Short Legacy Effects of Growing Season Nitrogen Addition and Reduced Precipitation alter Soil Respiration during Nongrowing Season


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Soil Respiration Measurement
2.4. Statistical Analyses
3. Results
3.1. Abiotic Factors
3.2. Soil Respiration and Its Components
3.3. Relationships of Soil Respiration with Environmental Factors
4. Discussion
4.1. Effects of N Addition on Nongrowing Season Soil Respiration and Its Components
4.2. Effects of Reduced Precipitation on Nongrowing Season Soil Respiration and Its Components
4.3. Combined Effects of N Addition and Reduced Precipitation on Nongrowing Season Soil Respiration and Its Components
4.4. Adjustment of Soil Respiration and Its Components by N Addition and Reduced Precipitation
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Brooks, P.; Mcknight, D.; Elder, K. Carbon limitation of soil respiration under non-growing season snowpacks: Potential feedbacks between growing season and non-growing season carbon fluxes. Glob. Chang. Biol. 2004, 11, 231–238. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhou, X.; Zhang, B.; Lu, M.; Luo, Y.; Liu, L.; Li, B. Different responses of soil respiration and its components to nitrogen addition among biomes: A meta-analysis. Glob. Chang. Biol. 2014, 20, 2332–2343. [Google Scholar] [CrossRef]
- Feng, J.; Wang, J.; Ding, L.; Yao, P.; Qiao, M.; Yao, S. Meta-analyses of the effects of major global change drivers on soil respiration across China. Atmos. Environ. 2017, 150, 181–186. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, X.; Shao, J.; Nie, Y.; He, Y.; Jiang, L.; Wu, Z.; Bai, S. Interactive effects of global change factors on soil respiration and its components: A meta-analysis. Glob. Chang. Biol. 2016, 22, 3157–3169. [Google Scholar] [CrossRef]
