

Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress
 , , , ,
, , , ,  , , , ,
, , , ,
Abstract
1. Introduction
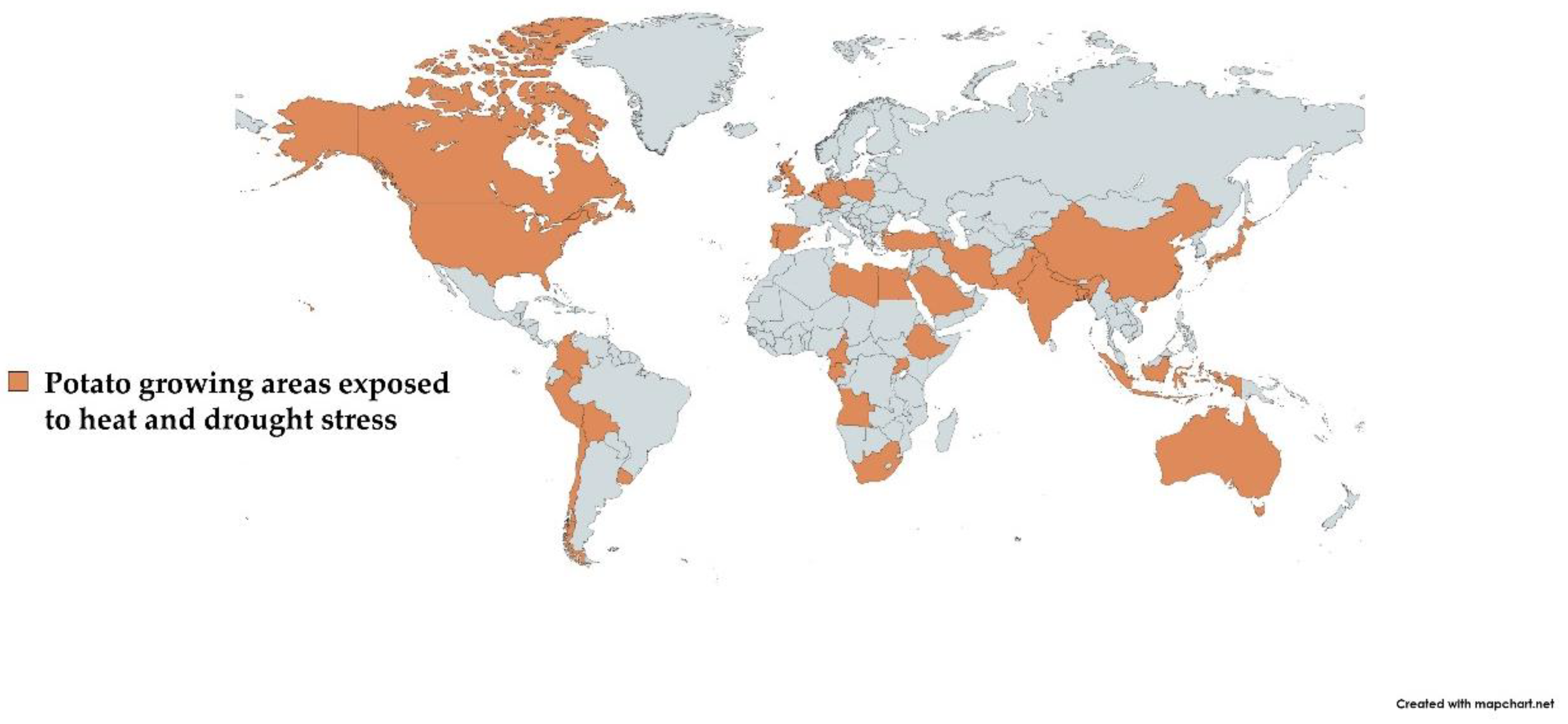
2. Production and Productivity of Potato Affected by Heat and Drought Stress
3. Physiological Changes and Responses of Potato Plants under Heat and Drought Conditions
3.1. Anatomy and Morphology
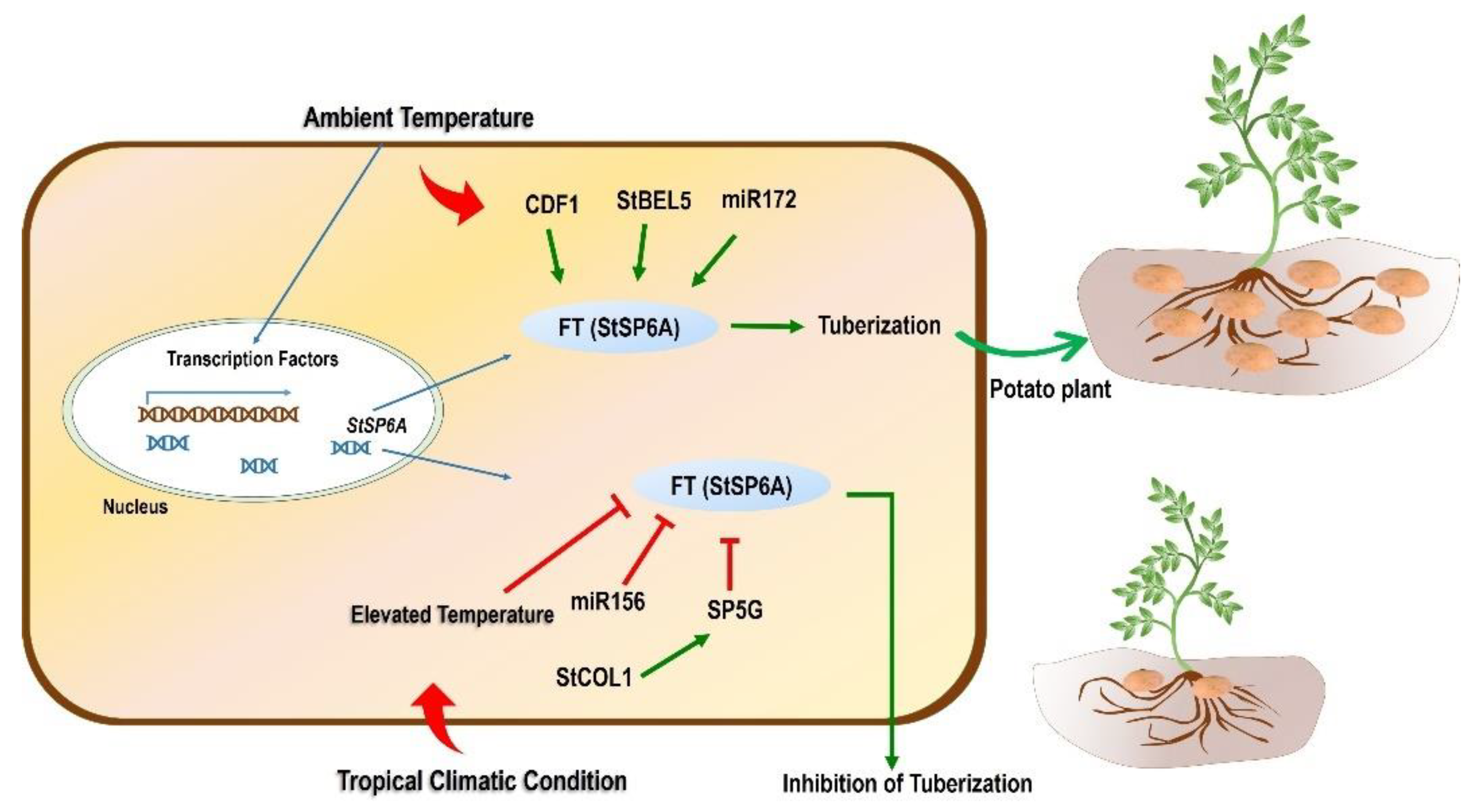
3.2. Role of Photoperiod
3.3. Carbon Partitioning and Source–Sink Relationship
3.4. Tuber Development
3.5. Photosynthesis
3.6. Senescence
3.7. Respiration
3.8. Transpiration
4. Biochemical Changes and Responses of Potato Plants
4.1. Carbohydrate Metabolism
4.2. Ionic and Osmolyte Balance
4.3. Heat Shock Proteins (HSPs)
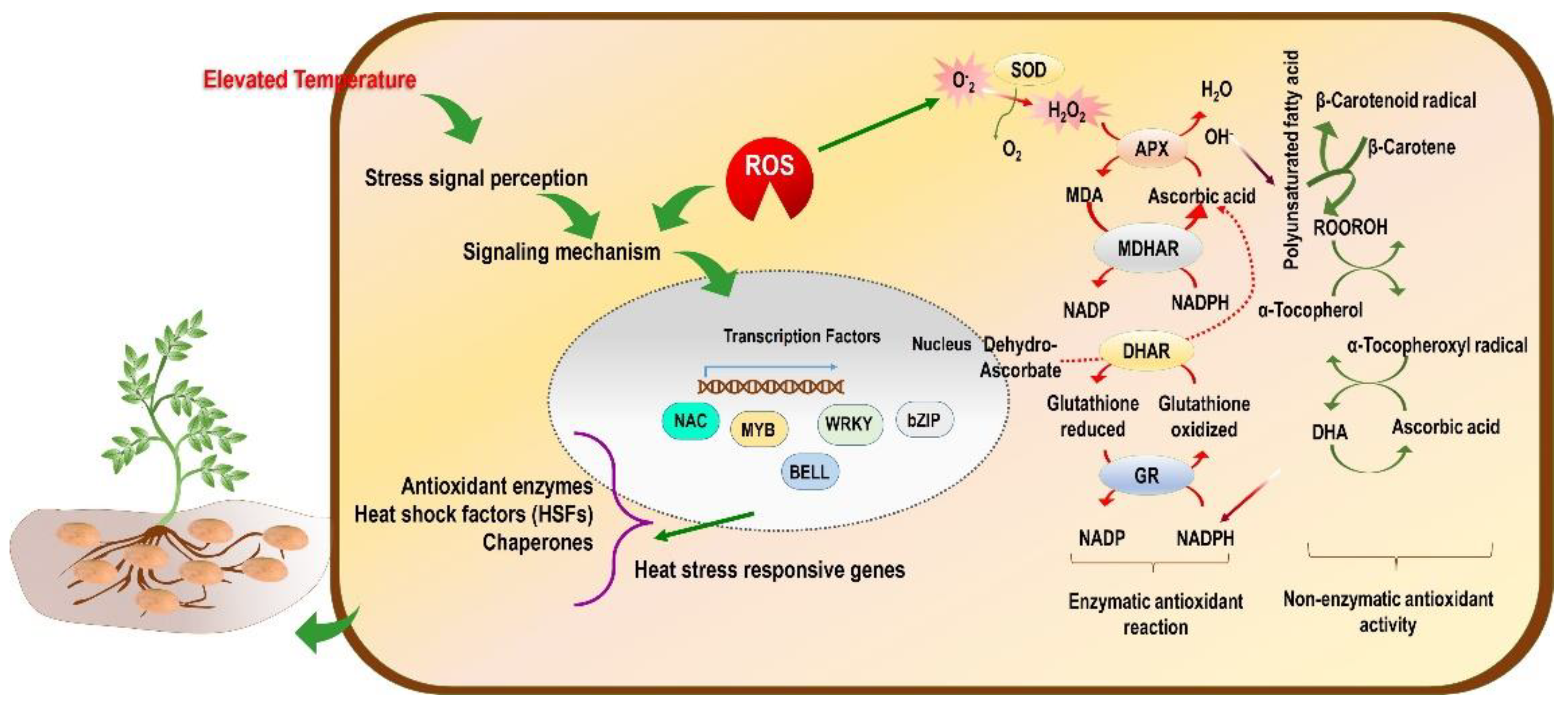
4.4. ROS, RNS, RSS, and Antioxidant System
5. Approaches for Adaptation of Potato Plant
5.1. Agrometeorology-Based Crop Modelling and Agronomic Practices
5.2. Exploring Genetic Diversity and Breeding
5.3. Molecular and Transgenic Approaches
5.4. CRISPR Approach for Molecular Insight and Functional Gene Analysis
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 14 March 2019).
- Lal, M.K.; Kumar, A.; Kardile, H.B.; Raigond, P.; Changan, S.S.; Thakur, N.; Dutt, S.; Tiwari, R.K.; Chourasia, K.N.; Kumar, D.; et al. Biofortification of Vegetables. Adv. Agri-Food Biotechnol. 2020, 105–129. [Google Scholar] [CrossRef]
- Lutaladio, N.B.; Castaldi, L. Potato: The Hidden Treasure. J. Food Compos. Anal. 2009, 22, 491–493. [Google Scholar] [CrossRef]
- Gomez-Zavaglia, A.; Mejuto, J.C.; Simal-Gandara, J. Mitigation of Emerging Implications of Climate Change on Food Production Systems. Food Res. Int. 2020, 134, 109256. [Google Scholar] [CrossRef] [PubMed]
- Stadel, C. Cultivated Landscapes of Native Amazonia and the Andes; Oxford University Press: Oxford, MI, USA, 2005; Volume 25. [Google Scholar]
- Beales, H.L.; Salaman, R.N. The History and Social Influence of the Potato; Cambridge University Press: Cambridge, UK, 1950; Volume 1. [Google Scholar]
- Hijmans, R.J. Global Distribution of the Potato Crop. Am. J. Potato Res. 2001, 78, 403–412. [Google Scholar] [CrossRef]
- Dutt, S.; Manjul, A.S.; Raigond, P.; Singh, B.; Siddappa, S.; Bhardwaj, V.; Kawar, P.G.; Patil, V.U.; Kardile, H.B. Key Players Associated with Tuberization in Potato: Potential Candidates for Genetic Engineering. Crit. Rev. Biotechnol. 2017, 37, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.L. The Vegetabe Crops Agribusiness in Brazil: Current Status and Future Trends. In Proceedings of the Embrapa Hortaliças-Resumo em Anais de Congresso (ALICE), Embrapa, Brazil, 2012. [Google Scholar]
- Staiger, C. International Year of the Potato. Pharm. J. 2008, 281, 743–744. [Google Scholar]
- Devaux, A.; Goffart, J.P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Springer International Publishing: New York, NY, USA, 2019; pp. 3–35. ISBN 9783030286835. [Google Scholar]
- Dahal, K.; Li, X.Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving Potato Stress Tolerance and Tuber Yield under a Climate Change Scenario—A Current Overview. Front. Plant Sci. 2019, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Lal, M.K.; Naga, K.C.; Kumar, R.; Chourasia, K.N.; Subhash, S.; Kumar, D.; Sharma, S. Emerging Roles of Melatonin in Mitigating Abiotic and Biotic Stresses of Horticultural Crops. Sci. Hortic. 2020, 272, 109592. [Google Scholar] [CrossRef]
- Francini, A.; Sebastiani, L. Abiotic Stress Effects on Performance of Horticultural Crops. Horticulturae 2019, 5, 67. [Google Scholar] [CrossRef]
- Muthoni, J.; Kabira, J.N. Potato Production in the Hot Tropical Areas of Africa: Progress Made in Breeding for Heat Tolerance. J. Agric. Sci. 2015, 7, 220–227. [Google Scholar] [CrossRef]
- Hancock, R.D.; Morris, W.L.; Ducreux, L.J.M.; Morris, J.A.; Usman, M.; Verrall, S.R.; Fuller, J.; Simpson, C.G.; Zhang, R.; Hedley, P.E.; et al. Physiological, Biochemical and Molecular Responses of the Potato (Solanum Tuberosum L.) Plant to Moderately Elevated Temperature. Plant Cell Environ. 2014, 37, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J. The Effect of Climate Change on Global Potato Production. Am. J. Potato Res. 2003, 80, 271–279. [Google Scholar] [CrossRef]
- Tang, R.; Niu, S.; Zhang, G.; Chen, G.; Haroon, M.; Yang, Q.; Rajora, O.P.; Li, X.Q. Physiological and Growth Responses of Potato Cultivars to Heat Stress. Botany 2018, 96, 897–912. [Google Scholar] [CrossRef]
- Battisti, D.S.; Naylor, R.L. Historical Warnings of Future Food Insecurity with Unprecedented Seasonal Heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Longmei, N.; Gill, G.K.; Zaidi, P.H.; Kumar, R.; Nair, S.K.; Hindu, V.; Vinayan, M.T.; Vikal, Y. Genome Wide Association Mapping for Heat Tolerance in Sub-Tropical Maize. BMC Genom. 2021, 22, 154. [Google Scholar] [CrossRef]
- Singh, B.; Kukreja, S.; Goutam, U. Impact of Heat Stress on Potato (Solanum Tuberosum L.): Present Scenario and Future Opportunities. J. Hortic. Sci. Biotechnol. 2020, 95, 407–424. [Google Scholar] [CrossRef]
- Steyn, J.M.; du Plessis, H.F.; Fourie, P.; Hammes, P.S. Yield Response of Potato Genotypes to Different Soil Water Regimes in Contrasting Seasons of a Subtropical Climate. Potato Res. 1998, 41, 239–254. [Google Scholar] [CrossRef]
