
Synanthropic Process Evaluation (with Factors Affecting Propensity to Parasitism) and Host Range within the Genus Ganoderma in Central Europe


Abstract
:1. Introduction
2. Materials and Methods
2.1. Temporal and Spatial Sampling Trends of Ganoderma Species
2.2. Host Specificity Differences among Ganoderma Species
2.3. Propensity of Ganoderma Species to Parasitism
3. Results
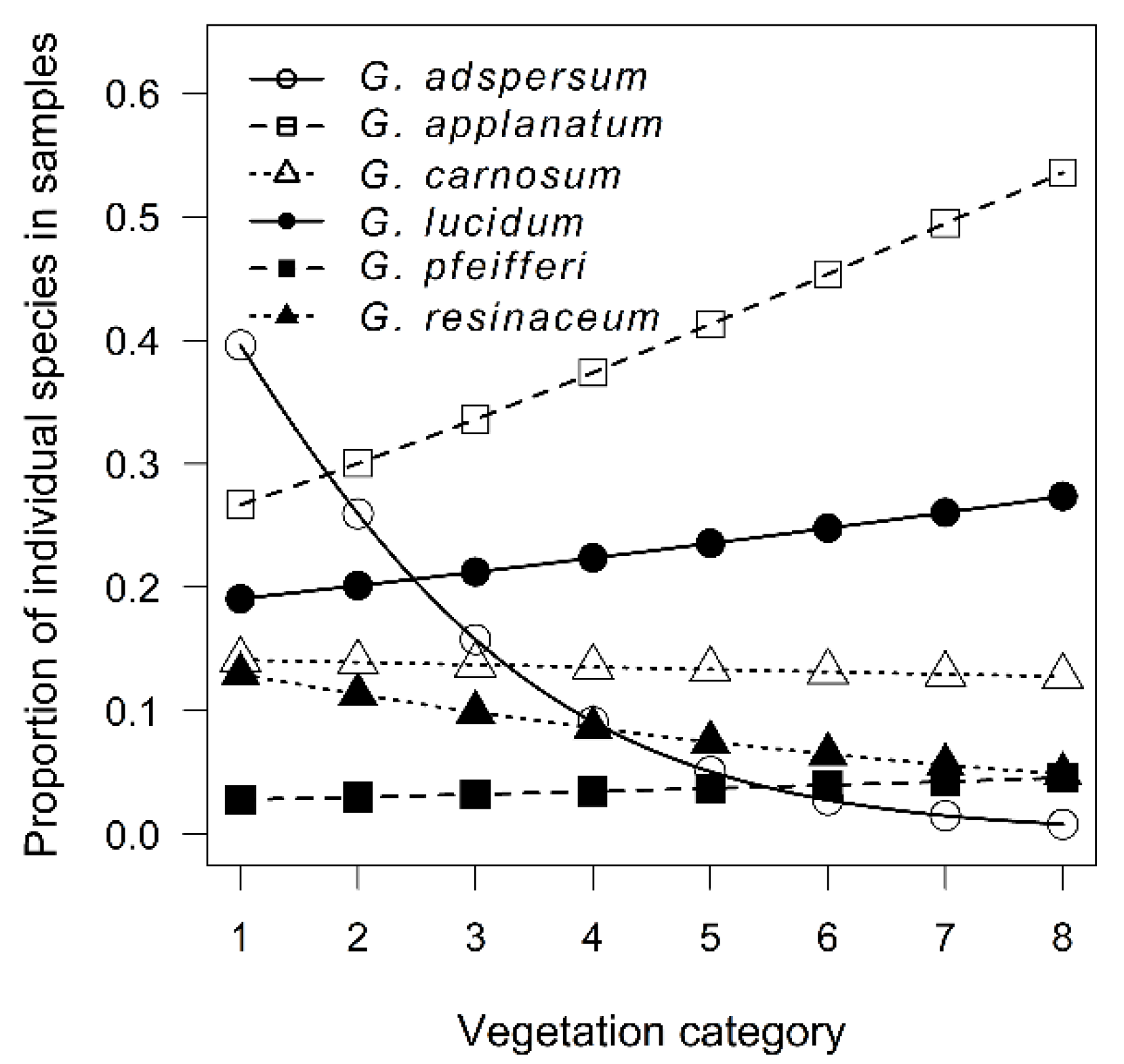
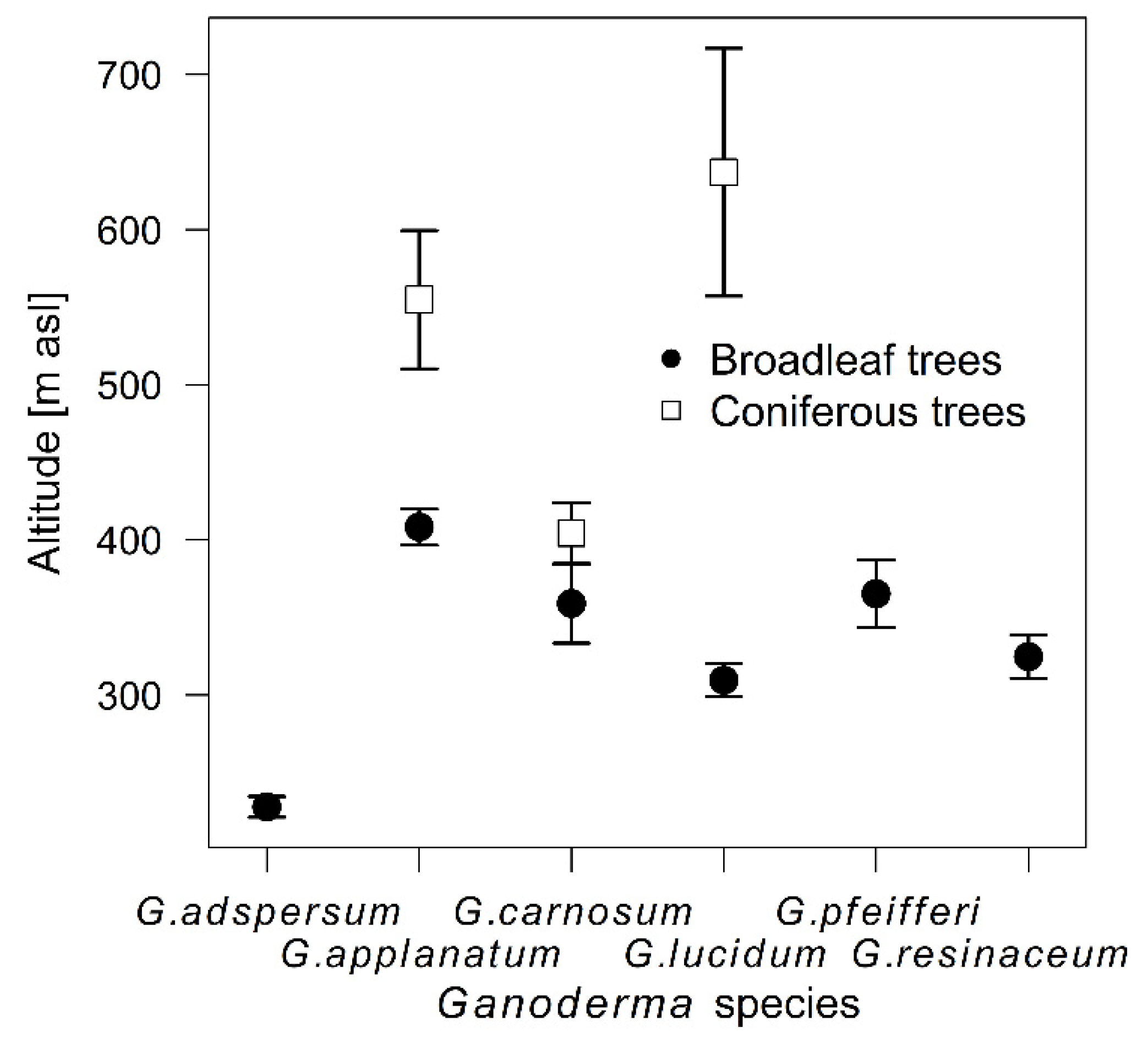
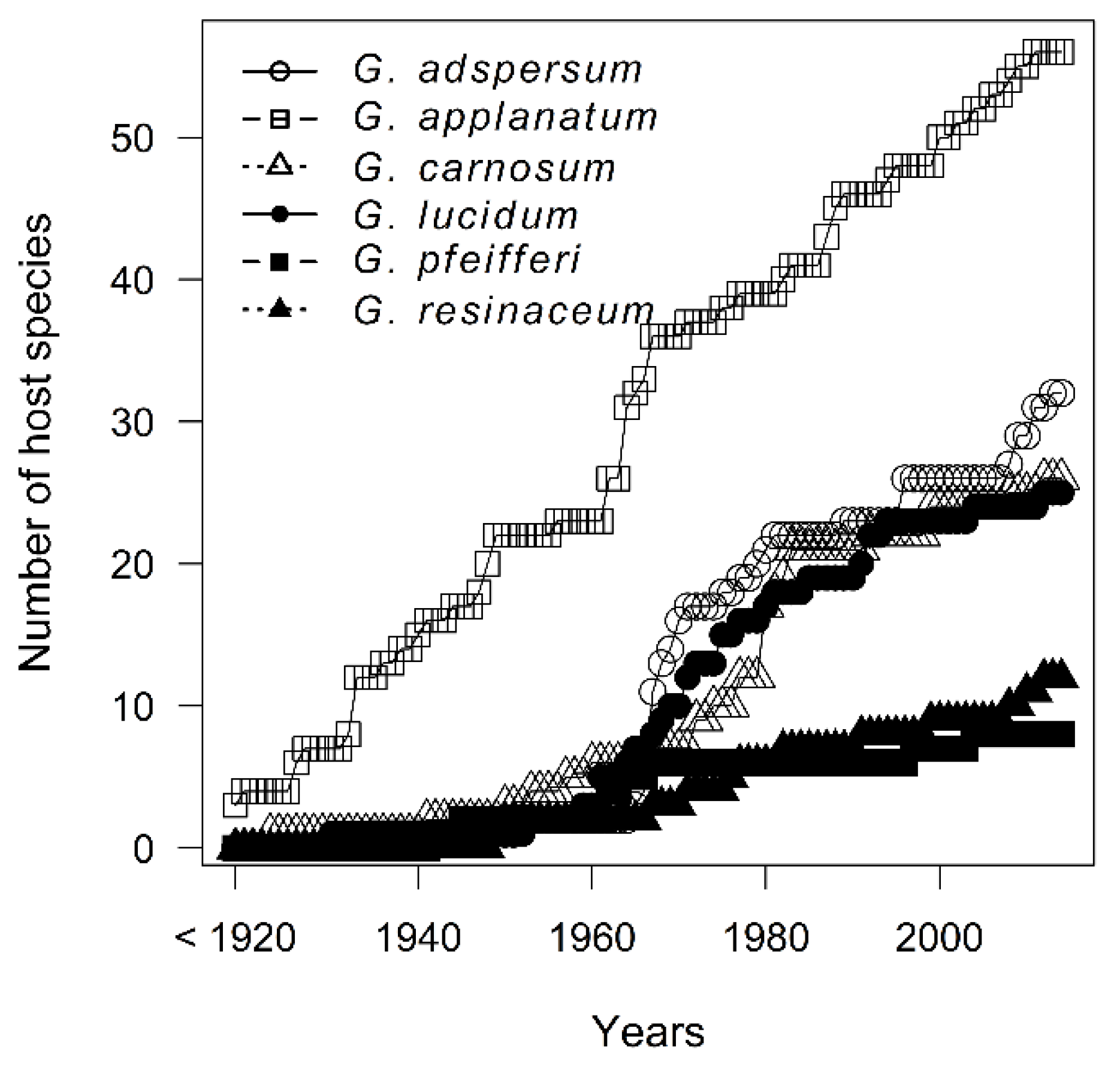
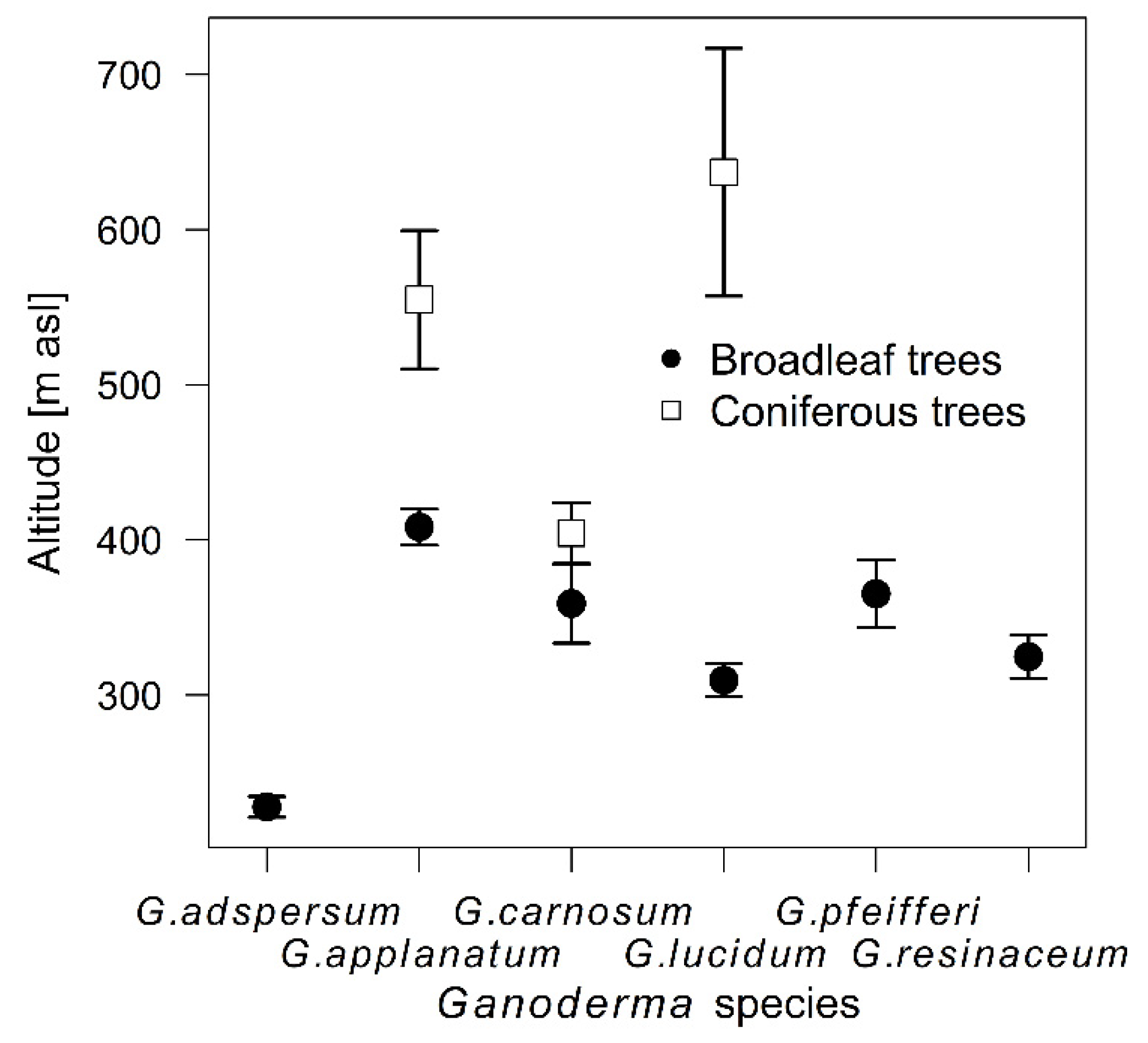
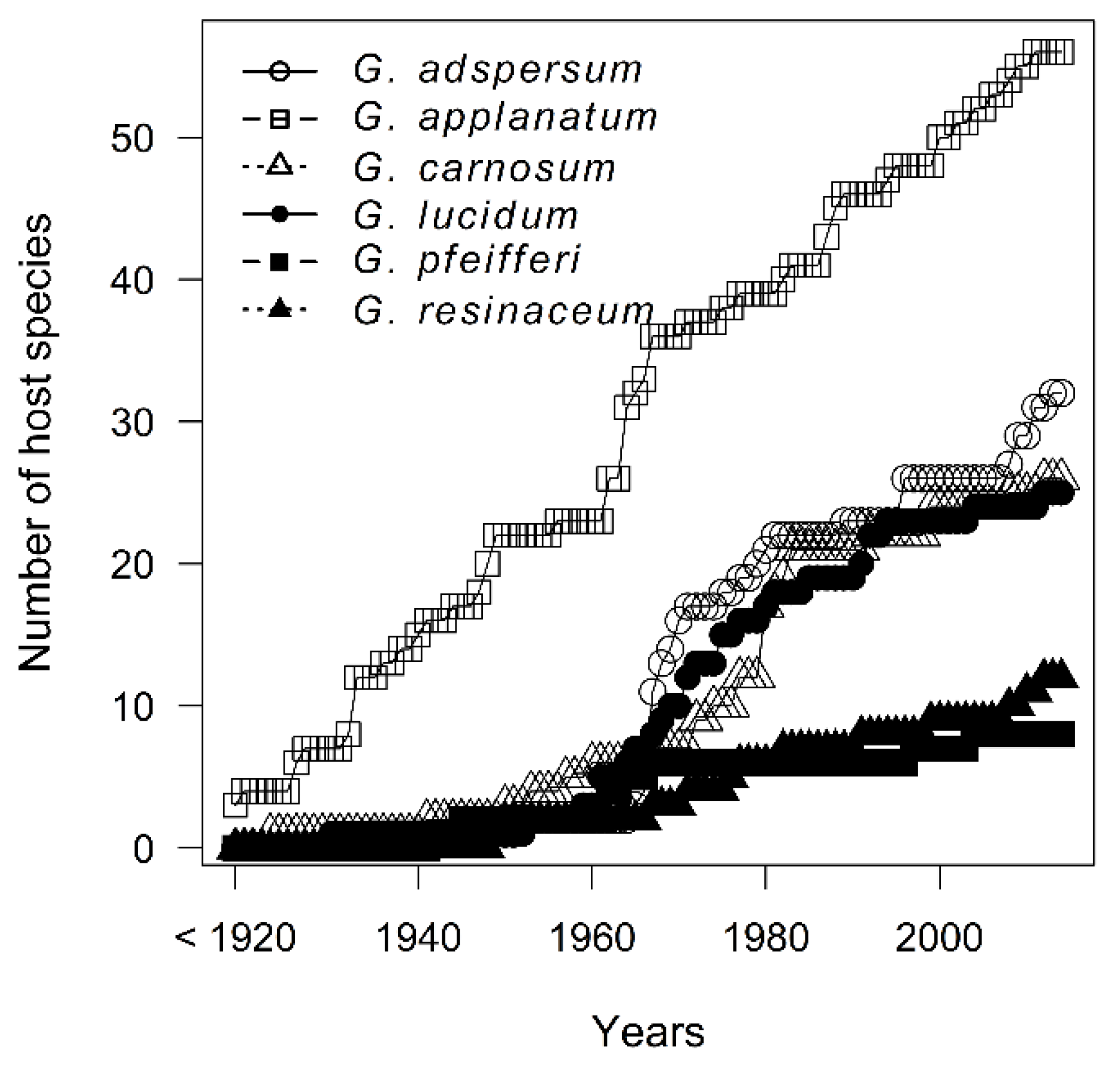
3.1. Temporal and Spatial Sampling Trends of Ganoderma Species
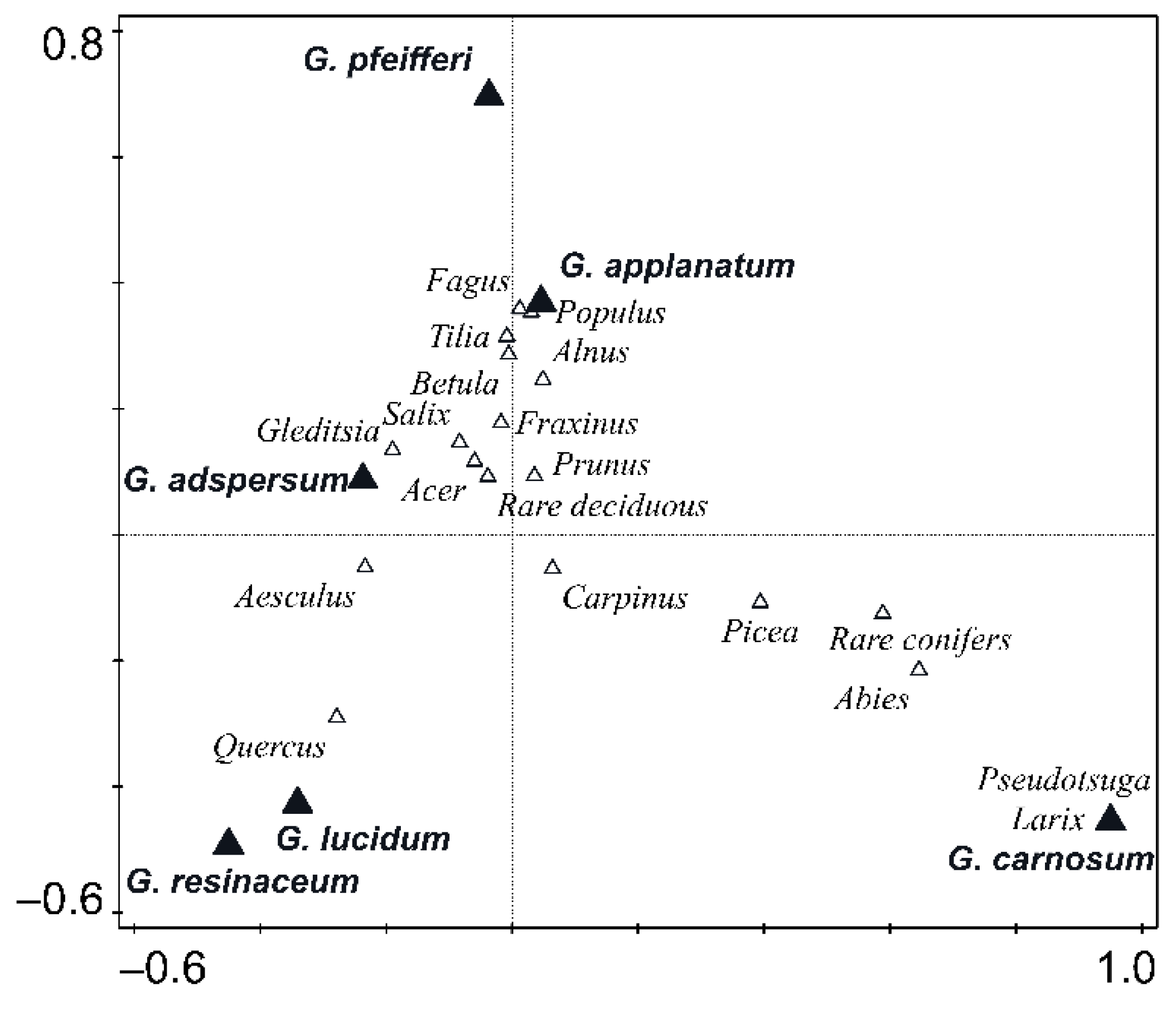
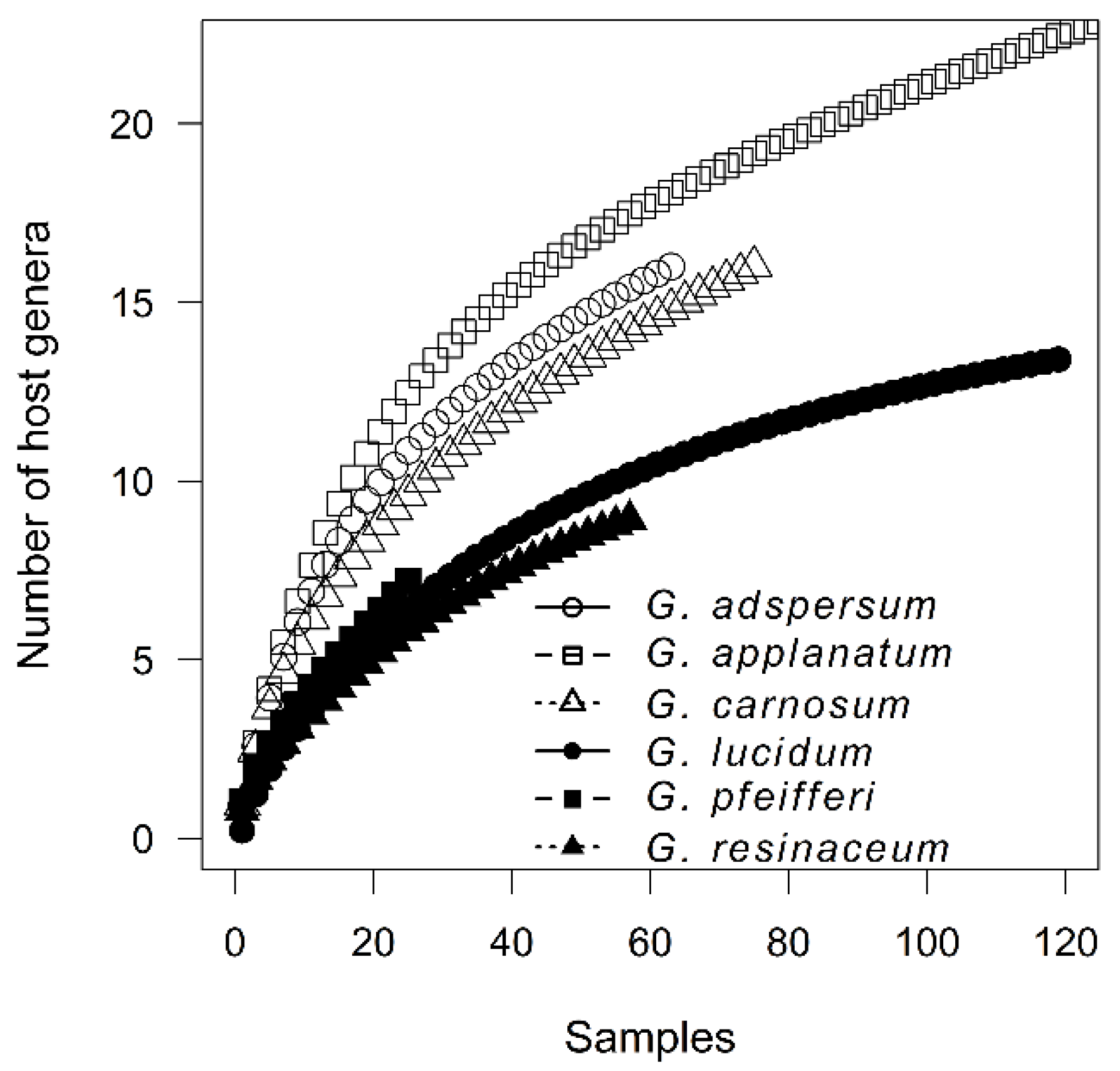
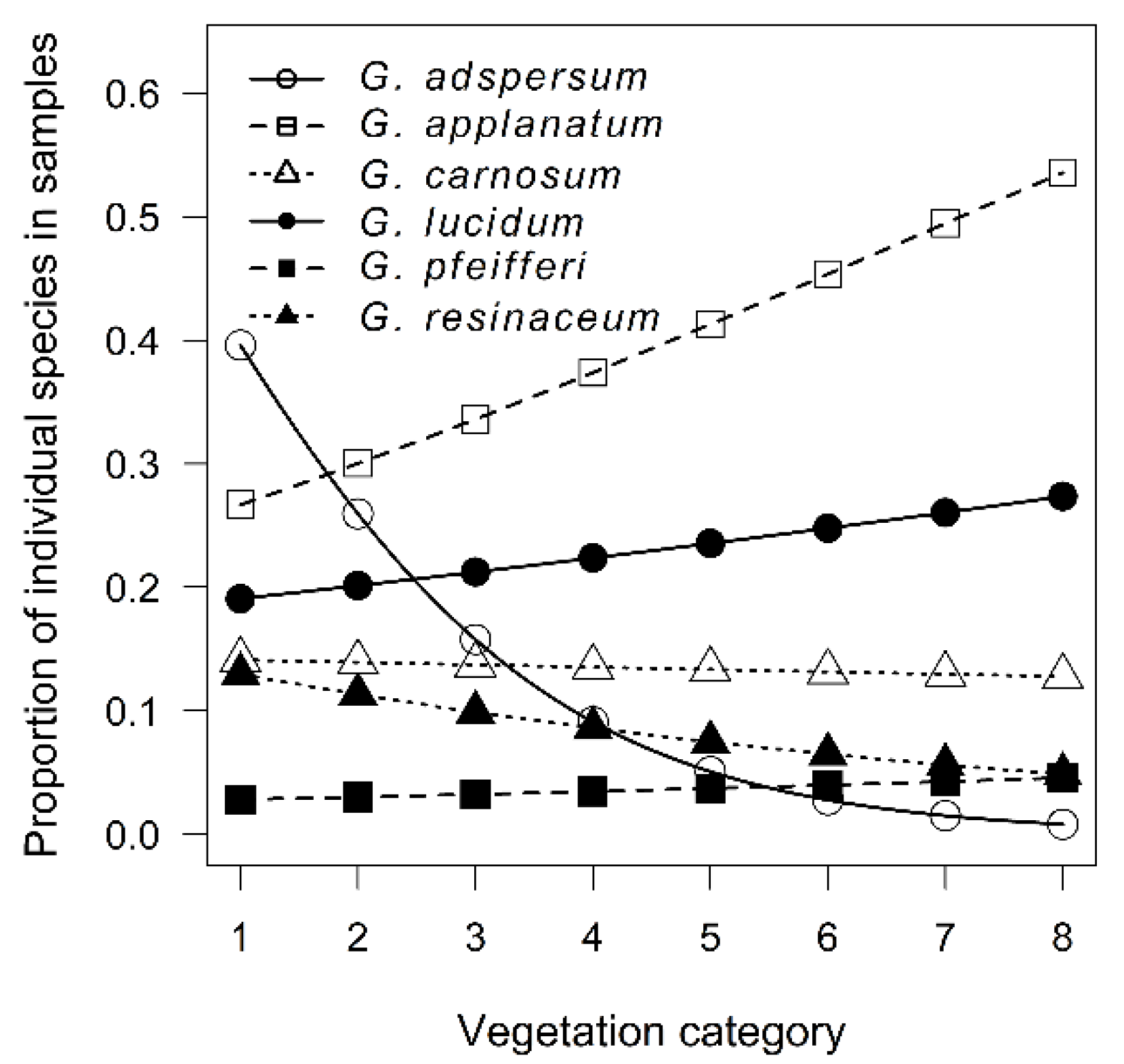
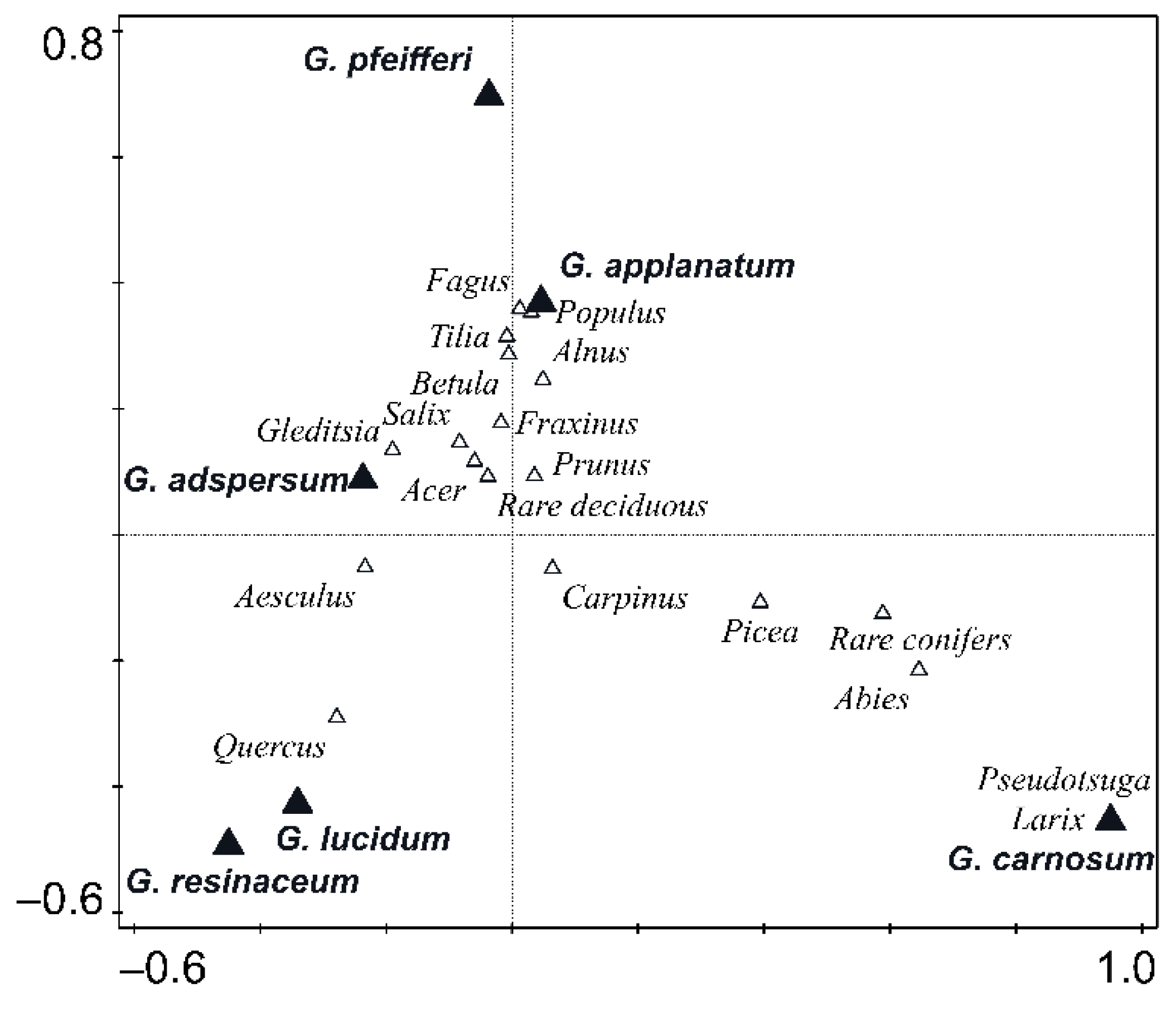
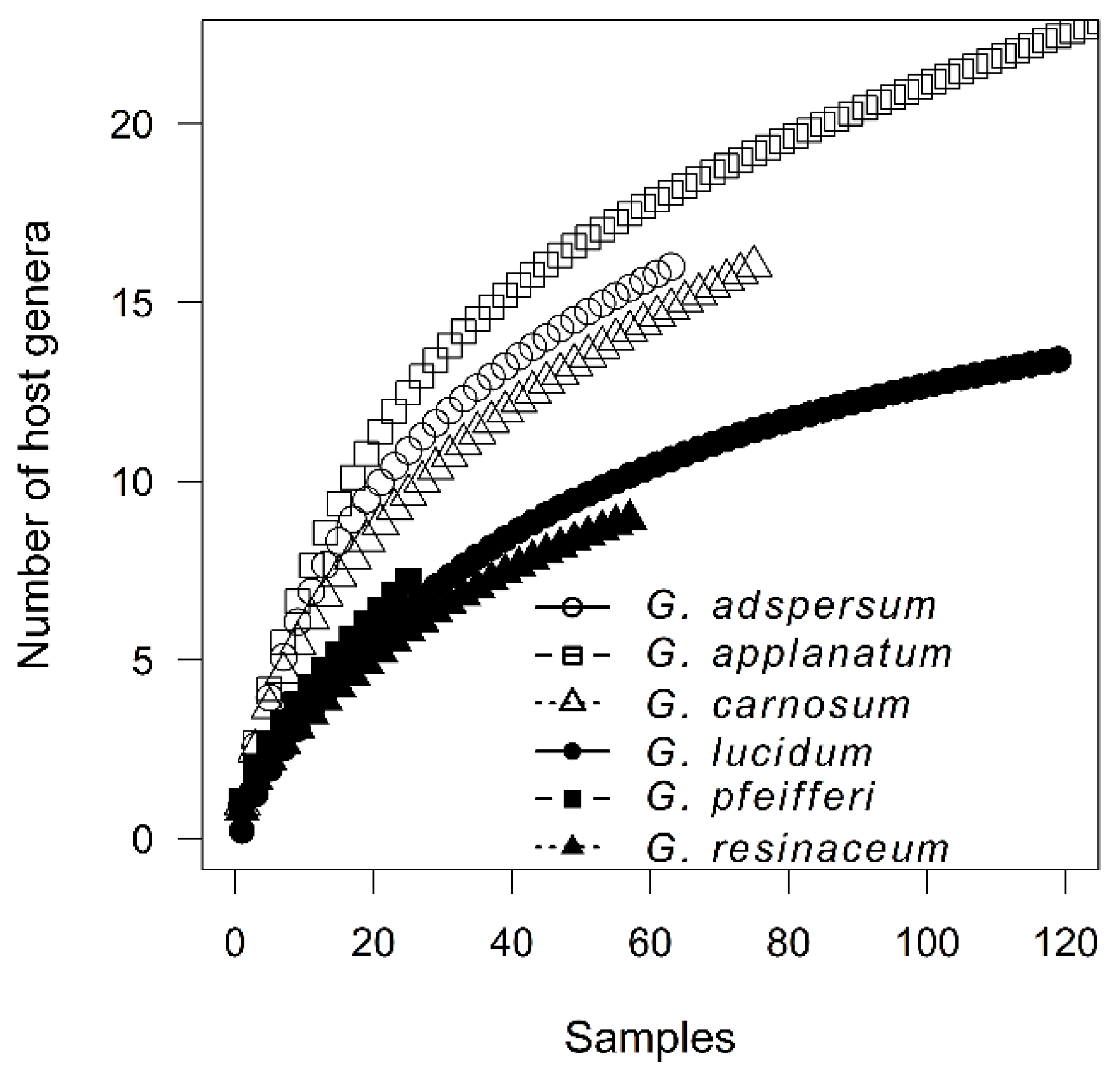
3.2. Host Specificity Differences among Ganoderma Species
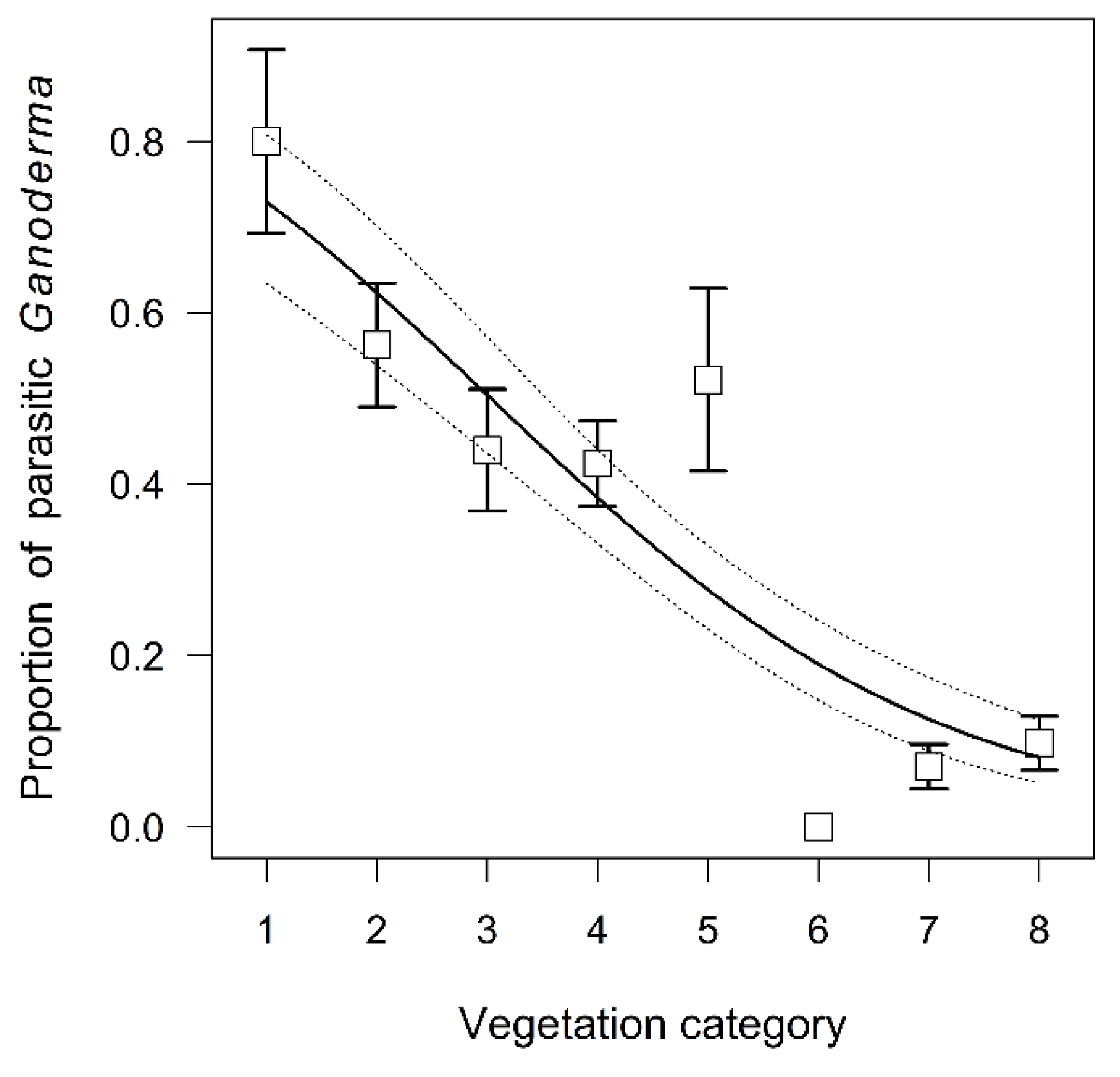
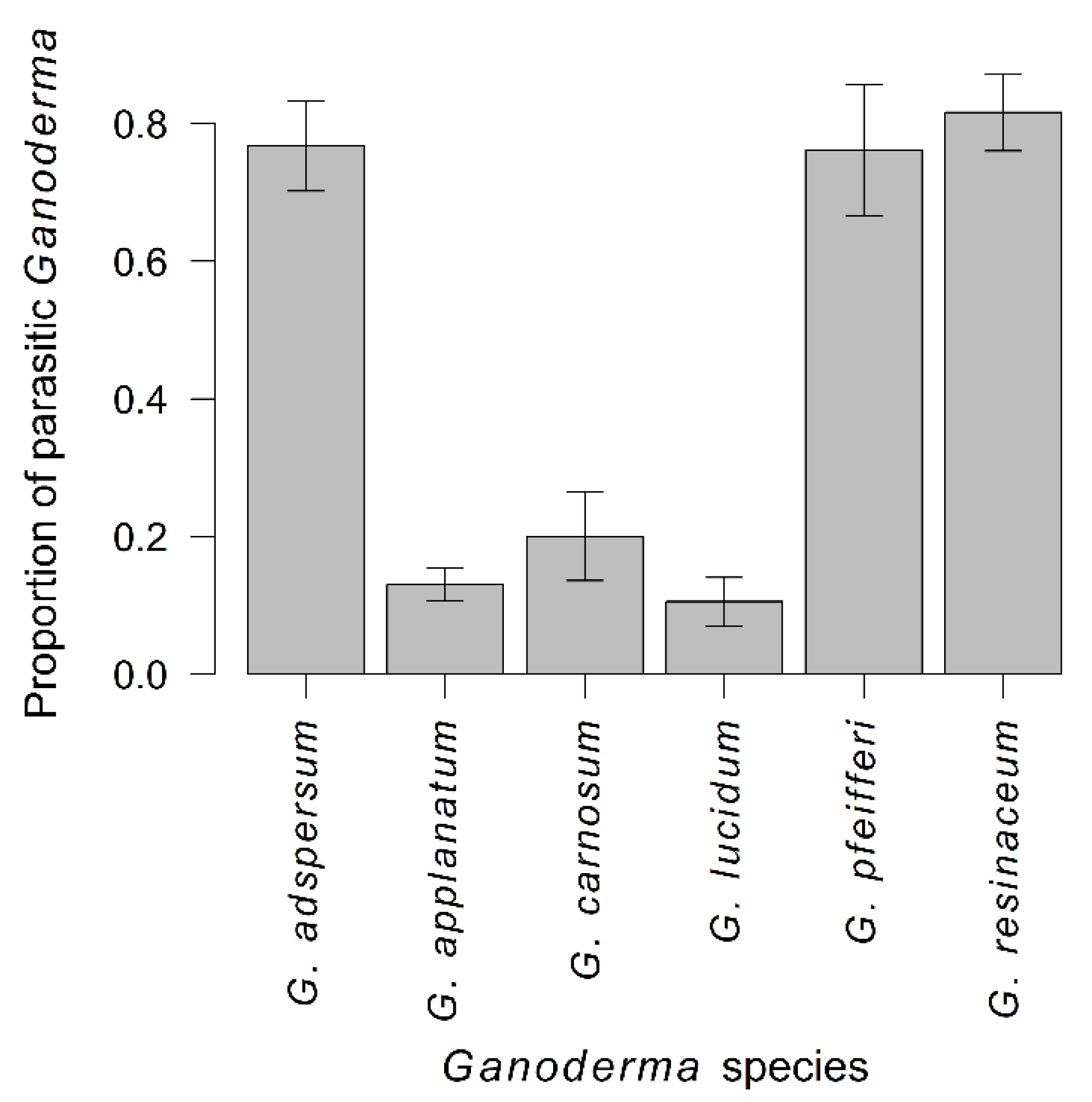
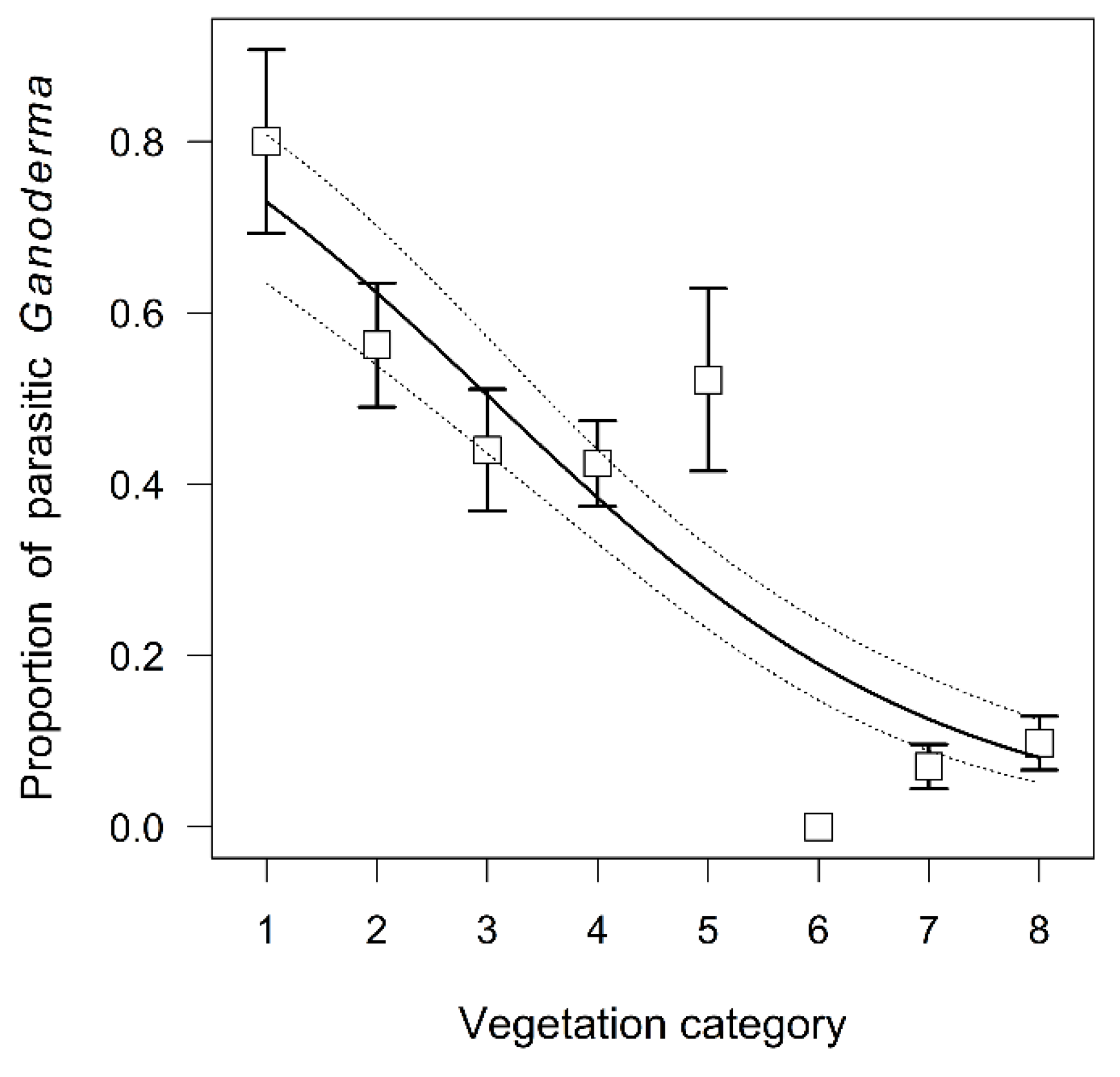
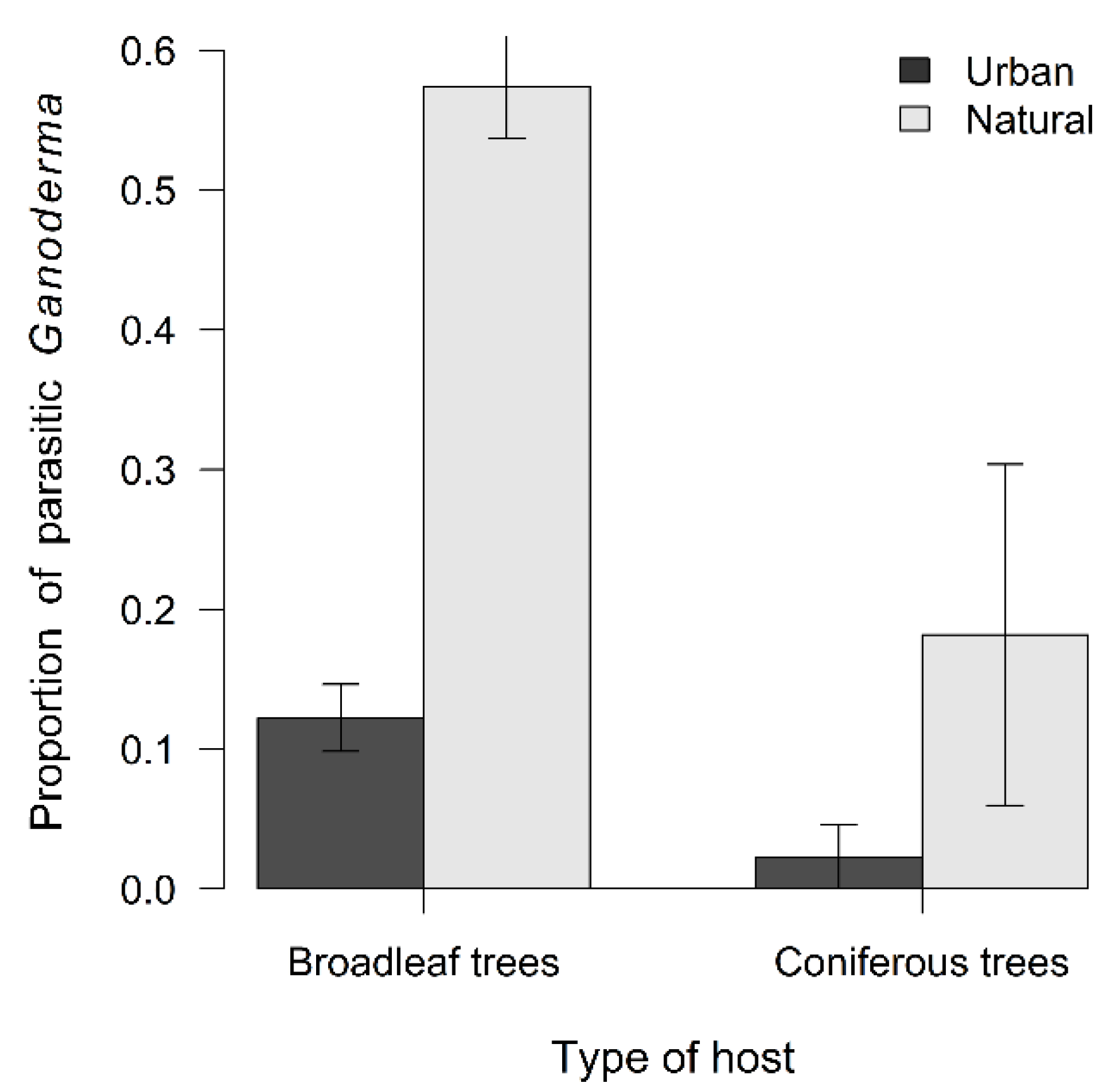
3.3. Propensity of Ganoderma Species to Parasitism
4. Discussion
4.1. Temporal and Spatial Sampling Trends of Ganoderma Species
4.2. Host Specificity Differences among Ganoderma Species
4.3. Propensity of Ganoderma Species to Parasitism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moncalvo, J.-M.; Buchanan, P.K. Molecular evidence for long distance dispersal across the Southern Hemisphere in the Ganoderma applanatum-australe species complex (Basidiomycota). Mycol. Res. 2008, 112, 425–436. [Google Scholar] [CrossRef]
- Park, Y.-J.; Kwon, O.C.; Son, E.S.; Yoon, D.E.; Han, W.; Nam, J.-Y.; Yoo, Y.; Lee, C.-S. Genetic diversity analysis of Ganoderma species and development of a specific marker for identification of medicinal mushroom Ganoderma lucidum. Afr. J. Microbiol. Res. 2012, 6, 5417–5425. [Google Scholar] [CrossRef]
- Wang, X.-C.; Xi, R.-J.; Li, Y.; Wang, N.-M.; Yao, Y.-J. The Species Identity of the Widely Cultivated Ganoderma, ‘G. lucidum’ (Ling-zhi), in China. PLoS ONE 2012, 7, e40857. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wu, S.-H.; Dai, Y.-C. Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”. Fungal Divers. 2012, 56, 49–62. [Google Scholar] [CrossRef]
- Zhou, L.-W.; Cao, Y.; Wu, S.-H.; Vlasák, J.; Li, D.-W.; Li, M.-J.; Dai, Y.-C. Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry 2015, 114, 7–15. [Google Scholar] [CrossRef]