- Yan, G.; Chen, F.; Zhang, X.; Wang, J.; Han, S.; Xing, Y.; Wang, Q. Spatial and temporal effects of nitrogen addition on root morphology and growth in a boreal forest. Geoderma 2017, 303, 178–187. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Sun, T.; Chen, L.; Pang, X.; Wang, Y.; Xiao, F. N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Chen, F.; Yan, G.; Xing, Y.; Zhang, J.; Wang, Q.; Wang, H.; Liu, T. Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest. Agric. For. Meteorol. 2019, 271, 336–345. [Google Scholar] [CrossRef]
- Yan, L.; Chen, S.; Huang, J.; Lin, G. Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Glob. Chang. Biol. 2010, 16, 2345–2357. [Google Scholar] [CrossRef]
- Liu, L.; Wang, X.; Lajeunesse, M.J.; Miao, G.; Piao, S.; Wan, S.; Deng, M. A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes. Glob. Chang. Biol. 2016, 22, 1394–1405. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, T.X.; Zhang, J.; Li, X.W.; Hu, H.L.; Liu, L.; Xiao, Y.L. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biol. Biochem. 2013, 58, 255–264. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Z.; Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2014, 15, 184–195. [Google Scholar] [CrossRef]
- Zhang, C.; Niu, D.; Hall, S.J.; Wen, H.; Li, X.; Fu, H.; Wan, C.; Elser, J.J. Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol. Biochem. 2014, 75, 113–123. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Chen, S.; Tan, X.; Wang, S.; Chen, M.; Ren, T.; Xia, J.; Huang, J.; Han, X. Changing precipitation exerts greater influence on soil heterotrophic than autotrophic respiration in a semiarid steppe. Agric. Forest Meteorol. 2019, 217, 413–421. [Google Scholar] [CrossRef]
