
Austroboletus brunneisquamus (Boletaceae, Boletales), a New Ectomycorrhizal Fungus from a Tropical Rainforest, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. Molecular Procedures
2.3. Dataset Assembly
2.4. Phylogenetic Analyses
3. Results
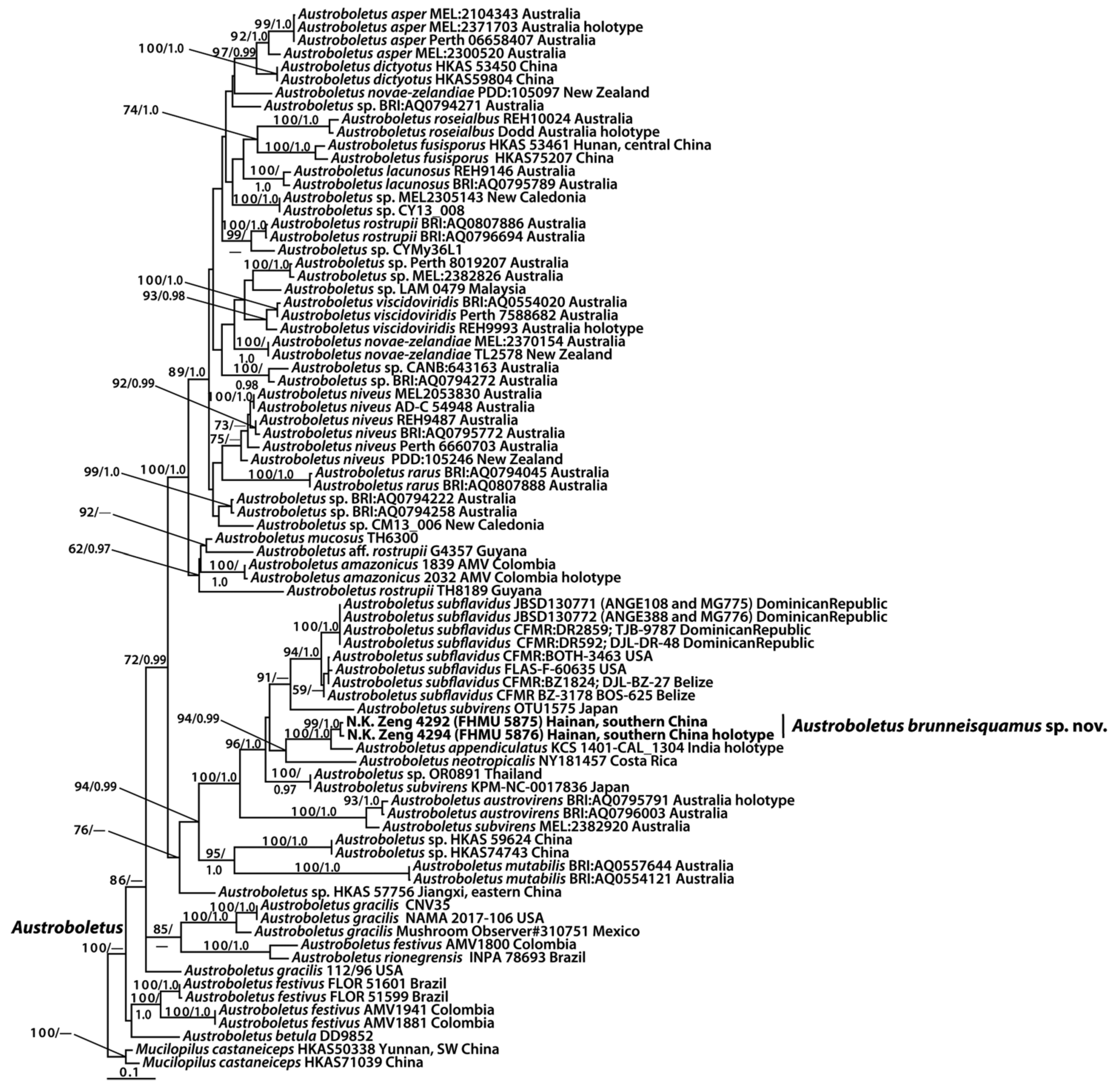
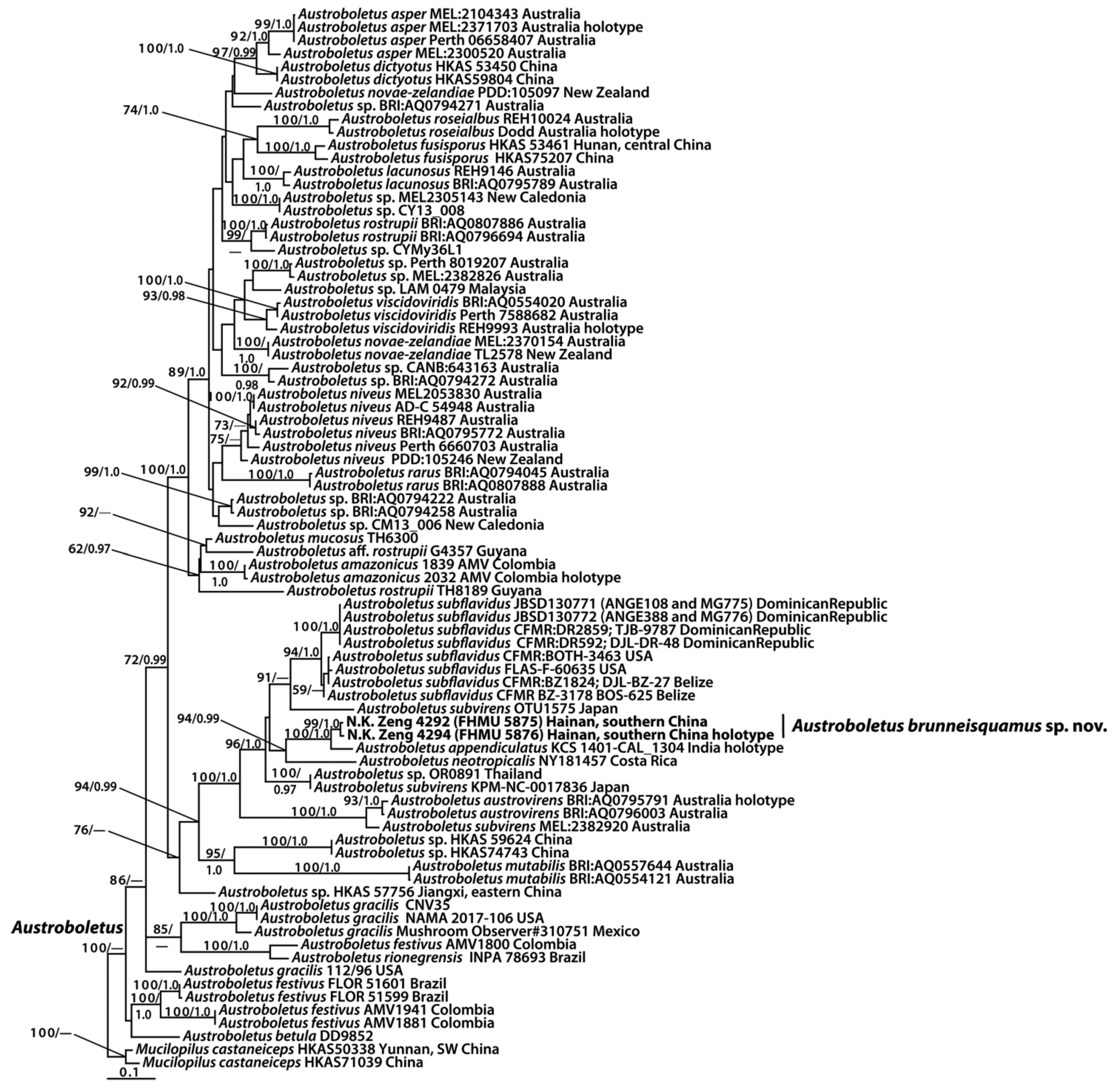
3.1. Molecular Data
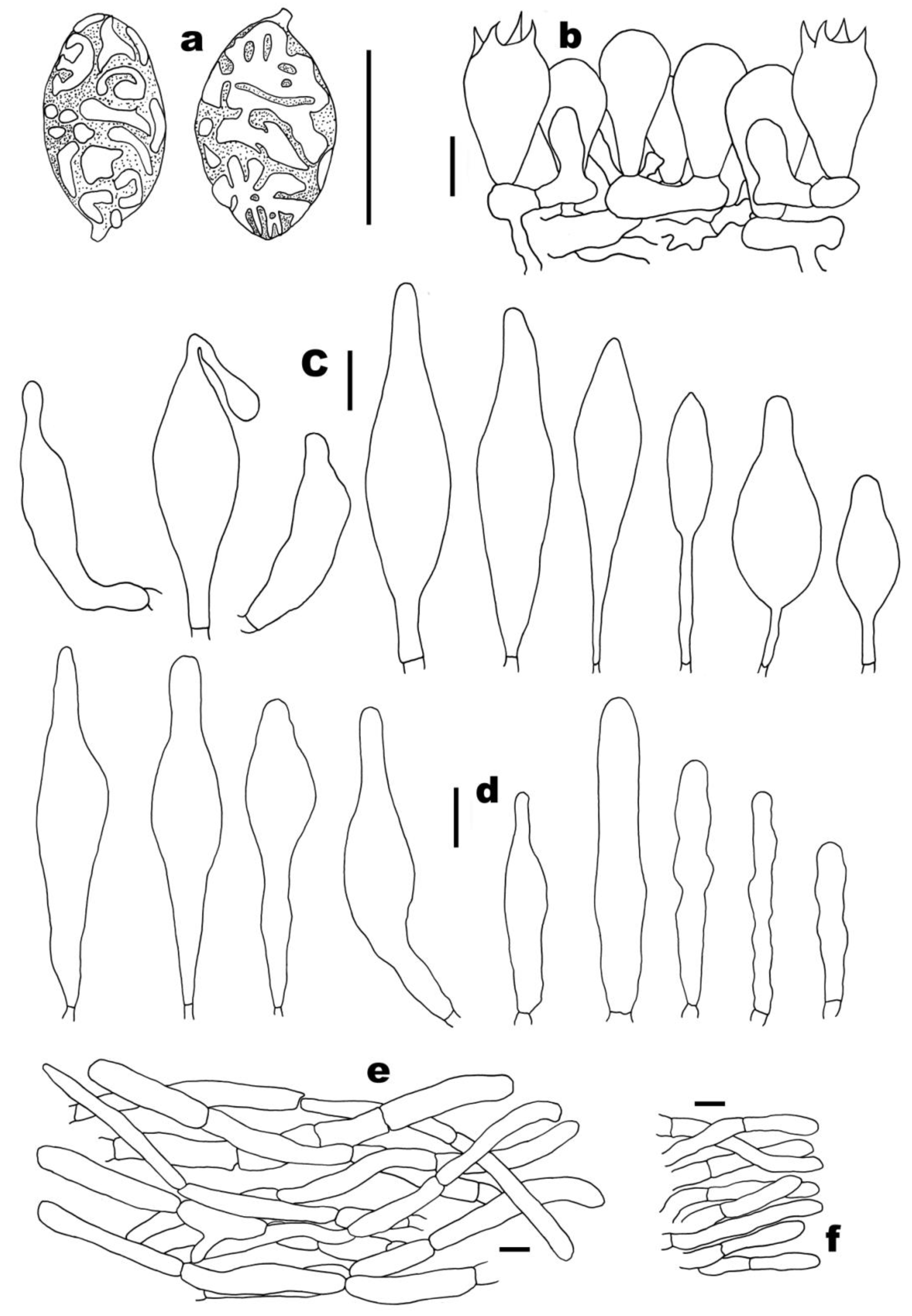
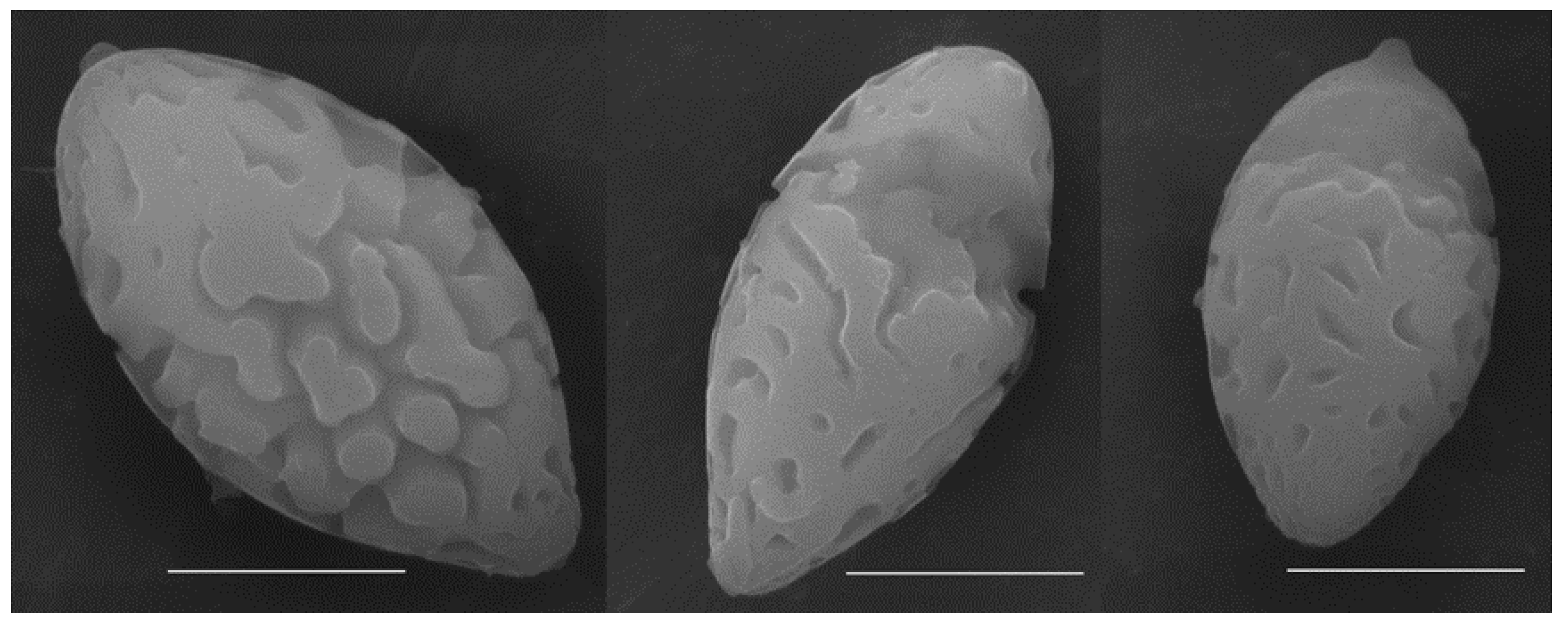
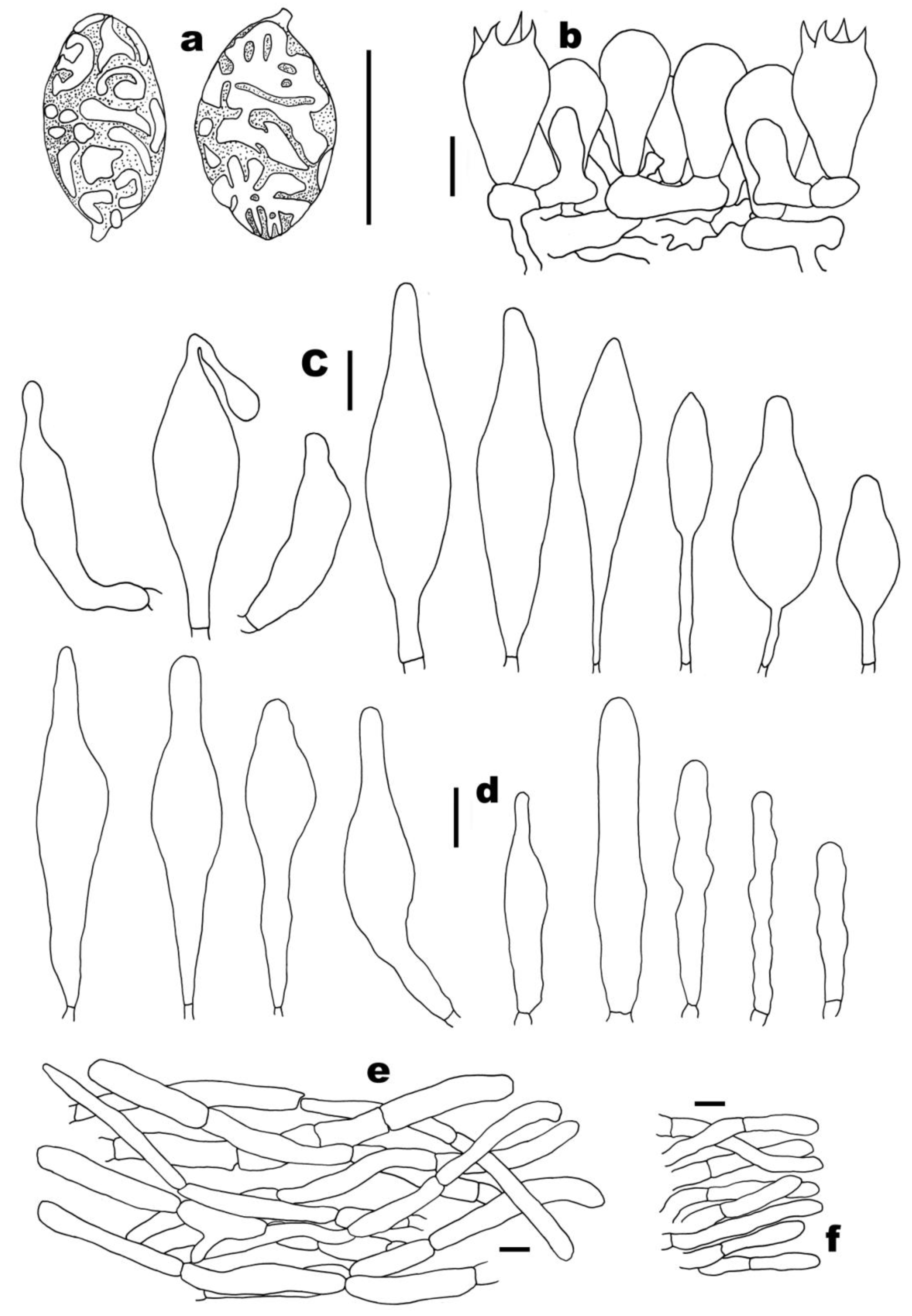
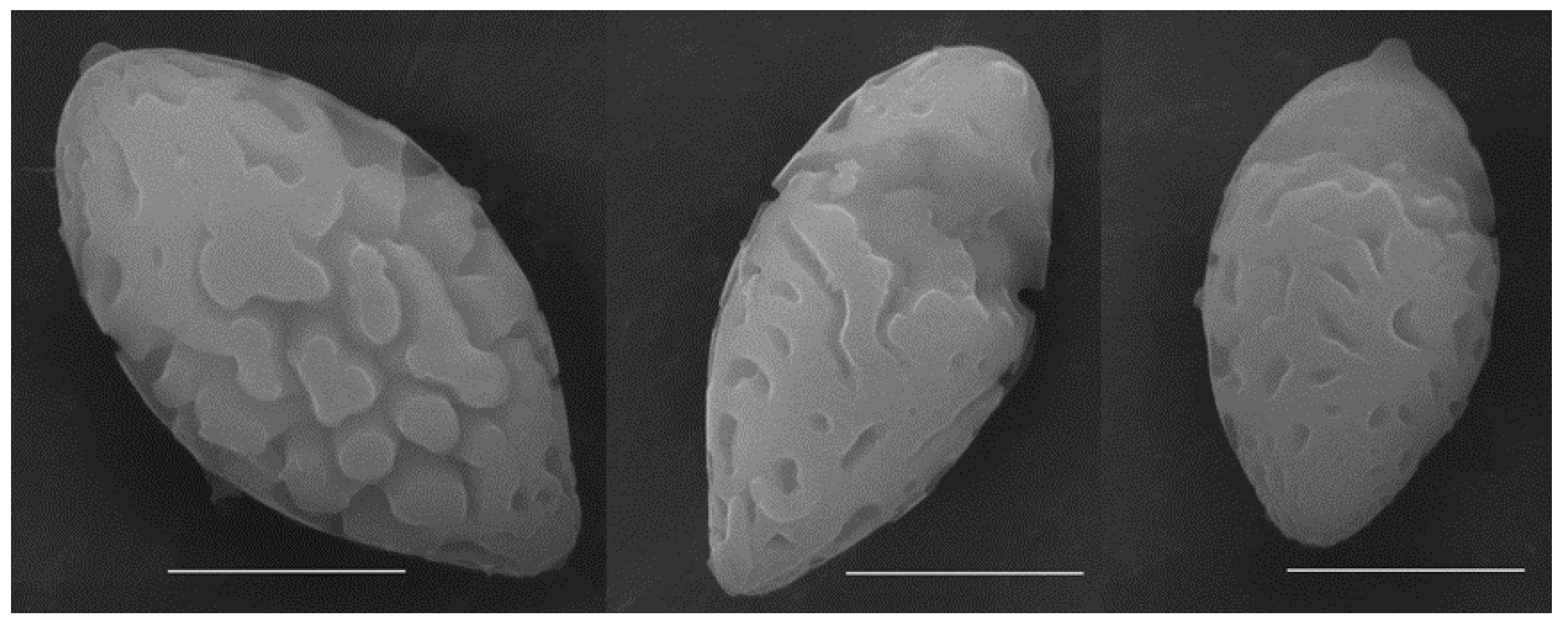
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Corner, E.J.H. Boletus in Malaysia; Government Printing Office: Singapore, 1972; p. 263.
- Wolfe, C.B. Austroboletus and Tylopilus subgenus. Porphyrellus with emphasis on North American taxa. Bibl. Haematol. 1979, 69, 1–148. [Google Scholar]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181 (Austroboletus asper K. Syme, Bonito, T. Lebel, Fechner & Halling, sp. nov.). Persoonia 2020, 45, 251–409. [Google Scholar] [PubMed]
- Gelardi, M.; Angelini, C.; Costanzo, F.; Ercole, E.; Ortiz-Santana, B.; Vizzini, A. Outstanding pinkish brown-spored neotropical boletes: Austroboletus subflavidus and Fistulinella gloeocarpa (Boletaceae, Boletales) from the Dominican Republic. Mycobiology 2020, 49, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Horak, E. Boletellus and Porphyrellus in Papua New Guinea. Kew Bull. 1977, 31, 645–652. [Google Scholar] [CrossRef]
- Fulgenzi, T.D.; Halling, R.E.; Henkel, T.W. Fistulinella cinereoalba sp. nov. and new distribution records for Austroboletus from Guyana. Mycologia 2010, 102, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Vasco-Palacios, A.D.M.; López-Quintero, C.; Franco-Molano, A.E.; Boekhout, T. Austroboletus amazonicus sp. nov. and Fistulinella campinaranae var. scrobiculata, two commonly occurring boletes from a forest dominated by Pseudomonotes tropenbosii (Dipterocarpaceae) in Colombian Amazonia. Mycologia 2014, 106, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Dentinger, B.T.M. Austroboletus olivaceoglutinosus, a new mushroom species from Sikkim, India with a distinctive green, glutinous pileus. Kew Bull. 2015, 70, 15. [Google Scholar] [CrossRef]
