
Proof-of-Principle That Cellular Automata Can Be Used to Predict Infestation Risk by Reticulitermes grassei (Blattodea: Isoptera)

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
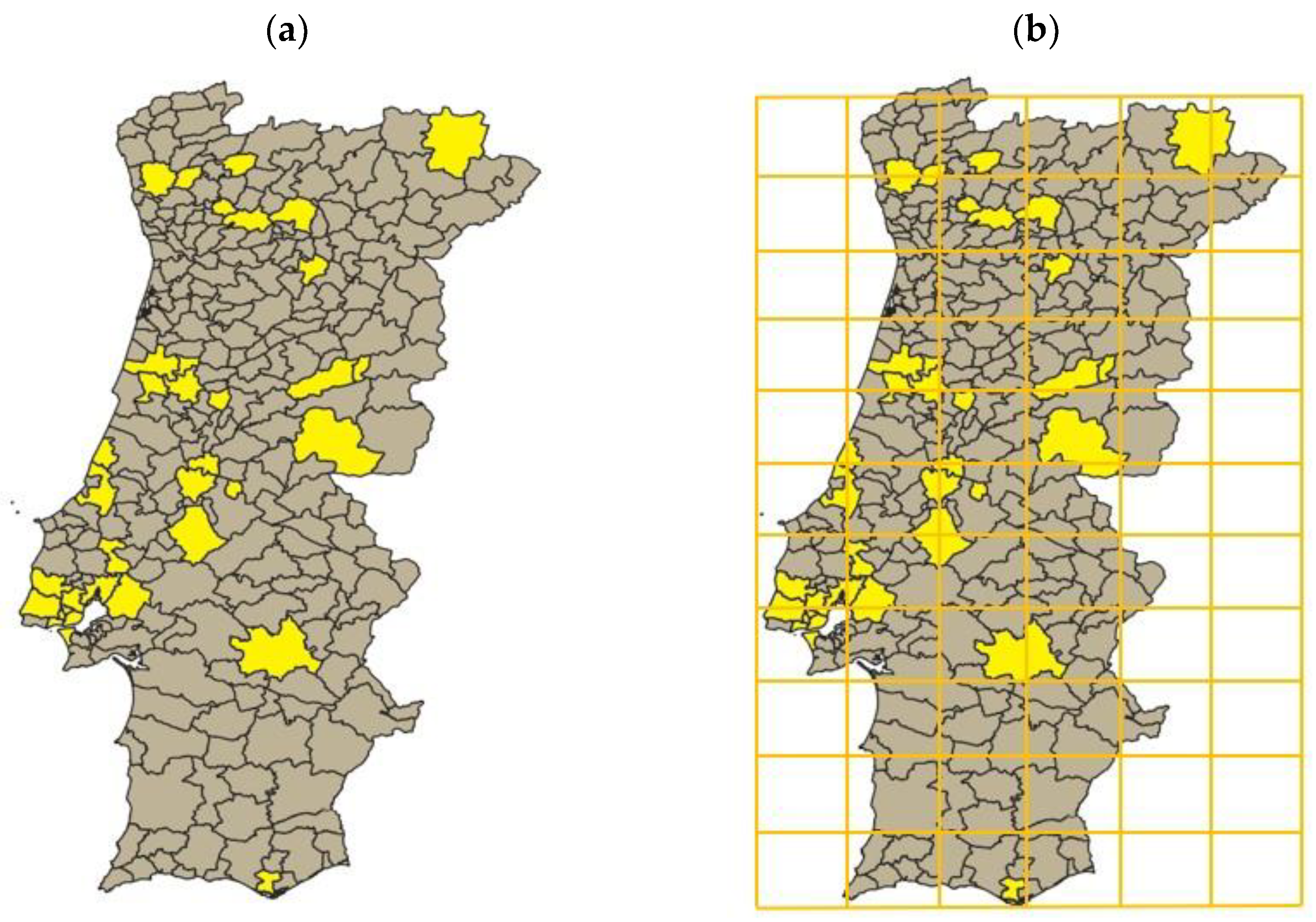
- (a)
- The classification of each cell for LS, IS, and DR was undertaken according to the most frequent class present in the totality of the region. However, if smaller cells had been considered, the classes of each zone might have been different, which would have led to larger, or smaller, probabilities for termite infestation;
- (b)
- Relatively large cells were considered; thus, the HP class of each cell was invariably high (larger cells comprehend more counties and, hence, more people), meaning that the class given for a specific cell was, most of the times, larger than 8. This had a very significant impact as, according to Nobre and Nunes [25], HP weight in the probability for R. grassei existence increases exponentially with class increase;
- (c)
- Termite presence in the neighboring cells was postulated to double the probability of termite establishment in each cell. However, this value was not mathematically estimated, meaning that the real predicted influence on the R. grassei probability could possibly be different (and vary with the number of surrounding cells with termite pest infestation, something that was also not considered in the developed simulation tool).
