Abstract
Heavy metal contamination is currently a major environmental concern, as most agricultural land is being polluted from municipal discharge. Among various other pollutants, cadmium (Cd), one of the most harmful heavy metals, enters into the food chain through the irrigation of crops with an industrial effluent. In the present study, a pot experiment was designed to assess the effect of different nitrogen (N)-fertilizer forms in the phytoremediation of Cd through Solanum nigrum L. Two types of N fertilizers (NH4NO3 and urea) were applied to the soil in different ratios (0:0, 100:0, 0:100, and 50:50 of NH4NO3 and urea, individually) along with different Cd levels (0, 25, and 50 mg kg−1). The plants were harvested 70 days after sowing the seeds in pots. Cadmium contamination significantly inhibited the growth of leaves and roots of S. nigrum plants. Cadmium contamination also induced oxidative stress; however, the application of N-fertilizers increased the plant biomass by inhibiting oxidative stress and enhancing antioxidants’ enzymatic activities. The greatest plant growth was observed in the urea-treated plants compared with the NH4NO3-treated plants. In addition, urea-fed plants also accumulated higher Cd concentrations than NH4NO3-fed plants. It is concluded that urea is helpful for better growth of S. nigrum under Cd stress. Thus, an optimum concentration of N-fertilizers might be effective in the phytoremediation of heavy metals through S. nigrum.
1. Introduction
Anthropogenic activities have resulted in environmental degradation and a significant reduction in soil productivity [1]. Abiotic stress, such as heavy metals, extreme temperature, and salinity, affect crop production [2]. Industrial discharge, sewage sludge, mining, and agriculture have become major precursors of heavy metal discharge into water bodies and soil [3]. Compared with all other heavy metals, cadmium (Cd) is considered lethal once it becomes part of the food cycle.
Around 70% of the dietary intake of Cd occurs from consuming the vegetables and grain crops grown in Cd-contaminated soils [4]. Cadmium is highly toxic for plant growth and seed germination, in addition to the antioxidant, photosynthetic, and enzyme activities [5]. It also causes visual toxicity symptoms including a reduction in plant height, necrosis, and leaf chlorosis [6]. Excessive concentration of Cd also impairs the uptake of some integral nutrients, such as iron, zinc, and manganese [7]. Increased electrolyte leakage (EL), malondialdehyde (MDA) content, and oxidative stress in plants, along with a reduced enzyme activity, are also evident in Cd-affected plants [8]. Therefore, it is imperative to control Cd pollution so as to minimize the Cd concentrations in the soil, as well as to reduce its uptake by plants, especially in the edible portions of plants [9]. Phytoremediation is a useful technique to cope with the heavy metals problem, through use of green plants to remove/degrade pollutants from the contaminated soils. Phytoremediation is considered an attractive substitute for soil remediation, because it is one of the cheapest and most ecofriendly methods compared with other techniques [10].
Different plants accumulate heavy metals when grown in a metal contaminated soil; an example of such an uptake is the Cd taken up by Brassica oleracea L. [11]. In addition, various hyperaccumulator plants can help to remediate the highly metal-polluted soils. A number of Cd-hyperaccumulator plant species have been reported, including S. nigrum [12], Malva rotundifolia [13], Bidens pilosa [14], Noccaea caerulescens [15], Sedum alfredii [16], and Jatropha curcas [17]. Among these Cd hyperaccumulators, S. nigrum is considered more efficient for its fast growth and higher tolerance to Cd, without compromising biomass production [9]. However, the growth of S. nigrum may be negatively affected at higher Cd concentrations, which require some amendments, especially in the early growth stages, in order to improve the phytoextraction used by this plant.
The application of N-fertilizers is considered an effective measure to improve the soil fertility and phytoremediation efficiency of hyperaccumulator plants [18]. Nitrogen is an integral component of plants that makes various secondary metabolites, and is also helpful in chlorophyll, which is a key component of photosynthesis [19,20]. Numerous effects of N fertilization on soil–Cd dynamics have been observed, including desorption and adsorption, chemical transformation, dilution effect, and transportation, which ultimately influence Cd uptake by plants [21]. Many studies have shown that the addition of different N compounds (NH4 and NO3) in different combinations and dosages affect plant growth and biomass [22,23,24,25,26].
The study was designed to evaluate the effect of various N-fertilizers on the phytoremediation of Cd through S. nigrum by measuring changes in plant growth, chlorophyll and carotenoid contents, photosynthetic parameters, oxidative stress, and antioxidant enzyme activities under different N fertilizer combinations and Cd contamination levels.
2. Materials and Methods
2.1. Experimental Design
A pot experiment was conducted in a botanical garden located in Government College University Faisalabad (Faisalabad, Pakistan; 31°25′0″ N, 73°5′28″ E). The soil texture was measured by the method of Bouyoucos [27]; the pH and electrical conductivity (EC) with pH and EC meters, respectively; organic carbon was measured using the Walkley–Black method [28]; and the sodium absorption ratio (SAR) and soluble ions were measured using the method described by Page et al. [29]. The soil physicochemical characteristics are shown in Table 1. The soil was spiked and entirely mixed with different Cd (CdCl2 2.5H2O) levels (0, 25, and 50 mg kg−1). Nitrogen fertilizers of two different kinds (NH4NO3 and urea) with varying ratios (0–0, 100–0, 0–100, and 50–50 mg kg−1) were added into the soil. It is noteworthy that the contents for Cd, Pb, Cu, and Zn were fundamentally low and unnoticeable in these fertilizers.