- Fechner, N.; Bonito, G.; Bougher, N.L.; Lebel, T.; Halling, R.E. New species of Austroboletus (Boletaceae) in Australia. Mycol. Prog. 2017, 16, 769–775. [Google Scholar] [CrossRef]
- García-Jiménez, J. Austroboletus neotropicalis Singer, J. Garcíay L. D. Gómez (Boletaceae), an interesting fungus associated to montane cloud forest and oak forest in Mexico. Árido-Ciencia 2019, 4, 3–8. [Google Scholar]
- Horak, E. Supplementary remarks to Austroboletus (Corner) Wolfe (Boletaceae). Sydowia 1980, 33, 71–87. [Google Scholar]
- Mao, X.L. The resources of macrofungi from the Mt. Namjagbarwa region in Xizang (Tibet), China. Acta Mycol. Sinica. 1985, 4, 197–207. [Google Scholar]
- Li, T.H.; Song, B. Bolete species known from China. Guizhou Sci. 2003, 21, 78–86. [Google Scholar]
- Hosen, M.I.; Feng, B.; Wu, G.; Zhu, X.T.; Li, Y.C.; Yang, Z.L. Borofutus, a new genus of Boletaceae from tropical Asia: Phylogeny, morphology and taxonomy. Fungal Divers. 2013, 58, 215–226. [Google Scholar] [CrossRef]
- Liang, Z.Q.; An, D.Y.; Jiang, S.; Su, M.S.; Zeng, N.K. Butyriboletus hainanensis (Boletaceae, Boletales), a new species from tropical China. Phytotaxa 2016, 267, 256–262. [Google Scholar] [CrossRef]
- Liang, Z.Q.; Chai, H.; Jiang, S.; Ye, Z.K.; Zeng, N.K. The genus Xanthoconium (Boletaceae, Boletales) in tropical China. Phytotaxa 2017, 295, 246–254. [Google Scholar] [CrossRef]
- An, D.Y.; Liang, Z.Q.; Jiang, S.; Su, M.S.; Zeng, N.K. Cantharellus hainanensis, a new species with a smooth hymenophore from tropical China. Mycoscience 2017, 58, 438–444. [Google Scholar] [CrossRef]
