
Home Range and Movement Patterns of Reintroduced White Lions (Panthera leo melanochaita) in the Kruger to Canyons Biosphere Reserve, South Africa



Abstract
Simple Summary
Abstract
1. Introduction
1.1. Home Range and Movements of Lions
1.2. Home Range and Movement Patterns of White Lions Compared to Tawny Lions
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Sampling
2.3. Data Analysis and Statistical Testing
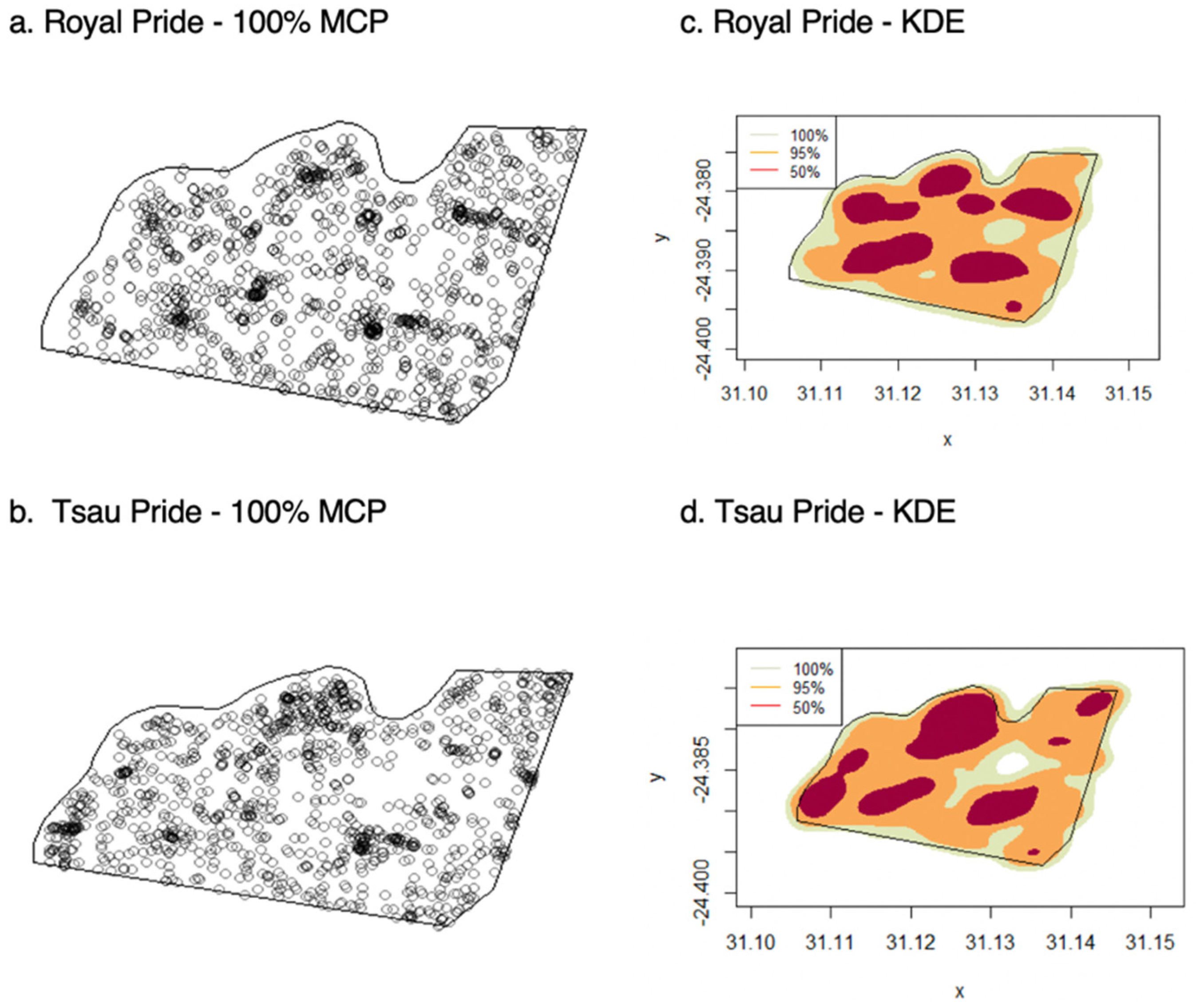
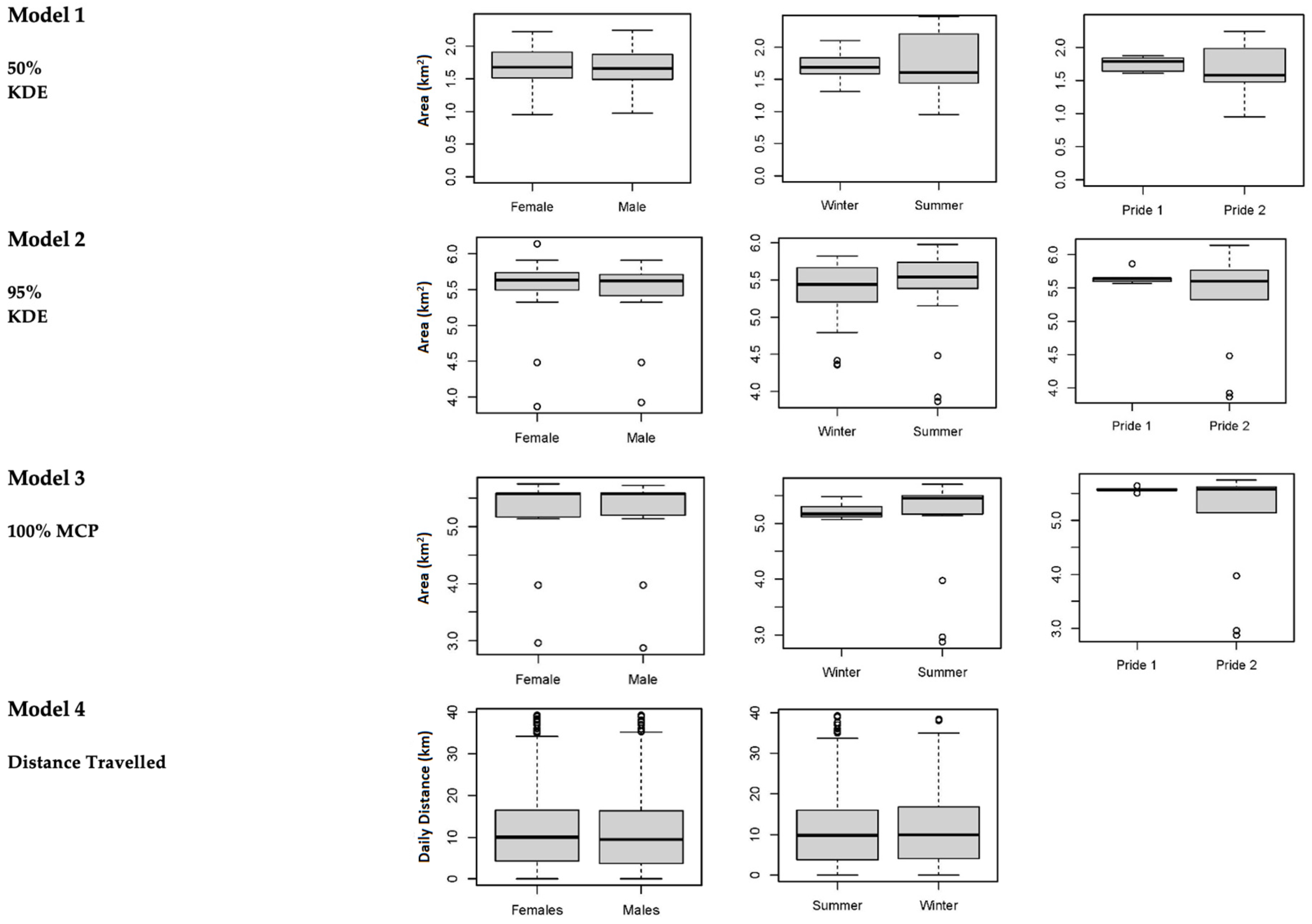
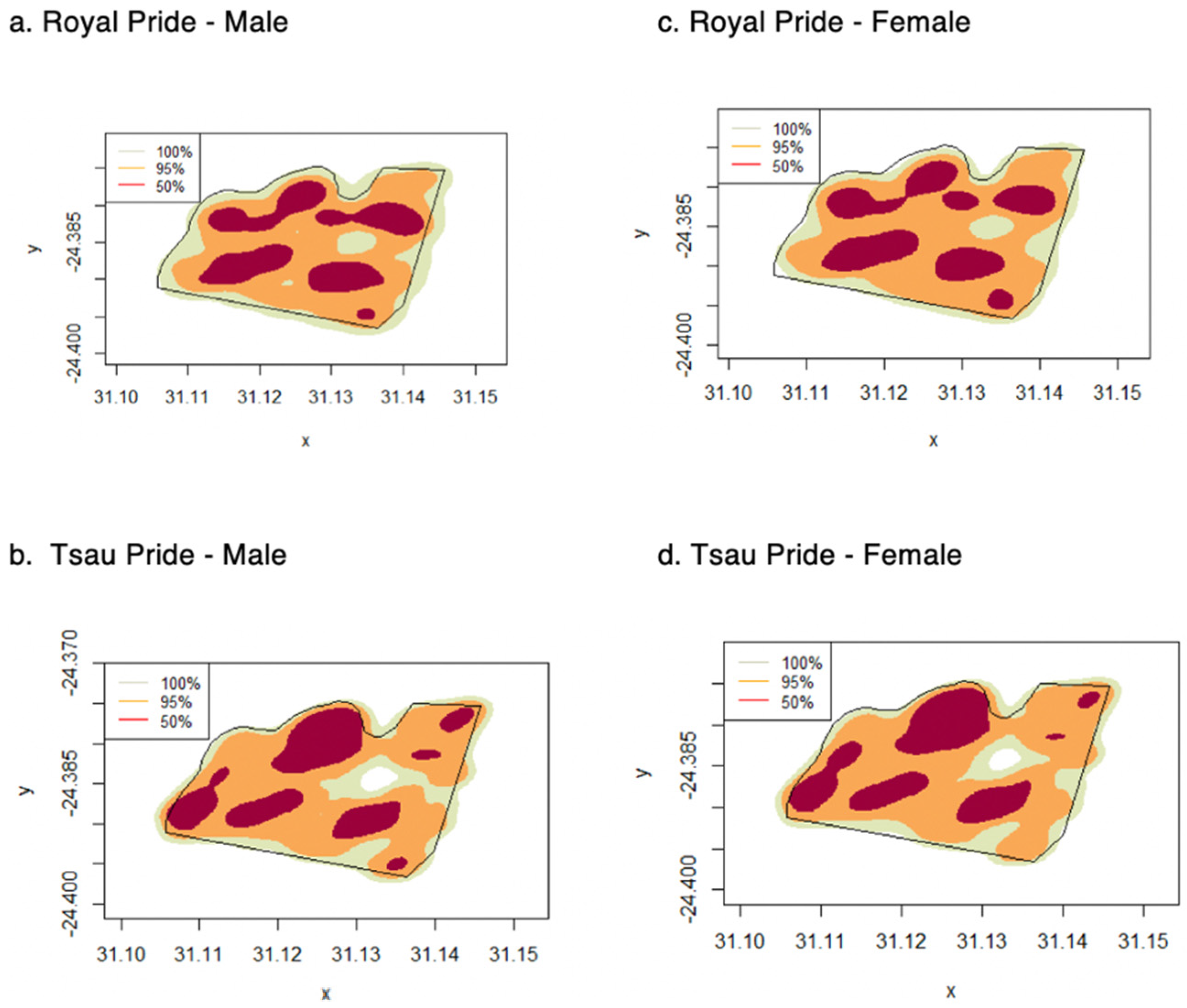
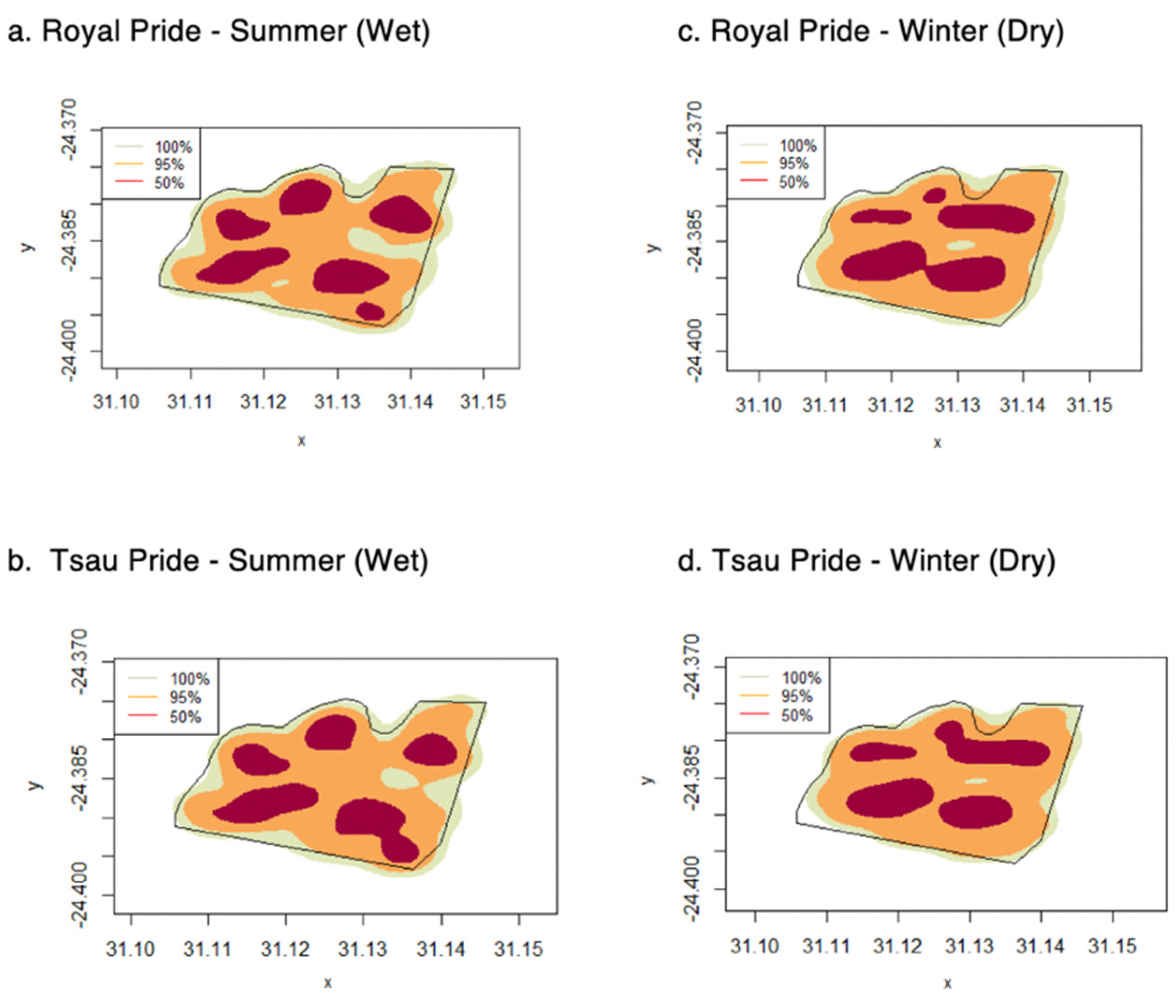
3. Results
3.1. Home Ranges and Movements
3.2. Habitat Selection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baillie, J.; Collen, B.; Griffiths, J.; Loh, J.; Turvey, S. Evolution Lost: Status and Trends of the World’s Vertebrates; Zoological Society of London: London, UK, 2010. [Google Scholar]
- Hunter, L.T.B.; Pretorius, K.; Carlisle, L.C.; Rickelton, M.; Walker, C.; Slotow, R.; Skinner, J.D. Restoring lions Panthera leo to northern KwaZulu-Natal, South Africa: Short-term biological and technical success but equivocal long-term conservation. Oryx 2007, 41, 196–204. [Google Scholar] [CrossRef]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed]
- Woodroffe, R. Predators and people: Using human densities to interpret decline of large carnivores. Anim. Conserv. 2000, 3, 165–173. [Google Scholar] [CrossRef]
- Cardillo, M.; Purvis, A.; Sechrest, W.; Gittleman, J.L.; Biebly, J.; Mace, G.M. Human population density and extinction risk in the world’s carnivores. PLoS Biol. 2004, 2, 909–914. [Google Scholar] [CrossRef]
