Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds


Abstract
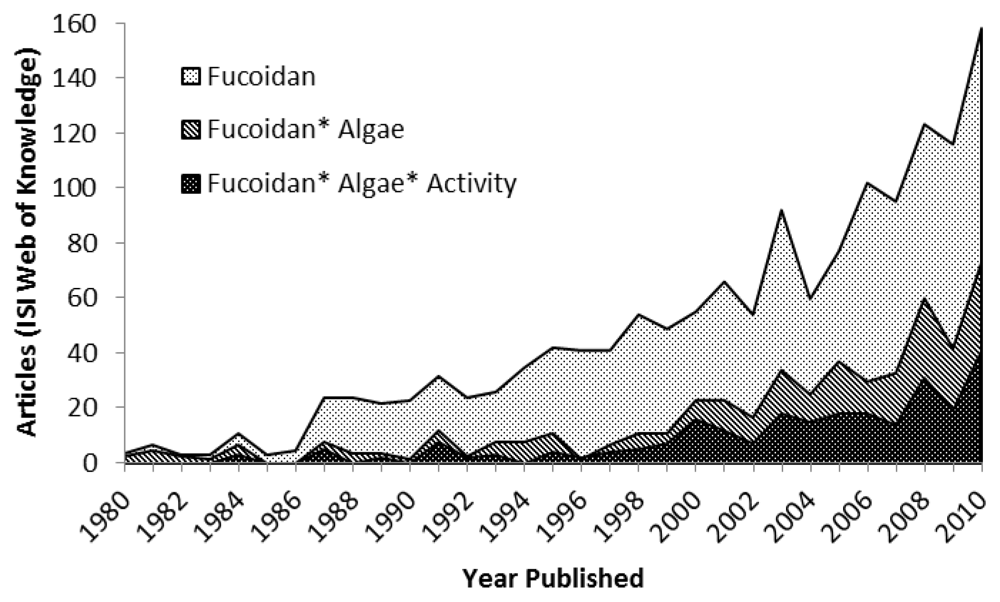
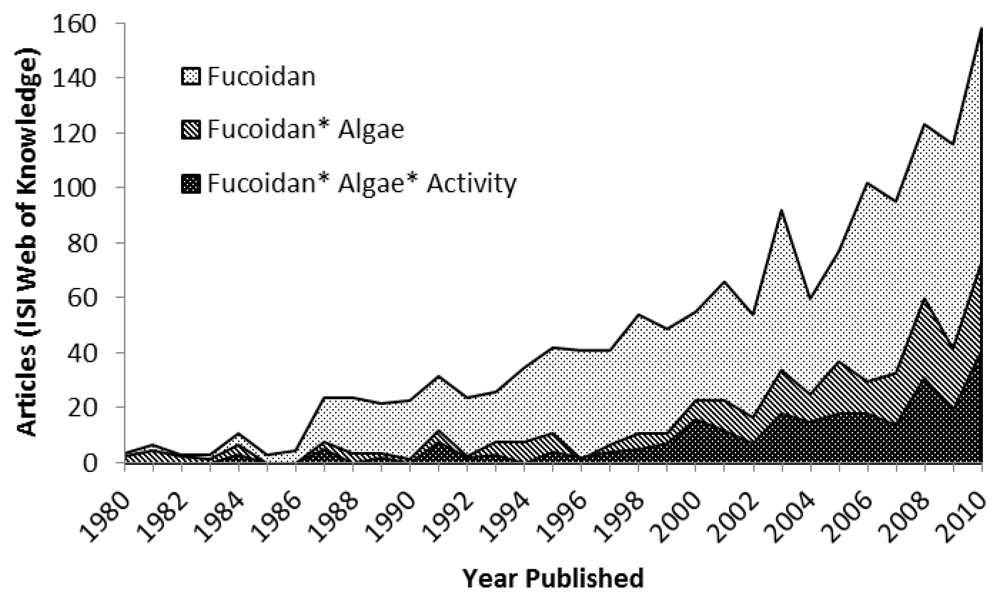
:1. Introduction
2. Historic Overview: FCSPs Extraction Procedures and Chemical Analyses
3. Taxonomic Comparison of Fucoidan or FCSPs Structure
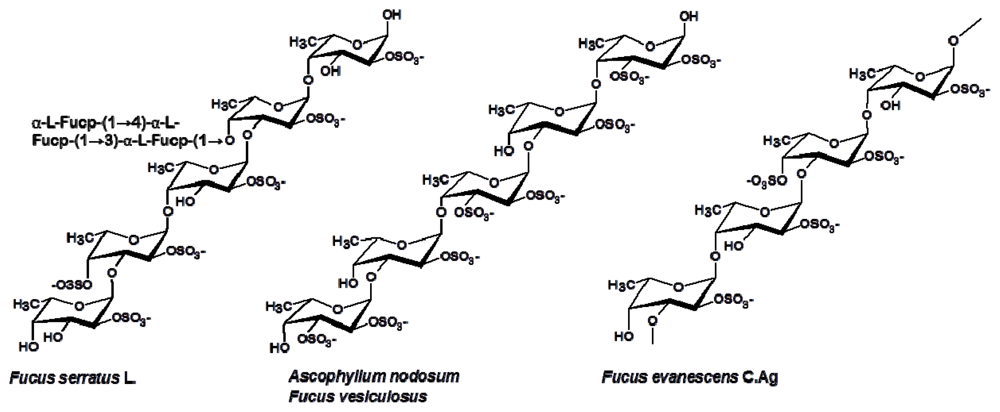
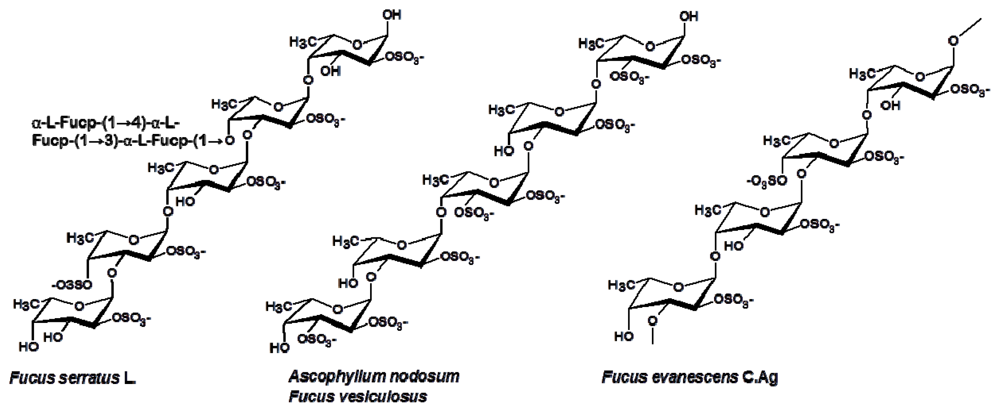
3.1. Fucales
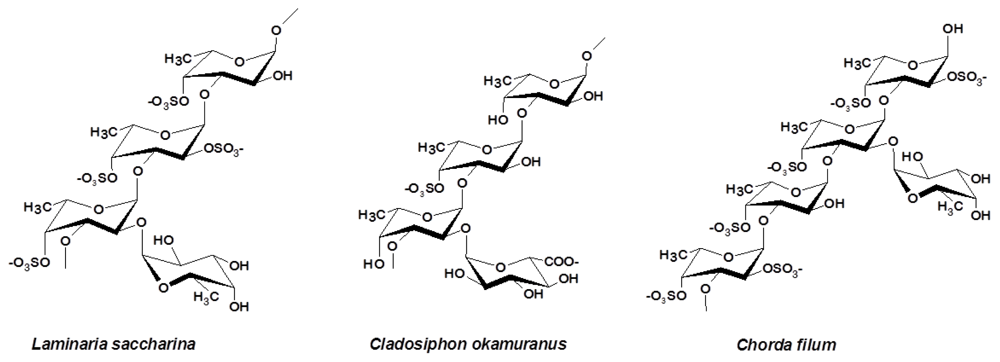
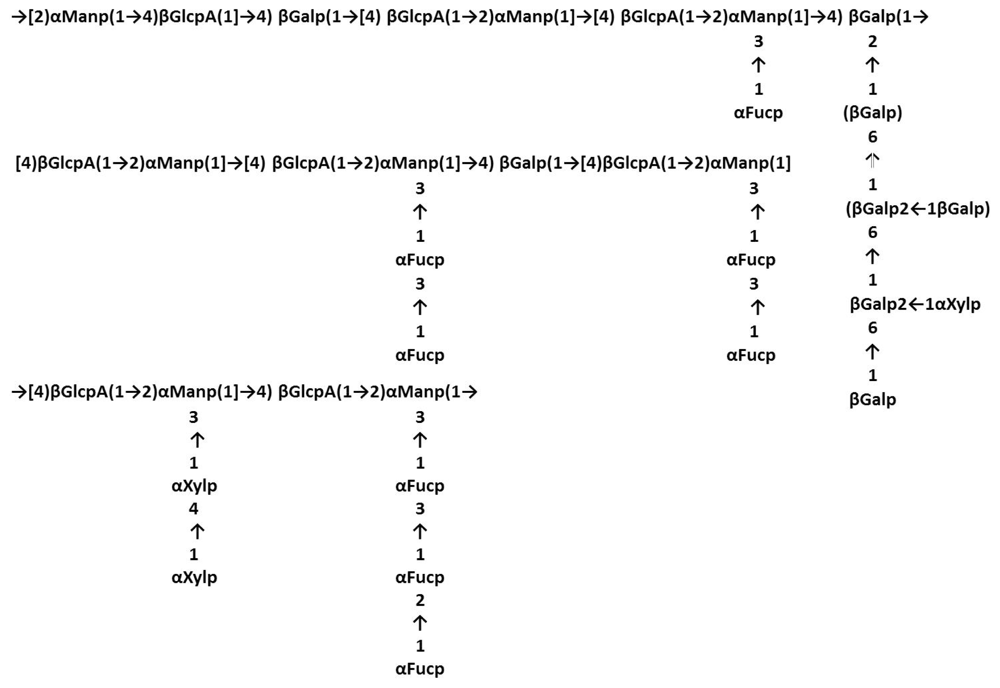
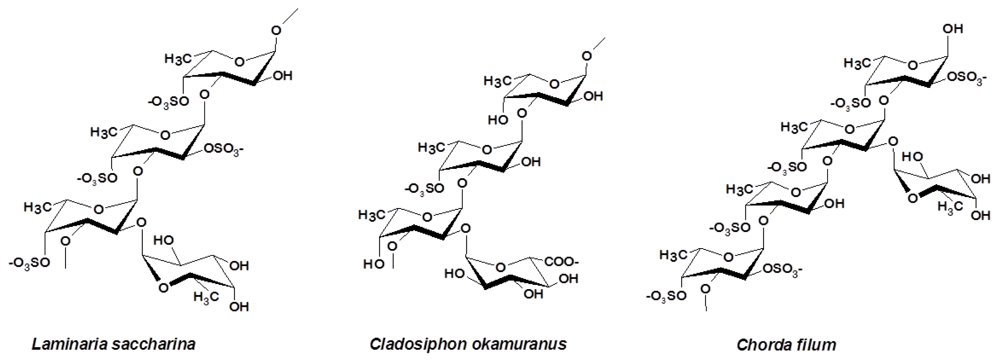
3.2. Laminariales and Other Brown Seaweed
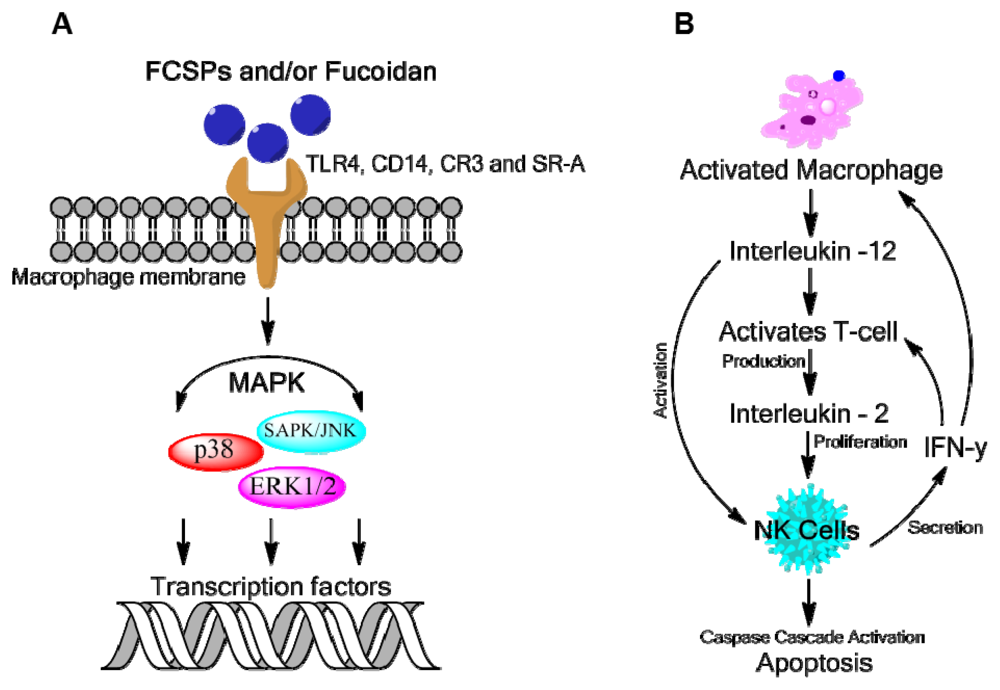
4. Bioactivity of Fucoidan or FCSPs
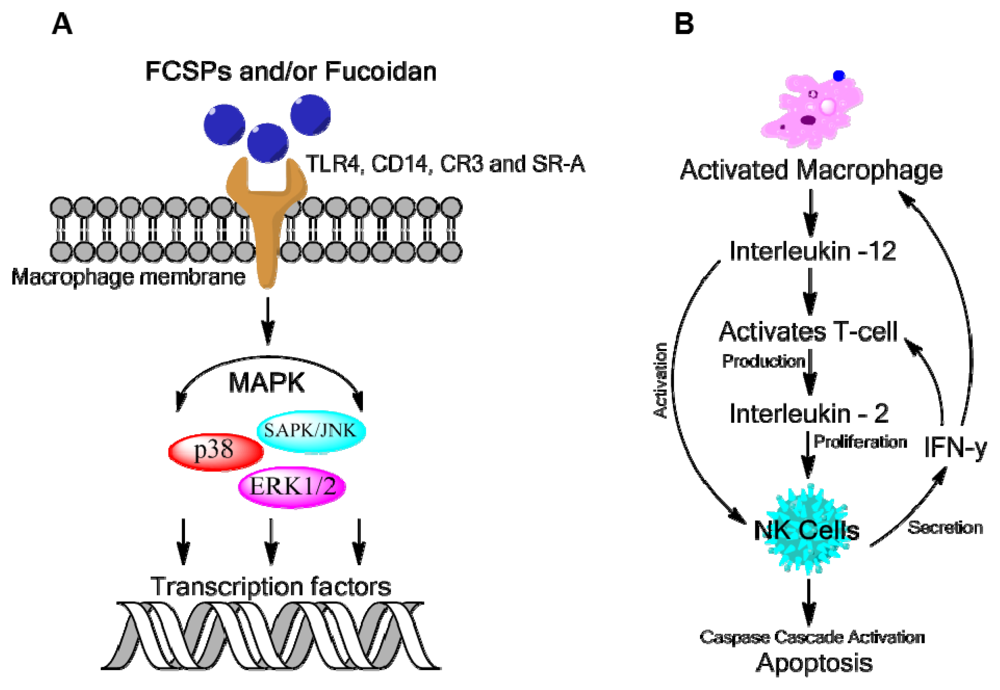
4.1. Antitumor and Immune-Response Activities
4.2. Anticoagulant and Antithrombotic Activities
4.3. Bioactivities and Oversulfation of FCSPs
5. Conclusions
References
- Jiao, G; Yu, G; Zhang, J; Ewart, SE. Chemical structure and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar]
- Kylin, H. Biochemistry of sea algae. H. Z. Physiol. Chem 1913, 83, 171–197. [Google Scholar]
- Kylin, H. Analysis of the biochemistry of the seaweed. H. Z. Physiol. Chem 1915, 94, 337–425. [Google Scholar]
- Hoagland, DR; Lieb, LL. The complex carbohydrates and forms of sulphur in marine algae of the Pacific coast. J. Biol. Chem 1915, 23, 287–297. [Google Scholar]
- Bird, GM; Hass, P. On the nature of the cell wall constituents of Laminaria sp. mannuronic acid. Biochem. J 1931, 25, 403–411. [Google Scholar]
- Nelson, WL; Cretcher, LH. The carbohydrate acid sulfate of Macrocystis pyrifera. J. Biol. Chem 1931, 94, 147–154. [Google Scholar]
- Lunde, G; Heen, E; Oy, E. Uber fucoidin. H. Z. Physiol. Chem 1937, 247, 189–196. [Google Scholar]
- Percival, EGV; Ross, AG. Fucoidin. Part 1. The isolation and purification of fucoidin from brown seaweeds. J Chem Soc 1950, 717–720. [Google Scholar]
- Black, WAP; Dewar, ET; Woodward, FN. Manufacturing of algal chemicals 4: Laboratory scale isolation of fucoidan from brown marine algae. J. Sci. Food Agric 1952, 3, 122–129. [Google Scholar]
- Conchie, J; Percival, EGV. Fucoidin. Part II. The hydrolysis of a methylated fucoidin prepared from Fucus vesiculosus. J Chem Soc 1950, 827–832. [Google Scholar]
- Chevolot, L; Foucault, A; Chaubet, F; Kervarec, N; Sinquin, C; Fisher, AM; Boisson-Vidal, C. Further data on the structure of brown seaweed fucans: Relationships with anticoagulant activity. Carbohydr. Res 1999, 319, 154–165. [Google Scholar]
- Mian, J; Percival, E. Carbohydrates of the brown seaweeds Himanthalia lorea and Bifurcaria bifurcate Part II. Structural studies of “fucans”. Carbohydr. Res 1973, 26, 147–161. [Google Scholar]
- Abdel-fattah, AF; Hussein, MD; Salem, HM. Studies of purification and some properties of Sargassan, a sulfated heteropolysaccharide from Sargassum linifolium. Carbohydr. Res 1974, 33, 9–17. [Google Scholar]
- Nagaoka, M; Shibata, H; Kimura-Takagi, I; Hashimoto, S; Kimura, K; Makino, T; Aiyama, R; Ueyama, S; Yokokura, T. Structural study of fucoidan from Cladosiphon okamuranus TOKIDA. Glycoconj. J 1999, 16, 19–26. [Google Scholar]
