The Persisting Influence of Edge on Vegetation in Hemiboreal Alnus Glutinosa (L.) Gaertn. Swamp Forest Set-Asides Adjacent to Recently Disturbed Stands

 ,
,  , and
, and
Abstract
1. Introduction
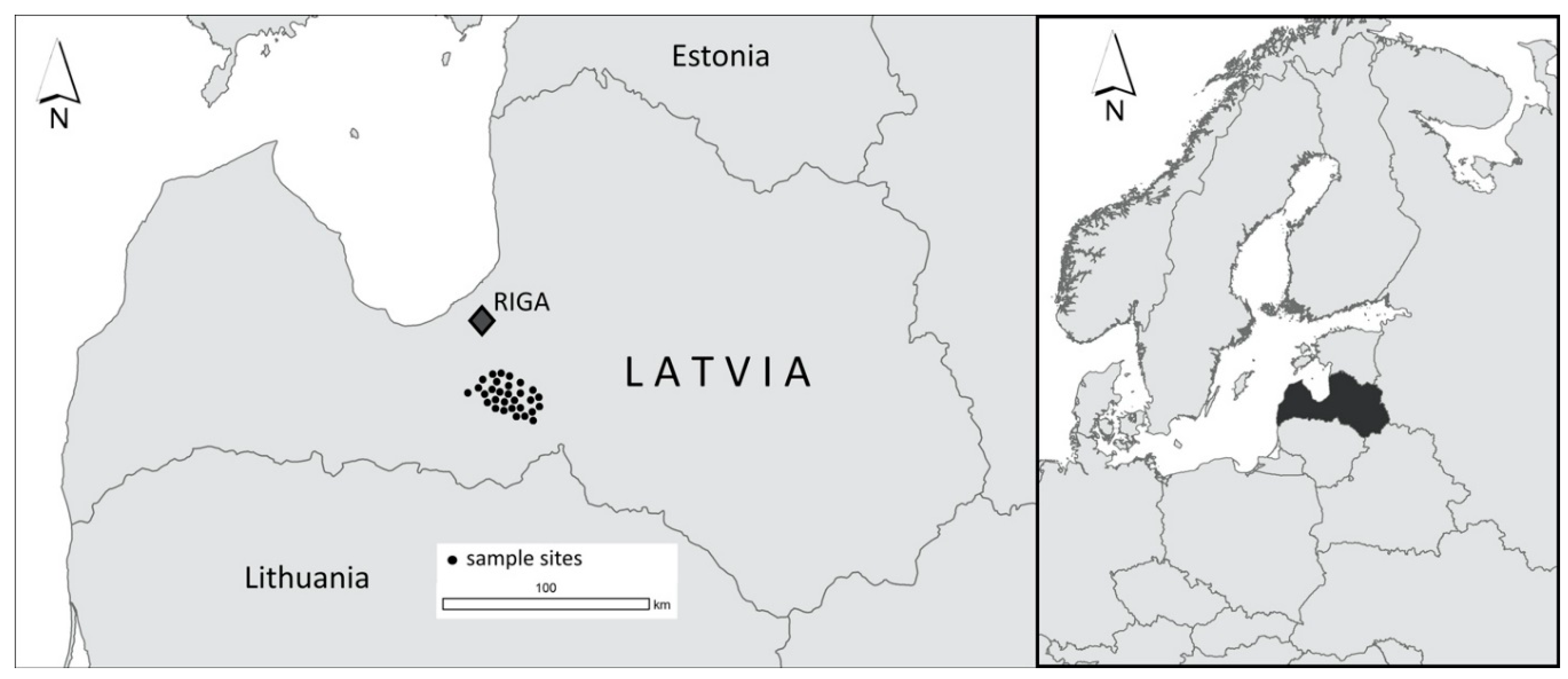
2. Materials and Methods
3. Results
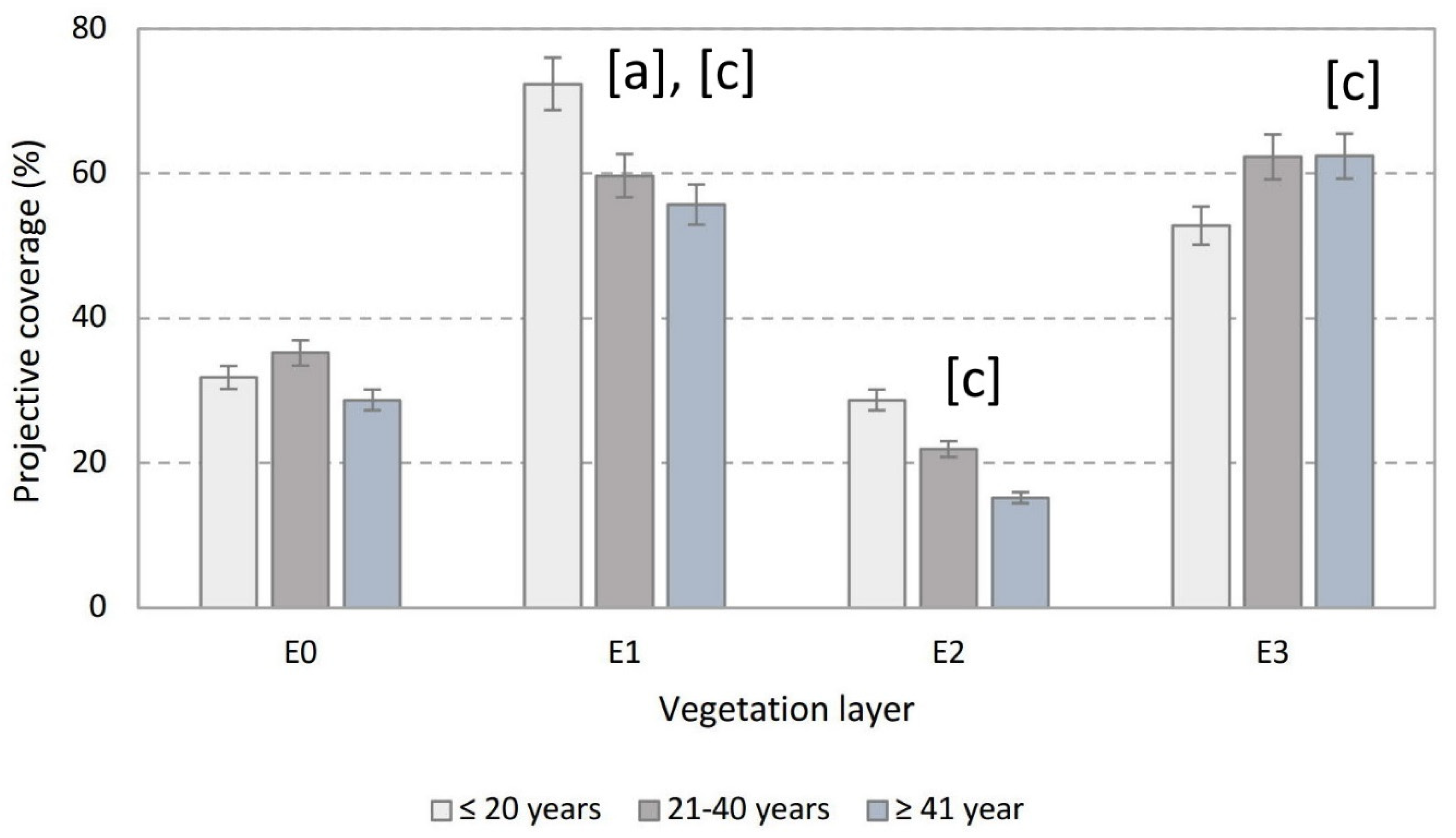
3.1. Projective Coverage
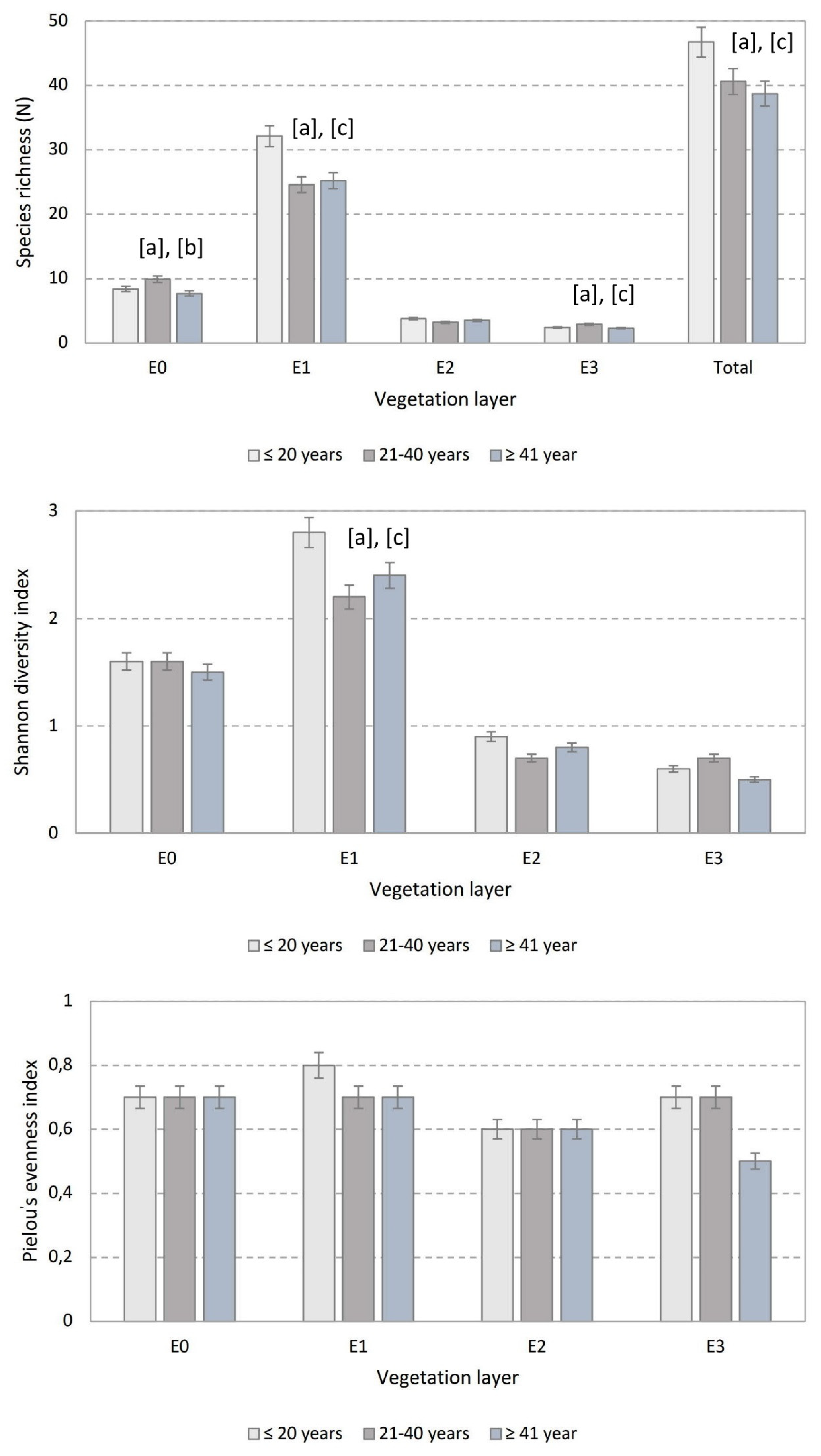
3.2. Species Richness, Shannon-Wiener Index and Evenness
3.3. Plant Functional Types
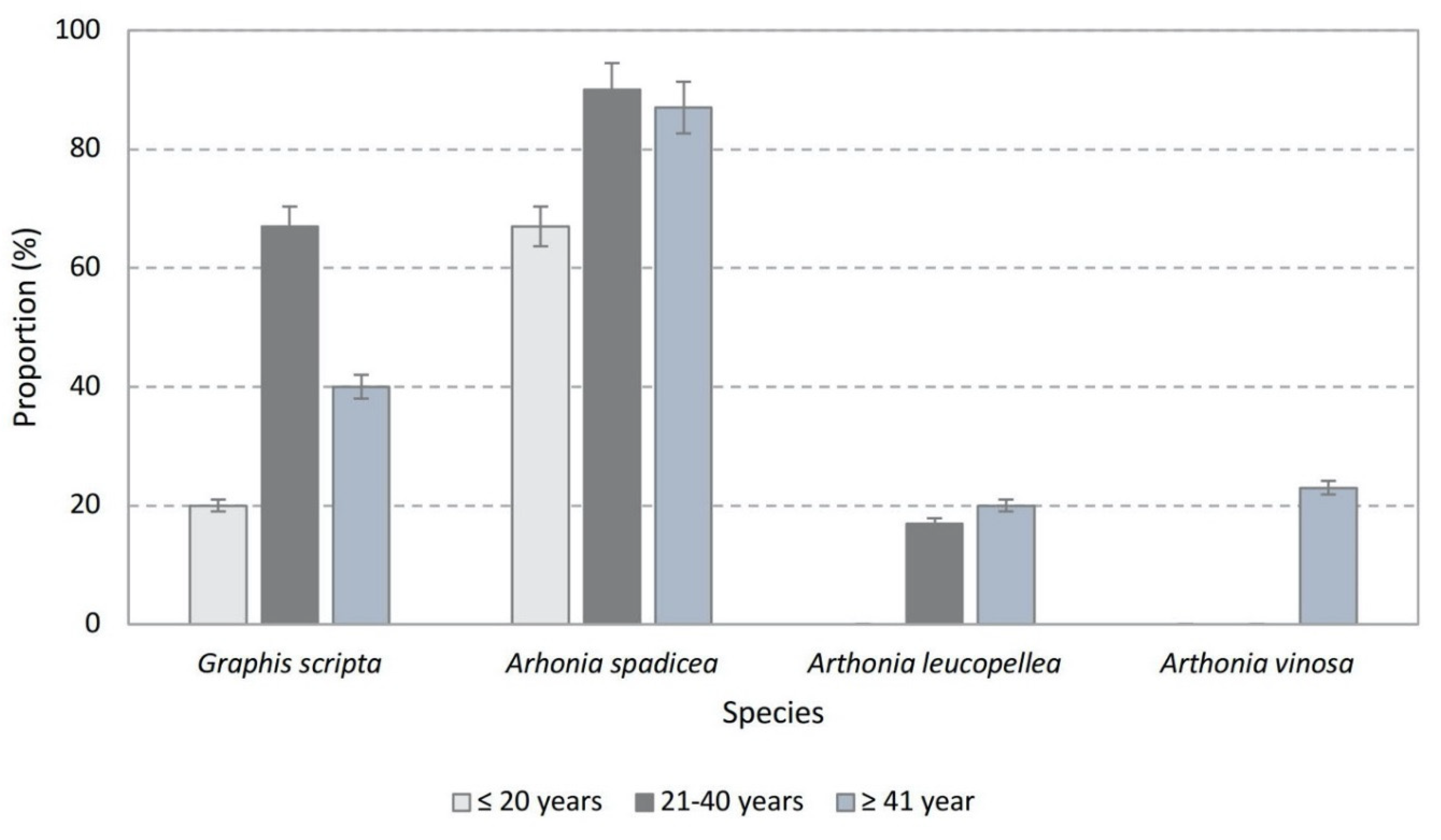
3.4. Proportion of Protected Epiphytic Lichen Species and WKH Indicator Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adjacent Stand Age Group | Mean Age | Range |
|---|---|---|
| ≤20 years (n = 10) | 7.3 | 1–20 |
| 21–40 years (n = 10) | 31.0 | 21–40 |
| ≥41 years (n = 10) | 63.3 | 41–87 |
| Variable | Species Group | Age Group | ||
|---|---|---|---|---|
| ≤20 Years | 21–40 Years | ≥41 Years | ||
| Habitat group | Boreal species | 24.06 ± 1.20 | 35.42 ± 1.77 | 26.88 ± 1.34 |
| Nemoral species | 15.73 ± 0.79 | 15.81 ± 0.79 | 14.85 ± 0.74 | |
| Nitrophylus species | 38.08 ± 1.90 | 31.57 ± 1.58 | 38.05 ± 1.90 | |
| Meadow species | 2.81 ± 0.14 | 1.50 ± 0.07 | 2.88 ± 0.14 | |
| Swamp forest species | 17.50 ± 0.88 | 14.77 ± 0.74 | 15.28 ± 0.76 | |
| Adventive species | 1.82 ± 0.09 | 0.93 ± 0.05 | 2.06 ± 0.10 | |