- Contosta, A.R.; Burakowski, E.A.; Varner, R.K.; Frey, S.D. Non-growing season soil respiration in a humid temperate forest: The roles of moisture, temperature, and snowpack. J. Geophys. Res. 2016, 121, 3072–3088. [Google Scholar] [CrossRef]
- Contosta, A.R.; Frey, S.D.; Cooper, A.B. Seasonal dynamics of soil respiration and N mineralization in chronically warmed and fertilized soils. Ecosphere 2011, 2, 1–21. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change 2013-The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; 1552p. [Google Scholar]
- Ravn, N.R.; Elberling, B.; Michelsen., A. Arctic soil carbon turnover controlled by experimental snow addition, summer warming and shrub removal. Soil Biol. Biochem. 2020, 142, 107698. [Google Scholar] [CrossRef]
- Zheng, J.; Guo, R.; Li, D.; Zhang, J.; Han, S. Nitrogen addition, drought and mixture effects on litter decomposition and nitrogen immobilization in a temperate forest. Plant Soil 2017, 416, 165–179. [Google Scholar] [CrossRef]
- Yan, G.; Dong, X.; Huang, B.; Wang, H.; Hong, Z.; Zhang, J.; Wang, Q. Effects of nitrogen deposition on litter decomposition and nutrient release mediated by litter types and seasonal change in a temperate forest. Can. J. Soil Sci. 2020, 100, 11–25. [Google Scholar] [CrossRef]
- Muhr, J.; Borken, W.; Matzner, E. Effects of soil frost on soil respiration and its radiocarbon signature in a Norway spruce forest soil. Glob. Chang. Biol. 2009, 15, 782–793. [Google Scholar] [CrossRef]