- Power, J.R. Evaluating how many lions a small reserve can sustain. S. Afr. J. Wildl. Res. 2003, 33, 3–11. [Google Scholar]
- Linnaeus, C. The system of nature through three kingdoms of nature, according to the classes, orders, genera, and species with characters, differences, synonyms, and places. In Systema Naturae, 10th ed.; Reform. Holmiae.; Linnean Society of London: London, UK, 1758; Volume 1, pp. 1–4. [Google Scholar]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN Cat Specialist Group. Cat News Spec. Issue 2016, 11, 80. [Google Scholar]
- Yamaguchi, N.; Kitchener, A.; Driscoll, C.; Macdonald, D. Divided infraorbital foramen in the lion (Panthera leo): Its implications for colonisation history, population bottlenecks, and conservation of the Asian lion (P. l. persica). Contrib. Zool. 2009, 78, 77–83. [Google Scholar] [CrossRef]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; IUCN/SSC: Gland, Switzerland, 1996; 382p. [Google Scholar]
- Miller, S.M.; Harper, C.K.; Bloomer, P.; Hofmeyr, J.; Funston, P.J. Fenced and fragmented: Conservation value of managed metapopulations. PLoS ONE 2015, 10, e0144605. [Google Scholar] [CrossRef]
- Miller, S.; Riggio, J.; Funston, P.; Power, R.J.; Williams, V.; Child, M.F. A Conservation Assessment of Panthera leo. In The Red List of Mammals of South Africa, Swaziland and Lesotho; Child, M.F., Roxburgh, L., Do Linh San, E., Raimondo, D., Davies-Mostert, H.T., Eds.; South African National Biodiversity Institute: Pretoria, South Africa; Endangered Wildlife Trust: Midrand, South Africa, 2016; p. 48. [Google Scholar]