- Ponce, NM; Pujol, CA; Damonte, EB; Flores, ML; Stortz, CA. Fucoidans from the brown seaweed Adenocystis utricularis: Extraction methods, antiviral activity and structural studies. Carbohydr. Res 2003, 338, 153–165. [Google Scholar]
- Marais, MF; Joseleau, JP. A fucoidan fraction from Ascophyllum nodosum. Carbohydr. Res 2001, 336, 155–159. [Google Scholar]
- Percival, E. Glucuronoxylofucan, a cell-wall component of Ascophyllum nodosum. Carbohydr. Res 1968, 7, 272–277. [Google Scholar]
- Duarte, ME; Cardoso, MA; Noseda, MD; Cerezo, AS. Structural studies on fucoidans from the brown seaweed Sargassum stenophyllum. Carbohydr. Res 2001, 333, 281–293. [Google Scholar]
- Ale, MT; Mikkelsen, JD; Meyer, AS. Designed optimization of a single-step extraction of fucose-containing sulfated polysaccharides from Sargassum sp. J Appl Phycol 2011. [Google Scholar] [CrossRef]
- Bilan, MI; Grachev, AA; Ustuzhanina, NE; Shashkov, AS; Nifantiev, NE; Usov, AI. Structure of a fucoidan from brown seaweed Fucus evanescens. Carbohydr. Res 2002, 337, 719–730. [Google Scholar]
- Cumashi, A; Ushakova, NA; Preobrazhenskaya, ME; D’Incecco, A; Piccoli, A; Totani, L; Tinari, N; Morozevich, GE; Berman, AE; Bilan, MI; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 5, 541–552. [Google Scholar]
- Bilan, MI; Grachev, AA; Shashkov, AS; Nifantiev, NE; Usov, AI. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohydr. Res 2006, 341, 238–245. [Google Scholar]
- Li, B; Wei, XJ; Sun, JL; Xu, SY. Structural investigation of a fucoidan containing a fucose-free core from the brown seaweed, Hizikia fusiforme. Carbohydr. Res 2006, 341, 1135–1146. [Google Scholar]
- Usov, AI; Smirnova, GP; Bilan, MI; Shashkov, AS. Polysaccharides of algae: 53. Brown alga Laminaria saccharina (L.) Lam. as a source of fucoidan. Bioorg. Khim 1998, 24, 382–389. [Google Scholar]
- Chizhov, AO; Dell, A; Morris, HR; Haslam, SM; McDowell, RA; Shashkov, AS; Nifant’ev, NE; Khatuntseva, EA; Usov, AI. A study of fucoidan from the brown seaweed Chorda filum. Carbohydr. Res 1999, 320, 108–119. [Google Scholar]
- Hemmingson, JA; Falshow, R; Furneaux, RH; Thompsom, K. Structure and antiviral activity of the galactofucans sulfates extracted from Undaria pinnatifida (Phaeophyta). J. Appl. Phycol 2006, 18, 185–193. [Google Scholar]
- Maruyama, H; Yamamoto, I. An antitumor fraction from an edible brown seaweed Laminaria religiosa. Hydrobiologia 1984, 116/177, 534–536. [Google Scholar]
- Rocha, HAO; Moraes, FA; Trindade, ES; Franco, CRC; Torquato, RJS; Veiga, SS; Valente, AP; Mourao, PAS; Leite, EL; Nader, HB; et al. Structural and hemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schroederi—An ideal antithrombotic agent? J. Biol. Chem 2005, 280, 41278–41288. [Google Scholar]
- O’Neill, AN. Degradative studies on fucoidin. J. Am. Chem. Soc 1954, 76, 5074–5076. [Google Scholar]
- Côte, RH. Disaccharides from fucoidin. J Chem Soc 1959, 2248–2254. [Google Scholar]
- Patankar, MS; Oehninger, S; Barnett, T; Williams, RL; Clark, GF. A revised structure for fucoidan may explain some of its biological activities. J. Biol. Chem 1993, 268, 21770–21776. [Google Scholar]
