

Forest–Fruticulture Conversion Alters Soil Traits and Soil Organic Matter Compartments


 , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Influence of Land Use on Soil Traits in a Tropical Ultisol from Brazil’s Legal Amazon
2.2. Influence of Land Use and Soil Depth on Soil Organic Matter Compartments in a Tropical Ultisol from Brazil’s Legal Amazon
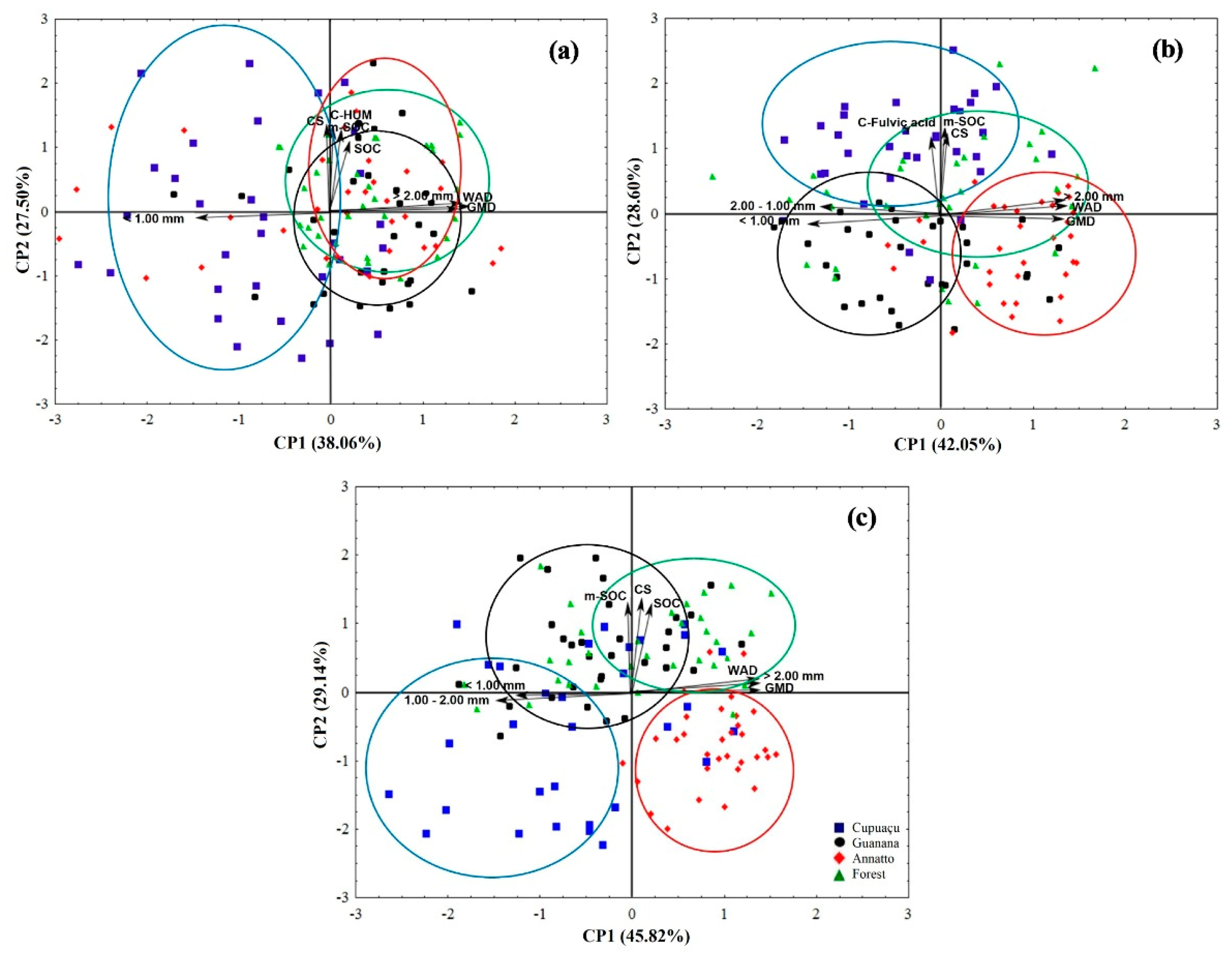
2.3. Multivariate Analysis
3. Discussion
4. Materials and Methods
4.1. Study Site, Climatic Conditions, and Soil Type
4.2. The Studied Crop Systems and Natural Ecosystem
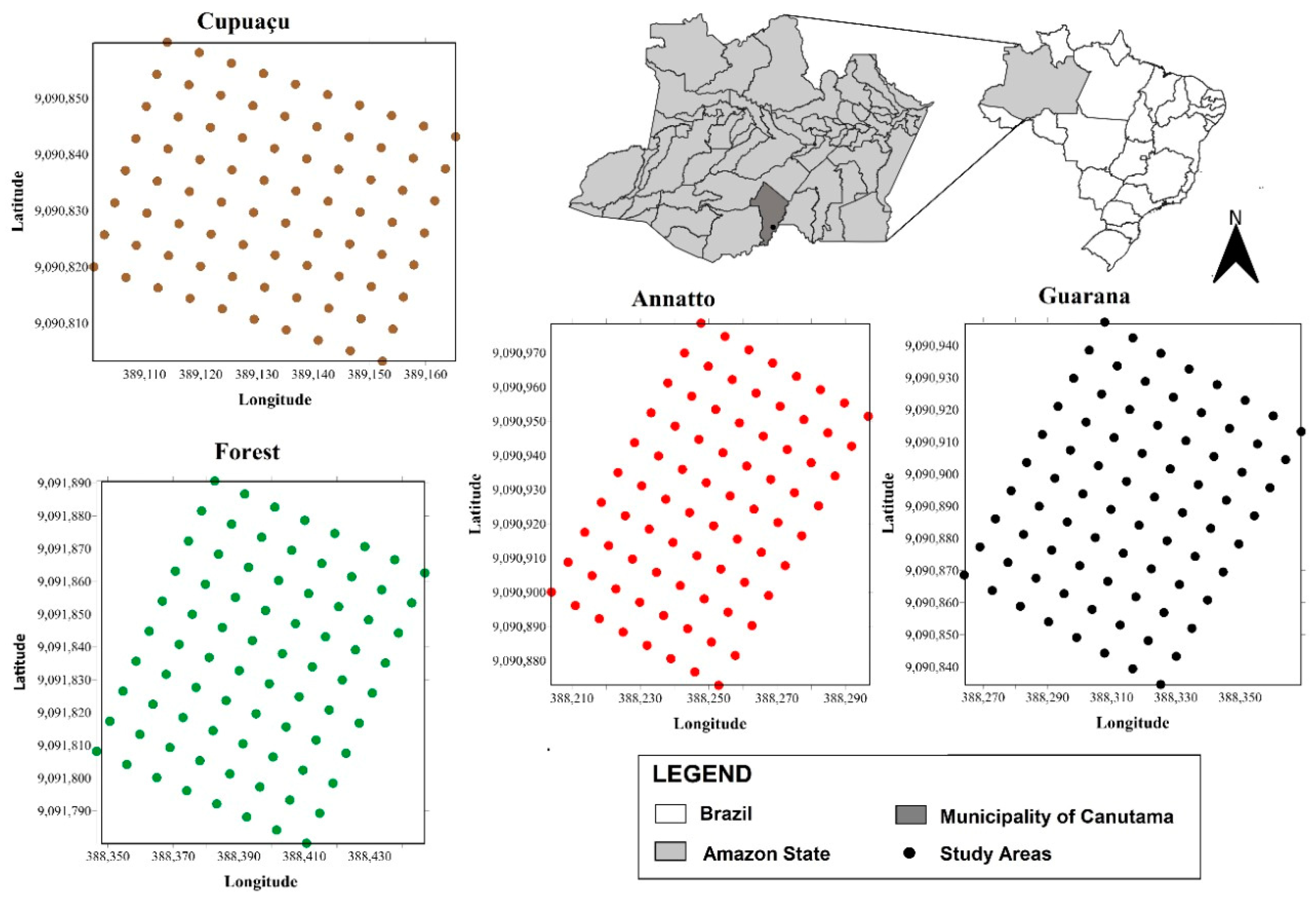
4.3. Experimental Design
4.4. Soil Sampling
4.5. SOM Fractioning
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Souza, T.; Barros, I.C.; Silva, L.J.R.; Laurindo, L.K.; Nascimento, G.S.; Lucena, E.O.; Martins, M.; Santos, V.B. Soil microbiota community assembling in native plant species from Brazil’s legal Amazon. Symbiosis 2022, 86, 93–109. [Google Scholar] [CrossRef]
- Brito, W.B.M.; Campos, M.C.C.; de Souza, F.G.; Silva, L.S.; da Cunha, J.M.; de Lima, A.F.L.; Martins, T.S.; de Oliveira, F.P.; de Oliveira, I.A. Spatial patterns of magnetic susceptibility optimized by anisotropic correction in different Alisols in southern Amazonas, Brazil. Precis. Agric. 2021, 23, 419–449. [Google Scholar] [CrossRef]
- Peralta, Z.M.C.; Alvarado, J.W.V.; Alvez-Valles, C.M. Biodiversity Islands and Dominant Species in Agricultural Landscapes of the South Western Amazon, Perú. In Biodiversity Islands: Strategies for Conservation in Human-Dominated Environments; Topics in Biodiversity and Conservation; Montagnini, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2022; Volume 20. [Google Scholar] [CrossRef]
- Marca-Zevallos, M.J.; Moulatlet, G.M.; Sousa, T.R.; Schietti, J.; Coelho, L.D.S.; Ramos, J.F.; Filho, D.D.A.L.; Amaral, I.L.; Matos, F.D.D.A.; Rincón, L.M.; et al. Local hydrological conditions influence tree diversity and composition across the Amazon basin. Ecography 2022, e06125. [Google Scholar] [CrossRef]
