
Leaf and Branch Hydraulic Plasticity of Two Light-Demanding Broadleaved Tree Species Differing in Water-Use Strategy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Trees
2.2. Measuring Hydraulic Conductance
2.3. Plasticity Index
2.4. Data Analysis
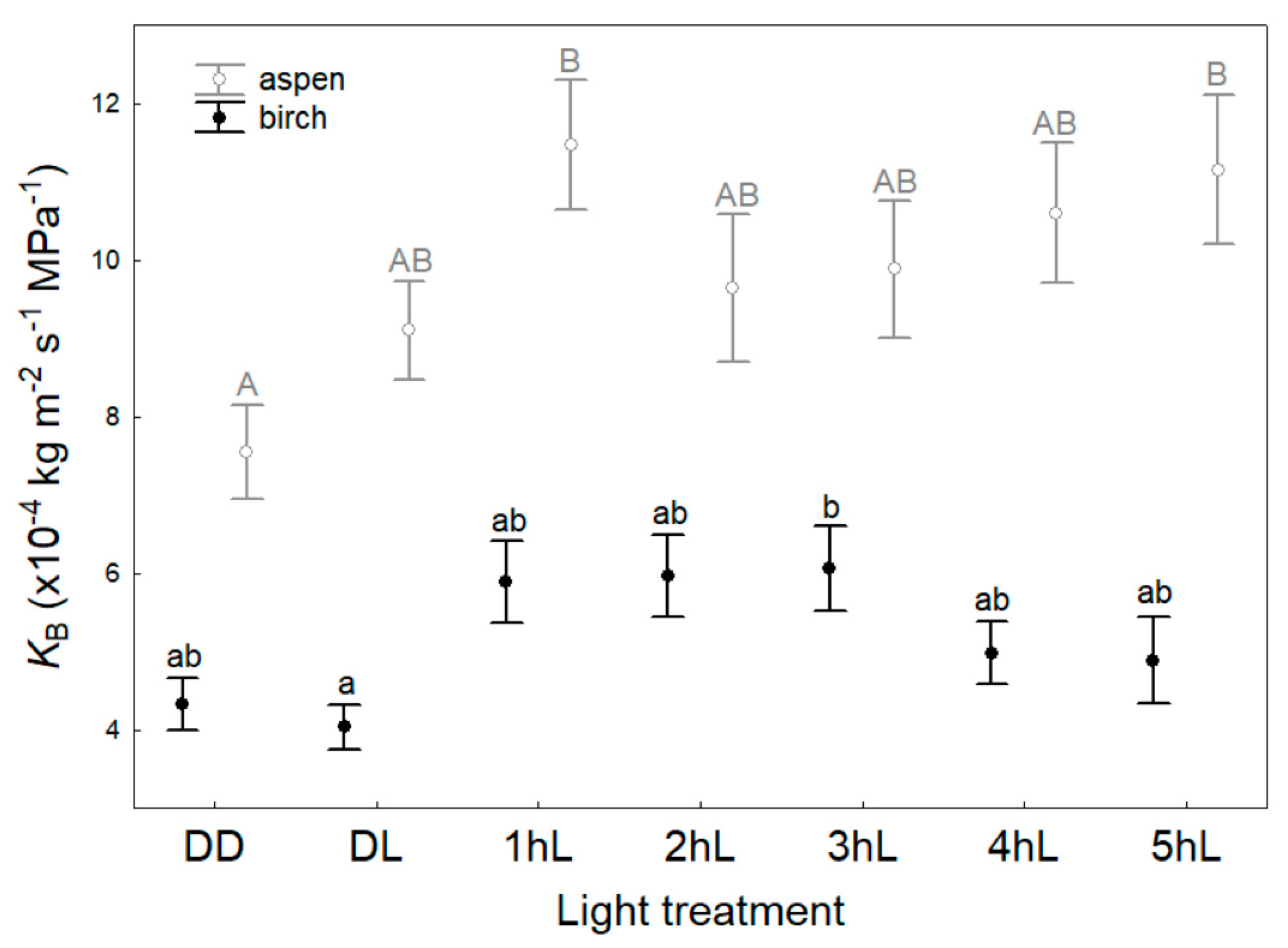
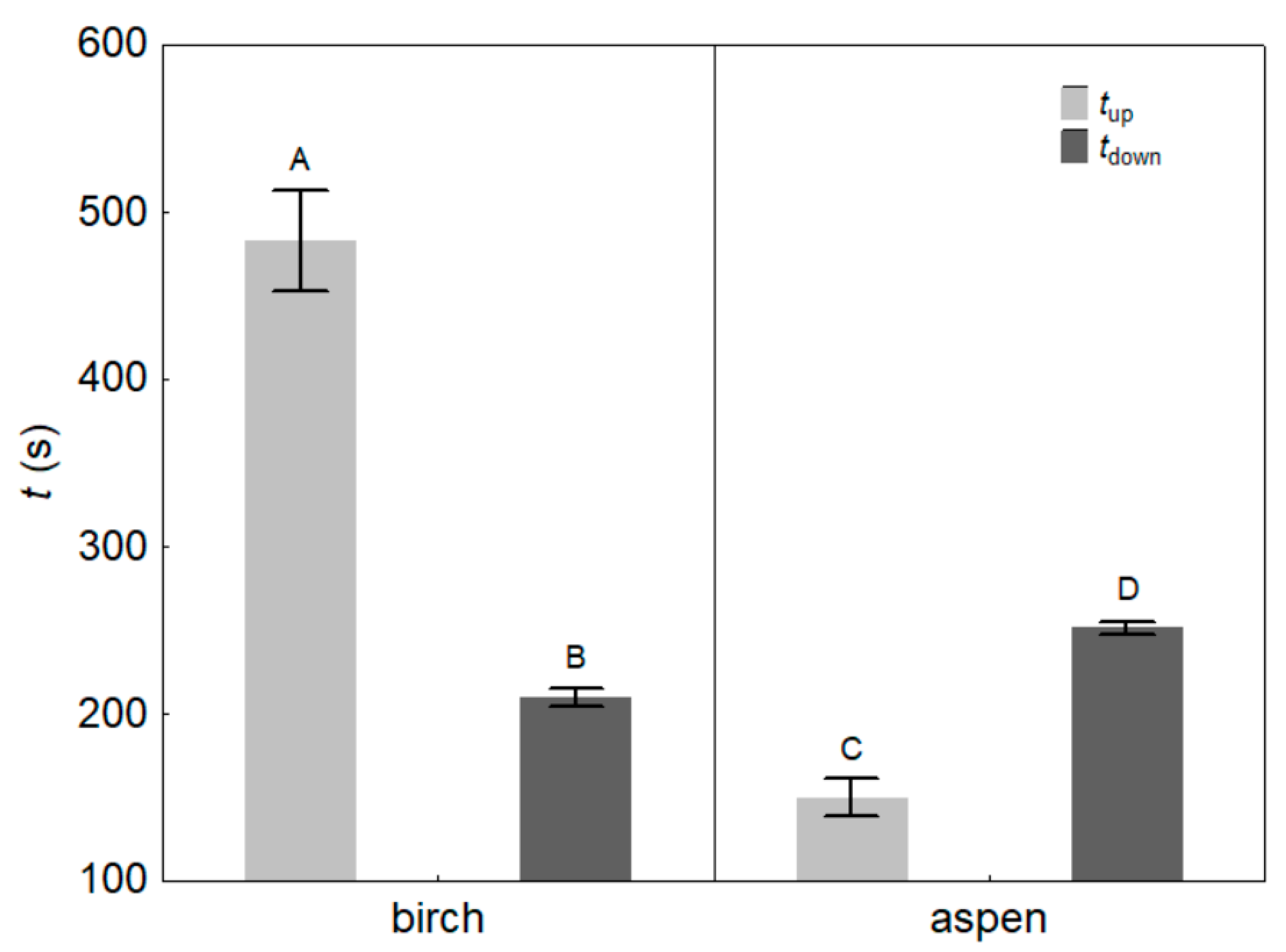
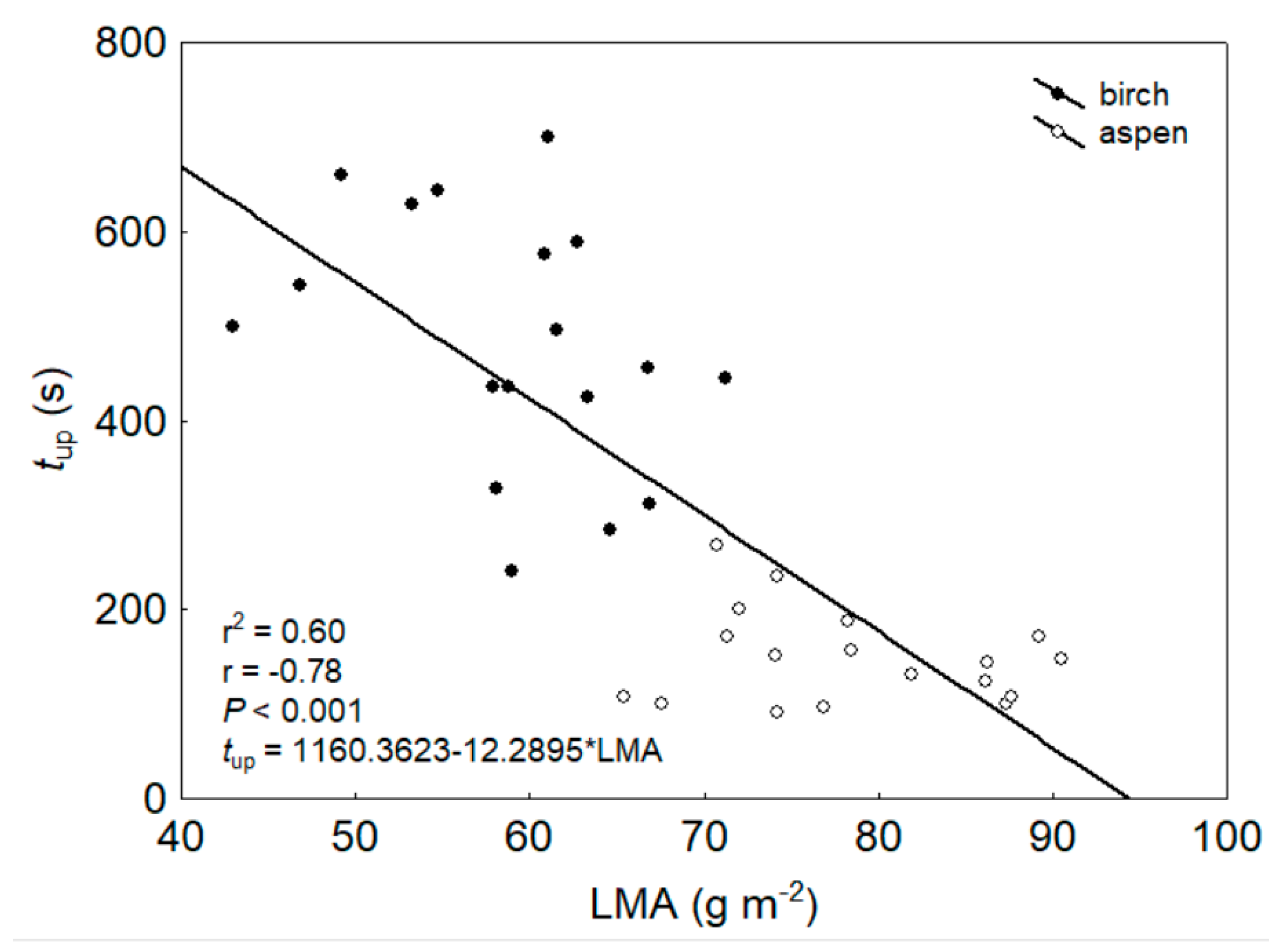
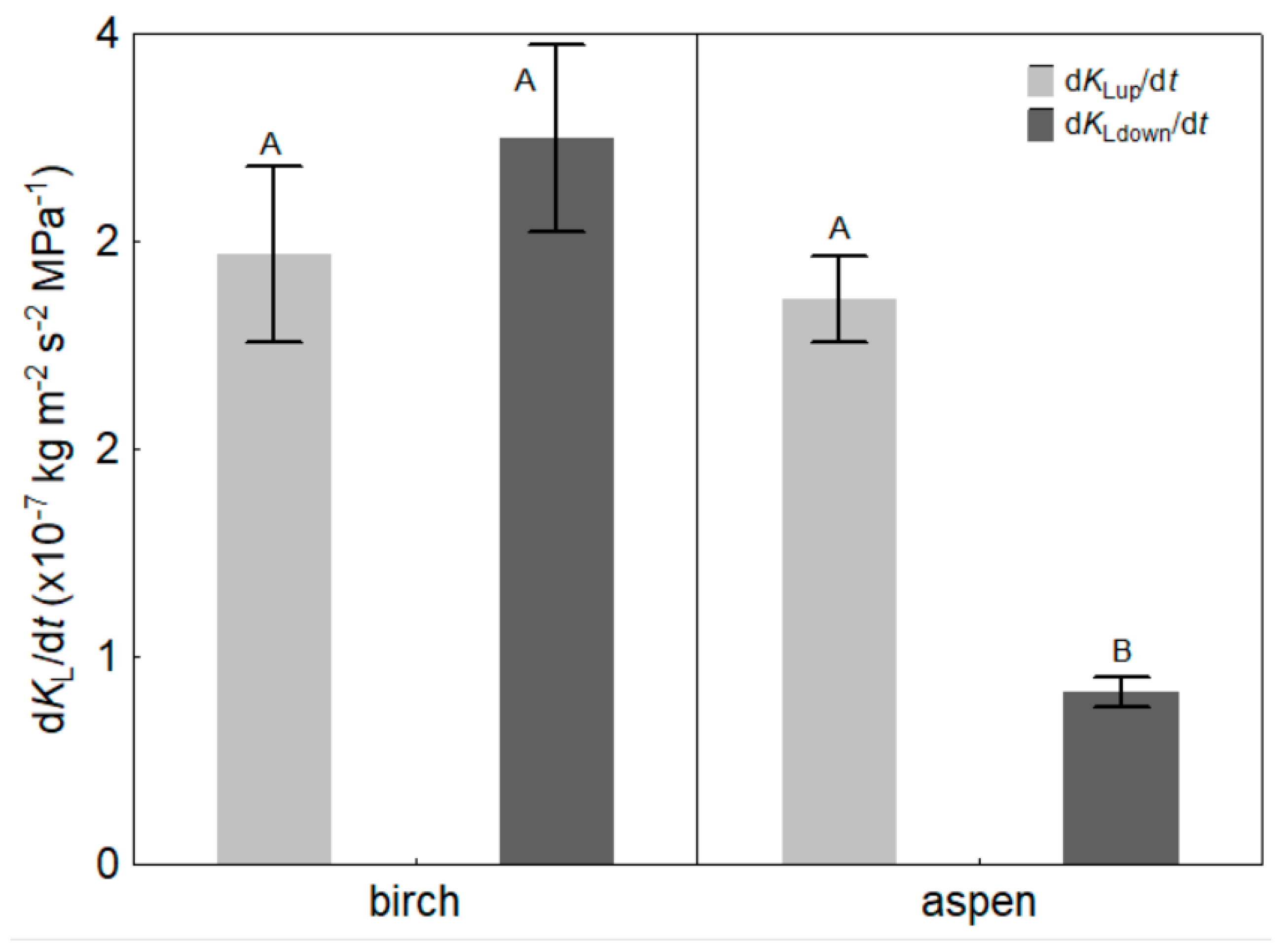
3. Results
4. Discussion
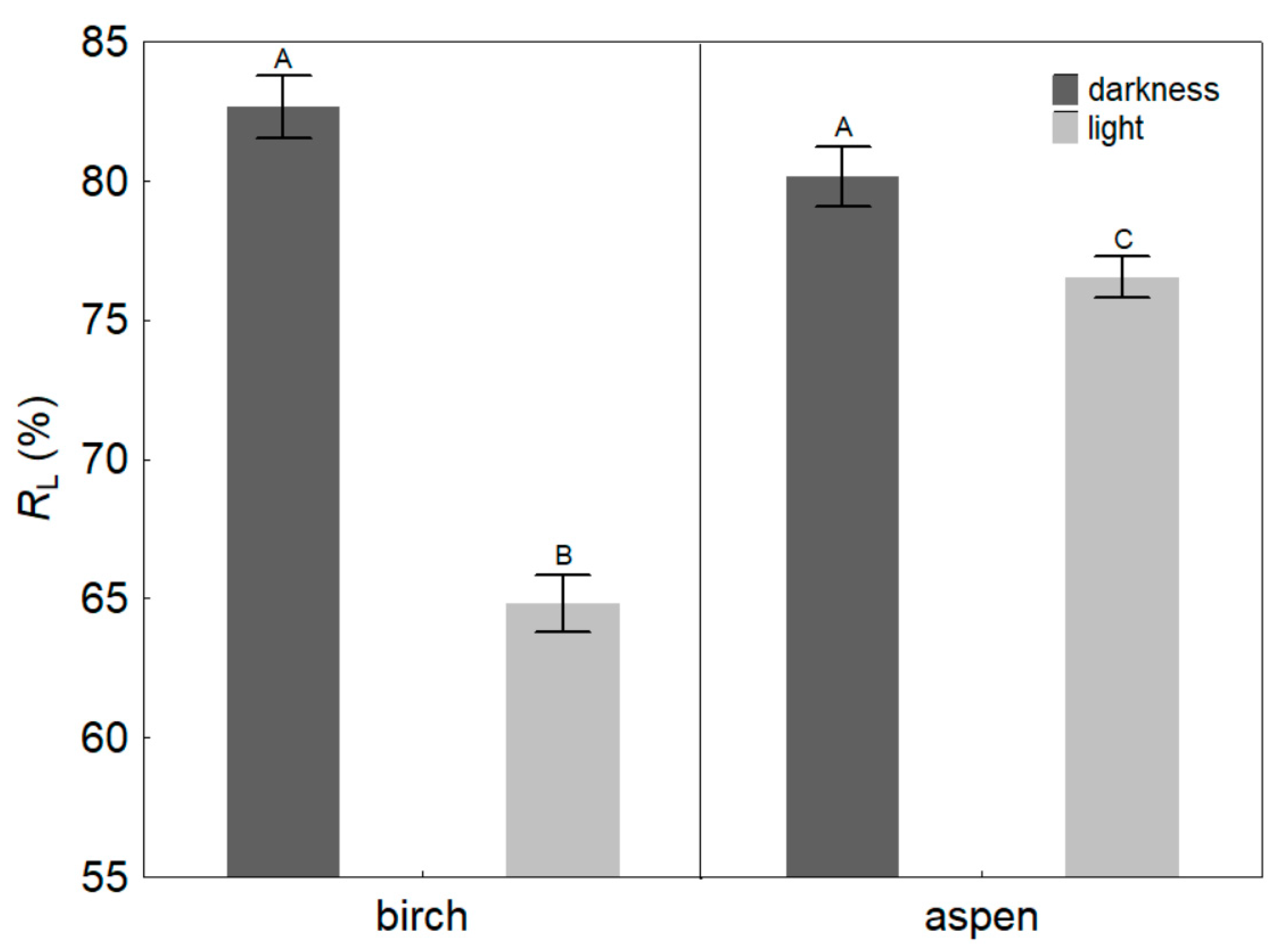
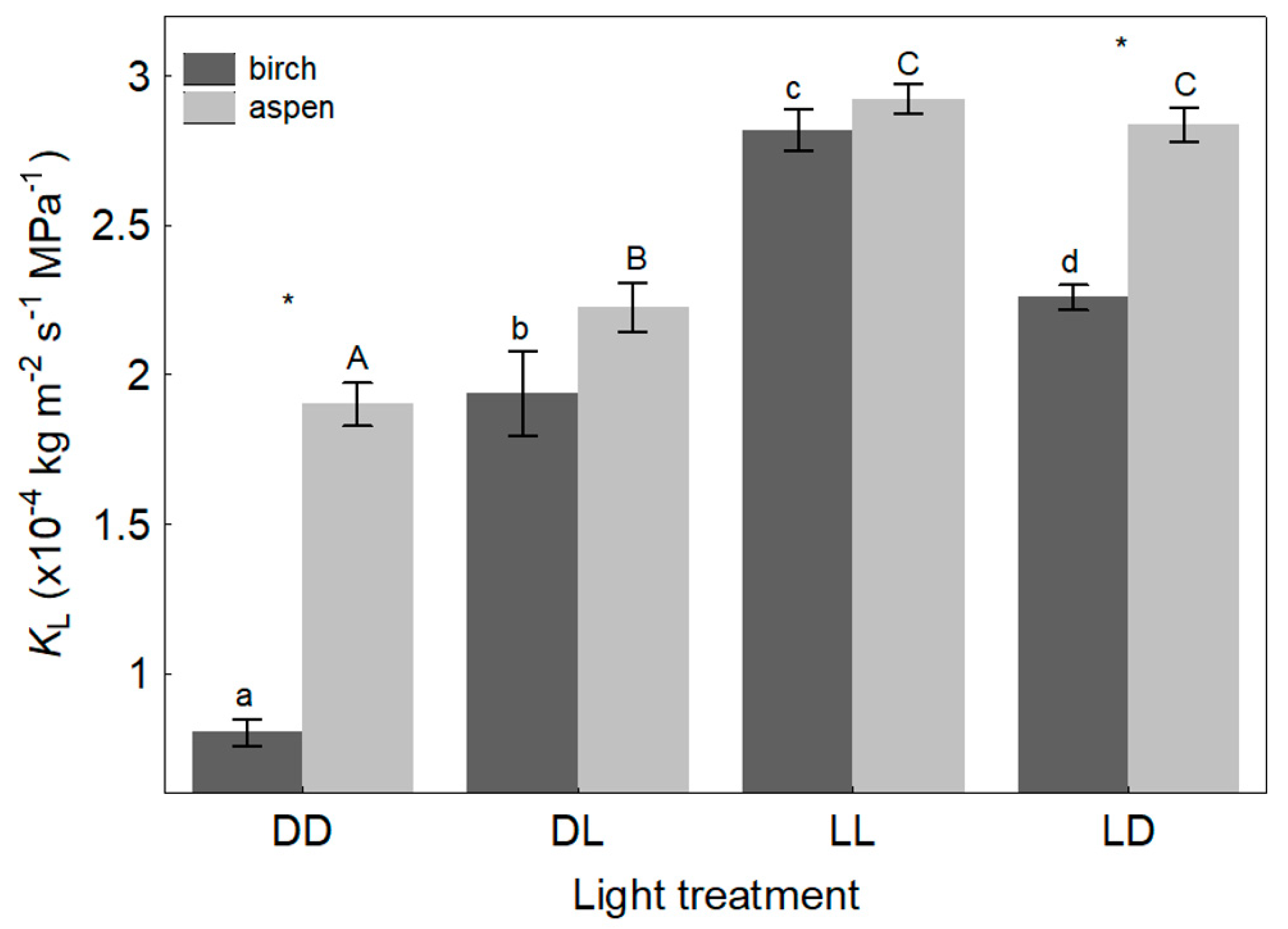
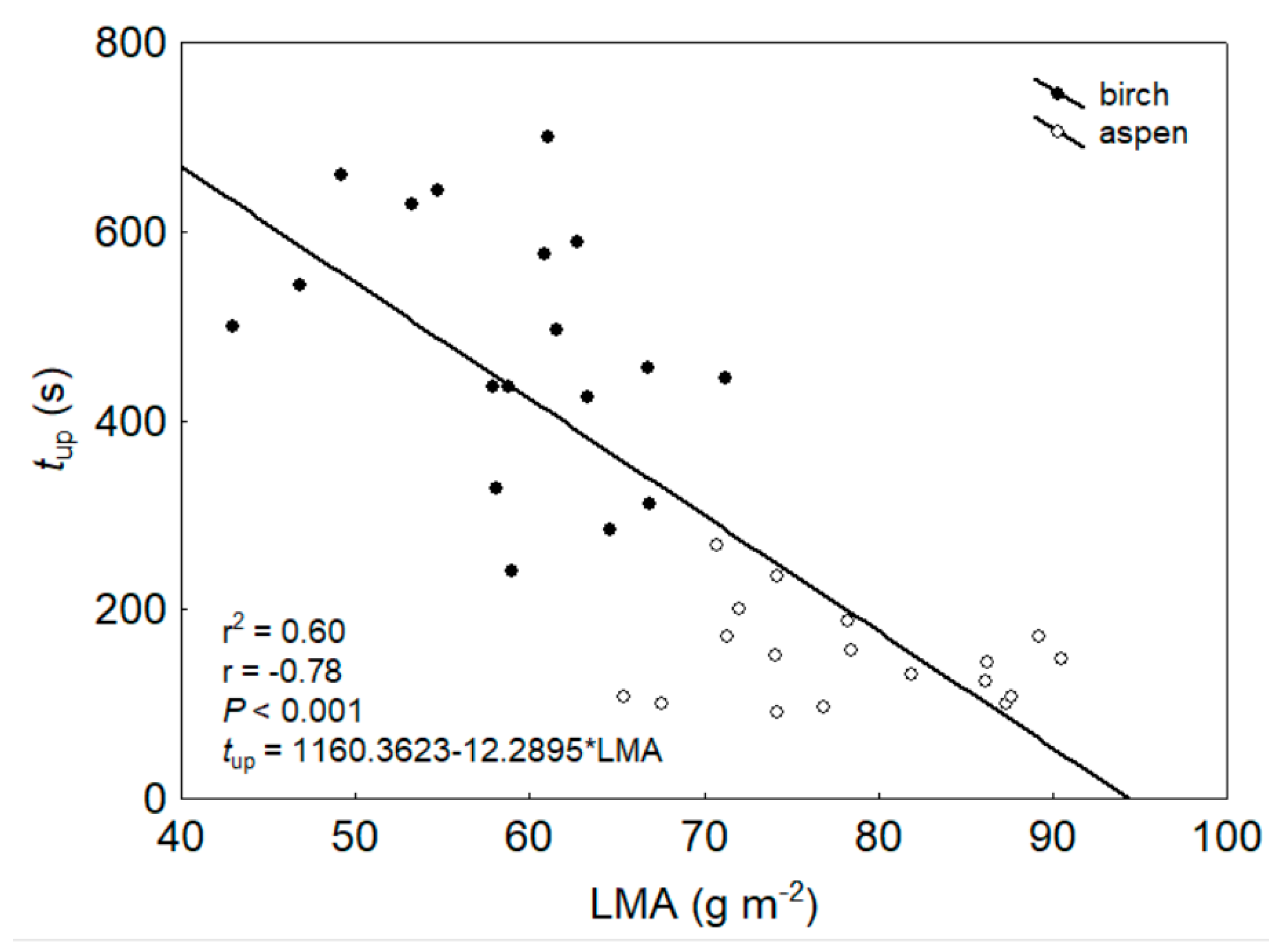
4.1. Light Effects on Leaf Hydraulic Properties
4.2. Light Effects on Branch Hydraulic Properties
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sack, L.; Ball, M.C.; Brodersen, C.; Davis, S.D.; Des Marais, D.L.; Donovan, L.A.; Givnish, T.J.; Hacke, U.G.; Huxman, T.; Jansen, S.; et al. Plant hydraulics as a central hub integrating plant and ecosystem function: Meeting report for ‘Emerging Frontiers in Plant Hydraulics’ (Washington, DC, May 2015). Plant Cell Environ. 2016, 39, 2085–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.L.; Kumar, M.; Katul, G.G.; Feng, X.; Konings, A.G. Plant hydraulics accentuates the effect of atmospheric moisture stress on transpiration. Nat. Clim. Chang. 2020, 10, 691–695. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Brodribb, T.J. Xylem hydraulic physiology: The functional backbone of terrestrial plant productivity. Plant Sci. 2009, 177, 245–251. [Google Scholar] [CrossRef]
- Scoffoni, C.; Chatelet, D.S.; Pasquet-kok, J.; Rawls, M.; Donoghue, M.J.; Edwards, E.J.; Sack, L. Hydraulic basis for the evolution of photosynthetic productivity. Nat. Plants 2016, 2, 16072. [Google Scholar] [CrossRef]
- Xiong, D.; Nadal, M. Linking water relations and hydraulics with photosynthesis. Plant J. 2020, 101, 800–815. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Annu. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Cowan, P.D.; Jaikumar, N.; Holbrook, N.M. The ‘hydrology’ of leaves: Co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef] [Green Version]
- Õunapuu, E.; Sellin, A. Daily dynamics of leaf and soil-to-branch hydraulic conductance in silver birch (Betula pendula) measured in situ. Plant Physiol. Biochem. 2013, 68, 104–110. [Google Scholar] [CrossRef]
- Newman, J.A.; Anand, M.; Henry, H.A.L.; Hunt, S.; Gedalof, Z. Climate Change Biology; CABI Publishing: Wallingford, UK, 2011. [Google Scholar]