Table 1.
Soil physicochemical properties used for the experiment.
2.2. Soil Pot Experiments
In this study, 5% sodium hypochlorite (NaClO) was applied for 10 min in order to sterilize the S. nigrum seeds, which were then washed four times with deionized water. The washed and blot-dried seeds were planted in a plastic tray filled with sand, and half-strength Hoagland solution was applied. After three weeks of germination, the uniform seedlings were transferred into pots (four seedlings in each pot). Every single pot was filled with 5 kg of soil. All of the pots were set following complete randomized design (CRD) with four repeats for each treatment, and the water was topped up with tap water in order to maintain the 70% soil water-holding limit.
2.3. Plant Harvesting
The plants were harvested 70 days after sowing the seeds and were sectioned into shoots and roots. The length of the roots and shoots, numbers of leaves, leaf area, and fresh weight of plants were measured. The root and shoot samples were further dried in an oven for 72 h at 70 °C, and the dry weights were measured.
2.4. Determination of Photosynthetic, Chlorophyll, and Carotenoid Content Parameters
Fresh leaf samples (0.5 g) were soaked in acetone (85%, v/v, Sigma) and placed in the dark. The soaked samples were centrifuged (4000× g for 10 min, 4 °C) and the supernatant was collected. Data were recorded on a spectrophotometer at wavelengths of 470, 647, and 664.5 nm, separately. The chlorophyll content, i.e., chlorophyll a, chlorophyll b, and total chlorophyll, and carotenoids were recorded [30]. On a sunny day (10:00 a.m. to 12:00 p.m.), an infrared gas analyzer (IRGA) was used for the assessment of the conductance of the stomata water-use efficiency, rate of transpiration, and photosynthetic rate in the plants’ leaves.
2.5. Determination of EL, MDA, H2O2, and Antioxidant Enzyme Concentration
The measurement of both the oxidative stress markers and the activities of antioxidant enzymes were done 70 days after seed sowing. The shoot and root samples were placed in glass tubes vertically and heated at 32 °C for 2 h in distilled water of a known volume in order to measure the electrolyte leakage. This solution was termed EC1. Then, the same solution was heated at 121 °C for 20 min, and EC of this second solution was recorded and named EC2. The Dionisio-Sese and Tobita [31] equation was used to estimate the EL content.
For the estimation of the H2O2 content, a phosphate buffer solution (3.0 mL) was added to the sample (50 mg) and centrifuged at 6000× g for 30 min, keeping the temperature at 4 °C. Then, 1 mL of titanium sulfate (0.1%) was mixed in a supernatant and centrifuged at 6000× g for 20 min at 4 °C. Absorption was determined at a wavelength of 410 nm and a coefficient of extinction for H2O2 of 0.28 µmol−1 cm−1. The peroxidase (POD), catalase (CAT), superoxide dismutase (SOD), and ascorbate peroxidase (APX) enzyme activities were determined following the recommendations of Zhang [32] and Aebi [33]. The samples were prepared in a phosphate buffer (0.05 mmol) and the supernatant was gained through centrifuging for 10 min at 12,000× g and 4 °C.
2.6. Cadmium Determination in Plants, Translocation Factor and Bioaccumulation Factor
Crushed root and shoot samples were digested using HNO3-HClO4 (3:1, v/v). The samples were kept in 65% HNO3-HClO4 (3:1, v/v) for one night and then put on a hot plate after adding HNO3 (5.0 mL). The clear solution obtained after digestion was filtered and the Cd concentration was measured using an atomic absorption spectrophotometer. Translocation factor (TF) was calculated by using Equation (1):
and the bioaccumulation factor (BCF) was calculated using Equation (2):
TF = Cd (Plant shoot)/Cd (Plant root)
TF = Cd concentration in root/Total Cd concentration in soil
2.7. Statistical Analysis
A two-way analysis of variance test (ANOVA) was applied in order to test the significance of the N fertilizers and Cd concentrations. Tukey’s posthoc test was applied for the multiple means comparison technique. The statistical analysis was performed with SPSS for Windows Software v. 19 (IBM, Armonk, NY, USA).
3. Results
3.1. Assessment of N-Fertilizer on Growth and Biomass
Stunted growth was observed in the control group, while no such signs were observed in the N-fertilizer-treated group (Figure 1). The roots’ and shoots’ dry weight, the number of leaves per plant shoot length and root length, and the leaf area of the S. nigrum plants significantly increased in the N-fertilizer treatments (Figure 1). A maximum plant growth was observed in the urea-treated plants at a Cd level (0 mg kg−1), with respect to the NH4NO3-treated and control plants. In addition, more leaves were observed in the urea-treated plants compared with the NH4NO3-treated plants (Figure 1).