- Zhou, M.; Yan, G.; Xing, Y.; Chen, F.; Zhang, X.; Wang, J.; Wang, Q. Nitrogen deposition and decreased precipitation does not change total nitrogen uptake in a temperate forest. Sci. Total Environ. 2019, 651, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Zhou, M.; Wang, M.; Han, S.; Liu, G.; Zhang, X.; Wang, Q. Nitrogen deposition and decreased precipitation altered nutrient foraging strategies of three temperate trees by affecting root and mycorrhizal traits. Catena 2019, 181, 104094. [Google Scholar] [CrossRef]
- Lü, C.; Tian, H. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef]
- Yan, G.; Xing, Y.; Lü, X.T.; Xu, L.; Zhang, J.; Dai, G.; Wang, Q. Effects of artificial nitrogen addition and reduction in precipitation on soil CO2 and CH4 effluxes and composition of the microbial biomass in a temperate forest. Eur. J. Soil Sci. 2019, 70, 1197–1211. [Google Scholar]
- Liu, B.; Mou, C.; Yan, G.; Xu, L.; Jiang, S.; Xing, Y.; Wang, Q. Annual soil CO2 efflux in a cold temperate forest in northeastern China: Effects of non-growing season snowpack and artificial nitrogen deposition. Sci. Rep. 2016, 6, 1–9. [Google Scholar]
- Zhang, T.; Wang, G.; Yang, Y.; Mao, T.; Chen, X. Non-growing season soil CO2 flux and its contribution to annual soil CO2 emissions in two typical grasslands in the permafrost region of the Qinghai-Tibet Plateau. Eur. J. Soil Biol. 2015, 71, 45–52. [Google Scholar] [CrossRef]
- Gaumont-Guay, D.; Black, T.A.; Griffis, T.J.; Barr, A.G.; Jassal, R.S.; Nesic, Z. Interpreting the dependence of soil respiration on soil temperature and water content in a boreal aspen stand. Agric. For. Meteorol. 2006, 140, 220–235. [Google Scholar] [CrossRef]
- Chen, W.; Wolf, B.; Zheng, X.; Yao, Z.; Butterbach-Bahl, K.; Brüggemann, N.; Han, X. Carbon dioxide emission from temperate semiarid steppe during the non-growing season. Atmos. Environ. 2013, 64, 141–149. [Google Scholar] [CrossRef]