- Pereira, MS; Mulloy, B; Mourão, PA. Structure and anticoagulant activity of sulfated fucans. Comparison between the regular, repetitive, and linear fucans from echinoderms with the more heterogeneous and branched polymers from brown algae. J. Biol. Chem 1999, 274, 7656–7667. [Google Scholar]
- Anastyuk, SD; Shevchenko, NM; Nazarenko, EL; Dmitrenok, PS; Zvyagintseva, TN. Structural analysis of a fucoidan from the brown alga Fucus evanescens by MALDI-TOF and tandem ESI mass spectrometry. Carbohydr. Res 2009, 21, 779–787. [Google Scholar]
- Bilan, MI; Usov, AI. Structural analysis of fucoidans. Nat. Prod. Commun 2008, 3, 1639–1648. [Google Scholar]
- Zhu, W; Ooi, VEC; Chan, PKS; Ang, PO. Isolation and characterization of a sulfated polysaccharide from the brown alga Sargassum patens and determination of its anti-herpes activity. Biochem. Cell Biol 2003, 81, 25–33. [Google Scholar]
- Berteau, O; Mulloy, B. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29–40. [Google Scholar]
- Ribeiro, AC; Vieira, RP; Mourão, PA; Mulloy, B. A sulfated alpha-l-fucan from sea cucumber. Carbohydr. Res 1994, 255, 225–240. [Google Scholar]
- Chandía, NP; Matsuhiro, B. Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol 2008, 42, 235–240. [Google Scholar]
- Bilan, MI; Zakharova, AN; Grachev, AA; Shashkov, AS; Nifantiev, NE; Usov, AI. Polysaccharides of algae: 60. Fucoidan from the pacific brown alga Analipus japonicus (Harv.) winne (Ectocarpales, Scytosiphonaceae). Russ. J. Bioorg. Chem 2007, 33, 38–46. [Google Scholar]
- Alekseyenko, TV; Zhanayeva, SY; Venediktova, AA; Zvyagintseva, TN; Kuznetsova, TA; Besednova, NN; Korolenko, TA. Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk Sea Fucus evanescens brown alga. Bull. Exp. Biol. Med 2007, 143, 730–732. [Google Scholar]
- Maruyama, H; Tamauchi, H; Iizuka, M; Nakano, T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta Med 2006, 72, 1415–1417. [Google Scholar]
- Ale, MT; Maruyama, H; Tamauchi, H; Mikkelsen, JD; Meyer, A. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol 2011, 49, 331–336. [Google Scholar]
- Makarenkova, ID; Deriabin, PG; L’vov, DK; Zviagintseva, TN; Besednova, NN. Antiviral activity of sulfated polysaccharide from the brown algae Laminaria japonica against avian influenza A (H5N1) virus infection in the cultured cells. Vopr. Virusol 2010, 55, 41–45. [Google Scholar]
- Zhu, Z; Zhang, Q; Chen, L; Ren, S; Xu, P; Tang, Y; Luo, D. Higher specificity of the activity of low molecular weight fucoidan for thrombin-induced platelet aggregation. Thromb. Res 2010, 125, 419–426. [Google Scholar]
- Semenov, AV; Mazurov, AV; Preobrazhenskaia, ME; Ushakova, NA; Mikhaı̆lov, VI; Berman, AE; Usov, AI; Nifant’ev, NE; Bovin, NV. Sulfated polysaccharides as inhibitors of receptor activity of P-selectin and P-selectin-dependent inflammation. Vopr Med Khim 1998, 44, 135–144. (in Russian). [Google Scholar]
- Wang, J; Zhang, Q; Zhang, Z; Song, H; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol 2010, 46, 6–12. [Google Scholar]