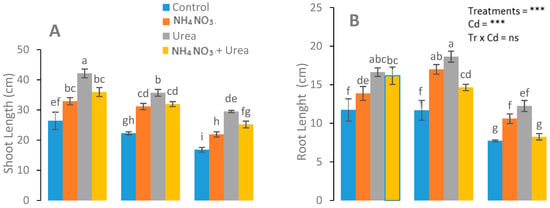
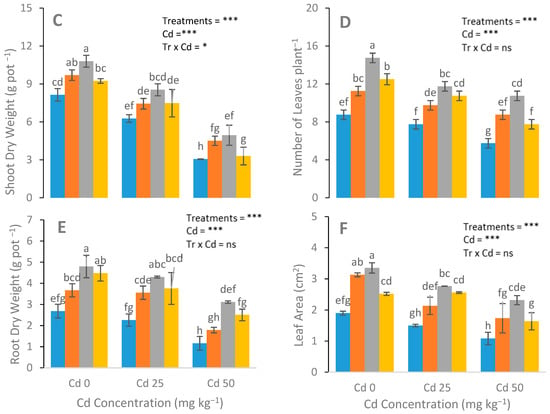
Figure 1.
Effect of the Cd (0, 25, and 50 mg kg−1) and N fertilizers (ratios of 0:0, 100:0, 0:100, and 50:50 mg kg−1 urea, NH4NO3, and urea + NH4NO3) on shoot length (A), root length (B), shoot dry weight (C), number of leaves (D), root dry weight (E), and leaf area (F) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
3.2. Assessment of Gas Exchange, Chlorophyll, and Carotenoid Content Attributes
Differential responses of chlorophyll and gas exchange attributes were recorded in urea-treated and NH4NO3-treated plants. Significant changes with the maximum values of Chl a, Chl b, total Chl, and carotenoid contents were observed in the urea-fed plants (Figure 2). A nonsignificant increase in chlorophyll content was observed in the urea-treated plants at 25 mg kg−1 Cd concentrations, in contrast to the NH4NO3-treated and control plants. In addition, a higher expression of gas attributes was recorded in the urea-treated plants than that of the NH4NO3-fed plants (Figure 3).
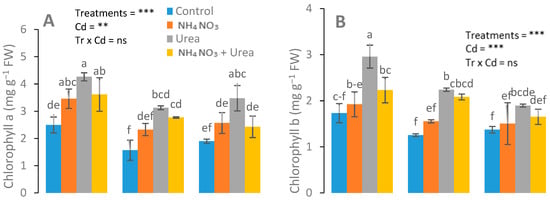
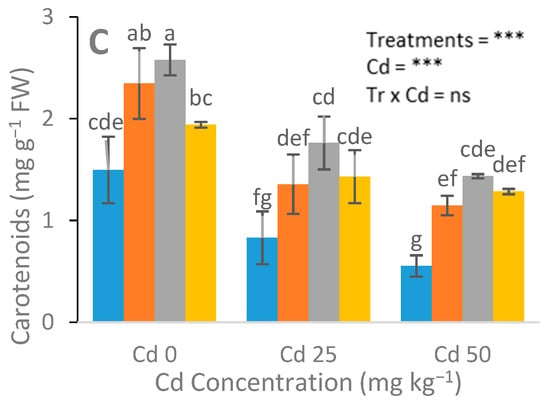
Figure 2.
Effects of Cd (0, 25, and 50 mg kg−1) and N fertilizers (ratios of 0:0, 100:0, 0:100, and 50:50 mg kg−1 urea, NH4NO3, and urea + NH4NO3) on chlorophyll a (A), chlorophyll b (B), and carotenoid (C) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
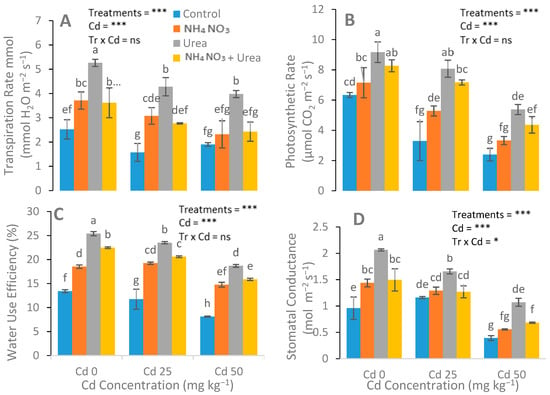
Figure 3.
Effects of Cd stress (0, 25, and 50 mg kg−1) and N fertilizers (ratios of 0:0, 100:0, 0:100, and 50:50 mg kg−1 urea, NH4NO3, and urea + NH4NO3) on the transpiration rate (A), photosynthetic rate (B), water-use efficiency (C), and stomata conductance (D) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
3.3. Assessment of Antioxidant Enzyme Activities and EL, MDA, and H2O2
After N-fertilizer supplementation, a significant reduction was noticed in the MDA, H2O2, and electrolyte leakage parameters (Figure 4). Reductions in EL in the leaves of the urea, NH4NO3, and urea + NH4NO3 treated plants were 15%, 33%, and 22%, respectively, compared with the control plants. The catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX), and superoxide dismutase (SOD) enzyme activities in the S. nigrum leaf were significantly increased after the addition of N-fertilizers (Figure 5). In contrast to the control group, the increases in the POD, CAT, APX, and SOD values of the N-fertilized plants were 41%, 21%, and 64%; 47%, 22%, and 66%; 15%, 28%, and 42%; and 48%, 69%, and 25%, respectively.