- Zeng, N.K.; Liang, Z.Q.; Tang, L.P.; Li, Y.C.; Yang, Z.L. The genus Pulveroboletus (Boletaceae, Boletales) in China. Mycologia 2017, 109, 422–442. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.K.; Chai, H.; Jiang, S.; Xue, R.; Wang, Y.; Hong, D.; Liang, Z.Q. Retiboletus nigrogriseus and Tengioboletus fujianensis, two new boletes from the south of China. Phytotaxa 2018, 367, 45–54. [Google Scholar] [CrossRef]
- Chai, H.; Liang, Z.Q.; Jiang, S.; Fu, X.L.; Zeng, N.K. Lanmaoa rubriceps, a new bolete from tropical China. Phytotaxa 2018, 347, 71–80. [Google Scholar] [CrossRef]
- Chai, H.; Liang, Z.Q.; Xue, R.; Jiang, S.; Luo, S.H.; Wang, Y.; Wu, L.L.; Tang, L.P.; Chen, Y.; Hong, D.; et al. New and noteworthy boletes from subtropical and tropical China. MycoKeys 2019, 46, 55–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, R.; Wu, L.L.; Jiang, S.; Hao, Y.J.; Chai, H.; Liang, Z.Q.; Zeng, N.K.; Su, M.S. Two new species of the genus Leccinellum (Boletaceae, Boletales) from the south of China. Phytotaxa 2019, 411, 93–104. [Google Scholar] [CrossRef]
- Wang, Y.; Su, M.S.; Jiang, S.; Xue, R.; Wu, L.L.; Xie, H.J.; Zhang, Y.Z.; Liang, Z.Q.; Zeng, N.K. The genus Hourangia in China and a description of Aureoboletus erythraeus sp. nov. Phytotaxa 2020, 472, 87–106. [Google Scholar] [CrossRef]
- Xie, H.J.; Lin, W.F.; Jiang, S.; Xue, R.; Wu, L.L.; Zhang, Y.Z.; Liang, Z.Q.; Zeng, N.K.; Su, M.S. Two new species of Hortiboletus (Boletaceae, Boletales) from China. Mycol. Prog. 2020, 19, 1377–1386. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Taschenlexikon der Farben. 3. Aufl.; Muster-Schmidt Verlag: Göttingen, Germany, 1981; p. 242. [Google Scholar]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2013, 58, 73–101. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.Y.; Kauff, F.; Schoch, C.; Matheny, P.B.; Hofstetter, V.; Cox, C.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of the fungi using a six gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenies. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analyses program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Wu, G.; Feng, B.; Xu, J.P.