- Veena, CK; Josephine, A; Preetha, SP; Varalakshmi, P; Sundarapandiyan, R. Renal peroxidative changes mediated by oxalate: The protective role of fucoidan. Life Sci 2006, 79, 1789–1795. [Google Scholar]
- Hayakawa, K; Nagamine, T. Effect of fucoidan on the biotinidase kinetics in human hepatocellular carcinoma. Anticancer Res 2009, 4, 1211–1217. [Google Scholar]
- Zhang, Q; Li, N; Zhao, T; Qi, H; Xu, Z; Li, Z. Fucoidan inhibits the development of proteinuria in active Heymann nephritis. Phytother. Res 2005, 19, 50–53. [Google Scholar]
- Hlawaty, H; Suffee, N; Sutton, A; Oudar, O; Haddad, O; Ollivier, V; Laguillier-Morizot, C; Gattegno, L; Letourneur, D; Charnaux, N. Low molecular weight fucoidan prevents intimal hyperplasia in rat injured thoracic aorta through the modulation of matrix metalloproteinase-2 expression. Biochem. Pharmacol 2011, 81, 233–243. [Google Scholar] [Green Version]
- Cashman, JD; Kennah, E; Shuto, A; Winternitz, C; Springate, CM. Fucoidan film safely inhibits surgical adhesions in a rat model. J Surg Res 2011. [Google Scholar] [CrossRef]
- Schneider, U; Stipper, A; Besserer, J. Dose-response relationship for lung cancer induction at radiotherapy dose. Z. Med. Phys 2010, 20, 206–214. [Google Scholar]
- Grossi, F; Kubota, K; Cappuzzo, F; de Marinis, F; Gridelli, C; Aita, M; Douillard, JY. Future scenarios for the treatment of advanced non-small cell lung cancer: Focus on taxane-containing regimens. Oncologist 2010, 15, 1102–1112. [Google Scholar]
- Cho, ML; Lee, BY; You, SG. Relationship between oversulfation and conformation of low and high molecular weight fucoidans and evaluation of their in vitro anticancer activity. Molecules 2011, 16, 291–297. [Google Scholar]
- Synytsya, A; Kim, WJ; Kim, SM; Pohl, R; Synytsya, A; Kvasnicka, F; Copikova, J; Park, YI. Structure and antitumor activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydr. Polym 2010, 81, 41–48. [Google Scholar]
- Yamamoto, I; Nagumo, T; Yagi, K; Tominaga, H; Aoki, M. Antitumor effect of seaweeds, 1. Antitumor effect of extract from Sargasum and Laminaria. Jpn. J. Exp. Med 1974, 44, 543–546. [Google Scholar]
- Yamamoto, I; Nagumo, T; Takahashi, M; Fujihara, M; Suzuki, Y; Iizima, N. Antitumor effect of seaweeds, 3. Antitumor effect of an extract from Sargassum kjellmanianum. Jpn. J. Exp. Med 1981, 51, 187–189. [Google Scholar]
- Yamamoto, I; Takahashi, M; Tamura, E; Maruyama, H; Mori, H. Antitumor activity of edible marine algae: Effect of crude fucoidan fractions prepared from edible brown seaweed against L-1210 leukemia. Hydrobiology 1984, 116/117, 145–148. [Google Scholar]
- Maruyama, H; Tamauchi, H; Hashimoto, M; Nakano, T. Antitumor activity and immune response of Mekabu fucoidan extracted from Sporophyll of Undaria pinnatifida. Vivo 2003, 17, 245–249. [Google Scholar]
- Takahashi, M. Studies on the mechanism of host mediated antitumor action of fucoidan from a brown alga Eisenia bicyclis. J. Jpn. Soc. Reticuloendothel. Syst 1983, 22, 269–283. [Google Scholar]
- Teruya, T; Tatemoto, H; Konishi, T; Tako, M. Structural characteristics and in vitro macrophage activation of acetyl fucoidan from Cladosiphon okamuranus. Glycoconj. J 2009, 26, 1019–1028. [Google Scholar]