- Bonierbale, M.W.; Amoros, W.R.; Salas, E.; de Jong, W. Potato Breeding. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Springer: Cham, Switzerland, 2019; pp. 163–217. ISBN 9783030286835. [Google Scholar]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Mangal, V.; Altaf, M.A.; Sharma, S.; Singh, B.; Kumar, M. Insight into Melatonin-Mediated Response and Signaling in the Regulation of Plant Defense under Biotic Stress. Plant Mol. Biol. 2021, 1, 385–399. [Google Scholar] [CrossRef]
- Lal, M.K.; Tiwari, R.K.; Gahlaut, V.; Mangal, V.; Kumar, A.; Singh, M.P.; Paul, V.; Kumar, S.; Singh, B.; Zinta, G. Physiological and Molecular Insights on Wheat Responses to Heat Stress. Plant Cell Rep. 2022, 41, 501–518. [Google Scholar] [CrossRef]
- Lal, M.K.; Sharma, N.; Adavi, S.B.; Sharma, E.; Altaf, M.A.; Tiwari, R.K.; Kumar, R.; Kumar, A.; Dey, A.; Paul, V.; et al. From Source to Sink: Mechanistic Insight of Photoassimilates Synthesis and Partitioning under High Temperature and Elevated [CO2]. Plant Mol. Biol. 2022, 1, 1–20. [Google Scholar] [CrossRef]
- Peng, J.; Manevski, K.; Kørup, K.; Larsen, R.; Zhou, Z.; Andersen, M.N. Environmental Constraints to Net Primary Productivity at Northern Latitudes: A Study across Scales of Radiation Interception and Biomass Production of Potato. Int. J. Appl. Earth Obs. Geoinf. 2021, 94, 102232. [Google Scholar] [CrossRef]
- Ávila-Valdés, A.; Quinet, M.; Lutts, S.; Martínez, J.P.; Lizana, X.C. Tuber Yield and Quality Responses of Potato to Moderate Temperature Increase during Tuber Bulking under Two Water Availability Scenarios. Field Crops Res. 2020, 251, 107786. [Google Scholar] [CrossRef]
- Lizana, X.C.; Avila, A.; Tolaba, A.; Martinez, J.P. Field Responses of Potato to Increased Temperature during Tuber Bulking: Projection for Climate Change Scenarios, at High-Yield Environments of Southern Chile. Agric. Meteorol. 2017, 239, 192–201. [Google Scholar] [CrossRef]
- Obiero, C.O.; Milroy, S.P.; Bell, R.W. Importance of Whole Plant Dry Matter Dynamics for Potato (Solanum Tuberosum L.) Tuber Yield Response to an Episode of High Temperature. Environ. Exp. Bot. 2019, 162, 560–571. [Google Scholar] [CrossRef]
- Haverkort, A.J. Ecology of Potato Cropping Systems in Relation to Latitude and Altitude. Agric. Syst. 1990, 32, 251–272. [Google Scholar] [CrossRef]
- Cantore, V.; Wassar, F.; Yamaç, S.S.; Sellami, M.H.; Albrizio, R.; Stellacci, A.M.; Todorovic, M. Yield and Water Use Efficiency of Early Potato Grown under Different Irrigation Regimes. Int. J. Plant Prod. 2014, 8, 409–428. [Google Scholar] [CrossRef]
- Fleisher, D.H.; Timlin, D.J.; Reddy, V.R. Temperature Influence on Potato Leaf and Branch Distribution and on Canopy Photosynthetic Rate. Agron. J. 2006, 98, 1442–1452. [Google Scholar] [CrossRef]
- Paul, S.; Das, M.K.; Baishya, P.; Ramteke, A.; Farooq, M.; Baroowa, B.; Sunkar, R.; Gogoi, N. Effect of High Temperature on Yield Associated Parameters and Vascular Bundle Development in Five Potato Cultivars. Sci. Hortic. 2017, 225, 134–140. [Google Scholar] [CrossRef]
- Adhikari, U.; Nejadhashemi, A.P.; Woznicki, S.A. Climate Change and Eastern Africa: A Review of Impact on Major Crops. Food Energy Secur. 2015, 4, 110–132. [Google Scholar] [CrossRef]
- Jarvis, A.; Ramirez-Villegas, J.; Campo, B.V.H.; Navarro-Racines, C. Is Cassava the Answer to African Climate Change Adaptation? Trop. Plant Biol. 2012, 5, 9–29. [Google Scholar] [CrossRef]
- Hill, D.; Nelson, D.; Hammond, J.; Bell, L. Morphophysiology of Potato (Solanum Tuberosum) in Response to Drought Stress: Paving the Way Forward. Front. Plant Sci. 2021, 11, 2258. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, R.; Asseng, S.; Robertson, R.; Petsakos, A.; Hoogenboom, G.; Quiroz, R.; Hareau, G.; Wolf, J. Climate Change Impact on Global Potato Production. Eur. J. Agron. 2018, 100, 87–98. [Google Scholar] [CrossRef]
- Wahid, A. Physiological Implications of Metabolite Biosynthesis for Net Assimilation and Heat-Stress Tolerance of Sugarcane (Saccharum Officinarum) Sprouts. J. Plant Res. 2007, 120, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Handayani, T.; Watanabe, K. The Combination of Drought and Heat Stress Has a Greater Effect on Potato Plants than Single Stresses. Plant Soil Environ. 2020, 66, 175–182. [Google Scholar] [CrossRef]
- Muthoni, J.; Shimelis, H. Heat and Drought Stress and Their Implications on Potato Production under Dry African Tropics. Aust J. Crop Sci. 2020, 14, 1405–1414. [Google Scholar] [CrossRef]
- Timlin, D.; Rahman, S.M.L.; Baker, J.; Reddy, V.R.; Fleisher, D.; Quebedeaux, B. Whole Plant Photosynthesis, Development, and Carbon Partitioning in Potato as a Function of Temperature. Agron. J. 2006, 98, 1195–1203. [Google Scholar] [CrossRef]
- Aliche, E.B.; Prusova-Bourke, A.; Ruiz-Sanchez, M.; Oortwijn, M.; Gerkema, E.; van As, H.; Visser, R.G.F.; van der Linden, C.G. Morphological and Physiological Responses of the Potato Stem Transport Tissues to Dehydration Stress. Planta 2020, 251, 45. [Google Scholar] [CrossRef]
- Singh, J.; Pandey, P.; James, D.; Chandrasekhar, K.; Achary, V.M.M.; Kaul, T.; Tripathy, B.C.; Reddy, M.K. Enhancing C3 Photosynthesis: An Outlook on Feasible Interventions for Crop Improvement. Plant Biotechnol. J. 2014, 12, 1217–1230. [Google Scholar] [CrossRef]