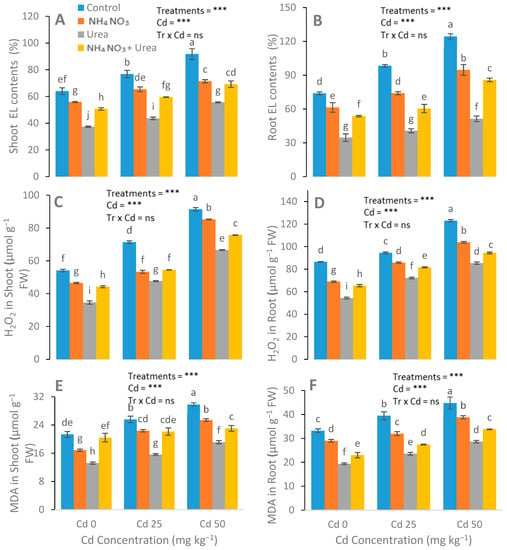
Figure 4.
Effect of Cd (0, 25, and 50 mg kg−1) and N fertilizers (ratios of 0:0, 100:0, 0:100, and 50:50 mg kg−1 urea, NH4NO3, and urea + NH4NO3) on electrolyte leakage (EL) in leaves (A), EL in roots (B), H2O2 in leaves (C), H2O2 in roots (D), malondialdehyde (MDA) in leaves (E), and MDA in the roots (F) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
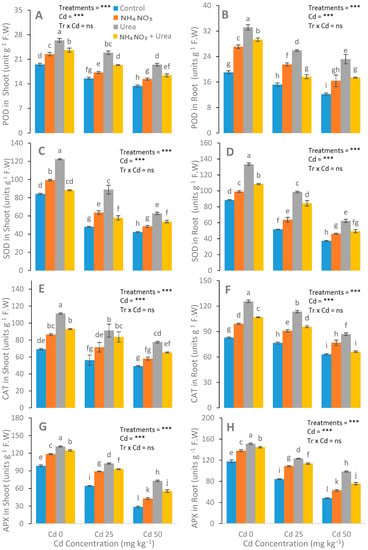
Figure 5.
Effects of Cd (0, 25, and 50 mg kg−1) and N fertilizers (levels 0–0, 100–0, 0–100, and 50–50 mg kg−1 for urea, NH4NO3, and urea + NH4NO3) on POD in leaves (A), POD in roots (B), SOD in leaves (C), SOD in roots (D), CAT in leaves (E), CAT in roots (F), APX in leaves (G), and APX in roots (H) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
3.4. Assessment of Concentration of Cd in Plants
The optimum induction of N-fertilizers remarkably enhanced the Cd concentration in the roots and shoots of S. nigrum compared with the respective controls (Figure 6). In order to assess the phytoremediation potential of Cd by S. nigrum, the TF and BCF were calculated. Results showed that plants supplemented with urea had higher BCF when compared with other treatments.
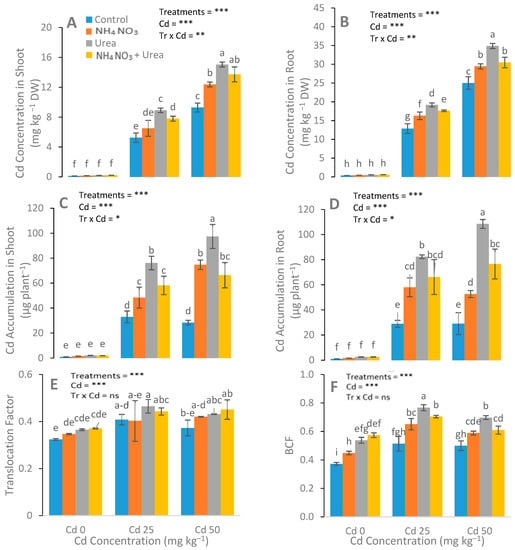
Figure 6.
Effect of Cd stress (0, 25, and 50 mg kg−1) and N fertilizers (levels 0–0, 100–0, 0–100, and 50–50 mg kg−1 for urea, NH4NO3, and urea + NH4NO3) on Cd uptake in shoots (A), Cd uptake in roots (B), Cd accumulation in shoots (C), Cd accumulation in roots (D) translocation factor (E) and bioaccumulation factor (BCF) (F) of S. nigrum. Different letters show a significance difference at p ˂ 0.05 along with n = 4.