| Seed dispersal agent | Animals | 12.65 ± 0.63 | 10.37 ± 0.52 | 14.07 ± 0.70 |
| Ants | 7.77 ± 0.39 | 8.28 ± 0.41 | 7.42 ± 0.37 | |
| Autochorous | 9.89 ± 0.49 | 8.44 ± 0.42 | 1.55 ± 0.58 | |
| Birds | 32.94 ± 1.65 | 37.45 ± 1.87 | 29.28 ± 1.46 | |
| Water | 20.10 ± 1.00 | 20.05 ± 1.00 | 20.42 ± 1.02 | |
| Wind | 16.66 ± 0.83 | 15.42 ± 0.77 | 17.27 ± 0.86 | |
| Plant strategy group | C | 29.05 ± 1.45 | 29.43 ± 1.47 | 27.17 ± 1.36 |
| CR | 7.08 ± 0.35 | 6.94 ± 0.35 | 6.13 ± 0.31 | |
| CS | 35.52 ± 1.78 | 37.48 ± 1.87 | 40.32 ± 2.02 | |
| CSR | 18.15 ± 0.91 | 14.61 ± 0.73 | 16.51 ± 0.83 | |
| R | 0.56 ± 0.03 | 0.35 ± 0.02 | 0.51 ± 0.03 | |
| S | 8.74 ± 0.44 | 9.63 ± 0.48 | 8.36 ± 0.42 | |
| SR | 0.90 ± 0.04 | 1.56 ± 0.08 | 1.01 ± 0.05 | |
| Raunkiaer life history form | Chamaephyte | 9.32 ± 0.47 | 11.54 ± 0.58 | 10.86 ± 0.54 |
| Geophyte | 10.51 ± 0.53 | 11.97 ± 0.60 | 12.19 ± 0.61 | |
| Helophyte | 16.55 ± 0.83 | 17.71 ± 0.89 | 18.01 ± 0.90 | |
| Hemicryptophyte | 40.90 ± 2.05 | 30.26 ± 1.51 | 36.40 ± 1.82 | |
| Hydrophyte | 0.95 ± 0.05 | 0.76 ± 0.04 | 0.79 ± 0.04 | |
| Phanerophyte | 17.43 ± 0.87 | 22.80 ± 1.14 | 17.00 ± 0.85 | |
| Therophyte | 4.34 ± 0.22 | 4.95 ± 0.25 | 4.76 ± 0.24 | |
References
- Ries, L.; Fletcher, R.J., Jr.; Battin, J.; Sisk, T.D. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 491–522. [Google Scholar] [CrossRef]