- Aisa, Y; Miyakawa, Y; Nakazato, T; Shibata, H; Saito, K; Ikeda, Y; Kizaki, M. Fucoidan induces apoptosis of human HS-sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol 2005, 78, 7–14. [Google Scholar]
- Whiteside, TL; Herberman, RB. The role of natural killer cells in human disease. Clin. Immunol. Immunopathol 1989, 53, 1–23. [Google Scholar]
- Raghavendran, HR; Srinivasan, P; Rekha, S. Immunomodulatory activity of fucoidan against aspirin-induced gastric mucosal damage in rats. Int. Immunopharmacol 2011, 11, 157–163. [Google Scholar]
- Kim, EJ; Park, SY; Lee, JY; Park, JH. Fucoidan present in brown algae induces apoptosis of human colon cancer cells. BMC Gastroenterol 2010, 10, 96. [Google Scholar]
- Yamasaki-Miyamoto, Y; Yamasaki, M; Tachibana, H; Yamada, K. Fucoidan induces apoptosis through activation of caspase-8 on human breast cancer MCF-7 cells. J. Agric. Food Chem 2009, 57, 8677–8682. [Google Scholar]
- Springer, GF; Wurzel, HA; McNeal, GM; Ansell, NJ; Doughty, MF. Isolation of anticoagulant fractions from crude fucoidin. Proc. Soc. Exp. Biol. Med 1957, 94, 404–409. [Google Scholar]
- Grauffel, V; Kloareg, B; Mabeau, S; Durand, P; Jozefonvicz, J. New natural polysaccharides with potent antithrombic activity: Fucans from brown algae. Biomaterials 1989, 10, 363–368. [Google Scholar]
- Nishino, T; Nagumo, T. The sulfate-content dependence of the anticoagulant activity of a fucan sulfate from the brown seaweed Ecklonia kurome. Carbohydr. Res 1991, 214, 193–197. [Google Scholar]
- Nishino, T; Kiyohara, H; Yamada, H; Nagumo, T. An anticoagulant fucoidan from the brown seaweed Ecklonia kurome. Phytochemistry 1991, 30, 535–539. [Google Scholar]
- Nishino, T; Nishioka, C; Ura, H; Nagumo, T. Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydr. Res 1994, 255, 213–224. [Google Scholar]
- Mourão, PA; Pereira, MS. Searching for alternatives to heparin: Sulfated fucans from marine invertebrates. Trends Cardiovasc. Med 1999, 9, 225–232. [Google Scholar]
- Nishino, T; Nagumo, T. Sugar constituents and blood coagulant activities of fucose-containing sulfated polysaccharide in nine brown seaweed species. Nippon Nog. Kaishi 1987, 61, 361–363. [Google Scholar]
- Kitamura, K; Matsuo, M; Yasui, T. Enzymic degradation of fucoidan by fucoidanase from the hepatopancreas of Patinopecten yessoensis. Biosci. Biotechnol. Biochem 1992, 56, 490–494. [Google Scholar]
- Nishino, T; Yokoyama, G; Dobashi, K; Fujihara, M; Nagumo, T. Isolation, purification, and characterization of fucose-containing sulfated polysaccharides from the brown seaweed Ecklonia kurome and their blood-anticoagulant activities. Carbohydr. Res 1989, 186, 119–129. [Google Scholar]
- Nishino, T; Aizu, Y; Nagumo, T. The influence of sulfate content and molecular weight of a fucan sulfate from the brown seaweed Ecklonia kurome on its antithrombin activity. Thromb. Res 1991, 64, 723–731. [Google Scholar]