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Magnago, A.C.; Neves, M.A.; Silveira, R.M.B.D. Fistulinella ruschii sp. nov. and a new record of Fistulinella campinaranae var. scrobiculata for the Atlantic Forest, Brazil. Mycologia 2018, 109, 1003–1013. [Google Scholar]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Halling, R.E.; Nuhn, M.; Osmundson, T.; Fechner, N.; Trappe, J.M.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Affinities of the Boletus chromapes group to Royoungia and the description of two new genera, Harrya and Australopilus. Aust. Syst. Bot. 2012, 25, 418–431. [Google Scholar] [CrossRef]
- Drehmel, D.; James, T.; Vilgalys, R. Molecular phylogeny and biodiversity of the boletes. Fungi 2008, 1, 17–23. [Google Scholar]
- Smith, M.E.; Henkel, T.W.; Aime, M.C.; Fremier, A.K.; Vilgalys, R. Ectomycorrhizal fungal diversity and community structure on three co-occurring leguminous canopy tree species in a neotropical rainforest. New Phytol. 2011, 192, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Vadthanarat, S.; Lumyong, S.; Raspé, O. Cacaoporus, a new Boletaceae genus, with two new species from Thailand. MycoKeys 2019, 54, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Carriconde, F.; Gardes, M.; Bellanger, J.M.; Letellier, K.; Gigante, S.; Gourmelon, V.; Thomas Ibanez, T.; McCoy, S.; Goxe, J.; Read, J.; et al. Host effects in high ectomycorrhizal diversity tropical rainforests on ultramafic soils in New Caledonia. Fungal Ecol. 2019, 39, 201–212. [Google Scholar] [CrossRef]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Orihara, T.; Smith, M.E.; Shimomura, N.; Iwase, K.; Maekawa, N. Diversity and systematics of the sequestrate genus Octaviania in Japan: Two new subgenera and eleven new species. Persoonia 2012, 28, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 184–190. [Google Scholar] [CrossRef]