- Patoine, G.; Eisenhauer, N.; Cesarz, S.; Phillips, H.R.P.; Xu, X.; Zhang, L.; Guerra, C.A. Drivers and trends of global soil microbial carbon over two decades. Nat. Commun. 2022, 13, e4195. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.F. Case Studies of Ecosystem Control and Stability. In Evolution from a Thermodynamic Perspective; Jordan, C.F., Ed.; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Souza, T. Soil Biology in Tropical Ecosystems; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Banerjee, S.; van der Heijden, M.G.A. Soil microbiomes and one health. Nat. Rev. Microbiol. 2022, 38, 1–15. [Google Scholar] [CrossRef]
- Cecagno, D.; Gomes, M.V.; Costa, S.E.V.G.A.; Martins, A.P.; Denardin, L.O.; Bayer, C.; Anghinoni, I.; Carvalho, P.C.F. Soil organic carbon in an integrated crop-livestock system under different grazing intensities. Rev. Bras. Ciênc. Agr. 2018, 13, 1–7. [Google Scholar] [CrossRef]
- Panchal, P.; Preece, C.; Peñuelas, J.; Giri, J. Soil carbon sequestration by root exudates. Trends Plant Sci. 2022, 27, 1360–1385. [Google Scholar] [CrossRef]
- Estrada, A.; Garber, P.A.; Gouveia, S.; Fernández-Llamazares, Á.; Ascensão, F.; Fuentes, A.; Garnett, S.T.; Shaffer, C.; Bicca-Marques, J.; Fa, J.E.; et al. Global importance of Indigenous Peoples, their lands, and knowledge systems for saving the world’s primates from extinction. Sci. Adv. 2022, 8, 32. [Google Scholar] [CrossRef]
- Darriaut, R.; Lailheugue, V.; Masneuf-Pomarède, I.; Marguerit, E.; Martins, G.; Compant, S.; Ballestra, P.; Upton, S.; Ollat, N.; Lauvergeat, V. Grapevine rootstock and soil microbiome interactions: Keys for a resilient viticulture. Hortic. Res. 2022, 9, uhac019. [Google Scholar] [CrossRef]
- Edwards, C.A.; Arancon, N.Q. The Role of Earthworms in Organic Matter and Nutrient Cycles. In Biology and Ecology of Earthworms; Springer: New York, NY, USA, 2022. [Google Scholar] [CrossRef]
- Teague, R.; Dowhower, S. Links of microbial and vegetation communities with soil physical and chemical factors for a broad range of management of tallgrass prairie. Ecol. Indic. 2022, 142, e109280. [Google Scholar] [CrossRef]
- Souza, T.; Dobner, M., Jr.; Schmitt, D.E.; Silva, L.J.R.; Schneider, K. Soil biotic and abiotic traits as driven factors for site quality of Araucaria angustifolia plantations. Biologia 2022, 77, 1219–1230. [Google Scholar] [CrossRef]