- Gratani, L. Plant Phenotypic Plasticity in Response to Environmental Factors. Adv. Bot. 2014, 208747. [Google Scholar] [CrossRef] [Green Version]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Henn, J.J.; Buzzard, V.; Enquist, B.J.; Halbritter, A.H.; Klanderuds, K.; Maitner, B.S.; Michaletz, S.T.; Pötsch, C.; Seltzer, L.; Telford, R.J.; et al. Intraspecific trait variation and phenotypic plasticity mediate alpine plant species response to climate change. Front. Plant Sci. 2018, 9, 1548. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, S.; Martin, I.; Babiano, J.; Escudero, A. Foliar plasticity related to gradients of heat and drought stress across crown orientations in three Mediterranean Quercus species. PLoS ONE 2019, 14, e0224462. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Gianoli, E.; Gomez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Kont, A.; Jaagus, J.; Aunap, R. Climate change scenarios and the effect of sea-level rise for Estonia. Glob. Planet. Chang. 2003, 36, 1–15. [Google Scholar] [CrossRef]
- Betts, A.K.; Desjardins, R.; Worth, D.; Beckage, B. Climate coupling between temperature, humidity, precipitation, and cloud cover over the Canadian Prairies. J. Geophys. Res. -Atmos. 2014, 119, 13305–13326. [Google Scholar] [CrossRef]
- Westra, S.; Fowler, H.J.; Evans, J.P.; Alexander, L.V.; Berg, P.; Johnson, F.; Kendon, E.J.; Lenderink, G.; Roberts, N.M. Future changes to the intensity and frequency of short-duration extreme rainfall. Rev. Geophys. 2014, 52, 522–555. [Google Scholar] [CrossRef]
- Busuioc, A.; Birsan, M.V.; Carbunaru, D.; Baciu, M.; Orzan, A. Changes in the large-scale thermodynamic instability and connection with rain shower frequency over Romania: Verification of the Clausius-Clapeyron scaling. Int. J. Climatol. 2016, 36, 2015–2034. [Google Scholar] [CrossRef]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Körner, O.; Nejad, A.R.; Delis, C.; Bouranis, D.; Koubouris, G.; Kambourakis, E.; et al. Stomatal behavior following mid or long-term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef]
- Niu, S.L.; Luo, Y.Q.; Li, D.J.; Cao, S.H.; Xia, J.Y.; Li, J.W.; Smith, M.D. Plant growth and mortality under climatic extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, E.; Lihavainen, J.; Keinänen, M.; Keski-Saari, S.; Kontunen-Soppela, S.; Sellin, A.; Sõber, A. Northern forest trees under increasing atmospheric humidity. In Progress in Botany; Canovas, F.M., Lüttge, U., Matyssek, R., Pretzsch, H., Eds.; Springer: Cham, Switzerland, 2018; Volume 80. [Google Scholar]