- Harper, K.A.; Lesieur, D.; Bergeron, Y.; Drapeau, P. Forest structure and composition at young fire and cut edges in black spruce boreal forest. Can. J. For. Res. 2004, 34, 289–302. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.A.R.; Jaiteh, M.S.; Esseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Hanson, J.J.; Stuart, J.D. Vegetation responses to natural and salvage logged fire edges in Douglas-fir/hardwood forests. For. Ecol. Manag. 2005, 214, 266–278. [Google Scholar] [CrossRef]
- Jansson, K.U.; Nilsson, M.; Esseen, P.A. Length and classification of natural and created forest edges in boreal landscapes throughout northern Sweden. For. Ecol. Manag. 2011, 262, 461–469. [Google Scholar] [CrossRef]
- Braithwaite, N.T.; Mallik, A.U. Edge effects of wildfire and riparian buffers along boreal forest streams. J. Appl. Ecol. 2012, 49, 192–201. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Mayerhofer, M.S.; Biswas, S.R.; Esseen, P.A.; Hylander, K.; Stewart, K.J.; Mallik, A.U.; Drapeau, P.; Jonsson, B.G.; et al. Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. J. Ecol. 2015, 103, 550–562. [Google Scholar] [CrossRef]
- Laurance, W.F.; Ferreira, L.V.; Rankin-de Merona, J.M.; Laurance, S.G. Rain forest fragmentation and the dynamics of Amazonian tree communities. Ecology 1998, 79, 2032–2040. [Google Scholar] [CrossRef]