- Brito, W.B.M.; Campos, M.C.C.; Mantovanelli, B.C.; Cunha, J.M.; Franciscon, U.; Soares, M.D.R. Spatial variability of soil physical properties in Archeological Dark Earths under different uses in southern Amazon. Soil Tillage Res. 2018, 182, 103–111. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuk, J.; Martin-Neto, L.; Ernani, P.R. Stocks and humification degree of organic matter fractions as affected by no-tillage on a subtropical soil. Plant Soil 2002, 238, 133–140. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Pivetta, L.A. Mechanical and biological approaches to alleviate soil compaction in tropical soils: Assessed by root growth and activity (Rb uptake) of soybean and maize grown in rotation with cover crops. Soil Use Manag. 2017, 33, 141–152. [Google Scholar] [CrossRef]
- Souza, F.G.; Campos, M.C.C.; Pinheiro, E.N.; Lima, A.F.L.D.; Brito Filho, E.G.D.; Cunha, J.M.; Brito, W.B.M. Aggregate stability and carbon stocks in Forest conversion to different cropping systems in Southern Amazonas, Brazil. Carbon Manag. 2020, 11, 81–96. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, V.K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Lima, A.F.L.; Campos, M.C.C.; Enck, B.F.; Simões, W.S.; Araújo, R.M.; Santos, L.A.C.; Cunha, J.M. Physical soil attributes in areas under forest/pasture conversion in northern Rondônia, Brazil. Environ. Monit. Dnd Assess. 2022, 194, 34–43. [Google Scholar] [CrossRef]
- Kukuļs, I.; Kļaviņš, M.; Nikodemus, O.; Kasparinskis, R.; Brūmelis, G. Changes in soil organic matter and soil humic substances following the afforestation of former agricultural lands in the boreal-nemoral ecotone (Latvia). Geoderma Reg. 2019, 16, e00213. [Google Scholar] [CrossRef]
- da Silva, L.J.R.; Souza, T.; Laurindo, L.K.; Nascimento, G.d.S.; de Lucena, E.O.; Freitas, H. Aboveground Biomass, Carbon Sequestration, and Yield of Pyrus pyrifolia under the Management of Organic Residues in the Subtropical Ecosystem of Southern Brazil. Agronomy 2022, 12, 231. [Google Scholar] [CrossRef]