- Lafta, A.M.; Lorenzen, J.H. Effect of High Temperature on Plant Growth and Carbohydrate Metabolism in Potato. Plant Physiol. 1995, 109, 637–643. [Google Scholar] [CrossRef]
- Schafleitner, R.; Gutierrez Rosales, R.O.; Gaudin, A.; Alvarado Aliaga, C.A.; Martinez, G.N.; Tincopa Marca, L.R.; Bolivar, L.A.; Delgado, F.M.; Simon, R.; Bonierbale, M. Capturing Candidate Drought Tolerance Traits in Two Native Andean Potato Clones by Transcription Profiling of Field Grown Plants under Water Stress. Plant Physiol. Biochem. 2007, 45, 673–690. [Google Scholar] [CrossRef]
- Hastilestari, B.R.; Lorenz, J.; Reid, S.; Hofmann, J.; Pscheidt, D.; Sonnewald, U.; Sonnewald, S. Deciphering Source and Sink Responses of Potato Plants (Solanum Tuberosum L.) to Elevated Temperatures. Plant Cell Environ. 2018, 41, 2600–2616. [Google Scholar] [CrossRef]
- Paul, S.; Bose, I.; Gogoi, N. Morphophysiological Responses: Criteria for Screening Heat Tolerance in Potato. Curr. Sci. 2016, 111, 1226–1231. [Google Scholar] [CrossRef]
- Monneveux, P.; Ramírez, D.A.; Pino, M.T. Drought Tolerance in Potato (S. Tuberosum L.). Can We Learn from Drought Tolerance Research in Cereals? Plant Sci. 2013, 205–206, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-Wide Analysis of the Potato Hsp20 Gene Family: Identification, Genomic Organization and Expression Profiles in Response to Heat Stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Trapero-Mozos, A.; Morris, W.L.; Ducreux, L.J.M.; McLean, K.; Stephens, J.; Torrance, L.; Bryan, G.J.; Hancock, R.D.; Taylor, M.A. Engineering Heat Tolerance in Potato by Temperature-Dependent Expression of a Specific Allele of HEAT-SHOCK COGNATE 70. Plant Biotechnol. J. 2018, 16, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.T.; Ávila, A.; Molina, A.; Jeknic, Z.; Chen, T.H.H. Enhanced in Vitro Drought Tolerance of Solanum Tuberosum and Solanum Commersonii Plants Overexpressing the ScCBF1 Gene. Cienc. Investig. Agrar. 2013, 40, 171–184. [Google Scholar] [CrossRef]
- Aien, A.; Khetarpal, S.; Singh, M.P. Higher glycinebetaine and antioxidant enzymes activity are associated with high temperature tolerance in potato investigating rice resilience to high Night Temperature and Elevated CO2 Under Warming Climate View Project National Innovations in Climate Res. Indian J. Plant Physiol. 2011, 16, 285. [Google Scholar]
- Saidi, Y.; Finka, A.; Muriset, M.; Bromberg, Z.; Weiss, Y.G.; Maathuis, F.J.M.; Goloubinoff, P. The Heat Shock Response in Moss Plants Is Regulated by Specific Calcium-Permeable Channels in the Plasma Membrane. Plant Cell 2009, 21, 2829–2843. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with Drought: Stress and Adaptive Responses in Potato and Perspectives for Improvement. Front. Plant Sci. 2015, 6, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Farooq, M.; Gogoi, N. Influence of High Temperature on Carbon Assimilation, Enzymatic Antioxidants and Tuber Yield of Different Potato Cultivars. Russ. J. Plant Physiol. 2016, 63, 339–345. [Google Scholar] [CrossRef]
- Minhas, J.S. Potato: Production Strategies under Abiotic Stress. In Improving Crop Resistance to Abiotic Stress; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; Volume 2, pp. 1155–1167. ISBN 9783527328406. [Google Scholar]
- Kumlay, A.M. Combination of the Auxins NAA, IBA, and IAA with GA3 Improves the Commercial Seed-Tuber Production of Potato (Solanum Tuberosum L.) under in Vitro Conditions. Biomed. Res. Int. 2014, 2014, 439259. [Google Scholar] [CrossRef]
- Rolando, J.L.; Ramírez, D.A.; Yactayo, W.; Monneveux, P.; Quiroz, R. Leaf Greenness as a Drought Tolerance Related Trait in Potato (Solanum Tuberosum L.). Environ. Exp. Bot. 2015, 110, 27–35. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Ahmad, P. Role of Mineral Nutrients in Abiotic Stress Tolerance: Revisiting the Associated Signaling Mechanisms. In Plant Signaling Molecules: Role and Regulation under Stressful Environments; Elsevier: Amsterdam, The Netherlands, 2019; pp. 269–285. ISBN 9780128164518. [Google Scholar]
- Joshi, R.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Ectopic Expression of Pokkali Phosphoglycerate Kinase-2 (OsPGK2-P) Improves Yield in Tobacco Plants under Salinity Stress. Plant Cell Rep. 2016, 35, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Nibau, C.; Gibbs, D.J.; Coates, J.C. Branching out in New Directions: The Control of Root Architecture by Lateral Root Formation. New Phytol. 2008, 179, 595–614. [Google Scholar] [CrossRef] [PubMed]
- George, T.S.; Taylor, M.A.; Dodd, I.C.; White, P.J. Climate Change and Consequences for Potato Production: A Review of Tolerance to Emerging Abiotic Stress. Potato Res. 2017, 60, 239–268. [Google Scholar] [CrossRef]
- Viola, R. Roberts a G, Haupt S, Gazzani S, Hancock RD, Marmiroli N, Machray GC, Oparka KJ. Tuberization in Potato Involves a Switch from Apoplastic to Symplastic Phloem Unloading. Plant Cell 2001, 13, 98. [Google Scholar] [CrossRef] [PubMed]
- Ahsan Altaf, M.; Shahid, R.; Kumar, R.; Mohsin Altaf, M.; Kumar, A.; Ullah Khan, L.; Saqib, M.; Azher Nawaz, M.; Saddiq, B.; Bahadur, S.; et al. Phytohormones Mediated Modulation of Abiotic Stress Tolerance and Potential Crosstalk in Horticultural Crops. J. Plant Growth Regul. 2022, 1–27. [Google Scholar] [CrossRef]