- Camargo, J.L.; Kapos, V. Complex edge effects on soil moisture and microclimate in central Amazonian forest. J. Trop. Ecol. 1995, 11, 205–221. [Google Scholar] [CrossRef]
- Williams-Linera, G. Vegetation structure and environmental conditions of forest edges in Panama. J. Ecol. 1990, 78, 356–373. [Google Scholar] [CrossRef]
- Kruys, N.; Jonsson, B.G. Insular patterns of calicioid lichens in a boreal old-growth forest-wetland mosaic. Ecography 1997, 20, 605–613. [Google Scholar] [CrossRef]
- Esseen, P.A.; Renhorn, K.E. Edge effects on an epiphytic lichen in fragmented forests. Conserv. Biol. 2008, 12, 1307–1317. [Google Scholar] [CrossRef]
- Moen, J.; Jonsson, B.G. Edge effects on liverworts and lichens in forest patches in a mosaic of boreal forest and wetland. Conserv. Biol. 2003, 17, 380–388. [Google Scholar] [CrossRef]
- Hylander, K. Aspect modifies the magnitude of edge effects on bryophyte growth in boreal forests. J. Appl. Ecol. 2005, 42, 518–525. [Google Scholar] [CrossRef]
- Chen, J.; Franklin, J.F.; Spies, T.A. Vegetation responses to edge environments in old-growth Douglas-fir forests. Ecol. Appl. 1992, 2, 387–396. [Google Scholar] [CrossRef]
- Roberge, J.M.; Bengtsson, S.B.; Wulff, S.; Snäll, T. Edge creation and tree dieback influence the patch-tracking metapopulation dynamics of a red-listed epiphytic bryophyte. J. Appl. Ecol. 2011, 48, 650–658. [Google Scholar] [CrossRef]