- Marinho Junior, J.L.; Piscoya, V.C.; Fernandes, M.M.; Gonçalves, S.B.; Holanda, F.S.R.; Cunha Filho, M.; Gomes Filho, R.R.; Pedrotti, A.; Araújo Filho, R.N. Carbon dynamics in humic fractions of soil organic matter under different vegetation cover in southern Tocantins. Floresta Ambiente 2021, 28, e20200024. [Google Scholar] [CrossRef]
- Frozzi, J.C.; Cunha, J.M.; Campos, M.C.C.; Bergamin, A.C.; Brito, W.B.M.; Fraciscon, U.; Silva, D.M.P.; Lima, A.F.L.; Brito Filho, E.G. Physical attributes and organic carbon in soils under natural and anthropogenic environments in the South Amazon region. Environ. Earth Sci. 2020, 79, 251–266. [Google Scholar] [CrossRef]
- Jordão, H.W.C.; Campos, M.C.C.; Mantovanelli, B.C.; Frozzi, J.C.; Cunha, J.M.; Chechi, L.; Silva, J.F. Attributes of pedoindicator soils in areas cultivated with typical crops in the Western Amazon, Brazil. Biosci. J. 2021, 36, 97–108. [Google Scholar] [CrossRef]
- Briedis, C.; Sá, J.C.M.; De-Carli, R.S.; Antunes, E.A.P.; Simon, L.; Romko, M.L.; Elias, L.S.; Ferreira, A.O. Particulate soil organic carbon and stratification ratio increases in response to crop residue decomposition under no-till. Rev. Bras. Ciênc. Solo 2012, 36, 1483–1490. [Google Scholar] [CrossRef]
- Mantovanelli, B.C.; Alho, L.C.; Campos, M.C.C.; Cunha, J.M.; Oliveira, I.A. Pedoindicators attributes in the variation of CO efflux in Indian black earth and non-anthropic soils. Carbon Manag. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Moreiram, A.; Fageria, N.K. Soil Chemical Attributes of Amazonas State, Brazil. Commun. Soil Sci Plant 2009, 40, 2912–2925. [Google Scholar] [CrossRef]
- Marques, J.D.O.; Luizão, F.J.; Teixeira, W.G.; Vitel, C.M.; Marques, E.M.A. Soil organic carbon, carbon stock and their relationships to physical attributes under forest soils in Central Amazonia. Rev. Árvore 2016, 40, 197–208. [Google Scholar] [CrossRef]
- Spaccini, R.; Mbagwu, J.S.C.; Conte, P.; Piccolo, A. Changes of humic substances characteristics from forested to cultivated soils in Ethiopia. Geoderma 2005, 132, 9–19. [Google Scholar] [CrossRef]
- Marques, J.D.O.; Luizão, F.J.; Teixeira, W.G.; Nogueira, E.M.; Fearnside, P.M.; Sarrazin, M. Soil carbon stocks under amazonian forest: Distribution in the soil fractions and vulnerability to emission. Open J. For. 2017, 7, 121–142. [Google Scholar] [CrossRef][Green Version]
