Effect of Bark-Stripping on Mechanical Stability of Norway Spruce

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
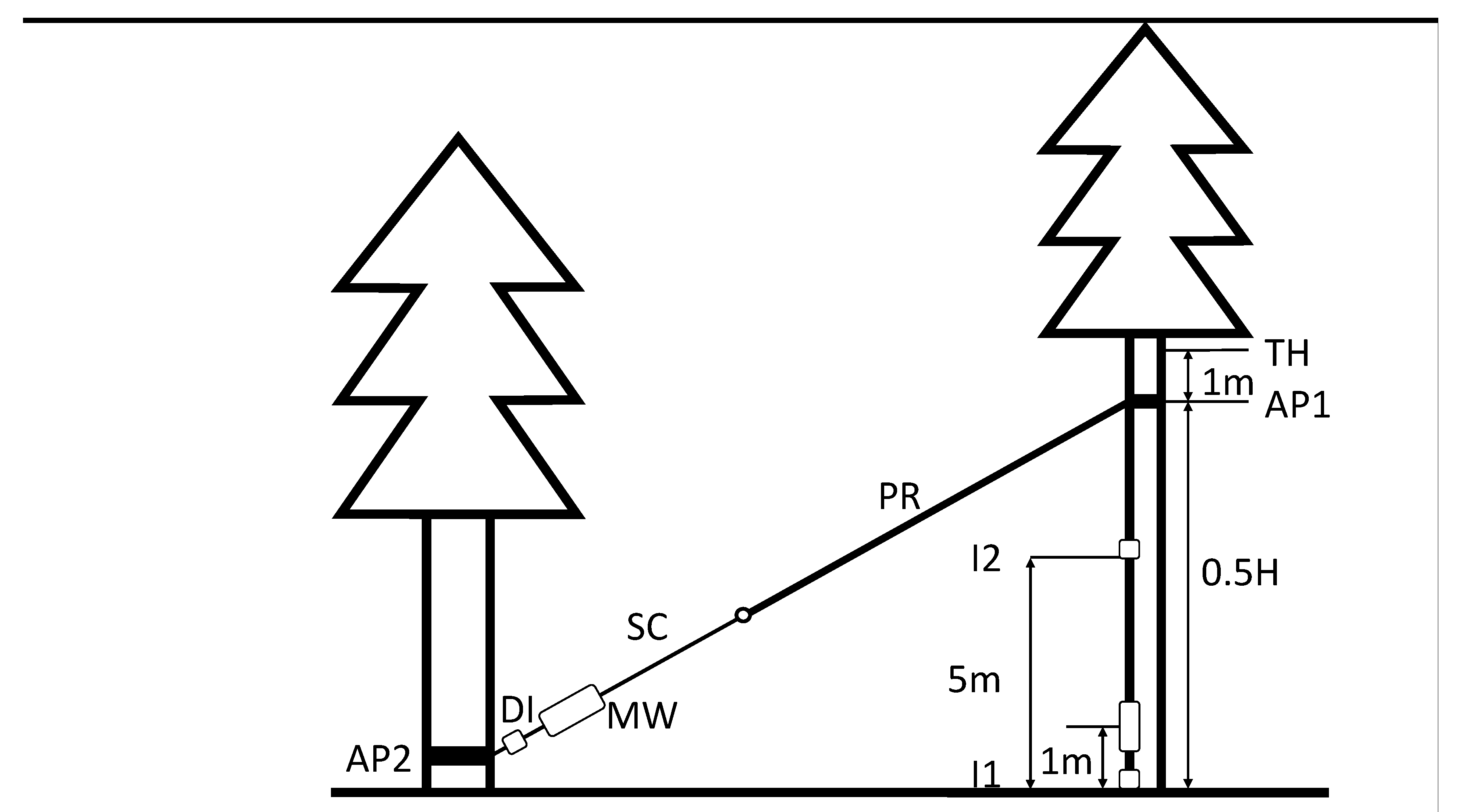
2.1. Study Site and Sample Trees
2.2. Static Pulling Tests
2.3. Data Processing and Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, B.; Schuck, A.R.T.; Schelhaas, M.J.; Orazio, C.; Blennow, K.; Nicoll, B. Living with Storm Damage to Forests; European Forest Institute: Joensuu, Finland, 2013; pp. 1–132. [Google Scholar]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Peltola, H.; Ikonen, V.P.; Gregow, H.; Strandman, H.; Kilpeläinen, A.; Venäläinen, A.; Kellomäki, S. Impacts of climate change on timber production and regional risks of wind-induced damage to forests in Finland. For. Ecol. Manag. 2010, 260, 833–845. [Google Scholar] [CrossRef]
- Albrecht, A.; Hanewinkel, M.; Bauhus, J.; Kohnle, U. How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur. J. For. Res. 2012, 131, 229–247. [Google Scholar] [CrossRef]
- Richter, C. Wood Characteristics: Description, Causes, Prevention, Impact on Use and Technological Adaptation; Springer Internationale Publishing: Basel, Switzerland, 2015; p. 222. [Google Scholar]
- Lodin, I. Choice of Tree Species in the Aftermath of Two Major Storms—A Qualitative Study of Private Forest Owners in Southern Sweden. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2016. [Google Scholar]
- Kurek, T.; Jacek, T.; Pazdrowski, W.; Szymański, M.; Łukowski, A. Intensity of stripping and sugar content in the bark and the bast of European beech (Fagus sylvatica). Open Life Sci. 2019, 14, 19–28. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley & Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. European Ungulates and Their Management in the 21st Century, 1st ed.; Cambridge University Press: Cambridge, UK, 2010; p. 618. [Google Scholar]
- Månsson, J.; Jarnemo, A. Bark-stripping on Norway spruce by red deer in Sweden: Level of damage and relation to tree characteristics. Scan. J. For. Res. 2013, 28, 117–125. [Google Scholar] [CrossRef]
- Verheyden, H.; Ballon, P.; Bernard, V.; Saint-Andrieux, C. Variations in bark-stripping by red deer Cervus elaphus across Europe. Mamm. Rev. 2006, 36, 217–234. [Google Scholar] [CrossRef]