- Pellissier, V.; Bergès, L.; Nedeltcheva, T.; Schmitt, M.C.; Avon, C.; Cluzeau, C.; Dupouey, J.L. Understorey plant species show long-range spatial patterns in forest patches according to distance-to-edge. J. Veg. Sci. 2013, 24, 9–24. [Google Scholar] [CrossRef]
- Baker, T.P.; Baker, S.C.; Dalton, P.J.; Fountain-Jones, N.M.; Jordan, G.J. Temporal persistence of edge effects on bryophytes within harvested forests. For. Ecol. Manag. 2016, 375, 223–229. [Google Scholar] [CrossRef]
- Gjerde, I.; Sætersdal, M.; Blom, H.H. Complementary hotspot inventory—A method for identification of important areas for biodiversity at the forest stand level. Biol. Conserv. 2007, 137, 549–557. [Google Scholar] [CrossRef]
- Timonen, J.; Siitonen, J.; Gustafsson, L.; Kotiaho, J.S.; Stokland, J.N.; Sverdrup-Thygeson, A.; Mönkkönen, M. Woodland key habitats in northern Europe: Concepts, inventory and protection. Scand. J. For. Res. 2010, 25, 309–324. [Google Scholar] [CrossRef]
- Ericsson, T.S.; Berglund, H.; Östlund, L. History and forest biodiversity of woodland key habitats in south boreal Sweden. Biol. Conserv. 2005, 122, 289–303. [Google Scholar] [CrossRef]
- Timonen, J.; Gustafsson, L.; Kotiaho, J.S.; Mönkkönen, M. Hotspots in cold climate: Conservation value of woodland key habitats in boreal forests. Biol. Conserv. 2011, 144, 2061–2067. [Google Scholar] [CrossRef]