- Oliveira, I.A.; Campos, M.C.C.; Siqueira, D.S.; Freitas, L.; Aquino, R.E.; Oliveira, F.P. Pedoenvironmental indicators of soil inWestern Amazonia, Brazil. Environ. Monit. Assess 2020, 192, 768–770. [Google Scholar] [CrossRef]
- Guimarães, D.V.; Gonzaga, M.I.S.; Silva, T.O.; Silva, T.L.; Dias, N.S.; Matias, M.I.S. Soil organic matter pools and carbon fractions in soil under different land uses. Soil Till. Res. 2013, 126, 177–182. [Google Scholar] [CrossRef]
- Oliveira, D.M.S.; Paustian, K.; Davies, C.; Cherubin, M.R.; Franco, A.L.C.; Cerri, C.C.; Cerri, C.E.P. Soil carbon changes in areas undergoing expansion of sugarcane into pastures in Brazil. Agric. Ecosyst. Environ. 2016, 228, 38–48. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorolgische Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Brazil; Ministerio das Minas e Energia; Departamento Nacional da Produçao Mineral (DNPM) Brazil; Ministerio das Minas e Energia, Secretaria-Geral (MME/SG). Projeto RADAMBRASIL, Levantamento de Recursos Naturais; Departamento Nacional de Produção Mineral. Folha SC—20. MAPA: Porto Velho, Brazil; Rio de Janeiro, Brazil, 1978; Volume 16. [Google Scholar]
- WRB—IUSS Working Group. World Reference Base for Soil; World Soil Resources Reports: Rome, Italy, 2015. [Google Scholar]
- Black, C.A. Methods of Soil Analysis, Part 2. In Agronomy Monograph; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; Volume 9, pp. 771–1572. [Google Scholar]
- Teixeira, P.C.; Donagema, G.K.; Ademir, F.; Teixeira, W.G. Manual de Métodos de Análise de Solo, 3rd ed.; Embrapa: Brasília, Brazil, 2017. [Google Scholar]
- IITA. Selected Methods for Soil and Plant Analysis; IITA Manual Services: Ibadan, Nigeria, 1979. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties. In Soil Science Society of America; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Olsen, S.; Cole, C.; Watanabe, F.; Dean, L. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA Circular Nr 939; US Gov. Print. Office: Washington, WA, USA; Madison, WI, USA, 1954.