- Hongo, T. The Agaricales of Japan 1–2. Rhodophyllaceae, Paxillaceae, Gomphidiaceae, Boletaceae and Strobilomycetaceae. Acta Phytotax. Geobot. 1960, 18, 97–112. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher | Locality | GenBank Accession No. | References | ||
|---|---|---|---|---|---|---|
| 28S | ITS | TEF1 | ||||
| Austroboletus aff. fusisporus | HKAS 53461 | Hunan, central China | KF112486 | — | KF112214 | [32] |
| Austroboletus aff. rostrupii | G4357 | Guyana | KJ786636 | — | — | Unpublished |
| Austroboletus amazonicus | 2032 AMV | Colombia | KF714510 | KF937309 | — | [8] |
| A. amazonicus | 1839 AMV | Colombia | KF714508 | KF937307 | — | [8] |
| Austroboletus asper | Perth 06658407 | Australia | KP242277 | KP242216 | — | [4] |
| A.asper | Perth 8018251 | Australia | KP242267 | — | — | [4] |
| A. asper | MEL:2104343 | Australia | KP242260 | KP242174 | — | [4] |
| A. asper | MEL:2300520 | Australia | KP242253 | KP242186 | — | [4] |
| A. asper | MEL:2371703 | Australia | — | KP242152 | — | [4] |
| Austroboletus austrovirens | BRI:AQ0796003 | Australia | KP242228 | KP242212 | — | [10] |
| A. austrovirens | BRI:AQ0795791 | Australia | KP242225 | KP242211 | — | [10] |
| Austroboletus brunneisquamus | N.K. Zeng 4292 (FHMU5875) | Hainan, southern China | MW506828 | MZ855494 | — | This study |
| A. brunneisquamus | N.K. Zeng 4294 (FHMU5876) | Hainan, southern China | MW506829 | MZ855495 | MW512637 | This study |
| Austroboletus dictyotus | HKAS59804 | China | JX901138 | — | — | [15] |
| A. dictyotus | HKAS 53450 | Hunan, central China | KF112487 | — | KF112215 | [32] |
| Austroboletus festivus | FLOR 51599 | Brazil | KY888001 | KY886202 | — | [33] |
| A. festivus | AMV1941 | Colombia | KT724096 | — | — | Unpublished |
| A. festivus | FLOR 51601 | Brazil | KY888000 | KY886203 | — | [33] |
| A. festivus | AMV1881 | Colombia | KT724095 | KT724086 | — | Unpublished |
| A. festivus | AMV1800 | Colombia | — | KT724085 | — | Unpublished |
| A. fusisporus | HKAS75207 | China | JX889720 | JX889719 | JX889718 | [15] |
| Austroboletus gracilis | 112/96 | USA | DQ534624 | — | KF030425 | [34] |