- Sellin, A.; Tullus, A.; Niglas, A.; Õunapuu, E.; Karusion, A.; Lõhmus, K. Humidity-driven changes in growth rate, photosynthetic capacity, hydraulic properties and other functional traits in silver birch (Betula pendula). Ecol. Res. 2013, 28, 523–535. [Google Scholar] [CrossRef]
- Sellin, A.; Taneda, H.; Alber, M. Leaf structural and hydraulic adjustment with respect to air humidity and canopy position in silver birch (Betula pendula). J. Plant Res. 2019, 132, 369–381. [Google Scholar] [CrossRef]
- Sellin, A.; Niglas, A.; Õunapuu-Pikas, E.; Kupper, P. Rapid and long-term effects of water deficit on gas exchange and hydraulic conductance of silver birch trees grown under varying atmospheric humidity. BMC Plant Biol. 2014, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Pritzkow, C.; Williamson, V.; Szota, C.; Trouve, R.; Arndt, S.K. Phenotypic plasticity and genetic adaptation of functional traits influences intra-specific variation in hydraulic efficiency and safety. Tree Physiol. 2020, 40, 215–229. [Google Scholar] [CrossRef]
- Tyree, M.T.; Nardini, A.; Salleo, S.; Sack, L.; El Omari, B. The dependence of leaf hydraulic conductance on irradiance during HPFM measurements: Any role for stomatal response? J. Exp. Bot. 2005, 56, 737–744. [Google Scholar] [CrossRef]
- Sellin, A.; Õunapuu, E.; Kupper, P. Effects of light intensity and duration on leaf hydraulic conductance and distribution of resistance in shoots of silver birch (Betula pendula). Physiol. Plant. 2008, 134, 412–420. [Google Scholar] [CrossRef]
- Voicu, M.C.; Zwiazek, J.J.; Tyree, M.T. Light response of hydraulic conductance in bur oak (Quercus macrocarpa) leaves. Tree Physiol. 2008, 28, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Guyot, G.; Scoffoni, C.; Sack, L. Combined impacts of irradiance and dehydration on leaf hydraulic conductance: Insights into vulnerability and stomatal control. Plant Cell Environ. 2012, 35, 857–871. [Google Scholar] [CrossRef]