- Ferreira, S.M.; Funston, P.J. Estimating lion population variables: Prey and disease effects in Kruger National Park, South Africa. Wildl. Res. 2010, 37, 194–206. [Google Scholar] [CrossRef]
- Castley, J.G.; Knight, M.H.; Mills, M.G.L.; Thouless, C. Estimation of the lion (Panthera leo) population in the southwestern Kgalagadi Transfrontier Park using a capture-recapture survey. Afr. Zool. 2002, 37, 27–34. [Google Scholar] [CrossRef][Green Version]
- Funston, P.J. Population characteristics of lions (Panthera leo) in the Kgalagadi Transfrontier Park. S. Afr. J. Wildl. Res. 2011, 41, 1–10. [Google Scholar] [CrossRef]
- Funston, P.J. Greater Mapungubwe TFCA Predator Management Plan; Unpublished Report; Shashe Limpopo Predator Research Group: Bulawayo, Zimbabwe, 2010. [Google Scholar]
- Funston, P.J.; Levendal, M. Draft Biodiversity Management Plan for the Lion (Panthera leo) in South Africa. 2014. Available online: https://www.gov.za/sites/default/files/gcis_document/201512/39468gen1190.pdf (accessed on 27 January 2022).
- Miller, S.M.; Funston, P.J. Rapid growth rates of lion (Panthera leo) populations in small, fenced reserves in South Africa: A management dilemma. S. Afr. J. Wildl. Res. 2014, 44, 43–55. [Google Scholar] [CrossRef]
- Tucker, L. Mystery of the White Lions: Children of the Sun God; Npenvu Press: White River, South Africa, 2003; p. 352. [Google Scholar]
- Robinson, R.; De Vos, V. Chinchilla mutant in the lion. Genetica 1982, 60, 61–63. [Google Scholar] [CrossRef]
- Cruickshank, K.M.; Robinson, T.J. Inheritance of the White Coat Colour Phenotype in African Lions (Panthera leo). In Proceedings of a Symposium on Lions and Leopards as Game Ranch Animals; Van Heerden, J., Ed.; Wildlife Group of the South African Veterinary Association: Pretoria, South Africa, 1997; pp. 92–95. [Google Scholar]