4. Discussion
4.1. Biomass and Plant Growth
It is an established fact that fertilizer application generally has positive effects on plant growth under heavy metal toxicity [34,35]. In the present study, the highest plant growth was observed in the urea-treated plants at 25 mg kg−1 Cd concentrations, in contrast to the NH4NO3-treated plants (Figure 1). The findings of our study are in accordance with the finding of Lin et al. [36], who found that 2.5 mmol N L−1 was the optimal concentration to boost S. alfredii shoot growth. Moreover, 1.0 mmol N L−1 was found to be an optimal concentration for Cd and Zn storage in the shoots of S. alfredii. Furthermore, stunted plant growth under heavy metal stress was noticed. For example, Rabelo et al. [37] reported that 2.0 mM Cd in Tanzania guinea grass inhibited the production of new tillers and leaves. A reduced nitrate uptake from the nutrient solution was reported by Gouia et al. [38] for Cd-exposed plants. It was investigated that the Cd concentration increased in S. nigrum with a supply of N-fertilizer (NH4NO3) without affecting the Cd speciation in plants [39]. Wei et al. [40] confirmed that the accretion of Cd in S. nigrum shoots and roots is augmented by applying urea in the growth medium. In our study, the root and shoot dry weights, shoot length, numbers of leaves per plant, and roots length of S. nigrum plants were significantly enhanced with the N-fertilizer application. In our current study, disparity in the Cd absorption in differently treated plants was observed, which is in line with the results of Ye et al. [41], who described an enhanced absorption of Cd and a phytoextraction efficiency of T. patula after N-fertilizer treatment. Moreover, our results are also supported by Yang et al. [42].
4.2. Photosynthetic Pigments
Maximum chlorophyll content was observed in the urea-treated plants at a 25 mg kg−1 Cd concentration, compared with the NH4NO3-treated and control plants (Figure 2). Our results support the previous reports [43], in which the author claimed that different N forms, e.g., ammonium nitrate (NH4NO3), ammonium sulfate ((NH4)2SO4), and calcium nitrate (Ca (NO3)2), significantly altered the Chl a, Chl b, total Chl, and carotenoid contents under Cd stress. Cadmium is a phytotoxic metal that causes growth inhibition. A high dose or prolonged exposure to Cd can lead to the death of plants, which is the result of disturbed respiration, reduced photosynthesis, and altered assimilation of N in plants [44]. Cadmium toxicity alters the N metabolism directly or indirectly [45]. It was found that an optimum N dosage may alleviate Cd toxicity to plants by improving the photosynthetic activity, stromal proteins, and the plant growth and biomass [46]. Under stress conditions, plants provoke antioxidant enzymes and certain metabolite activities for their survival in order to negate stress [47]. Jalloh et al. [48] reported that an appropriate dosage of urea and NH4+-N under Cd stress stimulated and increased the SOD and POD activities. Significantly increased MDA content in Cd-stressed plants were observed with addition of NH4+-N and/or urea in rice plants at the milking stage.
4.3. Oxidative Stress, Antioxidant Enzymes, and Cd Concentration
Malmir [49] found that Cr-induced oxidative stress, subsequently increasing the H2O2 and MDA contents. Similar evidence of Cr-induced oxidative stress was observed in Helianthus annuus L. [50] and Brassica. Various studies have shown that different amendments have reduced the Cd toxicity in different plant species, for example, EDTA reduced Cd toxicity in Brassica [51], silicon in Brassica napus L. [52], hydrogen peroxide in Brassica napus L. [53], supermine in Vigna radiate L. [54], and selenium in rapeseed seedlings [55]. The markedly reduced EL and MDA contents in the N-fertilizer-treated, Cd-stressed plants in our study demonstrate the significant role of N fertilizers for Cd stress. Our results revealed that SOD, POD, CAT, and APX activities of S. nigrum were recorded in response to Cd stress. Furthermore, the POD, CAT, SOD, and APX activities significantly increased after the application of N-fertilizers. Similar results were observed previously [43], indicating that N forms had a prominent impact on oxidative stress inflicted by Cr toxicity. Therefore, the Ca(NO3)2-fed plants had minimal oxidative stress compared with the (NH4)2SO4 and urea-treated plants.
Similar findings were observed for the N fertilizers for the phytoremediation of Cd, and even with different environmental conditions, soil textures, and experimental durations [56], and only a few analyses, we acknowledged the possible mechanisms for Cd phytoremediation with detailed analysis in our recent study. In addition, urea-treated plants translocated and accumulated a higher concentration of Cd than the NH4NO3-treated plants. However, the application of N-fertilizers inhibited oxidative damage and enhanced antioxidants’ enzyme activities. Our results depicted that at all levels of applied Cd, the both TF and BCF were enhanced with the application of urea alone, as compared with all other treatments (Figure 6E,F). This shows that among the various treatments, the plants treated with urea alone had greater potential for Cd phytoremediation. In conclusion, the suitable concentration of urea remarkably promoted and assisted the processes of Cd phytoremediation by S. nigrum.