- Pomin, VH; Pereira, MS; Valente, AP; Tollefsen, DM; Pavao, MSG; Mourao, PAS. Selective cleavage and anticoagulant activity of a sulfated fucan: Stereospecific removal of a 2-sulfate ester from the polysaccharide by mild acid hydrolysis, preparation of oligosaccharides, and heparin cofactor II-dependent anticoagulant activity. Glycobiology 2005, 15, 369–381. [Google Scholar]
- Haroun-Bouhedja, F; Ellouali, M; Sinquin, C; Boisson-Vidal, C. Relationship between sulfate groups and biological activities of fucans. Thromb. Res 2000, 100, 453–459. [Google Scholar]
- Dobashi, K; Nishino, T; Fujihara, M; Nagumo, T. Isolation and preliminary characterization of fucose-containing sulfated polysaccharides with blood-anticoagulant activity from the brown seaweed Hizikia fusiforme. Carbohydr. Res 1989, 194, 315–320. [Google Scholar]
- Pereira, MS; Melo, FR; Mourão, PAS. Is there a correlation between structure and anticoagulant action of sulfated galactans and sulfated fucans? Glycobiology 2002, 12, 573–580. [Google Scholar]
- Pereira, MS; Vilela-Silva, AES; Valente, A; Mourão, PAS. A 2-sulfated, 3-linked α-l-galactan is an anticoagulant polysaccharide. Carbohydr. Res 2002, 337, 2231–2238. [Google Scholar]
- Li, B; Lu, F; Wei, X; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Mourão, PAS. Use of sulfated fucans as anticoagulant and antithrombotic agents: Future perspectives. Curr. Pharmaceut. Des 2004, 10, 967–981. [Google Scholar]
- Boisson-Vidal, C; Chaubet, F; Chevolot, L; Sinquin, C; Theveniaux, J; Millet, J; Sternberg, C; Mulloy, B; Fischer, AM. Relationship between antithrombotic activities of fucans and their structure. Drug Dev. Res 2000, 51, 216–224. [Google Scholar]
- Qiu, XD; Amarasekara, A; Doctor, V. Effect of oversulfation on the chemical and biological properties of fucoidan. Carbohydr. Polym 2006, 63, 224–228. [Google Scholar]
- Soeda, S; Ishia, S; Shimeno, H; Nagamatsu, A. Inhibitory effect of oversulfated fucoidan on invasion through reconstituted basement membrane by murine Lewis lung carcinoma. Jpn. J. Cancer Res 1994, 85, 1144–1150. [Google Scholar]
- Soeda, S; Nobuaki, F; Shimeno, H; Nagamatsu, A. Oversulfated fucoidan and heparin suppress endotoxin induction of plasminogen activator inhivitor-1 in cultured human endothelial cells: Their possible mechanism of action. Biochim. Biophys. Acta 1995, 1269, 85–90. [Google Scholar]
- Koyanagi, S; Tanigawa, N; Nakagawa, H; Soeda, S; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol 2003, 65, 173–179. [Google Scholar]
- Teruya, T; Konishi, T; Uechi, S; Tamaki, H; Tako, M. Anti-proliferative activity of oversulfated fucoidan from commercially cultured Cladosiphon okamuranus TOKIDA in U937 cells. Int. J. Biol. Macromol 2007, 41, 221–226. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Brown seaweed sp. | Reported FCSPs composition | Extraction method | References |
|---|---|---|---|---|
| 1913 | Laminaria and Fucus | Fucoidan contains fucose, that occurs together with mannitol, alginate and laminaran | Dilute acetic acid extraction | Kylin, 1913 [2] |
| 1915 | Laminaria digitata | Fucoidan contains l-fucose and other pentoses | Dilute acetic acid extraction | Kylin, 1915 [3] |