- Steward, F.C.; Moreno, U.; Roca, W.M. Supplement 2: Growth, Form and Composition of Potato Plants as Affected by Environment. Ann. Bot. 1981, 1–45. [Google Scholar] [CrossRef]
- Kimball, B.A.; Kobayashi, K.; Bindi, M. Responses of Agricultural Crops to Free-Air CO2 Enrichment. Adv. Agron. 2002, 77, 293–368. [Google Scholar] [CrossRef]
- Abelenda, J.A.; Navarro, C.; Prat, S. Flowering and Tuberization: A Tale of Two Nightshades. Trends Plant Sci. 2014, 19, 115–122. [Google Scholar] [CrossRef]
- Kim, Y.U.; Seo, B.S.; Choi, D.H.; Ban, H.Y.; Lee, B.W. Impact of high temperatures on the marketable tuber yield and related traits of potato. Eur. J. Agron. 2017, 89, 46–52. [Google Scholar] [CrossRef]
- Rykaczewska, K. The Impact of High Temperature during Growing Season on Potato Cultivars with Different Response to Environmental Stresses. Am. J. Plant Sci. 2013, 04, 2386–2393. [Google Scholar] [CrossRef]
- Sarkar, D. The Signal Transduction Pathways Controlling in Planta Tuberization in Potato: An Emerging Synthesis. Plant Cell Rep. 2008, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, J.F.; Virgós-Soler, A.; Prat, S. Control of Photoperiod-Regulated Tuberization in Potato by the Arabidopsis Flowering-Time Gene CONSTANS. Proc. Natl. Acad. Sci. USA 2002, 99, 15211–15216. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Lin, Z.; Huang, L.; Damaris, R.N.; Li, M.; Yang, P. A CONSTANS-LIKE Gene of Nelumbo Nucifera Could Promote Potato Tuberization. Planta 2021, 253, 65. [Google Scholar] [CrossRef] [PubMed]
- Amador, V.; Monte, E.; García-Martínez, J.L.; Prat, S. Gibberellins Signal Nuclear Import of PHOR1, a Photoperiod-Responsive Protein with Homology to Drosophila Armadillo. Cell 2001, 106, 343–354. [Google Scholar] [CrossRef]
- Altaf, M.A.; Sharma, N.; Singh, J.; Samota, M.K.; Sankhyan, P.; Singh, B.; Kumar, A.; Naz, S.; Lal, M.K.; Tiwari, R.K.; et al. Mechanistic Insights on Melatonin-Mediated Plant Growth Regulation and Hormonal Cross-Talk Process in Solanaceous Vegetables. Sci. Hortic. 2023, 308, 111570. [Google Scholar] [CrossRef]
- Lin, L.; Zheng, Z.; Hua, T.; Ashraf, U.; Hamoud, Y.A.; Alaa, A.A.; Xiangru, T.; Meiyang, D.; Zaiman, W.; Shenggang, P. Nitrogen Deep Placement Combined with Straw Mulch Cultivation Enhances Physiological Traits, Grain Yield and Nitrogen Use Efficiency in Mechanical Pot-Seedling Transplanting Rice. Rice Sci. 2022, 29, 89–100. [Google Scholar] [CrossRef]
- Abeytilakarathna, P.D. Factors Affect to Stolon Formation and Tuberization in Potato: A Review. Agric. Rev. 2021, 43, 91–97. [Google Scholar] [CrossRef]
- Wolf, S.; Olesinski, A.A.; Rudich, J.; Marani, A. Effect of High Temperature on Photosynthesis in Potatoes. Ann. Bot. 1990, 65, 179–185. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E. Cracking and Fracture Properties of Potato (Solanum Tuberosum L.) Tubers and Their Relation to Dry Matter, Starch, and Mineral Distribution. J. Sci. Food Agric. 2019, 99, 3149–3156. [Google Scholar] [CrossRef] [PubMed]
- Appeldoorn, N.J.G.; de Bruijn, S.M.; Koot-Gronsveld, E.A.M.; Visser, R.G.F.; Vreugdenhil, D.; van der Plas, L.H.W. Developmental Changes of Enzymes Involved in Conversion of Sucrose to Hexose-Phosphate during Early Tuberisation of Potato. Planta 1997, 202, 220–226. [Google Scholar] [CrossRef]
- Salvucci, M.E.; Crafts-Brandner, S.J. Mechanism for Deactivation of Rubisco under Moderate Heat Stress. Physiol. Plant 2004, 122, 513–519. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, Biochemical, and Molecular Mechanisms of Heat Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Howarth, C.J. Genetic Improvements of Tolerance to High Temperature. In Abiotic Stresses: Plant Resistance through Breeding and Molecular Approaches; Ashraf, M., Harris, P.J.C., Eds.; Haworth Press Inc.: New York, NY, USA, 2005; pp. 277–300. [Google Scholar]
- Cao, Z.; Yao, X.; Liu, H.; Liu, B.; Cheng, T.; Tian, Y.; Cao, W.; Zhu, Y. Comparison of the Abilities of Vegetation Indices and Photosynthetic Parameters to Detect Heat Stress in Wheat. Agric. Meteorol. 2019, 265, 121–136. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Q.; Ci, D.; Shao, X.; Zhang, D. Effects of High Temperature on Photosynthesis and Related Gene Expression in Poplar. BMC Plant Biol. 2014, 14, 111. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.S.; Kjaer, K.H.; Rosenqvist, E.; Sharma, D.K.; Ottosen, C.O. Heat Stress and Recovery of Photosystem II Efficiency in Wheat (Triticum Aestivum L.) Cultivars Acclimated to Different Growth Temperatures. Environ. Exp. Bot. 2014, 99, 1–8. [Google Scholar] [CrossRef]