- Merbold, L.; Steinlin, C.; Hagedorn, F. Non-growing season greenhouse gas fluxes (CO2, CH4 and N2O) from a subalpine grassland. Biogeosciences 2013, 10, 3185–3203. [Google Scholar] [CrossRef]
- Eberwein, J.R.; Oikawa, P.Y.; Allsman, L.A.; Jenerette, G.D. Carbon availability regulates soil respiration response to nitrogen and temperature. Soil Biol. Biochem. 2015, 88, 158–164. [Google Scholar] [CrossRef]
- Zhang, X.; Xing, Y.; Wang, Q.; Yan, G.; Wang, M.; Liu, G.; Zhang, J. Effects of long-term nitrogen addition and decreased precipitation on the fine root morphology and anatomy of the main tree species in a temperate forest. For. Ecol. Manag. 2020, 455, 117664. [Google Scholar] [CrossRef]
- Noguchi, K.; Nagakura, J.; Konôpka, B.; Sakata, T.; Kaneko, S.; Takahashi, M. Fine-root dynamics in sugi (Cryptomeria japonica) under manipulated soil nitrogen conditions. Plant Soil 2013, 364, 159–169. [Google Scholar] [CrossRef]
- Thomas, R.Q.; Canham, C.D.; Weathers, K.C.; Goodale, C.L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 2010, 3, 13–17. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Rivkina, E.M.; Friedmann, E.I.; McKay, C.P.; Gilichinsky, D.A. Metabolic activity of permafrost bacteria below the freezing point. Appl. Environ. Microbiol. 2000, 66, 3230–3233. [Google Scholar] [CrossRef]
- Steinweg, J.M.; Fisk, M.C.; McAlexander, B.; Groffman, P.M.; Hardy, J.P. Experimental snowpack reduction alters organic matter and net N mineralization potential of soil macroaggregates in a northern hardwood forest. Biol. Fertil. Soils 2008, 45, 1–10. [Google Scholar] [CrossRef]