- Tchoumi, J.M.T.; Coetzee, M.P.A.; Rajchenberg, M.; Roux, J. Taxonomy and species diversity of Ganoderma species in the Garden Route National Park of South Africa inferred from morphology and multilocus phylogenies. Mycologia 2019, 111, 730–747. [Google Scholar] [CrossRef]
- Costa-Rezende, D.; Robledo, G.; Góes-Neto, A.; Reck, M.; Crespo, E.; Drechsler-Santos, E.R. Morphological reassessment and molecular phylogenetic analyses of Amauroderma s.lat. raised new perspectives in the generic classification of the Ganodermataceae family. Persoonia Mol. Phylogeny Evol. Fungi 2017, 39, 254–269. [Google Scholar] [CrossRef]
- Hapuarachchi, K.; Karunarathna, S.C.; Phengsintham, P.; Yang, H.D.; Kakumyan, P.; Hyde, K.D.; Wen, T.C. Ganodermataceae (Polyporales): Diversity in Greater Mekong Subregion countries (China, Laos, Myanmar, Thailand and Vietnam). Mycosphere 2019, 10, 221–309. [Google Scholar] [CrossRef]
- Loyd, A.L.; Barnes, C.W.; Held, B.W.; Schink, M.J.; Smith, M.E.; Smith, J.A.; Blanchette, R.A. Elucidating “lucidum”: Distinguishing the diverse laccate Ganoderma species of the United States. PLoS ONE 2018, 13, e0199738. [Google Scholar] [CrossRef]
- Kotlaba, F.; Pouzar, Z. Ganoderma adspersum (S. Schulz.) Donk lesklokorka tmavá, dvojník lesklokorky ploské G. ap-planatum (Pers. ex S. F. Gray). Pat. Czech. Mycol. 1971, 25, 88–102. [Google Scholar]
- Breintenbach, J.; Kränzlin, F. Fungi of Switzerland; Aphyllophorales, Verlag Mykologia: Lucerne, Switzerland, 1986; Volume 2. [Google Scholar]
- Niemelä, T.; Miettinen, O. The identity of Ganoderma applanatum (Basidiomycota). Taxon 2008, 57, 963–966. [Google Scholar] [CrossRef]
- Ryvarden, L.; Melo, I. Poroid Fungi of Europe; Fungiflora: Oslo, Norway, 2014; 455p. [Google Scholar]
- Kotlaba, F.; Pouzar, Z. Distribution and ecology of Ganoderma atkinsonii in Czechoslovakia. Czech. Mycol. 1981, 35, 121–133. [Google Scholar]
- Kotlaba, F. Common polypores (Polyporales s.l.) collected on uncommon hosts. Czech. Mycol. 1997, 49, 169–188. [Google Scholar] [CrossRef]
- Gottlieb, A.M.; Ferrer, E.; Wright, J.E. rDNA analyses as an aid to the taxonomy of species of Ganoderma. Mycol. Res. 2000, 104, 1033–1045. [Google Scholar] [CrossRef]