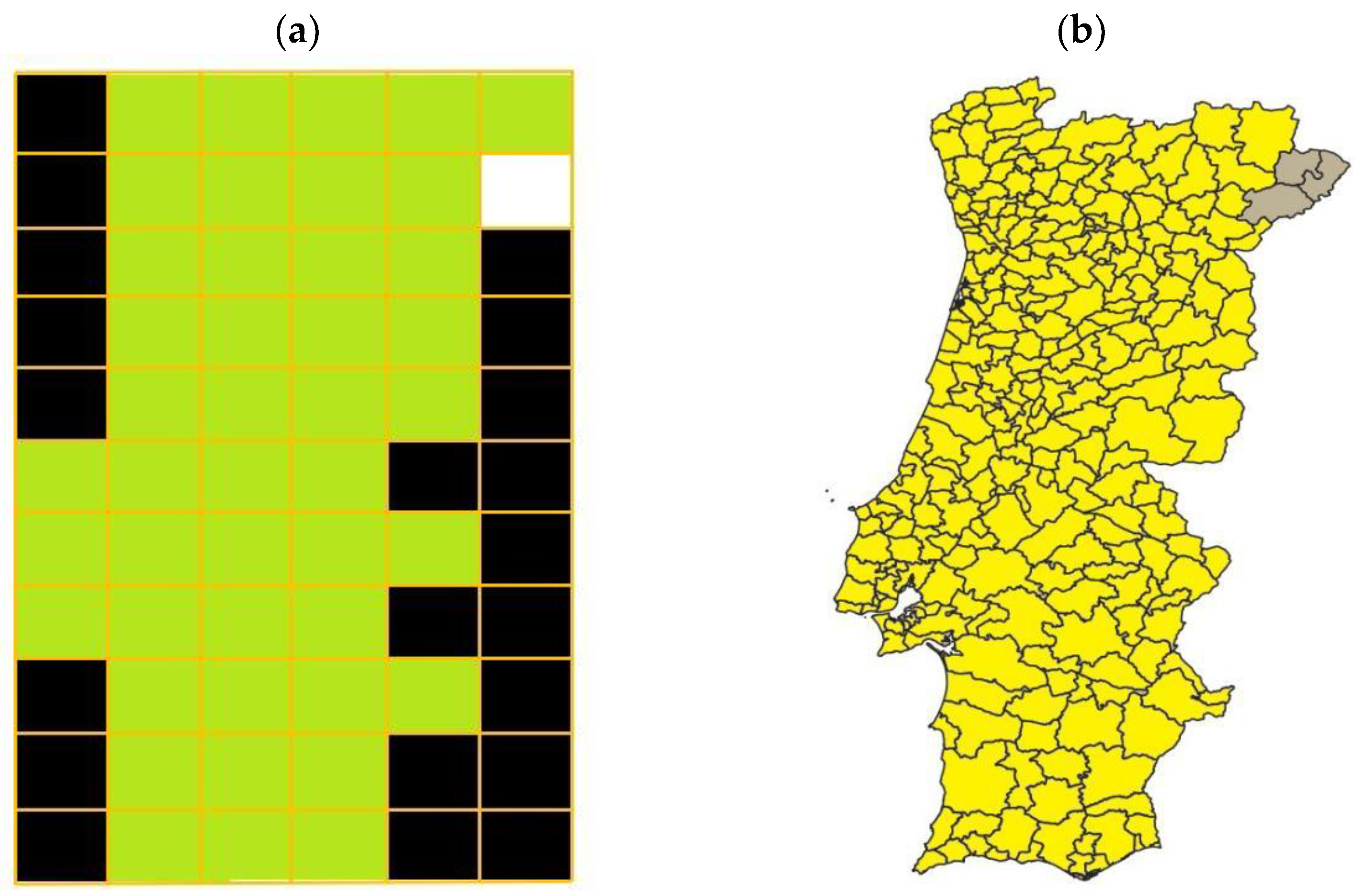
- (a)
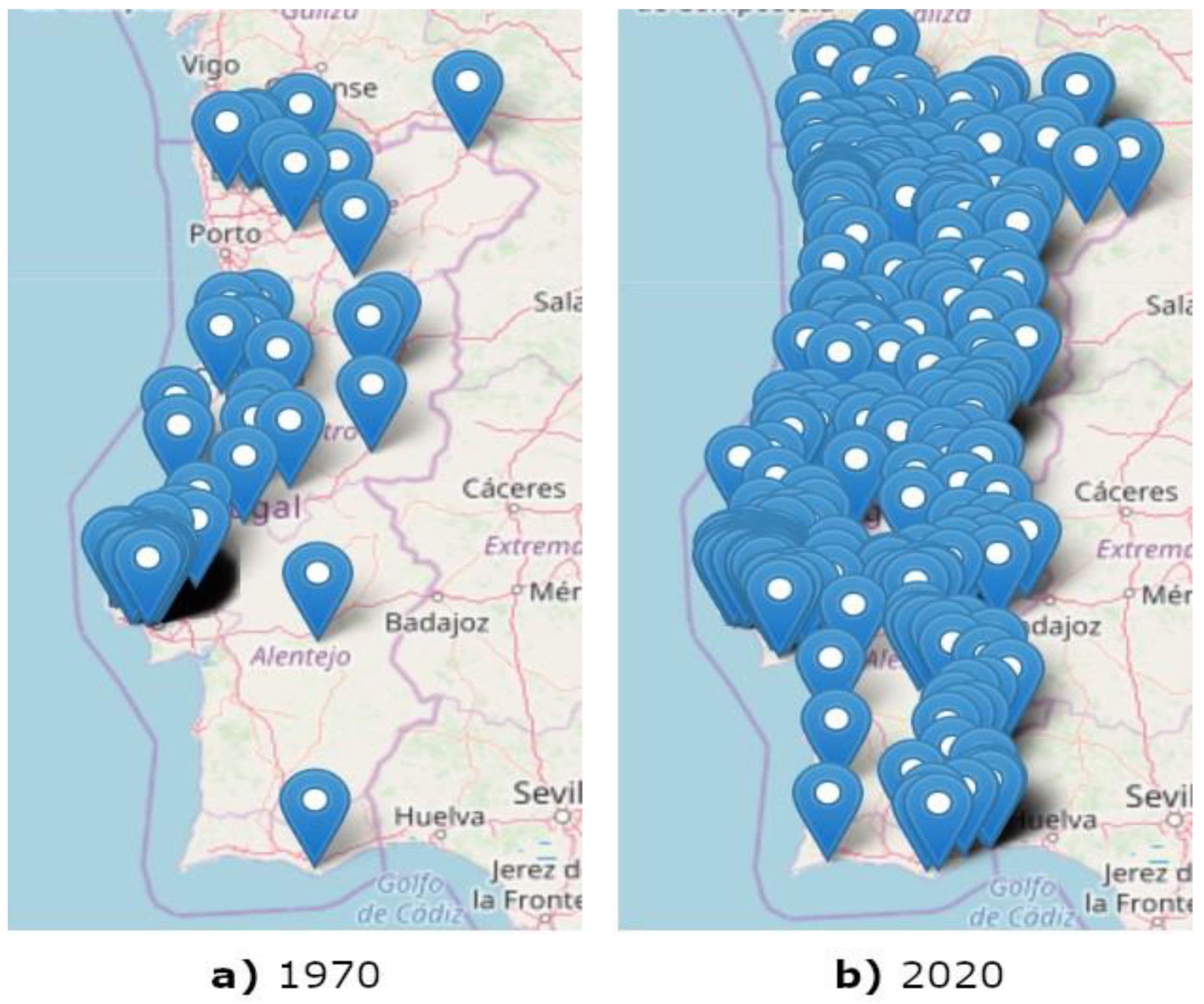
- Almost every reported observation of termites was performed due to an infestation of wood in buildings. This means that there could be many other locations where termites existed (and were deteriorating agents of wood in buildings) but were not reported to date. This would suggest that, as years go by and the number of reports increase, the number of affected counties should also increase. This is exactly what has happened and, nowadays, R. grassei distribution in applied wood in mainland Portugal covers a much larger area (as can be seen in Figure 4b).
- (b)
- The variables used were ecological variables only, whereas the presence data were almost exclusively from infestations reports. No variables considering timeframe and infestation dynamics as well as on the likelihood of infestation report and time since first occurrence were considered.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J. Predicting species distributions from museum and herbarium records using multiresponse models fitted with multivariate adaptive regression splines. Divers. Distrib. 2007, 13, 265–275. [Google Scholar] [CrossRef]
- Tonini, F.; Divino, F.; Lasinio, G.J.; Hochmair, H.H.; Scheffrahn, R.H. Predicting the geographical distribution of two invasive termite species from occurrence data. Environ. Entomol. 2014, 43, 1135–1144. [Google Scholar] [CrossRef][Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Warton, D.I.; Shepherd, L.C. Poisson point process models solve the “pseudo-absence problem” for presence-only data in ecology. Ann. Appl. Stat. 2010, 4, 1383–1402. [Google Scholar]