- Ohtsuka, A.; Sack, L.; Taneda, H. Bundle sheath lignification mediates the linkage of leaf hydraulics and venation. Plant Cell Environ. 2018, 41, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.L.; Douthe, C.; Flexas, J. Differential coordination of stomatal conductance, mesophyll conductance, and leaf hydraulic conductance in response to changing light across species. Plant Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Kunkle, J.; Pasquet-Kok, J.; Vuong, C.; Patel, A.J.; Montgomery, R.A.; Givnish, T.J.; Sack, L. Light-inducec plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads. New Phytol. 2015, 207, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Õunapuu-Pikas, E.; Sellin, A. Plasticity and light sensitivity of leaf hydraulic conductance to fast changes in irradiance in common hazel (Corylus avellana L.). Plant Sci. 2020, 290, 110299. [Google Scholar] [CrossRef] [PubMed]
- Possen, B.J.H.M.; Oksanen, E.; Rousi, M.; Ruhanen, H.; Ahonen, V.; Tervahauta, A.; Heinonen, J.; Heiskanen, J.; Kärenlampi, S.; Vapaavuori, E. Adaptability of birch (Betula pendula Roth) and aspen (Populus tremula L.) genotypes to different soil moisture conditions. For. Ecol. Manag. 2011, 262, 1387–1399. [Google Scholar] [CrossRef]
- Hynynen, J.; Niemisto, P.; Vihera-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Sellin, A.; Õunapuu, E.; Karusion, A. Experimental evidence supporting the concept of light-mediated modulation of stem hydraulic conductance. Tree Physiol. 2010, 30, 1528–1535. [Google Scholar] [CrossRef] [Green Version]
- Zapater, M.; Breda, N.; Bonal, D.; Pardonnet, S.; Granier, A. Differential response to soil drought among co-occurring broad-leaved tree species growing in a 15-to 25-year-old mixed stand. Ann. For. Sci. 2013, 70, 31–39. [Google Scholar] [CrossRef]
- Kupper, P.; Ivanova, H.; Sõber, A.; Rohula-Okunev, G.; Sellin, A. Night and daytime water relations in five fast-growing tree species: Effects of environmental and endogenous variables. Ecohydrology 2018, 11, e1927. [Google Scholar] [CrossRef]
- Aasamaa, K.; Sõber, A.; Hartung, W.; Niinemets, Ü. Drought acclimation of two deciduous tree species of different layers in a temperate forest canopy. Trees-Struct. Funct. 2004, 18, 93–101. [Google Scholar]
- Aasamaa, K.; Kõivik, K.; Kupper, P.; Sõber, A. Growth environment determines light sensitivity of shoot hydraulic conductance. Ecol. Res. 2014, 29, 143–151. [Google Scholar] [CrossRef]
- Russak, V. Päikesekiirgus. Tartu Kliima Ja Selle Muutumine Viimastel Kümnenditel; Ross, J., Ed.; Eesti Teaduste Akadeemia: Tartu, Estonia, 1990. [Google Scholar]