- Nölke, G.; Houdelet, M.; Kreuzaler, F.; Peterhänsel, C.; Schillberg, S. The Expression of a Recombinant Glycolate Dehydrogenase Polyprotein in Potato (Solanum Tuberosum) Plastids Strongly Enhances Photosynthesis and Tuber Yield. Plant Biotechnol. J. 2014, 12, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Aliche, E.B.; Theeuwen, T.P.J.M.; Oortwijn, M.; Visser, R.G.F.; van der Linden, C.G. Carbon Partitioning Mechanisms in POTATO under Drought Stress. Plant Physiol. Biochem. 2020, 146, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Da Ros, L.; Elferjani, R.; Soolanayakanahally, R.; Kagale, S.; Pahari, S.; Kulkarni, M.; Wahab, J.; Bizimungu, B. Drought-Induced Regulatory Cascades and Their Effects on the Nutritional Quality of Developing Potato Tubers. Genes 2020, 11, 864. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Ahmad, S.; Abbas, G.; Fatima, Z.; Hussain, S.; Ahmed, M.; Khan, M.A.; Khan, A.; Fahad, S.; Nasim, W.; et al. Modeling the Impact of Climate Warming on Potato Phenology. Eur. J. Agron. 2022, 132, 126404. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Mushtaq, A.; Ali, S.; Muhammad, H.M.D.; Saddiq, B.; Ahmad, R.; Zulfiqar, F.; Hayat, F.; Tiwari, R.K.; Lal, M.K.; et al. Foliar Application of Ascorbic Acid Enhances Growth and Yield of Lettuce (Lactuca Sativa) under Saline Conditions by Improving Antioxidant Defence Mechanism. Funct. Plant Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Nandy, S.; Mandal, S.; Gupta, S.K.; Anand, U.; Ghorai, M.; Mundhra, A.; Rahman, M.d.H.; Ray, P.; Mitra, S.; Ray, D.; et al. Role of Polyamines in Molecular Regulation and Cross-Talks Against Drought Tolerance in Plants. J. Plant Growth Regul. 2022, 1–17. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Kumar, R.; Lal, M.K.; Kumar, A.; Altaf, M.A.; Devi, R.; Mangal, V.; Naz, S.; Altaf, M.M.; Dey, A.; et al. Melatonin-Polyamine Interplay in the Regulation of Stress Responses in Plants. J. Plant Growth Regul. 2022, 1–17. [Google Scholar] [CrossRef]
- Ahmed, S.; Zhou, X.; Pang, Y.; Jin, L.; Bao, J. Improving Starch-Related Traits in Potato Crops: Achievements and Future Challenges; Wiley-Blackwell: Hoboken, NJ, USA, 2018; Volume 70, ISBN 8657186971932. [Google Scholar]
- Kumar, D.; Ezekiel, R. Developmental Changes in Sugars and Dry Matter Content of Potato Tuber under Sub-Tropical Climates. Sci. Hortic. 2006, 110, 129–134. [Google Scholar] [CrossRef]
- Edwards, A.; Fulton, D.C.; Hylton, C.M.; Jobling, S.A.; Gidley, M.; Rössner, U.; Martin, C.; Smith, A.M. A Combined Reduction in Activity of Starch Synthases II and III of Potato Has Novel Effects on the Starch of Tubers. Plant J. 1999, 17, 251–261. [Google Scholar] [CrossRef]
- Lovell, P.H.; Booth, A. Effects of Gibberellic Acid on Growth, Tuber Formation and Carbohydrate Distribution in Solanum Tuberosum. New Phytol. 1967, 66, 525–537. [Google Scholar] [CrossRef]
- Huber, S.C.; Israel, D.W. Biochemical Basis for Partitioning of Photosynthetically Fixed Carbon between Starch and Sucrose in Soybean (Glycine Max Merr.) Leaves. Plant Physiol. 1982, 69, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Shahid, R.; Altaf, M.M.; Kumar, R.; Naz, S.; Kumar, A.; Alam, P.; Tiwari, R.K.; Lal, M.K.; Ahmad, P. Melatonin: First-Line Soldier in Tomato under Abiotic Stress Current and Future Perspective. Plant Physiol. Biochem. 2022, 185, 188–197. [Google Scholar] [CrossRef]
- Liu, B.; Kong, L.; Zhang, Y.; Liao, Y. Gene and Metabolite Integration Analysis through Transcriptome and Metabolome Brings New Insight into Heat Stress Tolerance in Potato (Solanum Tuberosum L.). Plants 2021, 10, 103. [Google Scholar] [CrossRef]
- Dimenstein, L.; Lisker, N.; Kedar, N.; Levy, D. Changes in the Content of Steroidal Glycoalkaloids in Potato Tubers Grown in the Field and in the Greenhouse under Different Conditions of Light, Temperature and Daylength. Physiol. Mol. Plant Pathol. 1997, 50, 391–402. [Google Scholar] [CrossRef]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and Physiological Responses of Potato (Solanum Tuberosum L) Plantlets to Gradient Saline Stress. Front. Plant Sci. 2015, 5, 787. [Google Scholar] [CrossRef]
- Demirel, U.; Morris, W.L.; Ducreux, L.J.M.; Yavuz, C.; Asim, A.; Tindas, I.; Campbell, R.; Morris, J.A.; Verrall, S.R.; Hedley, P.E.; et al. Physiological, Biochemical, and Transcriptional Responses to Single and Combined Abiotic Stress in Stress-Tolerant and Stress-Sensitive Potato Genotypes. Front. Plant Sci. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Ranjeet, R. Kumar Protection against Heat Stress in Wheat Involves Change in Cell Membrane Stability, Antioxidant Enzymes, Osmolyte, H2O2 and Transcript of Heat Shock Protein. Int. J. Plant Physiol. Biochem. 2012, 4, 83–91. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst Phytohormones from Planta and PGPR under Biotic and Abiotic Stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Sakamoto, A.; Murata, N. The Role of Glycine Betaine in the Protection of Plants from Stress: Clues from Transgenic Plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a Multifaceted Signalling Molecule in Plant Responses to Abiotic Stress: Understanding the Physiological Mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- Devi, R.; Behera, B.; Raza, M.B.; Mangal, V.; Altaf, M.A.; Kumar, R.; Kumar, A.; Tiwari, R.K.; Lal, M.K.; Singh, B. An Insight into Microbes Mediated Heavy Metal Detoxification in Plants: A Review. J. Soil Sci. Plant Nutr. 2021, 41, 1–23. [Google Scholar] [CrossRef]
- Park, C.J.; Seo, Y.S. Heat Shock Proteins: A Review of the Molecular Chaperones for Plant Immunity. Plant Pathol. J. 2015, 31, 323. [Google Scholar] [CrossRef]