- von Neumann, J. Theory of Self-Reproducing Automata; University of Illinois Press: Urbana, IL, USA, 1966; Volume 1102024. [Google Scholar]
- Breckling, B.; Peer, G.; Matsinos, Y.G. Modelling Complex Ecological Dynamics; Springer: Berlin/Heidelberg, Germany, 2011; pp. 105–117. [Google Scholar]
- Austin, J.W.; Szalanski, A.L.; Myles, T.G.; Borges, P.A.; Nunes, L.; Scheffrahn, R.H. First record of Reticulitermes flavipes (Isoptera: Rhinotermitidae) from Terceira Island (Azores, Portugal). Fla. Entomol. 2012, 95, 196–198. [Google Scholar] [CrossRef]
- Nobre, T.; Nunes, L.; Eggleton, P.; Bignell, D. Distribution and genetic variation of Reticulitermes (Isoptera: Rhinotermitidae) in Portugal. Heredity 2006, 96, 403–409. [Google Scholar] [CrossRef]
- Cruz, H.; Jones, D.; Nunes, L. Wood. In Materials for Construction and Civil Engineering; Gonçalves, M.C., Margarido, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 557–583. [Google Scholar]
- Meulenkamp, J.E.; Sissingh, W.; Calvo, J.P.; Daams, R.; Studencka, B.; Londeix, L.; Cahuzac, B.; Kovac, M.; Nagymarosy, A.; Rusu, A.; et al. Late Lutetian (44–41 Ma). In Atlas Peri-Tethys, Palaeogeographical Maps—Explanatory Notes; Crasquin, S., Ed.; CCGM/CGMW: Paris, France, 2000; pp. 163–170. [Google Scholar]
- Evans, T.A.; Forschler, B.T.; Grace, J.K. Biology of invasive termites: A worldwide review. Annu. Rev. Entomol. 2013, 58, 455–474. [Google Scholar] [CrossRef]
- Clément, J.-L.; Bagnères, A.-G.; Uva, P.; Wilfert, L.; Quintana, A.; Reinhard, J. Biosystematics of Reticulitermes termites in Europe: Morphological, chemical and molecular data. Insectes Soc. 2001, 48, 202–215. [Google Scholar] [CrossRef]
- Luchetti, A.; Marini, M.; Mantovani, B. Filling the European gap: Biosystematics of the eusocial system Reticulitermes (Isoptera, Rhinotermitidae) in the Balkanic Peninsula and Aegean area. Mol. Phylogenet. Evol. 2007, 45, 377–383. [Google Scholar] [CrossRef]