| A. gracilis | CNV35 | — | — | MT345212 | — | Unpublished |
| A. gracilis | NAMA 2017-106 | USA | — | MH979242 | — | Unpublished |
| A. gracilis | Mushroom Observer # 310751 | Mexico | — | MH167935 | — | Unpublished |
| Austroboletus lacunosus | BRI:AQ0795789 | Australia | KP242271 | KP242162 | — | Unpublished |
| A. lacunosus | REH9146 | Australia | JX889669 | — | JX889709 | [35] |
| Austroboletus mucosus | TH6300 | Guyana | AY612798 | — | — | [36] |
| Austroboletus mutabilis | BRI:AQ0554121 | Australia | KP242241 | KP242192 | — | Unpublished |
| A. mutabilis | BRI:AQ0557644 | Australia | KP242237 | KP242196 | — | Unpublished |
| Austroboletus neotropicalis | NY181457 | Costa Rica | JQ924334 | JQ924301 | — | Unpublished |
| Austroboletus niveus | AD-C 54948 | Australia | KP242280 | KP242220 | — | Unpublished |
| A. niveus | Perth 6660703 | Australia | KP242279 | KP242217 | — | Unpublished |
| A. niveus | BRI:AQ0795772 | Australia | KP242276 | KP242156 | — | Unpublished |
| A. niveus | MEL2053830 | Australia | KC552058 | KC552016 | KC552099 | Unpublished |
| A. niveus | REH9487 | Australia | JX889668 | — | JX889708 | [35] |
| A. niveus | PDD:105246 | New Zealand | — | — | MH594049 | Unpublished |
| Austroboletus novae-zelandiae | TL2578 | New Zealand | — | KP191803 | — | Unpublished |
| A. novae-zelandiae | PDD:105097 | New Zealand | — | — | MH594051 | Unpublished |
| A. novae-zelandiae | MEL:2370154 | Australia | KP242256 | KP242175 | — | Unpublished |
| Austroboletus rarus | BRI:AQ0794045 | Australia | KP242236 | KP242197 | — | Unpublished |
| A. rarus | BRI:AQ0807888 | Australia | — | KP242200 | — | Unpublished |
| Austroboletus rionegrensis | INPA 78693 | Brazil | — | KY886201 | — | [33] |
| Austroboletus roseialbus | Dodd | Australia | KY872650 | KY872653 | — | [10] |
| A. roseialbus | REH10024 | Australia | KY872651 | KY872652 | — | [10] |
| Austroboletus rostrupii | BRI:AQ0807886 | Australia | KP242270 | KP242163 | — | Unpublished |