- Al-Fatimi, M.; Wurster, M.; Lindequist, U. Chemical Composition, Antimicrobial and Antioxidant Activities of the Volatile Oil of Ganoderma pfeifferi Bres. Medicines 2016, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, P.; Buonocore, D.; Altobelli, E.; Brandalise, F.; Cesaroni, V.; Iozzi, D.; Savino, E.; Marzatico, F. Improving Training Condition Assessment in Endurance Cyclists: Effects of Ganoderma lucidum and Ophiocordyceps sinensis Dietary Supplementation. Evidence-Based Complement. Altern. Med. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Yang, Z.L.; Feng, B. What is the Chinese “Lingzhi”? A taxonomic mini-review. Mycology 2013, 4, 1–4. [Google Scholar] [CrossRef]
- Schmidt, O.; Gaiser, O.; Dujesiefken, D. Molecular identification of decay fungi in the wood of urban trees. Eur. J. For. Res. 2011, 131, 885–891. [Google Scholar] [CrossRef]
- Kotlaba, F. Geographical Distribution and Ecology of Polypores /Polyporales s.l./ in Czechoslovakia; Academia: Praha, Czech Republic, 1984. [Google Scholar]
- Gáper, J. Trúdniky na Území Slovenska a ich Šírenie v Ekosystémoch Bazídiospórami [Polypores of Slovakia and Their Distribution by Basidiospores]; Vedecké štúdie; Technical University of Zvolen: Zvolen, Slovak Republic, 1998. [Google Scholar]
- Gáperová, S. Synantropic species in the genus Ganoderma. Acta Fac. Ecol. 2001, 8, 93–98. [Google Scholar]
- Petersen, J.E. Ganoderma in Northern Europe. Lakporesvampene (Ganoderma) i Danmark og Europa. Svampe 1983, 7, 1–11. [Google Scholar]
- Ryvarden, L.; Gilbertson, R.L. European polypores—Abortiporus-Lindtneria Vol. 1. Synop. Fungorum 1993, 13, 1–387. [Google Scholar]
- Schwarze, F.W.; Ferner, D. Ganodermaon Trees—differentiation of species and studies of invasiveness. Arboric. J. 2003, 27, 59–77. [Google Scholar] [CrossRef]
- Kout, J. Lesklokorka tmavá (Ganoderma adspersum) z Podorlické oblasti. Acta Musei Richnoviensis Sect. Natur. 2011, 18, 25–28. [Google Scholar]
- Papp, V.; Szabó, I. Distribution and Host Preference of Poroid Basidiomycetes in Hungary I. Ganoderma. Acta Silv. Lignaria Hung. 2013, 9, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Zicha, O. BioLib: Biological Library. 2021. Available online: https://www.biolib.cz/cz/glossaryterm/id229/ (accessed on 15 January 2021).
- Novák, J.; Zicha, O. BioLib Nástroj Pro Výpočet Mapovacích čtverců Metodou KFME. Available online: https://www.biolib.cz/cz/toolKFME/ (accessed on 8 December 2020).
- Jeffrey, D. Biodiversity in the City. In Proceedings of the A One-Day International Conference, Dublin, Ireland, 12 September 2002. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.9.13. 2013. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 5 December 2020).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 5 December 2020).