- Luchetti, A.; Velona, A.; Mueller, M.; Mantovani, B. Breeding systems and reproductive strategies in Italian Reticulitermes colonies (Isoptera: Rhinotermitidae). Insectes Soc. 2013, 60, 203–211. [Google Scholar] [CrossRef]
- Lefebvre, T.; Chaline, N.; Limousin, D.; Dupont, S.; Bagnères, A.-G. From speciation to introgressive hybridization: The phylogeographic structure of an island subspecies of termite, Reticulitermes lucifugus corsicus. BMC Evol. Biol. 2008, 8, 38. [Google Scholar] [CrossRef]
- Lefebvre, T.; Vargo, E.L.; Zimmermann, M.; Dupont, S.; Kutnik, M.; Bagnères, A.-G. Subterranean termite phylogeography reveals multiple postglacial colonization events in southwestern Europe. Ecol. Evol. 2016, 6, 5987–6004. [Google Scholar] [CrossRef]
- Ghesini, S.; Marini, M. Morphology and phylogeny of Reticulitermes sp. (Isoptera, Rhinotermitidae) from Cyprus. Bull. Entomol. Res. 2012, 102, 672–681. [Google Scholar] [CrossRef]
- Baudouin, G.; Bech, N.; Bagnères, A.-G.; Dedeine, F. Spatial and genetic distribution of a north American termite, Reticulitermes flavipes, across the landscape of Paris. Urban Ecosyst. 2018, 21, 751–764. [Google Scholar] [CrossRef]
- Ferreira, M.T.; Borges, P.A.V.; Nunes, L.; Myles, T.G.; Guerreiro, O.; Scheffrahn, R.H. Termites (Isoptera) in the Azores: An overview of the four invasive species currently present in the archipelago. Arquipel. Life Mar. Sci. 2013, 30, 39–55. [Google Scholar]
- Hernández-Teixidor, D.; Suárez, D.; García, J.; Mora, D. First report of the invasive Reticulitermes flavipes (Kollar, 1837) (Blattodea, Rhinotermitidae) in the Canary Islands. J. Appl. Entomol. 2019, 143, 478–482. [Google Scholar] [CrossRef]