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Plant and Soil Analysis: A Working Manual; Technical Bulletin n. 1. Nairobi; Tropical Soil Biology and Fertility Programme. Soil Science Society East Africa: UNESCO, ROSTA, 1993. [Google Scholar]
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Soil Sci Plant 1988, 19, 1467–1476. [Google Scholar] [CrossRef]
- Swift, R.S. Organic matter characterization. In Methods of Soil Analysis: Chemical Methods. Soil Science Society of America; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1996; Volume 3, pp. 1011–1020. [Google Scholar] [CrossRef]
- Benites, V.M.; Madari, B.; Machado, P.L.O.A. Extração e Fracionamento Quantitativo de Substâncias Húmicas do Solo: Um Procedimento Simplificado de Baixo Custo; Embrapa Solos: Rio de Janeiro, Brazil, 2003. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing. 2018. Available online: http://www.r-project.org/ (accessed on 17 October 2021).



{kind=link}
{kind=link}
| Soil Traits | P. cupana | T. grandiflorum | B. orellana | Amazon Rainforest | F-Value |
|---|---|---|---|---|---|
| Physical traits | |||||
| Bulk Density (g cm−3) | 1.11 (0.01) a | 1.07 (0.02) b | 1.04 (0.02) b | 0.92 (0.05) c | 17.92 *** |
| Geometric mean diameter (mm) | 2.49 (0.04) b | 2.38 (0.01) c | 2.72 (0.04) a | 2.57 (0.02) b | 13.44 *** |
| Weighted average diameter (mm) | 2.96 (0.02) b | 2.90 (0.05) b | 3.12 (0.04) a | 3.04 (0.02) a | 9.47 ** |
| >2.00 (mm) | 86.43 (0.31) b | 83.89 (0.27) b | 92.37 (0.54) a | 89.53 (0.38) a | 9.98 ** |
| 1.00–2.00 (mm) | 3.36 (0.11) b | 4.33 (0.12) a | 1.44 (0.09) d | 2.52 (0.17) c | 18.35 *** |
| <1.00 (mm) | 10.27 (0.02) a | 11.57 (0.06) a | 6.18 (0.02) c | 8.35 (0.03) b | 15.91 *** |
| Soil macroporosity (mm3 mm−3) | 9.86 (0.05) c | 13.20 (0.12) a | 12.48 (0.10) b | 12.46 (0.01) b | 13.79 *** |
| Soil microporosity (mm3 mm−3) | 37.04 (0.21) a | 19.68 (0.14) c | 33.19 (0.25) b | 20.79 (0.11) c | 18.26 *** |
| Total soil porosity (mm3 mm−3) | 47.41 (0.51) a | 37.11 (0.47) b | 45.16 (0.45) a | 36.79 (0.61) b | 9.31 ** |
| MG (mm3 mm−3) | 33.35 (0.40) a | 27.51 (0.69) b | 32.49 (1.09) a | 27.59 (0.41) b | 9.53 ** |
| Sand (g kg−1) | 365.94 (11.01) b | 277.41 (9.01) c | 384.64 (8.37) a | 238.39 (2.11) d | 12.34 *** |