- Morales, M. Sciplot: Scientific Graphing Functions for Factorial Designs. R Package Version 1.2-0. Available online: https://CRAN.R-project.org/package=sciplot (accessed on 5 December 2020).
- Bowman, A.W.; Azzalini, A. R Package ‘sm’: Nonparametric Smoothing Methods (Version 2.2–5.6). Available online: http://www.stats.gla.ac.uk/~adrian/sm (accessed on 5 December 2020).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simspon, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5–4. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 December 2020).
- Cayuela, L.; Gotelli, N.J. RareNMtests: Ecological and Biogeographical Null Model Tests for Comparing Rarefaction Curves. R Package Version 1.1. 2014. Available online: https://CRAN.R-project.org/package=rareNMtests (accessed on 5 December 2020).
- Ter Braak, C.J.F.; Šmiauler, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); CRAN: Ithaca, NY, USA, 2012; 500p, Available online: www.canoco.com (accessed on 5 December 2020).
- Fox, J.; Weisberg, S.; Adler, D.; Bates, D. Package ‘Car’; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Bowman, A.W.; Azzalini, A. Applied Smoothing Techniques for Data Analysis: The Kernel Approach with S-Plus Illustrations; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Coleman, B.D.; Mares, M.A.; Willig, M.R.; Hsieh, Y.-H. Randomness, Area, and Species Richness. Ecology 1982, 63, 1121–1133. [Google Scholar] [CrossRef]
- Tulloch, A.I.T.; Szabo, J.K. A behavioural ecology approach to understand volunteer surveying for citizen science datasets. Emu Austral. Ornithol. 2012, 112, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Gower, G.; Fenderson, L.; Salis, A.T.; Helgen, K.; van Loenen, A.L.; Heiniger, H.; Hofman-Kamińska, E.; Kowalczyk, R.; Mitchell, K.J.; Llamas, B.; et al. Widespread male sex bias in mammal fossil and museum collections. Proc. Natl. Acad. Sci. USA 2019, 116, 19019–19024. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.E.; Birkhead, T.R. Avian egg collections: Museum collection bias driven by shape and size. J. Avian Biol. 2020, 51. [Google Scholar] [CrossRef]
- Pyke, G.H.; Ehrlich, P.R. Biological collections and ecological/environmental research: A review, some observations and a look to the future. Biol. Rev. 2010, 85, 247–266. [Google Scholar] [CrossRef]