- Cho, Y.S.; Hu, L.; Hou, H.; Lee, H.; Xu, J.; Kwon, S.; Oh, S.; Kim, H.-M.; Jho, S.; Kim, S.; et al. The tiger genome and comparative analysis with lion and snow leopard genomes. Nat. Commun. 2013, 4, 2433. [Google Scholar] [CrossRef]
- McBride, C. Operation White Lion; St. Martin’s Press: New York, NY, USA, 1981; p. 159. [Google Scholar]
- Cesare, M. Man-Eaters, Mambas and Marula Madness: A Game Ranger’s Life in the Lowveld; Jonathan Ball Publishing: Johannesburg, South Africa, 2011; p. 432. [Google Scholar]
- Turner, J.A.; Vasicek, C.A.; Somers, M.J. Effects of a colour variant on hunting ability: The white lion in South Africa. Open Sci. Repos. Biol. 2015, e45011830. [Google Scholar] [CrossRef]
- Smuts, G.L. Lion; MacMillan: Johannesburg, South Africa, 1982. [Google Scholar]
- Whitman, K.; Packer, C. The effect of sport hunting on the social organisation of the African lion (Panthera leo). In Proceedings of a Symposium on Lions and Leopards as Game Ranch Animals; Van Heerden, J., Ed.; The Wildlife Group, South African Veterinary Association: Pretoria, South Africa, 1997; pp. 177–183. [Google Scholar]
- Whitman, K.; Starfield, A.M.; Quadling, H.S.; Packer, C. Sustainable trophy hunting of African lions. Nature 2004, 428, 175–178. [Google Scholar] [CrossRef]
- Dicks, T. South Africa’s White Lions. Wandering through Photo Safaris and Expeditions. 2022. Available online: https://wanderingthru.com/the-white-lions-of-south-africa/ (accessed on 2 August 2022).