| Silt (g kg−1) | 405.31 (9.31) c | 450.31 (12.51) b | 386.74 (21.09) c | 524.70 (17.01) a | 14.58 *** |
| Clay (g kg−1) | 223.91 (3.31) d | 267.64 (2.38) a | 239.77 (5.57) c | 260.47 (3.67) b | 11.94 *** |
| Chemical traits | |||||
| Soil pH | 3.89 (0.01) a | 3.92 (0.02) a | 3.87 (0.01) a | 3.74 (0.02) a | 3.45 ns |
| Al3+ (cmolc dm−3) | 4.52 (0.01) b | 5.11 (0.04) a | 5.04 (0.04) a | 5.04 (0.06) a | 8.94 * |
| H+ + Al3+ (cmolc dm−3) | 9.34 (0.09) c | 10.42 (0.08) a | 9.96 (0.01) b | 9.95 (0.02) b | 9.31 ** |
| K+ (cmolc dm−3) | 0.11 (0.01) a | 0.06 (0.02) b | 0.07 (0.01) b | 0.08 (0.03) b | 8.56 * |
| Ca2+ (cmolc dm−3) | 1.21 (0.03) a | 0.86 (0.01) b | 0.92 (0.03) b | 0.71 (0.02) c | 11.49 *** |
| Mg2+ (cmolc dm−3) | 0.42 (0.02) a | 0.19 (0.01) c | 0.26 (0.01) b | 0.19 (0.02) c | 10.59 ** |
| P (mg dm−3) | 6.04 (0.08) b | 3.16 (0.11) d | 4.87 (0.09) c | 7.73 (0.10) a | 15.24 *** |
| SB (cmolc dm−3) | 1.94 (0.01) a | 1.14 (0.01) c | 1.29 (0.02) b | 1.04 (0.01) c | 11.13 *** |
| C.E.C (cmolc dm−3) | 10.83 (0.02) b | 11.62 (0.05) a | 11.36 (0.03) a | 11.14 (0.04) a | 10.16 ** |
| V (%) | 16.40 (0.31) a | 10.12 (0.15) c | 11.71 (0.27) b | 9.67 (0.23) c | 11.09 *** |
| m (%) | 71.75 (1.01) c | 82.71 (0.68) a | 79.75 (1.23) b | 83.97 (0.89) a | 11.59 *** |
| CS (t ha−1) | 8.67 (0.28) c | 9.18 (0.27) b | 9.99 (0.13) b | 11.44 (0.97) a | 12.36 *** |
| Soil Depth (m) | B. orellana | T. grandiflorum | P. cupana | Amazon Rainforest |
|---|---|---|---|---|
| SOC (g kg−1) | ||||
| 0.00–0.05 | 20.67 (0.93) Ac 1 | 22.31 (1.03) Ab | 22.43 (1.21) Ab | 26.02 (1.57) Aa |
| 0.05–0.10 | 12.32 (0.81) Bc | 17.20 (2.01) Bb | 14.59 (0.84) Bc | 21.28 (0.98) Aa |
| 0.10–0.20 | 10.79 (0.56) Bb | 6.68 (0.98) Cc | 7.73 (0.34) Cc | 12.86 (1.03) Ba |
| p-SOC (g kg−1) | ||||
| 0.00–0.05 | 6.27 (0.12) Ac | 7.14 (0.09) Ab | 7.66 (0.09) Ab | 8.53 (0.10) Aa |
| 0.05–0.10 | 2.67 (0.23) Bb | 2.80 (0.11) Bb | 3.44 (0.11) Bb | 7.27 (0.10) Aa |
| 0.10–0.20 | 2.54 (0.17) Bb | 1.30 (0.03) Bc | 3.45 (0.06) Bb | 4.18 (0.03) Ba |
| m-SOC (g kg−1) | ||||
| 0.00–0.05 | 14.62 (0.13) Ab | 13.45 (1.17) Ab | 14.58 (0.93) Ab | 16.18 (1.17) Aa |