| 1915 | Macrocystis pyrifera | Mainly alginic acid, with some fucose-sulfate | Soaking in 2% Na2CO3 for 24 h, filtration, HCl addition, recovery of precipitate by filtration, redissolution in 2% Na2CO3 | Hoagland and Lieb, 1915 [4] |
| 1931 | Laminaria digitata | Substantial amounts of calcium sulfate and uronic acid | Soaking of the seaweed in water, precipitation of crude, sulfated polysaccharides by ethanol | Bird and Haas, 1931 [5] |
| 1931 | Macrocystis pyrifera | Methylpentose monosulphate polymer with fucose and alginate contaminants | Repeated extraction with 2% HCl at room temperature for 48 h, precipitated with 90% ethanol | Nelson and Cretcher, 1931 [6] |
| 1937 | Laminaria digitata | Proposed (R-R′-O-SO2-OM)n with R = fucose, R′ as unknown, M being Na, K, Ca0.5, or Mg0.5 | Precipitation of droplets exuded from seaweed in boiling ethanol | Lunde et al., 1937 [7] |
| 1950 | Fucus vesiculosus, Fucus spirales, Himanthalia lorea, Laminaria clustoni | Substantial amounts of fucose and sulfate; small amounts of uronic acid, galactose and xylose; metals and ash were also detected, ash was mainly calcium sulfate | Aqueous extraction at ~100 °C for 24 h, extract treated with lead acetate (to precipitate alginate and proteins), filtrate solution treated with Ba(OH)2 to precipitate a “hydroxide-fucoidin complex” | Percival and Ross, 1950 [8] |
| 1952 | Fucus vesiculosus | Fucose, ash, sulfate | 0.1 M HCl at pH 2–2.5 and 70 °C for 1 h, 3-times, fractional precipitation with ethanol | Black et. al., 1952 [9] |
| Species | Order | Extraction method | Composition | Reference |
|---|---|---|---|---|
| Cladosiphon okamuranus | Chordariales | Seaweed-H2O suspension was treated with 30% HCl (pH 3) at 100 °C for 15 min. Supernatant was neutralized with NaOH, precipitated with CaCl2 and EtOH for 20 h at 4 °C, precipitate was dissolved with H2O then dried | fucose, glucose, uronic acid and sulfate | Nagaoka et al., 1999 [14] |
| Adenocystis utricularis | Ectocapales | 80% EtOH, 24 h, 70 °C pretreatment then extracted with water (or 2% CaCl2; or HCl) for 7 h at rt, followed by exhaustive extraction at 70 °C | fucose, rhamnose, glucose, galactose, xylose, mannose, uronic acid and sulfate | Ponce et al., 2003 [15] |
| Himanthalia lorea | Fucales | Acid + alkali + water-acid-alkali sequence in 70 °C, 4 h. | fucose, xylose, uronic acid, sulfate | Mian and Percival, 1973 [12] |
| Ascophyllum nodosum | Fucales | Extracted at rt and then 70 °C with 0.01 NaCl containing 1% CaCl | fucose, xylose, galactose, glucose, sulfate | Marais and Joseleau, 2001 [16] |
| Fucales | Extracted with hot water and dilute alkali, formaldehyde treatment, then extracted with ammonium oxalate-oxalic acid for 6 h at 80 °C | fucose, xylose, uronic acid sulfate | Percival, 1968 [17] | |
| Sargassum stenophyllum | Fucales | Extracted with water 7% w/v mL, 12 h, 3×. Precipitated with EtOH and CaCl2 and cetylpyridinium chloride. Soluble fraction (SF) was then fractionated (F1–F6) | fucose, xylose, mannose, galactose, glucose, sulfate and uronic acid | Duarte et al., 2001 [18] |
| Sargassum sp. | Fucales | Extracted with 0.03 M HCl at 90 °C for 4 h, single-step | Fucose, rhamnose, galactose, glucose, mannose, xylose, uronic acid, sulfate | Ale et al., 2011 [19] |