- Jaagus, J.; Mändla, K. Climate change scenarios for Estonia based on climate models from the IPCC Fourth Assessment Report. Est. J. Earth Sci. 2014, 63, 166–180. [Google Scholar] [CrossRef]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Kupper, P.; Sõber, J.; Sellin, A.; Lõhmus, K.; Tullus, A.; Räim, O.; Lubenets, K.; Tulva, I.; Uri, V.; Zobel, M.; et al. An experimental facility for free air humidity manipulation (FAHM) can alter water flux through deciduous tree canopy. Environ. Exp. Bot. 2011, 72, 432–438. [Google Scholar] [CrossRef]
- Tullus, A.; Kupper, P.; Sellin, A.; Parts, L.; Sõber, J.; Tullus, T.; Tullus, H. Climate change at Northern latitudes: Rising atmospheric humidity decreases transpiration, N-uptake and growth rate of hybrid aspen. PLoS ONE 2012, 7, e42648. [Google Scholar] [CrossRef] [PubMed]
- Sellin, A.; Alber, M.; Keinanen, M.; Kupper, P.; Lihavainen, J.; Lõhmus, K.; Tullus, A. Growth of northern deciduous trees under increasing atmospheric humidity: Possible mechanisms behind the growth retardation. Reg. Environ. Chang. 2017, 17, 2135–2148. [Google Scholar] [CrossRef]
- Heimonen, K.; Valtonen, A.; Kontunen-Soppela, S.; Keski-Saari, S.; Rousi, M.; Oksanen, E.; Roininen, H. Insect herbivore damage on latitudinally translocated silver birch (Betula pendula)—Predicting the effects of climate change. Clim. Chang. 2015, 131, 245–257. [Google Scholar] [CrossRef]
- Valladares, F.; Martinez-Ferri, E.; Balaguer, L.; Perez-Corona, E.; Manrique, E. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource-use strategy? New Phytol. 2000, 148, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Melcher, P.J.; Zwieniecki, M.A.; Holbrook, N.M. The hydraulic conductance of the angiosperm leaf lamina: A comparison of three measurement methods. J. Exp. Bot. 2002, 53, 2177–2184. [Google Scholar] [CrossRef] [Green Version]
- Nardini, A.; Salleo, S.; Andri, S. Circadian regulation of leaf hydraulic conductance in sunflower (Helianthus annuus L. cv Margot). Plant Cell Environ. 2005, 28, 750–759. [Google Scholar] [CrossRef]
- Scoffoni, C.; Pou, A.; Aasamaa, K.; Sack, L. The rapid light response of leaf hydraulic conductance: New evidence from two experimental methods. Plant Cell Environ. 2008, 31, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Venisse, J.-S.; Õunapuu-Pikas, E.; Dupont, M.; Gousset-Dupont, A.; Saadaoui, M.; Faize, M.; Chen, S.; Chen, S.; Petel, G.; Fumanal, B.; et al. Genome-wide identification, structure characterization, and expression pattern profiling of the aquaporin gene family in Betula pendula. Int. J. Mol. Sci. 2021, 22, 7269. [Google Scholar] [CrossRef] [PubMed]