- Vospernik, S. Probability of bark stripping damage by red deer (Cervus elaphus) in Austria. Silva Fenn. 2006, 40, 589–601. [Google Scholar] [CrossRef][Green Version]
- Randveer, T.; Heikkilä, R. Damage cause by moose (Alces alces L.) by bark stripping of Picea abies. Scan. J. For. Res. 1996, 11, 153–158. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Damage to trees due to forestry operations and its pathological significance in temperate forests: A literature review. Forestry 2001, 74, 319–336. [Google Scholar] [CrossRef]
- Burneviča, N.; Jansons, Ā.; Zaļuma, A.; Kļaviņa, D.; Jansons, J.; Gaitnieks, T. Fungi Inhabiting Bark Stripping Wounds Made by Large Game on Stems of Picea abies (L.) Karst. in Latvia. Balt. For. 2016, 22, 2–7. [Google Scholar]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Cukor, J.; Vacek, Z.; Linda, R.; Vacek, S.; Marada, P.; Šimůnek, V.; Havránek, F. Effects of bark stripping on timber production and structure of Norway spruce forests in relation to climatic factors. Forests 2019, 10, 320. [Google Scholar] [CrossRef]
- McLaughlin, J.; Šica, L. Aļņu radītie mizas plēsumi parastai eglei un ar tiem saistītās trupes izraisītāju infekcijas. Mežzinātne 1996, 39, 26–37. [Google Scholar]
- Vlad, R.; Sidor, C. Assessment of the wood volume with stem decay in Norway spruce stands damaged by red deer. In Proceedings of the Biennial International Symposium Forest and Sustainable Development, Brasov, Romania, 24–25 October 2014; Transylvania University Press: Brasov, Romania, 2015; pp. 381–386. [Google Scholar]
- Harris, I.; Jones, P.; Osborn, T.; Lister, D. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef]
- Peltola, H.M. Mechanical stability of trees under static loads. Am. J. Bot. 2006, 93, 1501–1511. [Google Scholar] [CrossRef]
- Detter, A.; Richter, K.; Rust, C.; Rust, S. Aktuelle Untersuchungen zum Primärversagen von grünem Holz—Current studies on primary failure in green wood. In Proceedings of the Conference Deutsche Baumpflegetage, Augsburg, Germany, 5–7 May 2015; pp. 156–167. [Google Scholar]
- Liepa, I. Pieauguma Mācība; [Increment Theory]; Latvia University of Agriculture: Jelgava, Latvia, 1996; p. 123. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 24 November 2019).