| Sargassum linifolium | Fucales | Extracted with water at pH 1 (HCl), for 3 h at 80 °C | mannose, galactose, xylose, uronic acid and fucose residues | Abel-fattah et al., 1974 [13] |
| Fucus evanescens; Fucus distichus | Fucales | Pretreatment: MeOH–CHCl3–H2O (4:2:1), then extracted 2% CaCl2 for 5 h at 85 °C, precipitated and the precipitate was washed with water, stirred with 20% ethanolic solution and dissolved with water [20] | fucose, xylose, galactose, uronic acid and sulfate | Cumashi et al., 2007 [21] |
| Fucus serratus | Fucales | Pretreatment: MeOH–CHCl3–H2O (4:2:1), then extracted 2% CaCl2 for 5 h at 85 °C, the extracts were collected by centrifugation, combined, dialyzed and lyophilized [22] | fucose, xylose, mannose, glucose, galactose, uronic acid and sulfate | Cumashi et al., 2007 [21] |
| Hizikia fusiforme | Fucales | Powdered seaweed was extracted with H2O (1:10), 3×, 2 h at 70 °C, precipitated with EtOH and CaCl2 then dried | fucose, mannose, galactose, xylose, glucose, rhamnose, arabinose, uronic acid and sulfate | Li et al., 2006 [23] |
| Laminaria saccharina; Laminaria digitata; F. vesiculosus; F. spiralis Ascophyllum nodosum | Laminariales and Fucales | Extracted with 2% CaCl2 for 5 h at 85 °C, precipitated with Cetavlon, transformation of Cetavlonic salts into calcium salts, and an alkaline treatment to remove acetyl groups and to transform fucoidan into sodium salts [24] | fucose, xylose, mannose, glucose, galactose, uronic acid and sulfate | Cumashi et al., 2007 [21] |
| Chorda filum | Laminariales | Extracted with CHCl3–MeOH–H2O (2:4:1) followed by 80% EtOH, then extracted successively with 2% CaCl2 at 20 and 70 °C, then with HCl (pH 2) and 3% Na2CO3, precipitated with calcium salt | fucose, xylose, mannose, glucose, galactose, uronic acid and sulfate | Chizhov et al., 1999 [25] |
| Undaria pinnatifida | Laminariales | Ground seaweed extracted twice at rt for 6 h with 1% H2SO4, neutralized with 10% NaOH and lyophilized | fucose, mannose, xylose, rhamnose, galactose, glucose and sulfate | Hemmingson et al., 2006 [26] |
| Laminaria religiosa | Laminariales | Water extraction at boiling temp. for 4 h, fucoidan fraction was obtained by using 0.09 HCl at 4 °C for 2 h, then precipitated with 85% EtOH and dried | fucose, xylose, mannose, glucose, rhamnose, uronic acid and sulfate | Maruyama and Yamamoto 1984 [27] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106-2130. https://doi.org/10.3390/md9102106
Ale MT, Mikkelsen JD, Meyer AS. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Marine Drugs. 2011; 9(10):2106-2130. https://doi.org/10.3390/md9102106
Chicago/Turabian StyleAle, Marcel Tutor, Jørn D. Mikkelsen, and Anne S. Meyer. 2011. "Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds" Marine Drugs 9, no. 10: 2106-2130. https://doi.org/10.3390/md9102106
APA StyleAle, M. T., Mikkelsen, J. D., & Meyer, A. S. (2011). Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Marine Drugs, 9(10), 2106-2130. https://doi.org/10.3390/md9102106