| CL | 378.98 ± 62.53 ab | 16.50% | 6.64 ± 2.76 bc | 41.57% | 0.75 ± 0.41 ab | 54.67% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, L.; Chen, J.; Feng, L.; Li, F.; Yu, L. Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China. Forests 2022, 13, 864. https://doi.org/10.3390/f13060864
Wang Y, Zhang L, Chen J, Feng L, Li F, Yu L. Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China. Forests. 2022; 13(6):864. https://doi.org/10.3390/f13060864
Chicago/Turabian StyleWang, Yang, Limin Zhang, Jin Chen, Ling Feng, Fangbing Li, and Lifei Yu. 2022. "Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China" Forests 13, no. 6: 864. https://doi.org/10.3390/f13060864
APA StyleWang, Y., Zhang, L., Chen, J., Feng, L., Li, F., & Yu, L. (2022). Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China. Forests, 13(6), 864. https://doi.org/10.3390/f13060864