- Jenkins, T.M.; Dean, R.E.; Verkerk, R.; Forschler, B.T. Phylogenetic analyses of two mitochondrial genes and one nuclear intron region illuminate European subterranean termite (Isoptera: Rhinotermitidae) gene flow, taxonomy, and introduction dynamics. Mol. Phylogenet. Evol. 2001, 20, 286–293. [Google Scholar] [CrossRef]
- Ghesini, S.; Müller, G.; Marini, M. First record of the subterranean termite Reticulitermes grassei in Switzerland. Bull. Insectol. 2020, 73, 149–151. [Google Scholar]
- Seabra, A. Quelques observations sur le Calotermes flavicollis (Fab.) et le Termes lucifugus Rossi. Bol. Soc. Port. Ciênc. Nat. 1907, 1, 122–123. [Google Scholar]
- Nobre, T.; Nunes, L. Preliminary assessment of the termite distribution in Portugal. Silva Lusit. 2001, 9, 217–224. [Google Scholar]
- Nunes, L. Termite infestation risk in Portuguese historic buildings. In Proceedings of the Cost Action IE0601 “Wood Science for Conservation of Cultural Heritage” Conference, Braga, Portugal, 5–7 November 2008; Firenze University Press: Firenze, Italy, 2010; pp. 1000–1006. [Google Scholar]
- Van Rossum, G.; Drake, F.L., Jr. Python Tutorial; Centrum voor Wiskunde en Informatica Amsterdam: Amsterdam, The Netherlands, 1995; Volume 620. [Google Scholar]
- McKinney, W. Data structures for statistical computing in Python. In Proceedings of the 9th Python in Science Conference (SciPy2010), Austin, TX, USA, 28 June–3 July 2010; pp. 56–61. Available online: https://conference.scipy.org/proceedings/scipy2010/pdfs/mckinney.pdf (accessed on 16 December 2021).
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Habitants | Class |
|---|---|
| <2000 | 1 |
| [2000–4000] | 2 |
| [4000–6000] | 3 |