- ÖQUIST, M.G.; Sparrman, T.; Klemedtsson, L.; Drotz, S.H.; Grip, H.; Schleucher, J.; Nilsson, M. Water availability controls microbial temperature responses in frozen soil CO2 production. Glob. Chang. Biol. 2009, 15, 2715–2722. [Google Scholar] [CrossRef]
- Greaver, T.L.; Clark, C.M.; Compton, J.E.; Vallano, D.; Talhelm, A.F.; Weaver, C.P.; Felker-Quinn, E. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836. [Google Scholar] [CrossRef]
- Hartmann, A.A.; Buchmann, N.; Niklaus, P.A. A study of soil methane sink regulation in two grasslands exposed to drought and N fertilization. Plant Soil 2011, 342, 265–275. [Google Scholar] [CrossRef]
- Jonasson, S.; Michelsen, A.; Schmidt, I.K.; Nielsen, E.V. Responses in microbes and plants to changed temperature, nutrient, and light regimes in the arctic. Ecology 1999, 80, 1828–1843. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Z.; Li, X.; Sun, Y.; Zhang, X.; Liang, A. N fertilization effects on soil respiration, microbial biomass and root respiration in Larix gmelinii and Fraxinus mandshurica plantations in China. Plant Soil 2010, 333, 325–336. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, Y.; Zhang, B.; Li, A.; Daryanto, S.; Wang, L.; Huang, J. The impacts of precipitation increase and nitrogen addition on soil respiration in a semiarid temperate steppe. Ecosphere 2017, 8, e01655. [Google Scholar] [CrossRef]
- Shi, W.Y.; Zhang, J.G.; Yan, M.J.; Yamanaka, N.; Du, S. Seasonal and diurnal dynamics of soil respiration fluxes in two typical forests on the semiarid Loess Plateau of China: Temperature sensitivities of autotrophs and heterotrophs and analyses of integrated driving factors. Soil Biol. Biochem. 2012, 52, 99–107. [Google Scholar] [CrossRef]