| A. rostrupii | BRI:AQ0796694 | Australia | KP242258 | KP242179 | — | Unpublished |
| A. rostrupii | TH8189 | Guyana | — | JN168683 | — | [37] |
| Austroboletus sp. | HKAS74743 | China | KT990527 | — | KT990730 | [1] |
| Austroboletus sp. | MEL:2382826 | Australia | KP242283 | KP242213 | — | Unpublished |
| Austroboletus sp. | BRI:AQ0794271 | Australia | KP242259 | — | — | Unpublished |
| Austroboletus sp. | BRI:AQ0794258 | Australia | KP242255 | KP242182 | — | Unpublished |
| Austroboletus sp. | Perth 8019207 | Australia | KP242245 | — | — | Unpublished |
| Austroboletus sp. | BRI:AQ0794222 | Australia | KP242234 | KP242215 | — | Unpublished |
| Austroboletus sp. | MEL2305143 | New Caledonia | KC552060 | KC552018 | KC552101 | Unpublished |
| Austroboletus sp. | CANB:643163 | Australia | — | KP242201 | — | Unpublished |
| Austroboletus sp. | BRI:AQ0794272 | Australia | — | KP242159 | — | Unpublished |
| Austroboletus sp. | HKAS 59624 | Yunnan, SW China | KF112485 | — | KF112217 | [32] |
| Austroboletus sp. | HKAS 57756 | Jiangxi, eastern China | KF112383 | — | KF112212 | [32] |
| Austroboletus sp. | OR0891 | Thailand | — | — | MH614706 | [38] |
| Austroboletus sp. | CM13_006 | New Caledonia | — | KY774009 | — | [39] |
| Austroboletus sp. | CY13_008 | — | — | KY774008 | — | Unpublished |
| Austroboletus sp. | CYMy36L1 | — | — | KY774007 | — | Unpublished |
| Austroboletus sp. | LAM 0479 | Malaysia | KY091070 | — | — | Unpublished |
| Austroboletus sp. | DD9852 | North America | AY612797 | — | — | Unpublished |
| Austroboletus subflavidus | CFMR BZ-3178 BOS-625 | Belize | MK601716 | — | MK721070 | Unpublished; [40] |
| A. subflavidus | FLAS-F-60635 | USA | — | MH016816 | — | Unpublished |
| A. subflavidus | JBSD130771 | Dominican Republic | MT580902 | MT581525 | — | [5] |
| A. subflavidus | JBSD130772 | Dominican Republic | MT580903 | MT581526 | — | [5] |
| A. subflavidus | CFMR:DR2859 | Dominican Republic | MT580901 | MT581523 | — | [5] |