| [6000–8000] | 4 |
| [8000–10,000] | 5 |
| [10,000–20,000] | 6 |
| [20,000–30,000] | 7 |
| [30,000–40,000] | 8 |
| [40,000–60,000] | 9 |
| [60,000–100,000] | 10 |
| [100,000–400,000] | 11 |
| >400,000 | 12 |
| Leptosols | Class |
|---|---|
| 0% | 1 |
| <25% | 2 |
| [25–50]% | 3 |
| [50–75]% | 4 |
| >75% | 5 |
| Hours of Sunshine (per Year) | Class |
|---|---|
| [1100–8002] | 1 |
| [2101–2400] | 2 |
| [2401–2700] | 3 |
| [2701–3000] | 4 |
| [3001–3200] | 5 |
| Days of Rainfall (per Year) | Class |
|---|---|
| 0 | 1 |
| [1–50] | 2 |
| [51–75] | 3 |
| [76–100] | 4 |
| [101–110] | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sequeira, J.G.N.; Nobre, T.; Duarte, S.; Jones, D.; Esteves, B.; Nunes, L. Proof-of-Principle That Cellular Automata Can Be Used to Predict Infestation Risk by Reticulitermes grassei (Blattodea: Isoptera). Forests 2022, 13, 237. https://doi.org/10.3390/f13020237
Sequeira JGN, Nobre T, Duarte S, Jones D, Esteves B, Nunes L. Proof-of-Principle That Cellular Automata Can Be Used to Predict Infestation Risk by Reticulitermes grassei (Blattodea: Isoptera). Forests. 2022; 13(2):237. https://doi.org/10.3390/f13020237
Chicago/Turabian StyleSequeira, João G. N., Tânia Nobre, Sónia Duarte, Dennis Jones, Bruno Esteves, and Lina Nunes. 2022. "Proof-of-Principle That Cellular Automata Can Be Used to Predict Infestation Risk by Reticulitermes grassei (Blattodea: Isoptera)" Forests 13, no. 2: 237. https://doi.org/10.3390/f13020237
APA StyleSequeira, J. G. N., Nobre, T., Duarte, S., Jones, D., Esteves, B., & Nunes, L. (2022). Proof-of-Principle That Cellular Automata Can Be Used to Predict Infestation Risk by Reticulitermes grassei (Blattodea: Isoptera). Forests, 13(2), 237. https://doi.org/10.3390/f13020237