| 0.05–0.10 | 9.42 (0.15) Bb | 14.12 (0.16) Aa | 10.25 (0.81) Bb | 13.47 (0.99) Ba |
| 0.10–0.20 | 8.74 (0.21) B | 5.23 (0.81) B | 3.98 (0.27) C | 8.94 (0.51) C |
| C-Fulvic acid (g kg−1) | ||||
| 0.00–0.05 | 2.71 (0.05) Ab | 2.95 (0.01) Ab | 1.92 (0.01) Ac | 4.53 (0.01) Aa |
| 0.05–0.10 | 2.31 (0.03) Ab | 3.31 (0.02) Aa | 2.04 (0.05) Ab | 2.71 (0.05) Bb |
| 0.10–0.20 | - | - | - | - |
| C-Humic acid (g kg−1) | ||||
| 0.00–0.05 | 2.91 (0.03) Aa | 3.50 (0.05) Aa | 2.66 (0.02) Ab | 2.98 (0.05) Ab |
| 0.05–0.10 | 2.03 (0.01) Ba | 2.86 (0.01) Ba | 1.19 (0.03) Bb | 1.48 (0.01) Bb |
| 0.10–0.20 | - | - | - | - |
| C-Humine (g kg−1) | ||||
| 0.00–0.05 | 15.37 (0.01) Ab | 15.73 (0.02) Ab | 17.53 (0.02) Aa | 17.41 (0.01) Aa |
| 0.05–0.10 | 7.83 (0.03) Bc | 11.12 (0.05) Bb | 10.82 (0.05) Bb | 16.86 (0.02) Aa |
| 0.10–0.20 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enck, B.F.; Campos, M.C.C.; Pereira, M.G.; de Souza, F.G.; Santos, O.A.Q.; Diniz, Y.V.d.F.G.; Martins, T.S.; Cunha, J.M.; Lima, A.F.L.d.; Souza, T.A.F.d. Forest–Fruticulture Conversion Alters Soil Traits and Soil Organic Matter Compartments. Plants 2022, 11, 2917. https://doi.org/10.3390/plants11212917
Enck BF, Campos MCC, Pereira MG, de Souza FG, Santos OAQ, Diniz YVdFG, Martins TS, Cunha JM, Lima AFLd, Souza TAFd. Forest–Fruticulture Conversion Alters Soil Traits and Soil Organic Matter Compartments. Plants. 2022; 11(21):2917. https://doi.org/10.3390/plants11212917
Chicago/Turabian StyleEnck, Bruna Firmino, Milton Cesar Costa Campos, Marcos Gervásio Pereira, Fernando Gomes de Souza, Otavio Augusto Queiroz Santos, Yan Vidal de Figueiredo Gomes Diniz, Thalita Silva Martins, José Mauricio Cunha, Alan Ferreira Leite de Lima, and Tancredo Augusto Feitosa de Souza. 2022. "Forest–Fruticulture Conversion Alters Soil Traits and Soil Organic Matter Compartments" Plants 11, no. 21: 2917. https://doi.org/10.3390/plants11212917
APA StyleEnck, B. F., Campos, M. C. C., Pereira, M. G., de Souza, F. G., Santos, O. A. Q., Diniz, Y. V. d. F. G., Martins, T. S., Cunha, J. M., Lima, A. F. L. d., & Souza, T. A. F. d. (2022). Forest–Fruticulture Conversion Alters Soil Traits and Soil Organic Matter Compartments. Plants, 11(21), 2917. https://doi.org/10.3390/plants11212917