- Aune, K.; Jonsson, B.G.; Moen, J. Isolation and edge effects among woodland key habitats in Sweden: Is forest policy promoting fragmentation? Biol. Conserv. 2005, 124, 89–95. [Google Scholar] [CrossRef]
- Paal, J.; Prieditis, N.; Rannik, R.; Jeletsky, E.M. Classification structure of floodplain forests in Estonia: A comparison of two classification approaches. Ann. Bot. Fenn. 2008, 45, 255–268. [Google Scholar] [CrossRef]
- Prieditis, N. Alnus glutinosa–dominated wetland forests of the Baltic Region: Community structure, syntaxonomy and conservation. Plant Ecol. 1997, 129, 49–94. [Google Scholar] [CrossRef]
- McVean, D.N. Ecology of Alnus Glutinosa (L.) Gaertn.: VII. Establishment of alder by direct seeding of shallow blanket bog. J. Ecol. 1959, 47, 615–618. [Google Scholar] [CrossRef]
- Gill, C.J. 1975. The Ecological significance of adventitious rooting as a response to flooding in woody species, with special reference to Alnus glutinosa (L.) Gaertn. Flora 1975, 164, 85–97. [Google Scholar] [CrossRef]
- Ahti, T.; Hämet-Ahti, L.; Jalas, J. Vegetation zones and their sections in northwestern Europe. Ann. Bot. Fenn. 1968, 5, 169–211. [Google Scholar]
- Latvian Environment, Geology and Meteorology Centre Home Page. Available online: http://www.meteo.lv (accessed on 3 March 2020).