- Isaac, N.J.B.; Pocock, M. Bias and information in biological records. Biol. J. Linn. Soc. 2015, 115, 522–531. [Google Scholar] [CrossRef] [Green Version]
- Isaac, N.J.B.; Van Strien, A.J.; August, T.; De Zeeuw, M.P.; Roy, D.B. Statistics for citizen science: Extracting signals of change from noisy ecological data. Methods Ecol. Evol. 2014, 5, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Tingley, M.W.; Beissinger, S.R. Detecting range shifts from historical species occurrences: New perspectives on old data. Trends Ecol. Evol. 2009, 24, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Isaac, N.; Cruickshanks, K.L.; Weddle, A.M.; Rowcliffe, M.; Brereton, T.M.; Dennis, R.L.H.; Shuker, D.M.; Thomas, C. Distance sampling and the challenge of monitoring butterfly populations. Methods Ecol. Evol. 2011, 2, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Kadmon, R.; Farber, O.; Danin, A. Effect of Roadside Bias on the accuracy of predictive maps produced by bioclimatic models. Ecol. Appl. 2004, 14, 401–413. [Google Scholar] [CrossRef]
- Schwarze, F.W.M.R.; Engels, J.; Maitheck, C. Holzzersetzende Pilze in Bäumen; Rombach Verlag: Freiburg, Germany, 1994; 245p. [Google Scholar]
- Jahn, H. Pilze an Bäumen; Patzer Verlag: Berlin-Hannover, Germany, 2005; 275p. [Google Scholar]
- Tello, M.L.; Tomalak, M.; Siwecki, R.; Gáper, J. Biotic urban growing conditions threats, pests and diseases. In Urban Forests and Trees; Konijnendijk, C.C., Nilsson, K., Randrup, T., Schipperijn, J., Eds.; Springer: Heidelberg, Germany, 2005; 520p. [Google Scholar]
- Kout, J.; Vlasák, J. Rare polypores of the Czech Republic, chiefly from South Bohemia. Czech. Mycol. 2009, 108, 22–33. [Google Scholar]
- Černý, A. Parazitické Dřevokazné Houby. (Parasitic Wood-Decaying Fungi); Státní zemědělské nakladatelství: Praha, Czech Republic, 1989. [Google Scholar]
- Kout, J.; Vlasák, J. New or rare polyporoid fungi from the Pilsen Region part 2. Erica 2013, 20, 55–66. [Google Scholar]
- Sokół, S. Ganodermataceae Polski: Taksonomia, Ekologia i Rozmieszczenie; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2000; 134p. [Google Scholar]
- Christensen, M.; Heilmann-Clausen, J.; Walleyn, R.; Adamčík, S. Wood-inhabiting fungi as indicators of conservation value in European beech forests. In Monitoring and Indicators of Forest Biodiversity in Europe from Ideas to Operationality; Marchetti, M., Ed.; European Forestry Institute: Saarijärvi, Finland, 2004; pp. 229–237. [Google Scholar]