- Cochard, H.; Venisse, J.-S.; Barigah, T.S.; Brunel, N.; Herbette, S.; Guilliot, A.; Tyree, M.T.; Sakr, S. Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol. 2007, 143, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Ben Baaziz, K.; Lopez, D.; Bouzid, S.; Cochard, H.; Venisse, J.S.; Sakr, S. Early gene expression in the walnut tree occurring during stimulation of leaf hydraulic conductance by irradiance. Biol. Plant. 2012, 56, 657–666. [Google Scholar] [CrossRef]
- Ben Baaziz, K.; Lopez, D.; Rabot, A.; Combes, D.; Gousset, A.; Bouzid, S.; Venisse, J.S. Light-mediated K-leaf induction and contribution of both the PIP1s and PIP2s aquaporins in five tree species: Walnut (Juglans regia) case study. Tree Physiol. 2012, 32, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Lopez, D.; Venisse, J.S.; Fumanal, B.; Chaumont, F.; Guillot, E.; Daniels, M.J.; Gousset-Dupont, A. Aquaporins and leaf hydraulics: Poplar sheds new light. Plant Cell Physiol. 2013, 54, 1963–1975. [Google Scholar] [CrossRef] [Green Version]
- Miniussi, M.; Del Terra, L.; Savi, T.; Pallavicini, A.; Nardini, A. Aquaporins in Coffea arabica L.: Identification, expression, and impacts on plant water relations and hydraulics. Plant Physiol. Biochem. 2015, 95, 92–102. [Google Scholar] [CrossRef]
- Prado, K.; Boursiac, Y.; Tournaire-Roux, C.; Monneuse, J.M.; Postaire, O.; Da Ines, O.; Maurel, C. Regulation of Arabidopsis leaf hydraulics involves light-dependent phosphorylation of aquaporins in veins. Plant Cell 2013, 25, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Prado, K.; Cotelle, V.; Li, G.W.; Bellati, J.; Tang, N.; Tournaire-Roux, C.; Martinière, A.; Santoni, V.; Maurel, C. Oscillating aquaporin phosphorylation and 14-3-3 proteins mediate the circadian regulation of leaf hydraulics. Plant Cell 2019, 31, 417–429. [Google Scholar] [CrossRef]
- Groszmann, M.; Osborn, H.L.; Evans, J.R. Carbon dioxide and water transport through plant aquaporins. Plant Cell Environ. 2017, 40, 938–961. [Google Scholar] [CrossRef] [PubMed]