- Pickett, S.T.A. Space-for-Time Substitution as an Alternative to Long-Term Studies. In Long-Term Studies in Ecology; Likens, G.E., Ed.; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Bušs, K. Latvijas PSR Meža Tipololoģijas Pamati; LRZTIPI: Rīga, Latvia, 1976. [Google Scholar]
- Ek, T.; Suško, U.; Auziņš, R. Mežaudžu Atslēgas Biotopu Inventarizācija. Metodika; Valsts Meža Dienests: Rīga, Latvija, 2002; pp. 1–76. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie: Grundzfige der Vegetationskunde; Springer: Wien, Austria, 1964. [Google Scholar]
- Gavrilova, G.; Šulcs, V. Flora of Latvian Vascular Plants: List of Taxa; Institute of Biology of University of Latvia: Rīga, Latvia, 1999; pp. 1–136. [Google Scholar]
- Āboliņa, A.; Piterāns, A.; Bambe, B. Latvijas Kērpji un Sūnas. Taksonu Saraksts; DU AA “Saule”: Salaspils, Latvia, 2015; pp. 1–213. ISBN 978-9984-14-735-2. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; pp. 9–79. ISBN 0-691-08485-8. [Google Scholar]
- Cain, S.A. Life-forms and phytoclimate. Bot. Rev. 1950, 16, 1–32. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Wilson, P.J.; Hunt, R.; Grime, J.P.; Thompson, K. Allocating CSR plant functional types: A soft approach to a hard problem. Oikos 1999, 85, 282–294. [Google Scholar] [CrossRef]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimeš, L.; Klimešová, J.; et al. The LEDA traitbase: A database of life-history traits of the northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Fitter, A.H.; Peat, H.J. The ecological flora database. J. Ecol. 1994, 82, 415–425. [Google Scholar] [CrossRef]
- Flora for Vascular Plants in Central Russia Home Page. Available online: https://www.impb.ru/eco/index.php (accessed on 1 February 2020).
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von pflanzen in mitteleuropa. Scr. Geobot. 1992, 18, 1–248. [Google Scholar]
- Ministru Kabineta Noteikumi Nr. 396. Noteikumi Par īpaši Aizsargājamo Sugu un Ierobežoti Izmantojamo īpaši Aizsargājamo Sugu Sarakstu. (In Latvian). Available online: http://m.likumi.lv/doc.php?id=12821 (accessed on 17 March 2020).
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org (accessed on 5 February 2020).
- Matlack, G.R. Microenvironment variation within and among forest edge sites in the eastern United States. Biol. Conserv. 1993, 66, 185–194. [Google Scholar] [CrossRef]
- Marozas, V.; Grigaitis, V.; Brazaitis, G. Edge effect on ground vegetation in clear-cut edges of pine-dominated forests. Scand. J. For. Res. 2005, 20, 43–48. [Google Scholar] [CrossRef]
- Matlack, G. Vegetation dynamics of the forest edge-trends in space and successional time. J. Ecol. 1994, 82, 113–123. [Google Scholar] [CrossRef]
- Hutchison, B.A.; Matt, D.R. The distribution of solar radiation within a deciduous forest. Ecol. Monogr. 1977, 47, 185–207. [Google Scholar] [CrossRef]
- Kneeshaw, D.; Bergeron, Y.; Kuuluvainen, T. Forest ecosystem structure and disturbance dynamics across the circumboreal forest. In The Sage Handbook of Biogeography; Millington, A., Blumler, M., Schickhoff, U., Eds.; Sage: Los Angeles, CA, USA, 2011; pp. 263–280. ISBN 978-141-291-951-7. [Google Scholar]
- Brothers, T.S.; Spingarn, A. Forest fragmentation and alien plant invasion of central Indiana old-growth forests. Conserv. Biol. 1992, 6, 91–100. [Google Scholar] [CrossRef]
- Hermy, M.; Honnay, O.; Firbank, L.; Grashof-Bokdam, C.; Lawesson, J.E. An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol. Conserv. 1999, 91, 9–22. [Google Scholar] [CrossRef]
- Honnay, O.; Verheyen, K.; Hermy, M. Permeability of ancient forest edges for weedy plant species invasion. For. Ecol. Manag. 2002, 161, 109–122. [Google Scholar] [CrossRef]
- Sekgororoane, G.B.; Dilworth, T.G. Relative abundance, richness, and diversity of small mammals at induced forest edges. Can. J. Zool. 1995, 73, 1432–1437. [Google Scholar] [CrossRef]