- Tortić, M. Distribution of Polypores in Yugoslavia, II. Ganoderma. Acta Bot. Croat. 1985, 44, 59–71. [Google Scholar]
- Kotlaba, F.; Pouzar, Z. Ecology of the polypore Ganoderma adspersum in Bohemia. Czech. Mycol. 2009, 109, 11–15. [Google Scholar]
- Kotlaba, F.; Pouzar, Z. Ecology of Ganoderma resinaceum and its expansion in Bohemia. Czech. Mycol. 2009, 109, 14–19. [Google Scholar]
- Bernicchia, A. Polyporaceae s.l. Fungi Europaei; Massimo Candusso: Alassio, Italy, 2005; 808p. [Google Scholar]
- Suarez, A.V.; Tsutsui, N.D. The value of museum collections for research and society. Bioscience 2004, 54, 66–74. [Google Scholar] [CrossRef]
- Lavoie, C. Biological collections in an ever changing world: Herbaria as tools for biogeographical and environmental studies. Perspect. Plant. Ecol. Evol. Syst. 2013, 15, 68–76. [Google Scholar] [CrossRef]
- Heberling, J.M.; Isaac, B.L. Herbarium specimens as exaptations: New uses for old collections. Am. J. Bot. 2017, 104, 963–965. [Google Scholar] [CrossRef] [Green Version]
- Meineke, E.K.; Davis, C.C.; Davies, T.J. The unrealized potential of herbaria for global change biology. Ecol. Monogr. 2018, 88, 505–525. [Google Scholar] [CrossRef]
- Downey, P.O. A regional examination of the mistletoe host species inventory. Cunninghamia 2004, 8, 354–361. [Google Scholar]
- Norton, D.A.; Carpenter, M. Mistletoes as parasites: Host specificity and speciation. Trends Ecol. Evol. 1998, 13, 101–105. [Google Scholar] [CrossRef]
- Norton, D.A.; De Lange, P.J. Host specificity in parasitic mistletoes (Loranthaceae) in New Zealand. Funct. Ecol. 1999, 13, 552–559. [Google Scholar] [CrossRef]
- Jeffree, C.; Jeffree, E. Redistribution of the Potential Geographical Ranges of Mistletoe and Colorado Beetle in Europe in Response to the Temperature Component of Climate Change. Funct. Ecol. 1996, 10, 562–577. [Google Scholar] [CrossRef]
- Norton, D.A.; Smith, M.S. Why might roadside mulgas be better mistletoe hosts? Austral. Ecol. 1999, 24, 193–198. [Google Scholar] [CrossRef]