- Funston, P.J.; Mills, M.G.L.; Richardson, P.R.K.; van Jaarsveld, A.S. Reduced dispersal and opportunistic territory acquisition in male lions Panthera leo. J. Zool. 2003, 259, 131–142. [Google Scholar] [CrossRef]
- Turner, J.A.; Iongh, H.d.; Dunston-Clarke, E.J. Assessing the Social Cohesion of a Translocated Pride of White Lions Integrated with Wild Tawny Lions in South Africa, Using Social Network Analysis. Animals 2022, 12, 1985. [Google Scholar] [CrossRef]
- Burt, W.H. Territoriality and home range concepts as applied to mammals. J. Mammal. 1943, 24, 346–352. [Google Scholar] [CrossRef]
- Packer, C.; Scheel, D.; Pusey, A.E. Why lions form groups: Food is not enough. Am. Nat. 1990, 136, 1–19. [Google Scholar] [CrossRef]
- Schaller, G.B. The Serengeti Lion; The University of Chicago Press: London, UK, 1972. [Google Scholar]
- Gittleman, J.L.; Harvey, P.H. Carnivore home-ranges size, metabolic needs and ecology. Behav. Ecol. Sociobiol. 1982, 10, 57–63. [Google Scholar] [CrossRef]
- Van Orsdol, K.G.; Hanby, J.P.; Bygott, J.D. Ecological correlates of lion social organisation (Panthera leo). J. Zool. 1985, 206, 97–112. [Google Scholar] [CrossRef]
- Viljoen, P.C. The effects of change in prey availability in lion predation in a large natural ecosystem in northern Botswana. Symp. Proc. Zool. Soc. 1993, 65, 1–15. [Google Scholar]
- Spong, G. Space use in lions, Panthera leo, in the Selous Game Reserve: Social and ecological factors. Behav. Ecol. Sociobiol. 2002, 52, 303–307. [Google Scholar] [CrossRef]
- Bauer, H.; De Iongh, H.H. Lion (Panthera leo) home ranges and livestock conflicts in Waza National Park, Cameroon. Afr. J. Ecol. 2005, 43, 208–214. [Google Scholar] [CrossRef]
- Celesia, G.G.; Townsend Peterson, A.; Kerbis Peterhans, J.C.; Gnoske, T.P. Climate and landscape correlates of African lion (Panthera leo) demography. Afr. J. Ecol. 2009, 48, 58–71. [Google Scholar] [CrossRef]
- Tumenta, P.N.; de Iongh, H.H.; Funston, P.J.; Udo de Haes, H.A. Livestock depredation and mitigation methods practised by resident and nomadic pastoralists around Waza National Park, Cameroon. Oryx 2013, 47, 237–242. [Google Scholar] [CrossRef]
- Loveridge, A.J.; Valeix, M.; Davidson, Z.; Murindagomo, F.; Fritz, H.; Macdonald, D.W. Changes in home range size of African lions in relation to pride size and prey biomass in a semi-arid savanna. Ecography 2009, 32, 953–962. [Google Scholar] [CrossRef]
- Tumenta, P.N.; Visser, H.D.; van Rijssel, J.; Müller, L.; de Iongh, H.H.; Funston, P.J.; de Haes, H.A.U. Lion predation on livestock and native wildlife in Waza National Park, northern Cameroon. Mammalia 2012, 77, 247–251. [Google Scholar] [CrossRef]
- Packer, C.; Hilborn, R.; Mosser, A.; Kissui, B.; Borner, M.; Hopcraft, G.; Wilmshurst, J.; Mduma, S.; Sinclair, A.R.E. Ecological change, group territoriality, and population dynamics in Serengeti lions. Science 2005, 307, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Mosser, A.; Packer, C. Group territoriality and the benefits of sociality in the African lion Panthera leo. Anim. Behav. 2009, 78, 359–370. [Google Scholar] [CrossRef]
- Zehnder, A.; Henley, S.; Weibel, R. Home ranges of lions in the Kalahari, Botswana exhibit vast sizes and high temporal variability. Zoology 2018, 128, 46–54. [Google Scholar] [CrossRef]
- Bell, R.H.V. The Effect of Soil Nutrient Availability on Community Structure in African Ecosystems. Ecol. Stud. 1982, 42, 193–216. [Google Scholar] [CrossRef]
- McNaughton, S.J. Serengeti Grassland Ecology: The Role of Composite Environmental Factors and Contingency in Community Organization. Ecol. Monogr. 1983, 53, 291–320. [Google Scholar] [CrossRef]
- Stander, P. Demography of lions in the Etosha National Park, Namibia. Madoqua 1991, 18, 1–9. [Google Scholar]
- Hanby, J.P.; Bygott, J.D.; Packer, C. Ecology, Demography, and Behavior of Lions in Two Contrasting Habitats: Ngorongoro Crater and Serengeti Plains. In Serengeti II: Dynamics, Management, and Conservation of an Ecosystem; Sinclair, A.R.E., Arcese, P., Eds.; The University of Chicago Press: Chicago, IL, USA, 1995; pp. 315–331. [Google Scholar]
- Packer, C.; Pusey, A.S.; Eberly, L.E. Egalitarianism in female African lions. Science 2001, 293, 690–693. [Google Scholar] [CrossRef]
- Hunter, L.T.B. Behavioural Ecology of Reintroduced Lion and Cheetahs in the Phinda Resource Reserve, Kwa-Zulu Natal, South Africa. Ph.D. Thesis, Pretoria University, Pretoria, South Africa, 1998. [Google Scholar]