- Al-Whaibi, M.H. Plant Heat-Shock Proteins: A Mini Review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Lee, S.; Kang, H.; Kim, Y. Performance Optimization of a Hybrid Cooler Combining Vapor Compression and Natural Circulation Cycles. Int. J. Refrig. 2009, 32, 800–808. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of Plant Heat-Shock Proteins and Molecular Chaperones in the Abiotic Stress Response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Li, W.; Chen, Y.; Ye, M.; Wang, D.; Chen, Q. Evolutionary History of the Heat Shock Protein 90 (Hsp90) Family of 43 Plants and Characterization of Hsp90s in Solanum Tuberosum. Mol. Biol. Rep. 2020, 47, 6679–6691. [Google Scholar] [CrossRef]
- Savić, J.; Dragićević, I.; Pantelić, D.; Oljača, J.; Momčilović, I. Expression of Small Heat Shock Proteins and Heat Tolerance in Potato (Solanum Tuberosum L.). Arch. Biol. Sci. 2012, 64, 135–144. [Google Scholar] [CrossRef]
- Rudić, J.; Pantelić, D.; Oljača, J.; Momčilović, I. Effects of Elevated Temperature and Salicylic Acid on Heat Shock Response and Growth of Potato Microplants. Horticulturae 2022, 8, 372. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant Responses of Wheat Plants under Stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant Enzymes Regulation in Plants in Reference to Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Olson, K.R. Reactive Oxygen Species or Reactive Sulfur Species: Why We Should Consider the Latter. J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef] [PubMed]
- Gruhlke, M.C.H.; Slusarenko, A.J. The Biology of Reactive Sulfur Species (RSS). Plant Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic Insights on Melatonin-Mediated Drought Stress Mitigation in Plants. Physiol. Plant 2021, 172, 1212–1226. [Google Scholar] [CrossRef]
- Paul, S.; Gogoi, N.; Sarma, B.; Baroowa, B. Biochemical Changes in Potato under Elevated Temperature. Indian J. Plant Physiol. 2014, 19, 36–42. [Google Scholar] [CrossRef]
- Arora, D.; Jain, P.; Singh, N.; Kaur, H.; Bhatla, S.C. Mechanisms of Nitric Oxide Crosstalk with Reactive Oxygen Species Scavenging Enzymes during Abiotic Stress Tolerance in Plants. Free Radic. Res. 2016, 50, 291–303. [Google Scholar] [CrossRef]
- Shin, D.; Moon, S.J.; Han, S.; Kim, B.G.; Park, S.R.; Lee, S.K.; Yoon, H.J.; Lee, H.E.; bin Kwon, H.; Baek, D.; et al. Expression of StMYB1R-1, a Novel Potato Single MYB-like Domain Transcription Factor, Increases Drought Tolerance. Plant Physiol. 2011, 155, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Haverkort, A.J.; Franke, A.C.; Steyn, J.M.; Pronk, A.A.; Caldiz, D.O.; Kooman, P.L. A Robust Potato Model: LINTUL-POTATO-DSS. Potato Res. 2015, 58, 313–327. [Google Scholar] [CrossRef]
- Kooman, P.L.; Haverkort, A.J. Modelling Development and Growth of the Potato Crop Influenced by Temperature and Daylength: LINTUL-POTATO.LINTUL-POTATO. In Potato Ecology and Modelling of Crops under Conditions Limiting Growth, Proceedings of the Second International Potato Modeling Conference, Wageningen, The Netherlands, 17–19 May 1994; Haverkort, A.J., MacKerron, D.K.L., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 41–59. [Google Scholar] [CrossRef]
- Griffin, T.S.; Johnson, B.S.; Ritchie, J.T. A Simulation Model for Potato Growth and Development: Substor-Potato Version 2.0; Michigan State University, Department of Crop and Soil Sciences: East Lansing, MI, USA, 1993. [Google Scholar]
- Goswami, B.; Hussain, R.; Kumar, P.; Saikia, U.; Banarjee, S. Impact assessment of climate change on potato productivity in Assam using SUBSTOR-Potato model. J. Agrometeorol. 2018, 20, 105–109. [Google Scholar] [CrossRef]
- Kumar, R.; Kaundal, P.; Tiwari, R.K.; Siddappa, S.; Kumari, H.; Lal, M.K.; Naga, K.C.; Sharma, S.; Sagar, V.; Kumar, M. Establishment of a One-Step Reverse Transcription Recombinase Polymerase Amplification Assay for the Detection of Potato Virus S. J. Virol. Methods 2022, 307, 114568. [Google Scholar] [CrossRef]
- Struik, P.C. Responses of the Potato Plant to Temperature. Potato Biol. Biotechnol. Adv. Perspect. 2007, 367–393. [Google Scholar] [CrossRef]
- Sharma, E.K.; Tiwari, R.K.; Lal, M.K.; Sharma, E.; Tiwari, R.K.; Devi, R.; Mishra, U.N.; Thakur, R.; Gupta, R.; Dey, A.; et al. Nutrient-Mediated Perception and Signalling in Human Metabolism: A Perspective of Nutrigenomics. Int. J. Mol. Sci. 2022, 23, 11305. [Google Scholar] [CrossRef]
- Rykaczewska, K. The Effect of High Temperature Occurring in Subsequent Stages of Plant Development on Potato Yield and Tuber Physiological Defects. Am. J. Potato Res. 2015, 92, 339–349. [Google Scholar] [CrossRef]
- Boguszewska-Mańkowska, D.; Zarzyńska, K.; Nosalewicz, A. Drought Differentially Affects Root System Size and Architecture of Potato Cultivars with Differing Drought Tolerance. Am. J. Potato Res. 2020, 97, 54–62. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Verhagen, A. Climate Change and Its Repercussions for the Potato Supply Chain. Potato Res. 2008, 51, 223–237. [Google Scholar] [CrossRef]
- Smillie, R.M.; Hetherington, S.E.; Ochoa, C.; Malagamba, P. Tolerances of Wild Potato Species from Different Altitudes to Cold and Heat. Planta 1983, 159, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Naga, K.C.; Subhash, S.; Sagar, V. Continuous and Emerging Challenges of Silver Scurf Disease in Potato. Int. J. Pest Manag. 2020, 1–13. [Google Scholar] [CrossRef]