- Heilmann-Clausen, J.; Maruyama, P.K.; Bruun, H.H.; Dimitrov, D. Citizen science data reveal ecological, historical and evolutionary factors shaping interactions between woody hosts and wood-inhabiting fungi. New Phytol. 2016, 212, 1072–1082. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Büche, B.; Szallies, A.; Thorn, S.; Ulyshen, M.D.; Müller, J. Microclimate and habitat heterogeneity as the major drivers of beetle diversity in dead wood. J. Appl. Ecol. 2016, 53, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Karadelev, M.; Rusevska, K.; Kajevska, I. Distribution and ecology of genus Ganoderma (Ganodermataceae) in the Republic of Macedonia. In Proceedings of the International Conference on Biological and Environmental Sciences, Hurghada, Egypt, 13–16 March 2008; Kule, D., Malollari, I., Haxhimihali, D., Çullaj, A., Miho, A., Babani, F., Dhora, D., Shumka, S., Mustafa, B., Abeshi, P., et al., Eds.; FNS Tirana University: Tirana, Albania, 2008; pp. 320–326. [Google Scholar]
- Vlasák, J. Polypores: Collection of Dr. Josef Vlasák, Hluboká nad Vltavou, Czech Republic, Edition 18. II. 2015. Available online: http://mykoweb.prf.jcu.cz/polypores/ (accessed on 20 May 2019).
- Kuthan, J.; Kotlaba, F. Makromyzeten der Bulgarischen Schwarzmeerküste und Einiger Orte im Landesinnern Bulgariens; Národní Muzeum: Praha, Czech Republic, 1988. [Google Scholar]
- De Simone, D.; Annesi, T. Occurrence of Ganoderma adspersum on Pinus pinea. Phytopathol. Mediterr. 2012, 51, 374–382. [Google Scholar]
- Niemelä, T.; Kotiranta, H. Polypore survey of Finland 4. Phaeolus, Fistulina, Ganoderma and Ischnoderma. Karstenia 1986, 26, 57–64. [Google Scholar] [CrossRef]
- Szczepkowski, A.; Piętka, J. New localities and new host of Ganoderma pfeifferi in Poland. Acta Mycol. 2014, 38, 59–63. [Google Scholar] [CrossRef]
- Krah, F.-S.; Bässler, C.; Heibl, C.; Soghigian, J.; Schaefer, H.; Hibbett, D.S. Evolutionary dynamics of host specialization in wood-decay fungi. BMC Evol. Biol. 2018, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.-W.; Dai, Y.-C. Recognizing ecological patterns of wood-decaying polypores on gymnosperm and angiosperm trees in northeast China. Fungal Ecol. 2011, 5, 230–235. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Level | Df | LogLik | AICc | AICc Δ |
|---|---|---|---|---|
| Genus | 216 | −995.025 | 2450.5 | 0.00 |
| Order | 66 | −1180.481 | 2495.5 | 45.01 |
| Family | 108 | −1136.384 | 2495.7 | 45.17 |
| Species | 510 | −823.424 | 2840.6 | 390.13 |
| Phyllum | 12 | −1446.361 | 2916.8 | 466.28 |
| Host Genera | G. adspersum | G. applanatum | G. carnosum | G. lucidum | G. pfeifferi | G. resinaceum |
|---|---|---|---|---|---|---|
| Quercus | 16 | 20 | 4 | 79 | 1 | 43 |