5. Conclusions
Our study concludes that the application of an appropriate dosage of N fertilizers (NH4NO3 and urea) could be a suitable practice to enhance the remediation of heavy-metal-polluted soils when growing S. nigrum. The plant biomass increased significantly with N fertilizer addition. Maximum plant growth was observed in the urea-treated plants compared with the NH4NO3-treated plants. In addition, the urea-treated plants also accumulated a higher Cd than the NH4NO3-treated plants. The application of N fertilizers inhibited oxidative stress and enhanced the antioxidants’ enzymatic activities. It is concluded that urea might be helpful for better growth of S. nigrum under Cd stress. Our results indicate that the best N-fertilizer is urea, and that it is recommended for the phytoremediation of Cd. However, the application of urea for heavy metals phytoremediation other than Cd is still unknown, and future studies can be helpful for this investigation.
Author Contributions
Conceptualization, A.M., S.A. (Shafaqat Ali), T.Y., S.N., S.A. (Saad Alkahtani), and M.M.A.-D.; Data curation, A.M., M.R. (Muhammad Riaz), and A.H.; Formal analysis, A.M., M.R. (Muhammad Rizwan), and A.H.; Funding acquisition, S.A. (Saad Alkahtani) and M.M.A.-D.; Investigation, M.R. (Muhammad Riaz); Methodology, A.M., T.Y., M.R. (Muhammad Riaz), A.H., and S.N.; Project administration, S.A. (Shafaqat Ali) and S.A. (Saad Alkahtani); Resources, S.A. (Saad Alkahtani) and M.M.A.-D.; Software, M.R. (Muhammad Rizwan), M.S.A., and A.H.; Supervision, S.A. (Shafaqat Ali); Validation, M.R. (Muhammad Rizwan) and M.S.A.; Visualization, M.S.A. and S.N.; Writing—original draft, A.M., A.H., and S.N.; Writing—review and editing, S.A. (Shafaqat Ali), T.Y., S.A. (Saad Alkahtani), and M.M.A.-D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was also funded by the Researchers Supporting Project (RSP 2019/26), King Saud University, Riyadh, Saudi Arabia.
Acknowledgments
The authors are highly thankful to the Government College University Faisalabad-38000, Pakistan. This work was also funded by the Researchers Supporting Project (RSP 2019/26), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goldstone, J.A. Demography, environment, and security. Environ. Confl. Routledge 2018, 8, 84–108. [Google Scholar]
- Murtaza, G.; Javed, W.; Hussain, A.; Wahid, A.; Murtaza, B.; Owens, G. Metal uptake via phosphate fertilizer and city sewage in cereal and legume crops in Pakistan. Environ. Sci. Pollut. Res. 2015, 22, 9136–9147. [Google Scholar] [CrossRef] [PubMed]
- Markovic, J.; Jovic, M.; Smiciklas, I.; Sljivic-Ivanovic, M.; Onjia, A.; Trivunac, K.; Popovic, A. Cadmium retention and distribution in contaminated soil: Effects and interactions of soil properties, contamination level, aging time and in situ immobilization agents. Ecotoxicol. Environ. Saf. 2019, 174, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.; Rehman, M.Z.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Gill, R.A.; Yang, S.; Gill, M.B.; Ali, S.; Rafiq, M.T.; Zhou, W. Hydrogen sulfide alleviates cadmium-induced morpho-physiological and ultrastructural changes in Brassica napus. Ecotoxicol. Environ. Saf. 2014, 110, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Khalid, H.; Akmal, F.; Ali, S.; Rizwan, M.; Qayyum, M.F.; Iqbal, M.; Khalid, M.U.; Azhar, M. Effect of limestone, lignite and biochar applied alone and combined on cadmium uptake in wheat and rice under rotation in and effluent irrigated field. Environ. Pollut. 2017, 227, 560–568. [Google Scholar] [CrossRef]
- Gill, R.A.; Ali, B.; Islam, F.; Farooq, M.A.; Gill, M.B.; Mwamba, T.M.; Zhou, W. Physiological and molecular analyses of black and yellow seeded Brassica napus regulated by 5-aminolivulinic acid under chromium stress. Plant Physiol. Biochem. 2015, 94, 130–143. [Google Scholar] [CrossRef]
- Xia, S.; Song, Z.; Jeyakumar, P.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; Wang, H. A critical review on bioremediation technologies for Cr (VI)-contaminated soils and wastewater. Critic. Rev. Environ. Sci. Technol. 2019, 49, 1027–1078. [Google Scholar] [CrossRef]
- Deng, Z.; Cao, L. Fungal endophytes and their interactions with plants in phytoremediation: A review. Chemosphere 2017, 168, 1100–1106. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R.; Juzoń, K.; Borek, M.; Antonkiewicz, J. Photosynthetic response of cabbage in cadmium-spiked soil. Photosynthetica 2019, 57, 731–739. [Google Scholar] [CrossRef]