- Scoffoni, C.; Albuquerque, C.; Cochard, H.; Buckley, T.N.; Fletcher, L.R.; Caringella, M.A.; Sack, L. The causes of leaf hydraulic vulnerability and its influence on gas exchange in Arabidopsis thaliana. Plant Physiol. 2018, 178, 1584–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifilo, P.; Petruzzellis, F.; Abate, E.; Nardini, A. The extra-vascular water pathway regulates dynamic leaf hydraulic decline and recovery in Populus nigra. Physiol. Plant. 2021, 172, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Sellin, A.; Alber, M.; Jasińska, A.K.; Rosenvald, K. Adjustment of leaf anatomical and hydraulic traits across vertical canopy profiles of young broadleaved forest stands. Trees-Struct. Funct. 2022, 36, 67–80. [Google Scholar] [CrossRef]
- Sellin, A.; Kupper, P. Variation in leaf conductance of silver birch: Effects of irradiance, vapour pressure deficit, leaf water status and position within a crown. For. Ecol. Manag. 2005, 206, 153–166. [Google Scholar] [CrossRef]
- Sellin, A.; Eensalu, E.; Niglas, A. Is distribution of hydraulic constraints within tree crowns reflected in photosynthetic water-use efficiency? An example of Betula pendula. Ecol. Res. 2010, 25, 173–183. [Google Scholar] [CrossRef]
- Kangur, O.; Kupper, P.; Sellin, A. Predawn disequilibrium between soil and plant water potentials in light of climate trends predicted for northern Europe. Reg. Environ. Chang. 2017, 17, 2159–2168. [Google Scholar] [CrossRef]
- Sellin, A.; Alber, M.; Kupper, P. Increasing air humidity influences hydraulic efficiency but not functional vulnerability of xylem in hybrid aspen. J. Plant Physiol. 2017, 219, 28–36. [Google Scholar] [CrossRef]
- Azad, A.K.; Sawa, Y.; Ishikawa, T.; Shibata, H. Phosphorylation of plasma membrane aquaporin regulates temperature-dependent opening of tulip petals. Plant Cell Physiol. 2004, 45, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Hedfalk, K.; Tornroth-Horsefield, S.; Nyblom, M.; Johanson, U.; Kjellbom, P.; Neutze, R. Aquaporin gating. Curr. Opin. Struct. Biol. 2006, 16, 447–456. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Zwieniecki, M.A.; Melcher, P.J.; Holbrook, N.M. Hydrogel control of xylem hydraulic resistance in plants. Science 2001, 291, 1059–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardini, A.; Salleo, S.; Jansen, S. More than just a vulnerable pipeline: Xylem physiology in the light of ion-mediated regulation of plant water transport. J. Exp. Bot. 2011, 62, 4701–4718. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Secchi, F. Getting variable xylem hydraulic resistance under control: Interplay of structure and function. Tree Physiol. 2012, 32, 1431–1433. [Google Scholar] [CrossRef] [Green Version]