| Fagus | 2 | 39 | 2 | 4 | 17 | 1 |
| Tilia | 7 | 36 | 0 | 1 | 1 | 0 |
| Acer | 8 | 16 | 2 | 7 | 3 | 1 |
| Picea | 0 | 15 | 16 | 5 | 0 | 0 |
| Abies | 0 | 9 | 21 | 1 | 0 | 0 |
| Prunus | 6 | 12 | 3 | 2 | 0 | 1 |
| Carpinus | 0 | 11 | 4 | 7 | 0 | 1 |
| Salix | 2 | 14 | 0 | 3 | 0 | 2 |
| Aesculus | 6 | 4 | 0 | 4 | 1 | 4 |
| Populus | 1 | 17 | 0 | 0 | 0 | 0 |
| Alnus | 0 | 13 | 1 | 2 | 1 | 0 |
| Fraxinus | 3 | 9 | 1 | 0 | 1 | 2 |
| Betula | 1 | 11 | 0 | 1 | 0 | 0 |
| Pseudotsuga | 0 | 0 | 11 | 0 | 0 | 0 |
| Gleditsia | 5 | 1 | 0 | 0 | 0 | 0 |
| Larix | 0 | 0 | 5 | 0 | 0 | 0 |
| Pyrus | 1 | 2 | 1 | 0 | 0 | 0 |
| Robinia | 0 | 2 | 0 | 2 | 0 | 0 |
| Juglans | 2 | 1 | 0 | 0 | 0 | 0 |
| Platanus | 0 | 0 | 0 | 0 | 0 | 3 |
| Ulmus | 0 | 2 | 0 | 1 | 0 | 0 |
| Corylus | 1 | 1 | 0 | 0 | 0 | 0 |
| Malus | 1 | 0 | 1 | 0 | 0 | 0 |
| Pinus | 0 | 1 | 1 | 0 | 0 | 0 |
| Spiraea | 1 | 1 | 0 | 0 | 0 | 0 |
| Ailanthus | 0 | 1 | 0 | 0 | 0 | 0 |
| Cedrus | 0 | 0 | 1 | 0 | 0 | 0 |
| Cydonia | 0 | 1 | 0 | 0 | 0 | 0 |
| Davidia | 0 | 1 | 0 | 0 | 0 | 0 |
| Frangula | 0 | 1 | 0 | 0 | 0 | 0 |
| Hibiscus | 0 | 1 | 0 | 0 | 0 | 0 |
| Juniperus | 0 | 1 | 0 | 0 | 0 | 0 |
| Lonicera | 0 | 1 | 0 | 0 | 0 | 0 |
| Pterocarya | 0 | 1 | 0 | 0 | 0 | 0 |
| Taxus | 0 | 0 | 1 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Náplavová, K.; Beck, T.; Gáper, J.; Pyszko, P.; Gáperová, S. Synanthropic Process Evaluation (with Factors Affecting Propensity to Parasitism) and Host Range within the Genus Ganoderma in Central Europe. Forests 2021, 12, 1437. https://doi.org/10.3390/f12111437
Náplavová K, Beck T, Gáper J, Pyszko P, Gáperová S. Synanthropic Process Evaluation (with Factors Affecting Propensity to Parasitism) and Host Range within the Genus Ganoderma in Central Europe. Forests. 2021; 12(11):1437. https://doi.org/10.3390/f12111437
Chicago/Turabian StyleNáplavová, Kateřina, Terézia Beck, Ján Gáper, Petr Pyszko, and Svetlana Gáperová. 2021. "Synanthropic Process Evaluation (with Factors Affecting Propensity to Parasitism) and Host Range within the Genus Ganoderma in Central Europe" Forests 12, no. 11: 1437. https://doi.org/10.3390/f12111437
APA StyleNáplavová, K., Beck, T., Gáper, J., Pyszko, P., & Gáperová, S. (2021). Synanthropic Process Evaluation (with Factors Affecting Propensity to Parasitism) and Host Range within the Genus Ganoderma in Central Europe. Forests, 12(11), 1437. https://doi.org/10.3390/f12111437