- Guedes, M.L.; Haynes, K.G.; Vinyard, B.T.; Pinto, C.A.B.P. Heat Tolerance in Diploid Wild Potato Species In Vitro. Am. J. Potato Res. 2019, 96, 294–302. [Google Scholar] [CrossRef]
- Granado-Lorencio, F.; Olmedilla-Alonso, B.; Herrero-Barbudo, C.; Pérez-Sacristán, B.; Blanco-Navarro, I.; Blázquez-García, S. Comparative In Vitro Bioaccessibility of Carotenoids from Relevant Contributors to Carotenoid Intake. J. Agric. Food Chem. 2007, 55, 6387–6394. [Google Scholar] [CrossRef] [PubMed]
- Navarro, C.; Abelenda, J.A.; Cruz-Oró, E.; Cuéllar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of Flowering and Storage Organ Formation in Potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef]
- Eltayeb, A.E.; Yamamoto, S.; Habora, M.E.E.; Yin, L.; Tsujimoto, H.; Tanaka, K. Transgenic Potato Overexpressing Arabidopsis Cytosolic AtDHAR1 Showed Higher Tolerance to Herbicide, Drought and Salt Stresses. Breed Sci. 2011, 61, 3–10. [Google Scholar] [CrossRef]
- Tiburcio, A.F.; Altabella, T.; Bitrián, M.; Alcázar, R. The Roles of Polyamines during the Lifespan of Plants: From Development to Stress. Planta 2014, 240, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lehretz, G.G.; Sonnewald, S.; Hornyik, C.; Corral, J.M.; Sonnewald, U. Post-Transcriptional Regulation of FLOWERING LOCUS T Modulates Heat-Dependent Source-Sink Development in Potato. Curr. Biol. 2019, 29, 1614–1624.e3. [Google Scholar] [CrossRef] [PubMed]
- Abelenda, J.A.; Cruz-Oró, E.; Franco-Zorrilla, J.M.; Prat, S. Potato StCONSTANS-Like1 Suppresses Storage Organ Formation by Directly Activating the FT-like StSP5G Repressor. Curr. Biol. 2016, 26, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene Networks Involved in Drought Stress Response and Tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Luan, S. ABA Signal Transduction at the Crossroad of Biotic and Abiotic Stress Responses. Plant Cell Environ. 2012, 35, 53–60. [Google Scholar] [CrossRef]
- Kloosterman, B.; Abelenda, J.A.; Gomez, M.D.M.C.; Oortwijn, M.; de Boer, J.M.; Kowitwanich, K.; Horvath, B.M.; van Eck, H.J.; Smaczniak, C.; Prat, S.; et al. Naturally Occurring Allele Diversity Allows Potato Cultivation in Northern Latitudes. Nature 2013, 495, 246–250. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Kim, D.; Bae, S.; Park, J.; Kim, E.; Kim, S.; Yu, H.R.; Hwang, J.; Kim, J.-I.; Kim, J.S. Digenome-Seq: Genome-Wide Profiling of CRISPR-Cas9 off-Target Effects in Human Cells. Nat. Methods 2015, 12, 237–243. [Google Scholar] [CrossRef]
- Puchta, H. The Repair of Double-Strand Breaks in Plants: Mechanisms and Consequences for Genome Evolution. J. Exp. Bot. 2005, 56, 1–14. [Google Scholar] [CrossRef]
- Mishra, R.; Joshi, R.K.; Zhao, K. Base Editing in Crops: Current Advances, Limitations and Future Implications. Plant Biotechnol. J. 2020, 18, 20–31. [Google Scholar] [CrossRef]
- Nishida, K.; Arazoe, T.; Yachie, N.; Banno, S.; Kakimoto, M.; Tabata, M.; Mochizuki, M.; Miyabe, A.; Araki, M.; Hara, K.Y.; et al. Targeted Nucleotide Editing Using Hybrid Prokaryotic and Vertebrate Adaptive Immune Systems. Science 2016, 353, aaf8729. [Google Scholar] [CrossRef] [PubMed]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable Base Editing of T to G C in Genomic DNA without DNA Cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Bánfalvi, Z.; Csákvári, E.; Villányi, V.; Kondrák, M. Generation of Transgene-Free PDS Mutants in Potato by Agrobacterium-Mediated Transformation. BMC Biotechnol. 2020, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Sevestre, F.; Facon, M.; Wattebled, F.; Szydlowski, N. Facilitating Gene Editing in Potato: A Single-Nucleotide Polymorphism (SNP) Map of the Solanum Tuberosum L. Cv. Desiree Genome. Sci. Rep. 2020, 10, 2045. [Google Scholar] [CrossRef]
- Zheng, Z.; Ye, G.; Zhou, Y.; Pu, X.; Su, W.; Wang, J. Editing Sterol Side Chain Reductase 2 Gene (StSSR2) via CRISPR/Cas9 Reduces the Total Steroidal Glycoalkaloids in Potato. All Life 2021, 14, 401–413. [Google Scholar] [CrossRef]
- Veillet, F.; Kermarrec, M.P.; Chauvin, L.; Chauvin, J.E.; Nogué, F. CRISPR-Induced Indels and Base Editing Using the Staphylococcus Aureus Cas9 in Potato. PLoS ONE 2020, 15, e0235942. [Google Scholar] [CrossRef] [PubMed]
- Veillet, F.; Chauvin, L.; Kermarrec, M.P.; Sevestre, F.; Merrer, M.; Terret, Z.; Szydlowski, N.; Devaux, P.; Gallois, J.L.; Chauvin, J.E. The Solanum Tuberosum GBSSI Gene: A Target for Assessing Gene and Base Editing in Tetraploid Potato. Plant Cell Rep. 2019, 38, 1065–1080. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kaundal, P.; Arjunan, J.; Sharma, S.; Chakrabarti, S.K. Development of a Visual Detection Method for Potato Virus S by Reverse Transcription Loop-Mediated Isothermal Amplification. 3 Biotech 2020, 10, 213. [Google Scholar] [CrossRef] [PubMed]