- Wagener, W.W. Judging Hazard from Native Trees in California Recreational Areas: A Guide for Professional Foresters; USFS Research Paper PSW-P1; CFS: Berkeley, CA, USA, 1963; p. 29. [Google Scholar]
- Shibata, E.; Torazawa, Y. Effects of bark stripping by sika deer, Cervus nippon, on wind damage to coniferous trees in subalpine forest of central Japan. J. For. Res. 2008, 13, 296–301. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Lehtonen, M.; Väisänen, H.; Peltola, H. Effects of wood decay by Heterobasidion annosum on the vulnerability of Norway spruce stands to wind damage: A mechanistic modelling approach. Can. J. For. Res. 2017, 47, 777–787. [Google Scholar] [CrossRef]
- Lundström, T.; Jonas, T.; Stöckli, S.; Ammann, W. Anchorage of mature conifers: Resistive turning moment, root–soil plate geometry and root growth orientation. Tree Physiol. 2007, 27, 1217–1227. [Google Scholar] [CrossRef]
- Jillich, S.; Köhler, J.; Rust, C.; Rust, S.; Detter, A. Brechen schlanke Fichten leichter? In Jahrbuch der Baumpflege; Dujesiefken, D., Ed.; Haymarket Media: Braunschweig, Germany, 2013; pp. 267–273. [Google Scholar]
- Szoradova, A.; Praus, L.; Kolarik, J. Evaluation of the root system resistance against failure of urban trees using principal component analysis. Biosyst. Eng. 2013, 115, 244–249. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Five basidiomycetes in living stems of Picea abies, associated with 7–25 year-old wounds. Balt. For. 1998, 1, 29–35. [Google Scholar]
- Deflorio, G.; Johnson, C.R.; Fink, S.; Schwarze, F.W.M.R. Decay development in living sapwood of coniferous and deciduous trees inoculated with six wood decay fungi. For. Ecol. Manag. 2008, 255, 2373–2383. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi; Springer: Berlin/Heidelberg, Germany, 2006; p. 334. [Google Scholar]
- Herrmann, S.; Huber, M.O.; Bont, Z.; Rigling, A.; Wunder, J. Decay detection in Norway spruce (Picea abies) with the Rotfinder instrument. For. Ecol. Manag. 2019, 448, 549–558. [Google Scholar] [CrossRef]
- Schelhaas, M.J. Impacts of natural disturbances on the development of European forest resources: Application of model approaches from tree and stand levels to large-scale scenarios. Alterra Sci. Contrib. 2008, 23, 168. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Undamaged | Bark-Stripped | |
|---|---|---|
| Number of trees | 11 | 8 |
| Stem circumference (cm) | 86.0 ± 13.7 | 82.2 ± 17.3 |
| Damage circumference (cm) | - | 22.5 ± 6.2 |
| Damage proportion (%) | - | 28.1 ± 7.3 |
| DBH (cm) | 27.3 ± 4.3 | 25.8 ± 5.1 |
| Tree height (m) | 24.3 ± 2.6 | 23.1 ± 3.3 |
| Stem volume (m3) | 0.76 ± 0.24 | 0.64 ± 0.27 |
| Pulling direction (cosine) | 0.33 ± 0.46 | 0.83 ± 0.23 |
| Parameter | F-Value | p-Value |
|---|---|---|
| Primary failure | ||
| Stem volume | 268.9 | <0.001 |
| Presence of bark-stripping wound | 159.3 | <0.001 |
| Stem volume by presence of bark-stripping interaction | 28.9 | <0.001 |
| R2 | 0.96 | |
| Model overall significance, p-value | <0.001 | |
| Secondary failure | ||
| Stem volume | 294.5 | <0.001 |
| Presence of bark-stripping wound | 38.0 | <0.001 |
| Stem volume by presence of bark-stripping interaction | 0.6441 | 0.43 |
| R2 | 0.95 | |
| Model overall significance, p-value | <0.001 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krisans, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests 2020, 11, 357. https://doi.org/10.3390/f11030357
Krisans O, Saleniece R, Rust S, Elferts D, Kapostins R, Jansons A, Matisons R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests. 2020; 11(3):357. https://doi.org/10.3390/f11030357
Chicago/Turabian StyleKrisans, Oskars, Renate Saleniece, Steffen Rust, Didzis Elferts, Rolands Kapostins, Aris Jansons, and Roberts Matisons. 2020. "Effect of Bark-Stripping on Mechanical Stability of Norway Spruce" Forests 11, no. 3: 357. https://doi.org/10.3390/f11030357
APA StyleKrisans, O., Saleniece, R., Rust, S., Elferts, D., Kapostins, R., Jansons, A., & Matisons, R. (2020). Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests, 11(3), 357. https://doi.org/10.3390/f11030357