- Aanderud, Z.T.; Jones, S.E.; Schoolmaster, D.R., Jr.; Fierer, N.; Lennon, J.T. Sensitivity of soil respiration and microbial communities to altered snowfall. Soil Biol. Biochem. 2013, 57, 217–227. [Google Scholar] [CrossRef]
- Mahecha, M.D.; Reichstein, M.; Carvalhais, N.; Lasslop, G.; Lange, H.; Seneviratne, S.I.; Vargas, R.; Ammann, C.; Arain, M.A.; Cescatti, A.; et al. Global convergence in the temperature sensitivity of respiration at ecosystem level. Science 2010, 329, 838–840. [Google Scholar] [CrossRef] [PubMed]






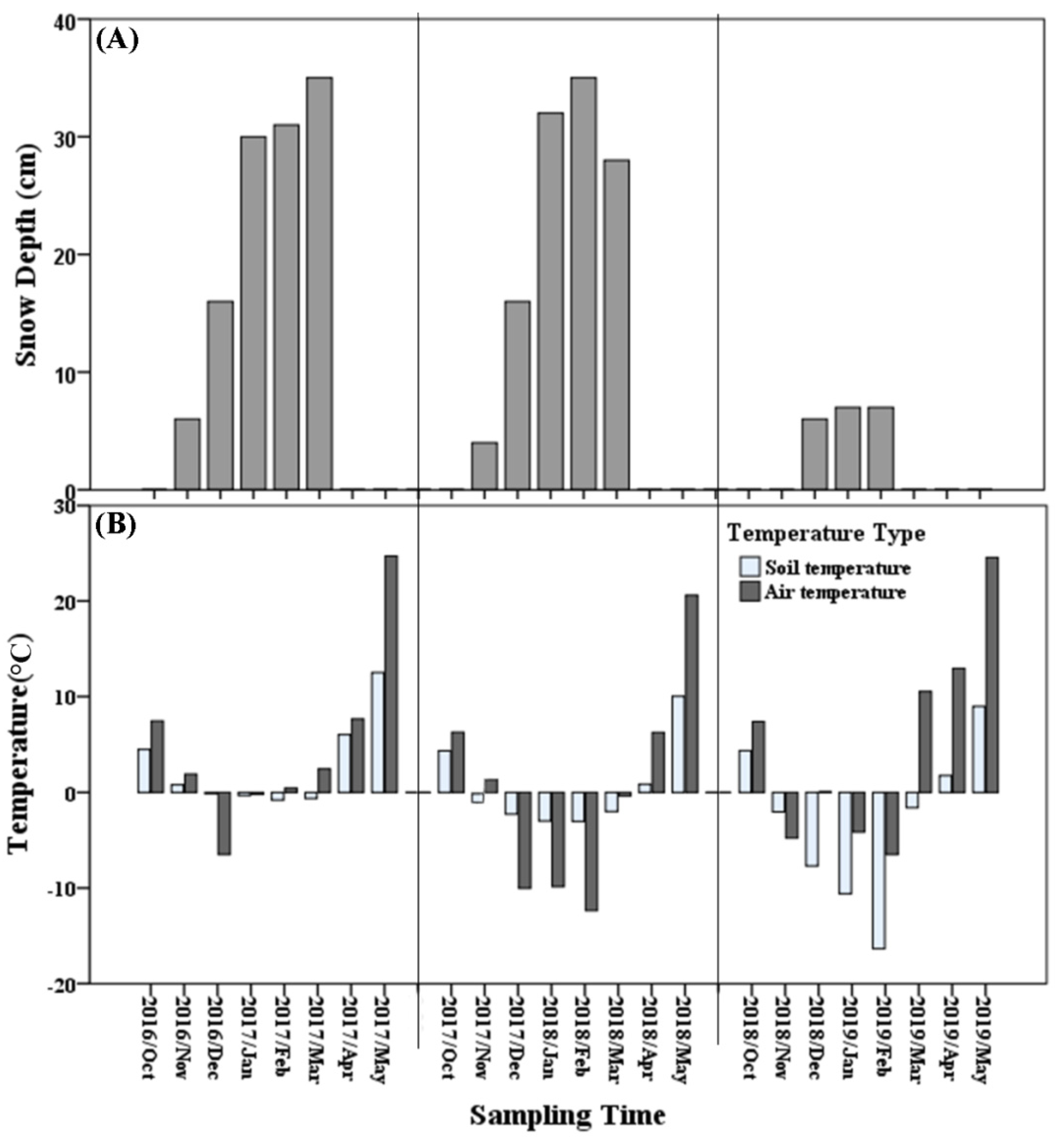{kind=link}
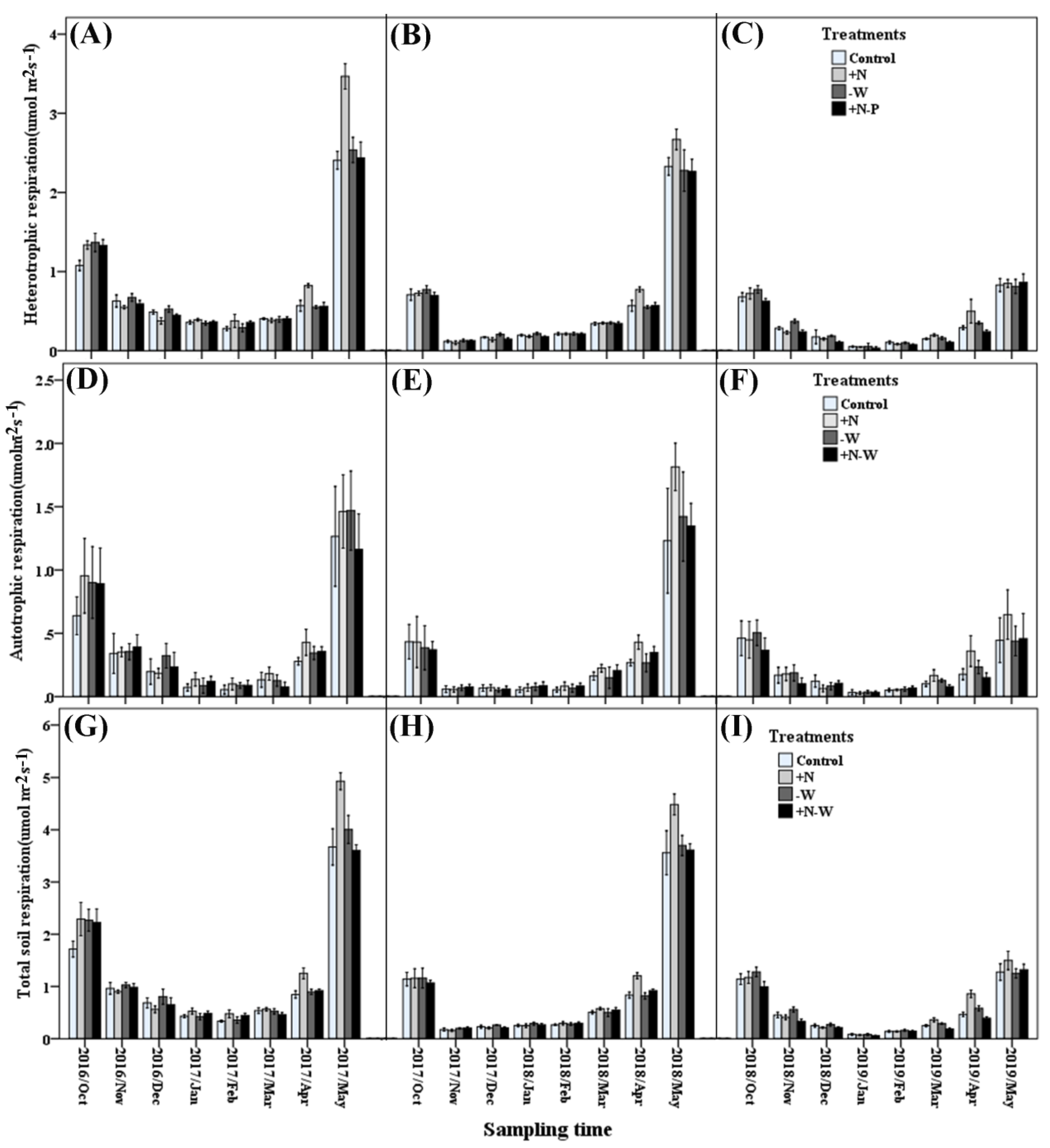{kind=link}
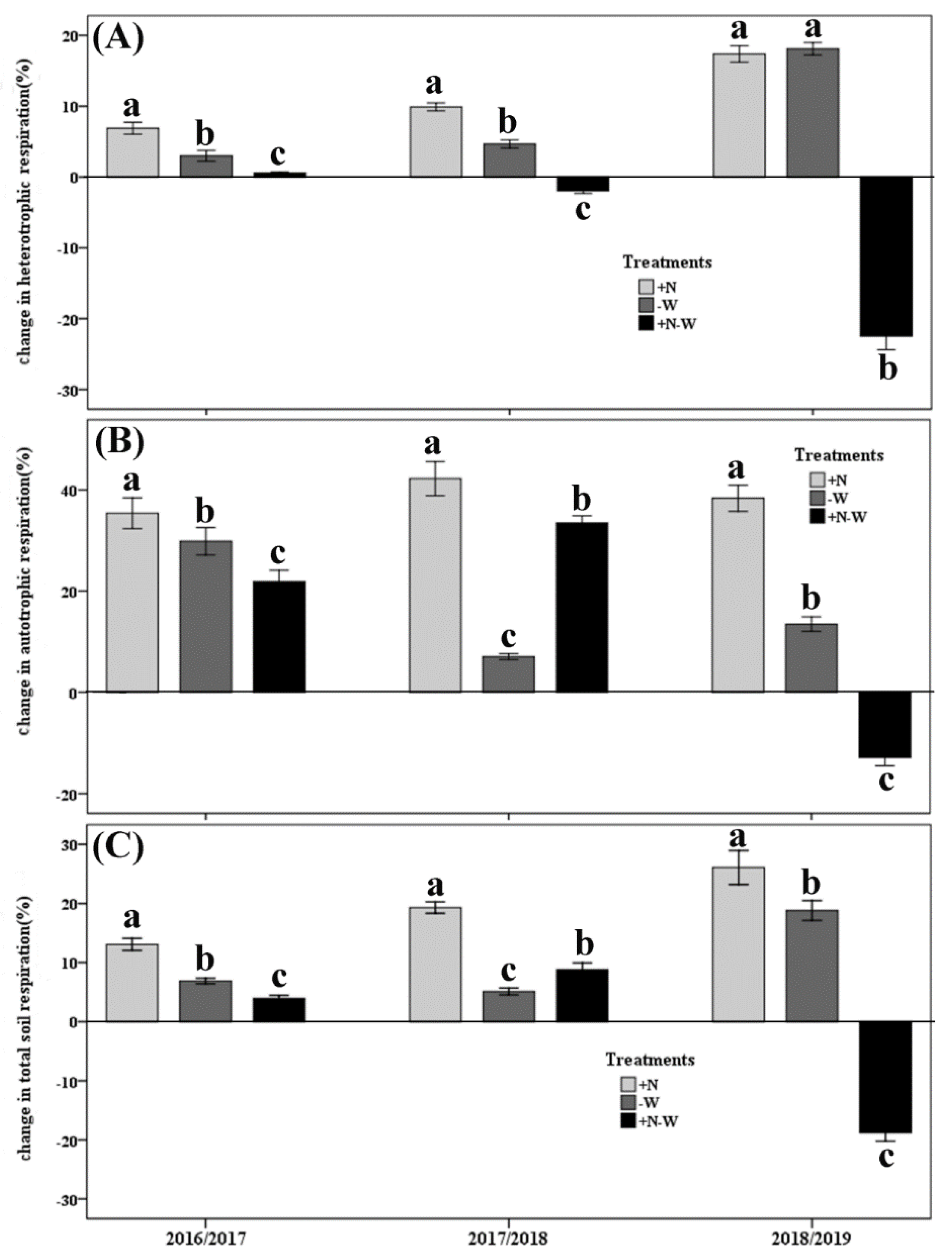{kind=link}
{kind=link}
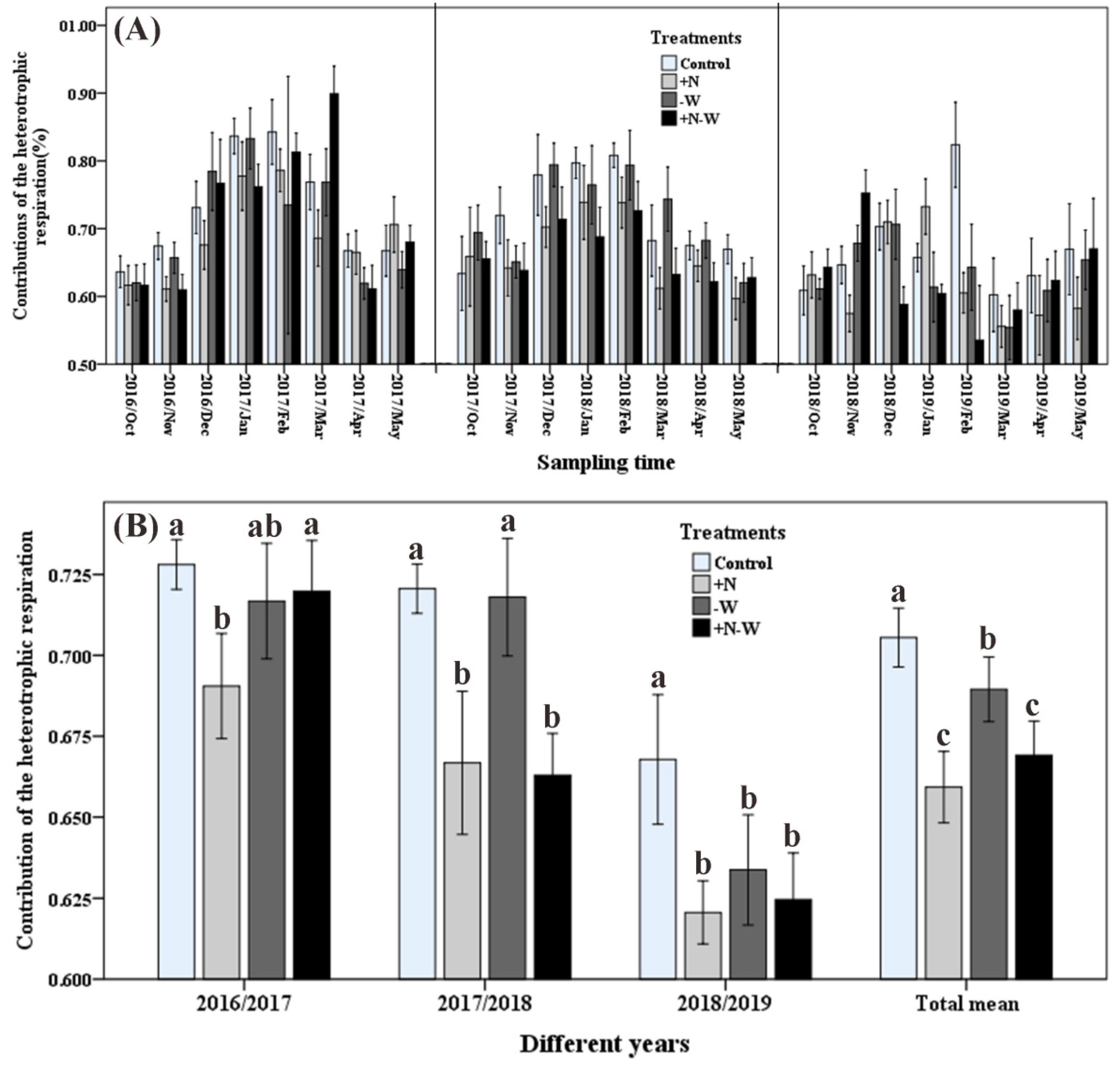{kind=link}
| Treatment or Sampling Times | p Value | |||
|---|---|---|---|---|
| df | Rs | Ra | Rh | |
| +N | 1 | <0.001 | 0.008 | <0.001 |
| -W | 1 | <0.001 | 0.201 | <0.001 |
| Sampling times | 23 | <0.001 | <0.001 | <0.001 |
| +N*-W | 1 | <0.001 | <0.001 | <0.001 |
| +N*sampling times | 23 | <0.001 | 0.067 | <0.001 |
| -W*sampling times | 23 | <0.001 | 0.848 | <0.001 |
| +N*-W*sampling times | 23 | <0.001 | 0.002 | <0.001 |
| Treatments | Soil Temperature | Air Temperature | Soil Moisture | |||
|---|---|---|---|---|---|---|
| R2 | p Value | R2 | p Value | R2 | p Value | |
| Control | 0.701 | <0.001 | 0.625 | <0.001 | 0.542 | <0.001 |
| +N | 0.731 | <0.001 | 0.659 | <0.001 | 0.612 | <0.001 |
| -W | 0.774 | <0.001 | 0.569 | <0.001 | 0.647 | <0.001 |
| +N-W | 0.561 | <0.001 | 0.551 | <0.001 | 0.605 | <0.001 |
| Models | a | b | c | R2 | p | Q10 |
|---|---|---|---|---|---|---|
| Rs = aebtMc | ||||||
| Control | 2.35 ± 0.09 *** | 0.14 ± 0.01 ** | 0.70 ± 0.03 *** | 0.80 | <0.001 | 4.07 ± 0.10a |