- Uranga, M.; Aragonés, V.; Selma, S.; Vázquez-Vilar, M.; Orzáez, D.; Daròs, J.A. Efficient Cas9 Multiplex Editing Using Unspaced SgRNA Arrays Engineering in a Potato Virus X Vector. Plant J. 2021, 106, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kaundal, P.; Tiwari, R.K.; Siddappa, S.; Kumari, H.; Chandra Naga, K.; Sharma, S.; Kumar, M. Rapid and Sensitive Detection of Potato Virus X by One-Step Reverse Transcription-Recombinase Polymerase Amplification Method in Potato Leaves and Dormant Tubers. Mol. Cell. Probes 2021, 58, 101743. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, D.; Jogam, P.; Allini, V.R.; Abbagani, S.; Alok, A. The Present and Potential Future Methods for Delivering CRISPR/Cas9 Components in Plants. J. Genet. Eng. Biotechnol. 2020, 18, 25. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.H.; Gaoa, Y.K.; Yuan, L.; Zhang, Q.X. Plant Genome Editing Made Efficient and Easy: Targeted Mutagenesis Using the CRISPR/Cas System. Acta Hortic. 2017, 1185, 209–213. [Google Scholar] [CrossRef]
- Koltun, A.; Corte, L.E.D.; Mertz-Henning, L.M.; Gonçalves, L.S.A. Genetic Improvement of Horticultural Crops Mediated by CRISPR/Cas: A New Horizon of Possibilities. Hortic. Bras. 2018, 36, 290–298. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Response to Tropicalization (Heat/Drought Stress) | Reference |
|---|---|---|
| Yield and total biomass production | Reduction in both yield and biomass | [46] |
| Root growth | Root growth was stimulated under drought stress | [46] |
| Photosynthesis | Reduction in the PSII efficiency and ultimately reduction in photosynthesis | [40,47] |
| Clorophyll and carotenoid content of leaves and tubers | Significant reduction in both chlorophyll and carotenoid content | [48] |
| Membrane stability index (MSI) and cell membrane stability (CMS) | Reduction in membrane stability index under heat and drought stress | [40,48] |
| Relative water content (RWC) | Reduced under both heat and water-deficient conditions | [40] |
| Water potential | Significant reduction in water potential | [49] |
| Water use efficiency (WUE) | Reduction in WUE mainly under drought conditions | [49] |
| Tuber bulking | Reduce dry matter partitioning, reduce tuber filing, increase the production of secondary tubers, increased russeting and cracking | [21] |
| Tuberization | High temperature reduces tuberization | [16] |
| HSP20 | ATP-independent molecular chaperones inhibit the irreversible aggregation of denaturing proteins, thus enhancing the thermotolerance of the plant | [50] |
| Starch synthesis | Heat leads to the production of reactive oxygen species that interfere with starch synthesis | [51] |
| Sucrose synthesis | A decline in sucrose content | [47] |
| Proline content | Acts as osmotic agent, protecting plant cells from dehydration | [52] |
| Glycine betaine | Acts as osmoprotectant in potato under heat and drought stress | [53] |
| Sucrose synthase | Degradation of sucrose into hexoses | [47] |
| Superoxide dismutase | Scavenges superoxide molecule under stress conditions and prevents oxidative damage in potato | [53] |
| Ascorbate peroxidase | Activity increased under heat and drought stress and provides tolerance in potato | [53] |
| Genes Expressed | Role of Gene and Molecular Response | Reference |
|---|---|---|
| DHAR1 | Synthesis of ascorbic acid, which acts as a strong antioxidant, protecting chlorophyll against degradation, and allowing faster removal of H2O2 | [142] |
| DREB1B | TF involved in enhancing drought tolerance | [49] |
| ScCBF1 | The CBFs bind to the cold/dehydration-responsive regulatory motif (CRT/DRE) and are present in the promoter of many drought and cold-responsive genes, such as those associated with osmoprotectant, cold-responsive (COR), and late embryogenesis-abundant (LEA) proteins | [52] |
| HSPs | Molecular chaperones prevent denaturing and aggregation of proteins under heat stress | [47] |
| SAM-DC | Mediates the polyamines, spermidine, and spermine involved in providing tolerance against stress | [143] |
| StSP6A | Master regulator of tuberization and is significantly affected by temperature | [16,47,144] |
| StCOL1 | Suppresses tuber formation by activation of FT-like StSP5G repressor | [145] |
| ABRE and DRE/CRT | The promoters of drought-tolerant gene. ABRE is a major cis-acting element in ABA-responsive gene expression. | [146] |
| LEA5 | Provides tolerance against drought stress | [46] |
| MYB96 | Involved in ABA and auxin cross-talk, increasing lateral root formation | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lal, M.K.; Tiwari, R.K.; Kumar, A.; Dey, A.; Kumar, R.; Kumar, D.; Jaiswal, A.; Changan, S.S.; Raigond, P.; Dutt, S.; et al. Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress. Plants 2022, 11, 2857. https://doi.org/10.3390/plants11212857
Lal MK, Tiwari RK, Kumar A, Dey A, Kumar R, Kumar D, Jaiswal A, Changan SS, Raigond P, Dutt S, et al. Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress. Plants. 2022; 11(21):2857. https://doi.org/10.3390/plants11212857
Chicago/Turabian StyleLal, Milan Kumar, Rahul Kumar Tiwari, Awadhesh Kumar, Abhijit Dey, Ravinder Kumar, Dharmendra Kumar, Arvind Jaiswal, Sushil Sudhakar Changan, Pinky Raigond, Som Dutt, and et al. 2022. "Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress" Plants 11, no. 21: 2857. https://doi.org/10.3390/plants11212857
APA StyleLal, M. K., Tiwari, R. K., Kumar, A., Dey, A., Kumar, R., Kumar, D., Jaiswal, A., Changan, S. S., Raigond, P., Dutt, S., Luthra, S. K., Mandal, S., Singh, M. P., Paul, V., & Singh, B. (2022). Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress. Plants, 11(21), 2857. https://doi.org/10.3390/plants11212857