- Ji, P.H.; Song, Y.F.; Jiang, Y.; Tang, X.; Tong, Y.A.; Gao, P.; Han, W. A two-year field study of phytoremediation using Solanum nigrum L. in China. Int. J. Phytorem. 2016, 18, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.X.; Luo, Q.; Liu, S.L.; Zhao, Y.; Long, Y.; Pan, Y.Z. Screening ornamental plants to identify potential Cd hyperaccumulators for bioremediation. Ecotoxicol. Environ. Saf. 2018, 162, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Wei, S.; Twardowska, I.; Han, R.; Xu, L. Hyperaccumulating potential of Bidens pilosa L. for Cd and elucidation of its translocation behavior based on cell membrane permeability. Environ. Sci. Pollut. 2017, 24, 23161–23167. [Google Scholar] [CrossRef]
- Jacobs, A.; Noret, N.; Van Baekel, A.; Liénard, A.; Colinet, G.; Drouet, T. Influence of edaphic conditions and nitrogen fertilizers on cadmium and zinc phytoextraction efficiency of Noccaea caerulescens. Sci. Total Environ. 2019, 665, 649–659. [Google Scholar] [CrossRef]
- Li, T.Q.; Di, Z.Z.; Yang, X.E.; Sparks, D.L. Effects of dissolved organic matter from the rhizosphere of the hyperaccumulator Sedum alfredii on sorption of zinc and cadmium by different soils. J. Hazard. Mater. 2011, 192, 1616–1622. [Google Scholar] [CrossRef]
- Álvarez-Mateos, P.; Alés-Álvarez, F.J.; García-Martín, J.F. Phytoremediation of highly contaminated mining soils by Jatropha curcas L. and production of catalytic carbons from the generated biomass. J. Environ. Manag. 2019, 231, 886–895. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Y.F.; Chang, S.X.; Yang, Y.F.; Fu, S.L.; Jiang, P.K.; Luo, Y.; Yang, M.; Chen, Z.H.; Hu, S.D.; et al. Biochar reduces soil heterotrophic respiration in a subtropical plantation through increasing soil organic carbon recalcitrancy and decreasing carbon-degrading microbial activity. Soil Biol. Biochem. 2018, 122, 173–185. [Google Scholar] [CrossRef]
- Pan, H.; Xie, K.; Zhang, Q.; Jia, Z.; Xu, J.; Di, H.; Li, Y. Archaea and bacteria respectively dominate nitrification in lightly and heavily grazed soil in a grassland system. Biol. Fertil. Soils 2018, 54, 41–54. [Google Scholar] [CrossRef]
- Xiao, M.L.; Zang, H.D.; Liu, S.L.; Ye, R.Z.; Zhu, Z.K.; Su, Y.R.; Wu, J.S.; Ge, T.D. Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice–soil systems: A 13C-CO2 pulse labeling study. Plant Soil 2019, 445, 101–112. [Google Scholar] [CrossRef]
- Arnamwong, S.; Wu, L.H.; Hu, P.J.; Yuan, C.; Thiravetyan, P.; Luo, Y.M.; Christie, P. Phytoextraction of cadmium and zinc by Sedum plumbizincicola using different nitrogen fertilizers, a nitrification inhibitor and a urease inhibitor. Int. J. Phytorem. 2015, 17, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ding, Y.; Zhang, Q.; Liu, X.; Xu, J.; Li, Y.; Di, H. Heterotrophic nitrification and denitrification are the main sources of nitrous oxide in two paddy soils. Plant Soil 2018, 445, 39–53. [Google Scholar] [CrossRef]
- Yang, Z.B.; Liu, L.X.; Lv, Y.F.; Cheng, Z.; Xu, X.X.; Xian, J.R.; Zhu, X.M.; Yang, Y.X. Metal availability, soil nutrient, and enzyme activity in response to application of organic amendments in Cd-contaminated soil. Environ. Sci. Pollut. Res. 2018, 25, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Yavari, S.; Malakahmad, A.; Sapari, N.B.; Yavari, S. Nutrients balancefor improvement of phytoremediation ability of teak seedlings (Tectona grandis). J. Plant Nutr. 2018, 41, 545–551. [Google Scholar] [CrossRef]
- Liu, Y.L.; Ge, T.D.; Jun, Y.; Liu, S.L.; Shibistova, O.; Wang, P.; Wang, J.K.; Li, Y.; Guggenberger, G.; Kuzyakov, Y.; et al. Initial utilization of rhizodeposits with rice growth in paddy soils: Rhizosphere and N fertilization effects. Geoderma. Res. 2019, 338, 30–39. [Google Scholar] [CrossRef]
- Yang, W.; Dai, H.; Skuza, L.; Wei, S. Strengthening role and the mechanism of optimum nitrogen addition in relation to Solanum nigrum L. Cd hyperaccumulation in soil. Ecotoxicol. Environ. saf. 2019, 182, 109444. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Constable and Co. Ltd.: London, UK, 1962; p. 219. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeny, D.R. Methods of Soil Analysis (Part 2). In Chemical and Microbiological Properties; SSSA: Madison, WI, USA, 1982; Volume 9. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids pigments of photosynthetic biomembranes. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Ecosyst. Environ. Res. 1998, 253, 122–130. [Google Scholar] [CrossRef]
- Zhang, X.Z. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. In Research Methodology of Crop Physiology; Agriculture Press: Beijing, China, 1992; pp. 208–211. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Zhang, R.R.; Liu, Y.; Xue, W.L.; Chen, R.X.; Du, S.T.; Jin, C.W. Slow release nitrogen fertilizers can improve yield and reduce Cd concentration in pakchoi (Brassica chinensis L.) grown in Cd contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 25074–25083. [Google Scholar] [CrossRef]