- Lovejoy, T.E.; Bierregaard, R.O., Jr.; Rylands, A.B.; Malcolm, J.R.; Quintela, C.E.; Harper, L.H.; Brown, K.S., Jr.; Powell, A.H.; Powell, G.V.N.; Schubart, H.O.R.; et al. Edge and other effects of isolation on Amazon forest fragments. In Conservation Biology; Soule, M., Ed.; Sinauer Associates: Sunderland, MA, USA, 1986; pp. 257–285. ISBN 978-087-893-794-3. [Google Scholar]
- Gignac, L.D.; Dale, M.R. Effects of size, shape, and edge on vegetation in remnants of the upland boreal mixed-wood forest in agro-environments of Alberta, Canada. Botany 2007, 85, 273–284. [Google Scholar] [CrossRef]
- Lawes, M.J.; Lamb, B.C.; Boudreau, S. Area-but no edge-effect on woody seedling abundance and species richness in old Afromontane forest fragments. J. Veg. Sci. 2005, 16, 363–372. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E. Structure and composition of edges next to regenerating clear-cuts in mixed-wood boreal forest. J. Veg. Sci. 2002, 13, 535–546. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]




| Variable | Species Group | Adjacent Stand Age Group | ||
|---|---|---|---|---|
| 0–20 vs 20–40 | 20–40 vs >41 | 0–20 vs >41 | ||
| Projective coverage | E0 | ns | ns | ns |
| E1 | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| E2 | ns | ns | p < 0.05 ↑ | |
| E3 | ns | ns | p < 0.05 ↓ | |
| Number of species | E0 | p < 0.05 ↑ | p < 0.05 ↑ | ns |
| E1 | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| E2 | ns | ns | ns | |
| E3 | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| Total | p < 0.001 ↑ | ns | p < 0.001 ↑ | |
| Plant functional types (E1 layer) | ||||
| Habitat group | Boreal species | ns | ns | ns |
| Nemoral species | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| Nitrophilous species | p < 0.001 ↑ | ns | p < 0.05 ↑ | |
| Adventive species | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| Meadow species | ns | ns | ns | |
| Swamp forest species | p < 0.05 ↑ | ns | p < 0.001 ↑ | |
| Seed dispersal agent | Wind | p < 0.05 ↑ | ns | ns |
| Water | ns | ns | ns | |
| Other animals | p < 0.05 ↓ | ns | p < 0.05 ↓ | |
| Birds | ns | ns | p < 0.001 ↑ | |
| Plant strategy group | C | p < 0.05 ↑ | ns | p < 0.05 ↑ |
| CS | ns | ns | ||
| CR | p < 0.05 ↑ | ns | p < 0.05 ↑ | |
| CSR | p < 0.001 ↑ | ns | p < 0.001 ↑ | |
| Raunkiaer life history form | Phanerophyte | p < 0.05 ↑ | p < 0.05 ↑ | ns |
| Hemicryptophyte | p < 0.001 ↑ | ns | p < 0.05 ↑ | |
| Therophyte | ns | ns | ns | |
| Geophyte | ns | ns | ns | |
| Environmental factor | Temperature | ns | ns | ns |
| Moisture | ns | ns | ns | |
| Soil pH | p < 0.05 ↑ | ns | ns | |
| Light-demanding species (≥6) | p < 0.001 ↑ | ns | p < 0.001 ↑ | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liepa, L.; Rendenieks, Z.; Jansons, Ā.; Straupe, I.; Dubrovskis, E.; Miezīte, O. The Persisting Influence of Edge on Vegetation in Hemiboreal Alnus Glutinosa (L.) Gaertn. Swamp Forest Set-Asides Adjacent to Recently Disturbed Stands. Forests 2020, 11, 1084. https://doi.org/10.3390/f11101084
Liepa L, Rendenieks Z, Jansons Ā, Straupe I, Dubrovskis E, Miezīte O. The Persisting Influence of Edge on Vegetation in Hemiboreal Alnus Glutinosa (L.) Gaertn. Swamp Forest Set-Asides Adjacent to Recently Disturbed Stands. Forests. 2020; 11(10):1084. https://doi.org/10.3390/f11101084
Chicago/Turabian StyleLiepa, Līga, Zigmārs Rendenieks, Āris Jansons, Inga Straupe, Edgars Dubrovskis, and Olga Miezīte. 2020. "The Persisting Influence of Edge on Vegetation in Hemiboreal Alnus Glutinosa (L.) Gaertn. Swamp Forest Set-Asides Adjacent to Recently Disturbed Stands" Forests 11, no. 10: 1084. https://doi.org/10.3390/f11101084
APA StyleLiepa, L., Rendenieks, Z., Jansons, Ā., Straupe, I., Dubrovskis, E., & Miezīte, O. (2020). The Persisting Influence of Edge on Vegetation in Hemiboreal Alnus Glutinosa (L.) Gaertn. Swamp Forest Set-Asides Adjacent to Recently Disturbed Stands. Forests, 11(10), 1084. https://doi.org/10.3390/f11101084