- Bertram, B. Lion population regulation. Afr. Wildl. J. 1973, 2, 215–225. [Google Scholar] [CrossRef]
- Funston, P.J.; Mills, M.G.L.; Biggs, H.C.; Richardson, P.R.K. Hunting by male lions: Ecological influences and socioecological implications. Anim. Behav. 1998, 56, 1333–1345. [Google Scholar] [CrossRef]
- Tuqa, J.H.; Funston, P.J.; Musyoki, C.; Ojwang, G.O.; Gichuki, N.N.; Bauer, H.; Tamis, W.; Dolrenry, S.; Van’t Zelfde, M.; de Snoo, G.R.; et al. Impact of severe climate variability on lion home range and movement patterns in the Amboseli ecosystem, Kenya. Glob. Ecol. Conserv. 2014, 2, 1–10. [Google Scholar] [CrossRef]
- Powell, R.A. Animal Home Ranges and Territories and Home Range Estimators. In Research Techniques in Animal Ecology: Controversies and Consequences; Boitani, L., Fuller, T.K., Eds.; Colombia University Press: New York, NY, USA, 2000; pp. 64–110. [Google Scholar]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Dunston, E.; Abell, J.; Doyle, R.; Duffy, D.; Poynter, C.; Kirk, J.; Hilley, V.; Forsyth AJenkins, E.; McAllister, D.; Freire, R. Does captivity influence and hunting behaviour? Assessment for an ex situ reintroduction program of African lions Panthera leo. Mamm. Rev. 2017, 47, 254–260. [Google Scholar] [CrossRef]
- Lehmann, M.B.; Funston, P.J.; Owen, C.R.; Slotow, R. Home range utilisation and territorial behaviour of lions (Panthera leo) on Karongwe Game Reserve, South Africa. PLoS ONE 2008, 3, e3998. [Google Scholar] [CrossRef] [PubMed]
- Yiu, S.-W.; Parrini, F.; Karczmarski, L.; Keith, M. Home range establishment and utilization by reintroduced lions (Panthera leo) in a small South African wildlife reserve. Integr. Zool. 2017, 12, 318–332. [Google Scholar] [CrossRef]
- Druce, D.; Genis, H.; Braak, J.; Greatwood, S.; Delsink, A.; Kettles, R.; Hunter, L.; Slotow, R. Population demography and spatial ecology of a reintroduced lion population in the Greater Makalali Conservancy, South Africa. Koedoe 2004, 47, 103–118. [Google Scholar] [CrossRef]
- Hayward, M.; Hayward, G.; Druce, D.; Kerley, G. Do fences constrain predator movements on an evolutionary scale? Home range, food intake and movement patterns of large predators reintroduced to Addo Elephant National Park, South Africa. Biodivers. Conserv. 2008, 18, 887–904. [Google Scholar] [CrossRef]
- Mucina, L.; Rutherford, M.C. (Eds.) The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006; p. 19. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 15 July 2022).
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R; Chapman and Hall/CRC Press: London, UK, 2015; Available online: https://www.routledge.com/Spatial-Point-Patterns-Methodology-and-Applications-with-R/Baddeley-Rubak-Turner/9781482210200/ (accessed on 15 July 2022).
- Bivand, R.; Lewin-Koh, N. Maptools: Tools for Handling Spatial Objects; R Package Version 1.1-4. 2022. Available online: https://r-forge.r-project.org/projects/maptools (accessed on 2 August 2022).
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 15 July 2022).
- Cheng, J.; Karambelkar, B.; Xie, Y. Leaflet: Create Interactive Web Maps with the JavaScript ‘Leaflet’ Library; R Package Version 2.0.4.1. 2021. Available online: https://CRAN.R-project.org/package=leaflet (accessed on 15 July 2022).
- Appelhans, T.; Detsch, F.; Reudenbach, C.; Woellauer, S. Mapview: Interactive Viewing of Spatial Data in R.; R Package Version 2.10.0. 2021. Available online: https://CRAN.R-project.org/package=mapview (accessed on 15 July 2022).
- Carslaw, D.C.; Ropkins, K. Openair—An R package for air quality data analysis. Environ. Model. Softw. 2012, 27–28, 52–61. [Google Scholar] [CrossRef]
- Duong, T.; Smoothing, K. R Package Version 1.13.2. 2021. Available online: https://CRAN.R-project.org/package=ks (accessed on 15 July 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Dunnington, D. Ggspatial: Spatial Data Framework for Ggplot2; R Package Version 1.1.5. 2021. Available online: https://CRAN.R-project.org/package=ggspatial (accessed on 18 July 2022).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling; R Package Version 3.4-13. 2021. Available online: https://CRAN.R-project.org/package=raster (accessed on 18 July 2022).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library; R Package Version 1.5-27. 2021. Available online: https://CRAN.R-project.org/package=rgdal (accessed on 18 July 2022).
- Blondel, E. Cleangeo: Cleaning Geometries from Spatial Objects; R Package Version 0.2-4. 2021. Available online: https://CRAN.R-project.org/package=cleangeo (accessed on 18 July 2022).