| +N | 1.97 ± 0.12 *** | 0.13 ± 0.01 * | 0.66 ± 0.04 *** | 0.76 | <0.001 | 3.71 ± 0.21b |
| -W | 2.12 ± 0.14 ** | 0.13 ± 0.00 ** | 0.68 ± 0.08 *** | 0.78 | <0.001 | 3.94 ± 0.19ab |
| +N-W | 1.88 ± 0.10 * | 0.13 ± 0.01 ** | 0.58 ± 0.04 | 0.64 | <0.001 | 3.70 ± 0.12b |
| Rh = aebtMc | ||||||
| Control | 1.78 ± 0.14 *** | 0.15 ± 0.01* | 0.56 ± 0.06 *** | 0.75 | <0.001 | 4.55 ± 0.09a |
| +N | 1.72 ± 0.09 *** | 0.14 ± 0.02 | 0.45 ± 0.08 *** | 0.62 | <0.001 | 4.12 ± 0.28b |
| -W | 1.69 ± 0.18 *** | 0.14 ± 0.01 * | 0.52 ± 0.10 *** | 0.59 | <0.001 | 4.15 ± 0.14b |
| +N-W | 1.63 ± 0.21 *** | 0.14 ± 0.01 * | 0.45 ± 0.04 *** | 0.48 | <0.001 | 3.87± 0.19b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, G.; Xing, Y.; Wang, Q.; Mu, C. Short Legacy Effects of Growing Season Nitrogen Addition and Reduced Precipitation alter Soil Respiration during Nongrowing Season. Forests 2020, 11, 358. https://doi.org/10.3390/f11030358
Yan G, Xing Y, Wang Q, Mu C. Short Legacy Effects of Growing Season Nitrogen Addition and Reduced Precipitation alter Soil Respiration during Nongrowing Season. Forests. 2020; 11(3):358. https://doi.org/10.3390/f11030358
Chicago/Turabian StyleYan, Guoyong, Yajuan Xing, Qinggui Wang, and Changcheng Mu. 2020. "Short Legacy Effects of Growing Season Nitrogen Addition and Reduced Precipitation alter Soil Respiration during Nongrowing Season" Forests 11, no. 3: 358. https://doi.org/10.3390/f11030358
APA StyleYan, G., Xing, Y., Wang, Q., & Mu, C. (2020). Short Legacy Effects of Growing Season Nitrogen Addition and Reduced Precipitation alter Soil Respiration during Nongrowing Season. Forests, 11(3), 358. https://doi.org/10.3390/f11030358