| A. subflavidus | CFMR:DR592 | Dominican Republic | — | MT581524 | — | [5] |
| A. subflavidus | CFMR:BZ1824 | Belize | — | MT581522 | — | [5] |
| A. subflavidus | CFMR:BOTH-3463 | Florida (USA) | MT580900 | MT581521 | — | [5] |
| Austroboletus subvirens | OTU1575 | Japan | — | MT594990 | — | Unpublished |
| A. subvirens | KPM-NC-0017836 | Japan | JN378518 | — | JN378458 | [41] |
| A. subvirens | MEL:2382920 | Australia | — | KP012789 | — | Unpublished |
| Austroboletus viscidoviridis | REH9993 | Australia | — | KY872649 | — | [10] |
| A. viscidoviridis | Perth 7588682 | Australia | KP242282 | KP242219 | — | Unpublished |
| A. viscidoviridis | BRI:AQ0554020 | Australia | KP242243 | KP242189 | — | Unpublished |
| Mucilopilus castaneiceps | HKAS50338 | China | KT990555 | — | KT990755 | [1] |
| M. castaneiceps | HKAS71039 | China | KT990547 | — | KT990748 | [1] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Liang, Z.-Q.; Su, M.-S.; Jiang, S.; Chen, Y.; Fan, Y.-G.; Zeng, N.-K. Austroboletus brunneisquamus (Boletaceae, Boletales), a New Ectomycorrhizal Fungus from a Tropical Rainforest, China. Forests 2021, 12, 1438. https://doi.org/10.3390/f12111438
Xu C, Liang Z-Q, Su M-S, Jiang S, Chen Y, Fan Y-G, Zeng N-K. Austroboletus brunneisquamus (Boletaceae, Boletales), a New Ectomycorrhizal Fungus from a Tropical Rainforest, China. Forests. 2021; 12(11):1438. https://doi.org/10.3390/f12111438
Chicago/Turabian StyleXu, Chang, Zhi-Qun Liang, Ming-Sheng Su, Shuai Jiang, Yun Chen, Yu-Guang Fan, and Nian-Kai Zeng. 2021. "Austroboletus brunneisquamus (Boletaceae, Boletales), a New Ectomycorrhizal Fungus from a Tropical Rainforest, China" Forests 12, no. 11: 1438. https://doi.org/10.3390/f12111438
APA StyleXu, C., Liang, Z.-Q., Su, M.-S., Jiang, S., Chen, Y., Fan, Y.-G., & Zeng, N.-K. (2021). Austroboletus brunneisquamus (Boletaceae, Boletales), a New Ectomycorrhizal Fungus from a Tropical Rainforest, China. Forests, 12(11), 1438. https://doi.org/10.3390/f12111438