- Kerley, G.; Behrens, K.; Carruthers, J.; Diemont, M.; Du Plessis, J.; Minnie, L.; Richardson, P.; Somers, M.; Tambling, C.; Turpie, J.; et al. Livestock predation in South Africa: The need for and value of a scientific assessment. S. Afr. J. Sci. 2017, 113, 1–3. [Google Scholar] [CrossRef]
- Powell, R.A.; Mitchell, M.S. What is home range? J. Mamm. 2012, 93, 948–958. [Google Scholar] [CrossRef]
- Lesilau, F.; Verschueren, S.; van’t Zelfde, M.; Musters, K.C.; de Snoo, G.R.; de Iongh, H.H. Spatial ecology of lions in a small, semi fenced park surrounded by dense human populations: The case study of Nairobi National Park, Kenya. Mammalia 2021, 85, 198–207. [Google Scholar] [CrossRef]
- Hayward, M.W.; Slotow, R. Temporal partitioning of activity in large African carnivores: Tests of multiple hypotheses. S. Afr. J. Wild. Res. 2009, 39, 109–125. [Google Scholar] [CrossRef]
- Rudnai, J.A. The Social Life of the Lion; Medical and Technical Publishing Co., Ltd.: Lancaster, UK, 1973; p. 122. [Google Scholar]
- McDonald, T. Ecological Management Plan: Tsau Conservancy; Unpublished Report; Monzweni Eco-Developers: Hoedspruit, South Africa, 2005. [Google Scholar]
- Manly, B.F.; McDonald, L.; Thomas, D.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals: Statistical Design and Analysis for Field Studies, 2nd ed.; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Davidson, Z.; Valeix, M.; Loveridge, A.J.; Hunt, J.E.; Johnson, P.J.; Madzikanda, H.; Macdonald, D.W. Environmental determinants of habitat and kill site selection in a large carnivore: Scale matters. J. Mammal. 2012, 93, 677–685. [Google Scholar] [CrossRef]
- Garai, M.E.; Slotow, R.; Reilly, B.; Carr, R.D. History and success of elephant reintroductions to small fenced reserves in South Africa. Pachyderm 2004, 37, 28–36. [Google Scholar]
- Gusset, M.; Ryan, S.J.; Hofmeyr, M.; Van Dyk, G.; Davies-Mostert, H.T.; Graf, J.A.; Owen, C.; Szykman, M.; Macdonald, D.W.; Monfort, S.L.; et al. Efforts going to the dogs? Evaluating attempts to re-introduce endangered wild dogs in South Africa. J. Appl. Ecol. 2008, 45, 100–108. [Google Scholar] [CrossRef]
- Slotow, R.; Hunter, L.T.B. Reintroduction Decisions Taken at the Incorrect Social Scale Devalue Their Conservation Contribution: African Lion in South Africa. In The Reintroduction of Top-Order Predators; Hayward, M.W., Somers, M.J., Eds.; Blackwells Publishing: Oxford, UK, 2008. [Google Scholar]
- Kettles, R.; Slotow, R. Management of free-ranging lions on an enclosed game reserve. S. Afr. J. Wild. Res. 2009, 39, 23–33. [Google Scholar] [CrossRef]
- Miller, S.M.; Bissett, C.; Parker, D.M.; Burger, A.; Courtenay, B.; Dickerson, T.; Naylor, S.; Druce, D.J.; Ferreira, S.; Slotow, R.; et al. Management of reintroduced lions in small, fenced reserves in South Africa: An assessment and guidelines. S. Afr. J. Wild. Res. 2013, 43, 138–154. [Google Scholar] [CrossRef]
- McEvoy, O.K.; Ferreira, S.M.; Parker, D.M. The influence of population demographics on lion (Panthera leo) growth rates in small, fenced wildlife reserves. Afr. J. Wild. Res. 2021, 51, 75–89. [Google Scholar] [CrossRef]
- MacDonald, D.W. The ecology of carnivore social behaviour. Nature 1983, 301, 379–383. [Google Scholar] [CrossRef]
- Nilsen, E.B.; Herfindal, I.; Linnell, J.D.C. Can intra-specific variation in carnivore home-range size be explained using remote sensing estimates of environmental productivity? Ecoscience 2005, 12, 68–75. [Google Scholar] [CrossRef]
- Funston, P.J. Predator-Prey Relationships between Lions and Large Ungulates in the Kruger National Park. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 1999. [Google Scholar]
- Kilian, P.J. The Ecology of Reintroduced Lions on the Welgevonden Private Game Reserve, Waterberg. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2003. [Google Scholar]
- Briers-Louw, W.; Leslie, A. Ecology of Three Apex Predators in Majete Wildlife Reserve, Malawi. Master’s Thesis, Universsity of Stellenbosch, Stellenbosch, South Africa, 2017. [Google Scholar] [CrossRef]
- Lesilau, F. Human–Lion Conflict around Nairobi National Park. Ph.D. Thesis, University of Leiden, Leiden, The Netherlands, 2019. [Google Scholar]
- Husseman, J.S.; Murray, D.L.; Power, G.; Mack, C.; Wenger, C.R.; Quigley, H. Assessing differential prey selection patterns between two sympatric large carnivores. Oikos 2003, 101, 591–601. [Google Scholar] [CrossRef]
- Gorini, L.; Linnell, J.D.C.; May, R.; Panzacchi, M.; Boitani, L.; Odden, M.; Nilsen, E.B. Habitat heterogeneity and mammalian predator-prey interactions. Mamm. Rev. 2012, 42, 55–77. [Google Scholar] [CrossRef]