- Nardini, A.; Grego, F.; Trifilo, P.; Salleo, S. Changes of xylem sap ionic content and stem hydraulics in response to irradiance in Laurus nobilis. Tree Physiol. 2010, 30, 628–635. [Google Scholar] [CrossRef]
- Lee, J.; Holbrook, N.M.; Zwieniecki, M.A. Ion induced changes in the structure of bordered pit membranes. Front. Plant Sci. 2012, 3, 55. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | B. pendula | P. × wettsteinii | Statistical Significance |
|---|---|---|---|
| LS (cm) | 27.9 ± 0.34 | 20.5 ± 0.38 | p < 0.05 |
| AL (cm2) | 167 ± 3.4 | 338 ± 9.1 | p < 0.05 |
| LMA (g m−2) | 60.8 ± 0.64 | 77.8 ± 0.74 | p < 0.05 |
| HV (×10−4; m2 m−2) | 1.24 ± 0.023 | 1.34 ± 0.035 | p < 0.05 |
| Trait | Factor | Statistical Significance |
|---|---|---|
| Leaf hydraulic conductance, KL (kg m−2 s−1 MPa−1) | Species Light treatment Species × Light tr. | p < 0.001 p < 0.001 p < 0.001 |
| Branch hydraulic conductance, KB (kg m−2 s−1 MPa−1) | Species Light treatment Species×Light tr. | p < 0.001 p = 0.001 p = 0.174 |
| Up-regulation time of KL, tup (s) | Species | p < 0.001 |
| Down-regulation time of KL, tdown (s) | Species | p < 0.001 |
| Up-regulation rate of KL, (dKLup/dt; kg m−2 s−2 MPa−1) | Species | p = 0.667 |
| Down-regulation rate of KL, (dKLdown/dt; kg m−2 s−2 MPa−1) | Species | p < 0.001 |
| Initial up-regulation rate of KL, (bup; kg m−2 s−2 MPa−1) | Species | p = 0.193 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Õunapuu-Pikas, E.; Venisse, J.-S.; Label, P.; Sellin, A. Leaf and Branch Hydraulic Plasticity of Two Light-Demanding Broadleaved Tree Species Differing in Water-Use Strategy. Forests 2022, 13, 594. https://doi.org/10.3390/f13040594
Õunapuu-Pikas E, Venisse J-S, Label P, Sellin A. Leaf and Branch Hydraulic Plasticity of Two Light-Demanding Broadleaved Tree Species Differing in Water-Use Strategy. Forests. 2022; 13(4):594. https://doi.org/10.3390/f13040594
Chicago/Turabian StyleÕunapuu-Pikas, Eele, Jean-Stéphane Venisse, Philippe Label, and Arne Sellin. 2022. "Leaf and Branch Hydraulic Plasticity of Two Light-Demanding Broadleaved Tree Species Differing in Water-Use Strategy" Forests 13, no. 4: 594. https://doi.org/10.3390/f13040594
APA StyleÕunapuu-Pikas, E., Venisse, J.-S., Label, P., & Sellin, A. (2022). Leaf and Branch Hydraulic Plasticity of Two Light-Demanding Broadleaved Tree Species Differing in Water-Use Strategy. Forests, 13(4), 594. https://doi.org/10.3390/f13040594