- Zhou, G.D.; Guo, J.M.; Yang, J.; Yang, J.X. Effect of fertilizers on Cd accumulation and subcellular distribution of two cosmos species (Cosmos sulphureus and Cosmos bipinnata). Int. J. Phytorem. Res. 2018, 20, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Dou, C.; Li, Y.; Wang, H.; Niazi, N.K.; Zhang, S.; Ye, Z. Nitrogen fertilizer enhances zinc and cadmium uptake by hyperaccumulator Sedum alfredii Hance. J. Soils Sediments 2019, 20, 320–329. [Google Scholar] [CrossRef]
- Rabelo, F.H.S.; Azevedo, R.A.; Monteiro, F.A. Proper supply of S increases GSH synthesis in the establishment and reduces tiller mortality during the regrowth of Tanzania guinea grass used for Cd phytoextraction. J. Soils Sediments. Res. 2017, 17, 1427–1436. [Google Scholar] [CrossRef]
- Gouia, H.; Ghorbala, M.H.; Meyer, C. Effects of cadmium on activity of nitrate reductase and on other enzymes of the nitrate assimilation pathway in bean. Plant Physiol. Biochem. Res. 2000, 38, 629–638. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, P.; Kopittke, P.M.; Wang, A.; Sale, P.W.; Tang, C. Cadmium accumulation is enhanced by ammonium compared to nitrate in two hyperaccumulators, without affecting speciation. J. Exp. Bot. 2016, 67, 5041–5050. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Li, Y.; Zhou, Q.; Srivastava, M.; Chiu, S.; Zhan, J.; Wu, Z.; Sun, T. Effect of fertilizer amendments on phytoremediation of Cd-contaminated soil by a newly discovered hyperaccumulator Solanum nigrum L. J. Hazard. Mater. 2010, 176, 269–273. [Google Scholar] [CrossRef]
- Ye, X.; Hu, H.; Li, H.; Xiong, Q.; Gao, H. Combined nitrogen fertilizer and wheat straw increases the cadmium phytoextraction efficiency of Tagetes patula. Ecotoxicol. Environ. Saf. 2019, 170, 210–217. [Google Scholar] [CrossRef]
- Yang, W.; Dai, H.; Dou, X.; Zhang, Q.; Wei, S. Effect and mechanism of commonly used four nitrogen fertilizers and three organic fertilizers on Solanum nigrum L. hyperaccumulating Cd. Environ. Sci. Pollut. Res. 2019, 26, 12940–12947. [Google Scholar] [CrossRef]
- Hassan, M.J.; Zhang, G.; Zhu, Z. Influence of cadmium toxicity on plant growth and nitrogen uptake in rice as affected by nitrogen form. J. Plant Nutr. 2008, 3, 251–262. [Google Scholar] [CrossRef]
- Sanita, L.; Di Toppi, S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar] [CrossRef]
- Kastori, R.; Petrovic, N.; Arsenijevic-Maksimovic, I. Heavy metals and plants. In Heavy Metals in the Environment; Kastori, R., Ed.; Feljton: Novi Sad, Serbia, 1997; pp. 195–257. [Google Scholar]
- Pakovic, D.; Plesnicar, M.; Arsenijevic-Marksimovic, I.; Petrovic, N. Effects of nitrogen nutrition on photosynthesis in Cd-treated plants. Ann. Bot. 2000, 86, 841–847. [Google Scholar] [CrossRef]
- Foyer, C.H.; Descourviers, P.; Kunert, K.J. Protection against oxygen radicals: An important defense mechanism studied in transgenic plants. Plant Cell Environ. Res. 1994, 17, 507–523. [Google Scholar] [CrossRef]
- Jalloh, M.A.; Chen, J.; Zhen, F.; Zhang, G. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard. Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef]
- Malmir, H.A. Comparison of antioxidant enzyme activities in leaves stems and roots of Sorghum (Sorghum bicolor L.) exposed to Chromium (VI). Afr. J. Plant Sci. 2011, 5, 436–444. [Google Scholar]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Pandey, V.; Dixit, V.; Shyam, R. Antioxidative responses in relation to growth of mustard (Brassica juncea cv. Pusa Jaikisan) plants exposed to hexavalent chromium. Chemosphere 2005, 61, 40–47. [Google Scholar] [CrossRef]
- Mahmud, J.A.; Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Fujita, M. EDTA reduces cadmium toxicity in mustard (Brassica juncea L.) by enhancing metal chelation, antioxidant defense and glyoxalase systems. Acta Agrobot. 2019, 72, 1722. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L.: An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Nahar, K.; Rahman, M.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Physiological and biochemical mechanisms of spermine-induced cadmium stress tolerance in mung bean (Vigna radiata L.) seedlings. Environ. Sci. Pollut. Res. 2016, 23, 21206–21218. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Exogenous selenium pretreatment protects rapeseed seedlings from cadmium-induced oxidative stress by upregulating antioxidant defense and methylglyoxal detoxification systems. Biol. Trace Element Res. 2012, 149, 248–261. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).