- Riggio, J.; Jacobson, A.; Dollar, L.; Bauer, H.; Becker, M.; Dickman, A.; Funston, P.; Groom, R.; Henschel, P.; de Iongh, H. The size of savannah Africa: A lion’s Panthera leo view. Biodivers. Conserv. 2013, 22, 17–35. [Google Scholar] [CrossRef]
- Western, D. Water availability and its influence on the structure and dynamics of the savanna large mammal community. Afr. J. Ecol. 1975, 13, 265–286. [Google Scholar] [CrossRef]
- Mills, M.; Retief, P. The effect of windmill closure on the movement patterns of ungulates along the Auob riverbed. Koedoe Afr. Prot. Area Conserv. Sci. 1984, 27, 107–118. [Google Scholar] [CrossRef]
- Owen-Smith, R.N. Ecological guidelines for waterpoints in extensive protected areas. S. Afr. J. Wild. Res. 1996, 26, 107–112. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pride | Adult (>4 yr) | Adult (>4 yr) | Sub Adult (2–4 yr) | Sub Adult (2–4 yr) | Cub (<2 yr) | Cub (<2 yr) | Total | Observation Period |
|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |||
| Royal | 1 * | 1 | 2 | 1 | 0 | 0 | 5 | 25 January 2010–12 December 2011 |
| Tsau | 2 * | 2 | 2 | 1 | 0 | 0 | 7 | 1 January 2018–31 December 2020 |
| Lion | MCP | KDE 95% | KDE 50% |
|---|---|---|---|
| Male 1 (RM1) | 6.81 | 5.47 | 1.69 |
| Female 1 (RF1) | 6.33 | 5.32 | 1.59 |
| Subadult Male 1 (RSM1) | 6.31 | 5.21 | 1.58 |
| Subadult Male 2 (RSM2) | 6.31 | 5.21 | 1.61 |
| Subadult Female 1 (RSF1) | 6.31 | 5.21 | 1.61 |
| Lion | MCP | KDE 95% | KDE 50% |
|---|---|---|---|
| Male 1 (TM1) | 6.58 | 5.64 | 1.62 |
| Male 2 (TM2) | 6.89 | 5.72 | 1.61 |
| Female 1 (TF1) | 6.52 | 5.53 | 1.58 |
| Female 2 (TF2) | 6.79 | 5.61 | 1.61 |
| Subadult Male 1 (TSM1) | 6.51 | 5.37 | 1.61 |
| Subadult Male 2 (TSM2) | 6.51 | 5.37 | 1.61 |
| Subadult Female 1 (RSF1) | 6.51 | 5.37 | 1.58 |
| Home Range | All Years | Males | Females |
|---|---|---|---|
| 50% Kernel Density Estimator (km2) | 1.76 ± 0.10 | 1.85 ± 0.31 | 1.79 ± 0.32 |
| (1.61–1.87) | (1.48–2.24) | (1.47–2.20) | |
| 95% Kernel Density Estimator (km2) | 5.53 ± 0.35 | 5.55 ± 0.21 | 5.50 ± 0.23 |
| (5.50–5.64) | (5.20–5.75) | (5.14–5.71) | |
| 100% Minimum Convex Polygon (km2) | 5.70 ± 0.23 | 5.75 ± 0.21 | 5.63 ± 0.41) |
| (5.32–6.25) | (5.41–6.14) | (5.60–5.66) | |
| Daily Distance (km) | 10.91 ± 4.90 | 11.54 ± 5.12 | 11.31 ± 4.94 |
| (0.01–19.22) | (0.01–19.30) | (0.01–19.30) |
| Home Range | All Seasons | Summer (Wet) | Winter (Dry) |
|---|---|---|---|
| 50% Kernel Density Estimator (km2) | 1.83 ± 0.28 | 1.89 ± 0.39 | 1.77 ± 0.17 |
| (1.37–2.47) | (1.37–2.47) | (1.58–2.10) | |
| 50% Kernel Density Estimator (km2) | 5.33 ± 0.34 | 5.44 ± 0.21 | 5.21 ± 0.11 |
| (5.07–5.70) | (5.14–5.70) | (5.07–5.35) | |
| 100% Minimum Convex Polygon (km2) | 5.57 ± 0.20 | 5.65 ± 0.21 | 5.48 ± 0.19 |
| (5.15–5.98) | (5.15–5.98) | (5.20–5.70) | |
| Daily Distance (km) | 9.46 ± 4.59 | 9.62 ± 4.75 | 9.29 ± 4.41 |
| (0.01–18.24) | (0.01–18.51) | (0.01–17.96) |
| Home Range | Royal Pride | Tsau Pride |
|---|---|---|
| 50% Kernel Density Estimator (km2) | 1.76 ± 0.10 | 1.81 ± 0.29 |
| (1.61–1.87) | (1.47–2.24) | |
| 50% Kernel Density Estimator (km2) | 5.57 ± 0.35 | 5.51 ± 0.21 |
| (5.50–5.64) | (5.14–5.75) | |
| 100% Minimum Convex Polygon (km2) | 5.65 ± 0.82 | 5.70 ± 0.23 |
| (5.57–5.86) | (5.32–6.25) |
| Vegetation Type | Proportion of TTPA | Royal Pride | Royal Pride | Tsau Pride | Tsau Pride |
|---|---|---|---|---|---|
| Wi | Bi | Wi | Bi | ||
| Woodland | 12% | 1.595 | 0.288 | 1.052 | 0.184 |
| Plains | 13% | 0.239 | 0.043 | 0.390 | 0.068 |
| Riverine | 27% | 1.193 | 0.216 | 1.299 | 0.227 |
| Thicket | 48% | 2.502 | 0.453 | 2.980 | 0.521 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turner, J.A.; Dunston-Clarke, E.J.; Fabris-Rotelli, I.; de Iongh, H. Home Range and Movement Patterns of Reintroduced White Lions (Panthera leo melanochaita) in the Kruger to Canyons Biosphere Reserve, South Africa. Animals 2022, 12, 2003. https://doi.org/10.3390/ani12152003
Turner JA, Dunston-Clarke EJ, Fabris-Rotelli I, de Iongh H. Home Range and Movement Patterns of Reintroduced White Lions (Panthera leo melanochaita) in the Kruger to Canyons Biosphere Reserve, South Africa. Animals. 2022; 12(15):2003. https://doi.org/10.3390/ani12152003
Chicago/Turabian StyleTurner, Jason A., Emma J. Dunston-Clarke, Inger Fabris-Rotelli, and Hans de Iongh. 2022. "Home Range and Movement Patterns of Reintroduced White Lions (Panthera leo melanochaita) in the Kruger to Canyons Biosphere Reserve, South Africa" Animals 12, no. 15: 2003. https://doi.org/10.3390/ani12152003
APA StyleTurner, J. A., Dunston-Clarke, E. J., Fabris-Rotelli, I., & de Iongh, H. (2022). Home Range and Movement Patterns of Reintroduced White Lions (Panthera leo melanochaita) in the Kruger to Canyons Biosphere Reserve, South Africa. Animals, 12(15), 2003. https://doi.org/10.3390/ani12152003