Actinomycetes from Sediments in the Trondheim Fjord, Norway: Diversity and Biological Activity

Abstract
:Introduction
Results
Collection and analysis of the sediment samples

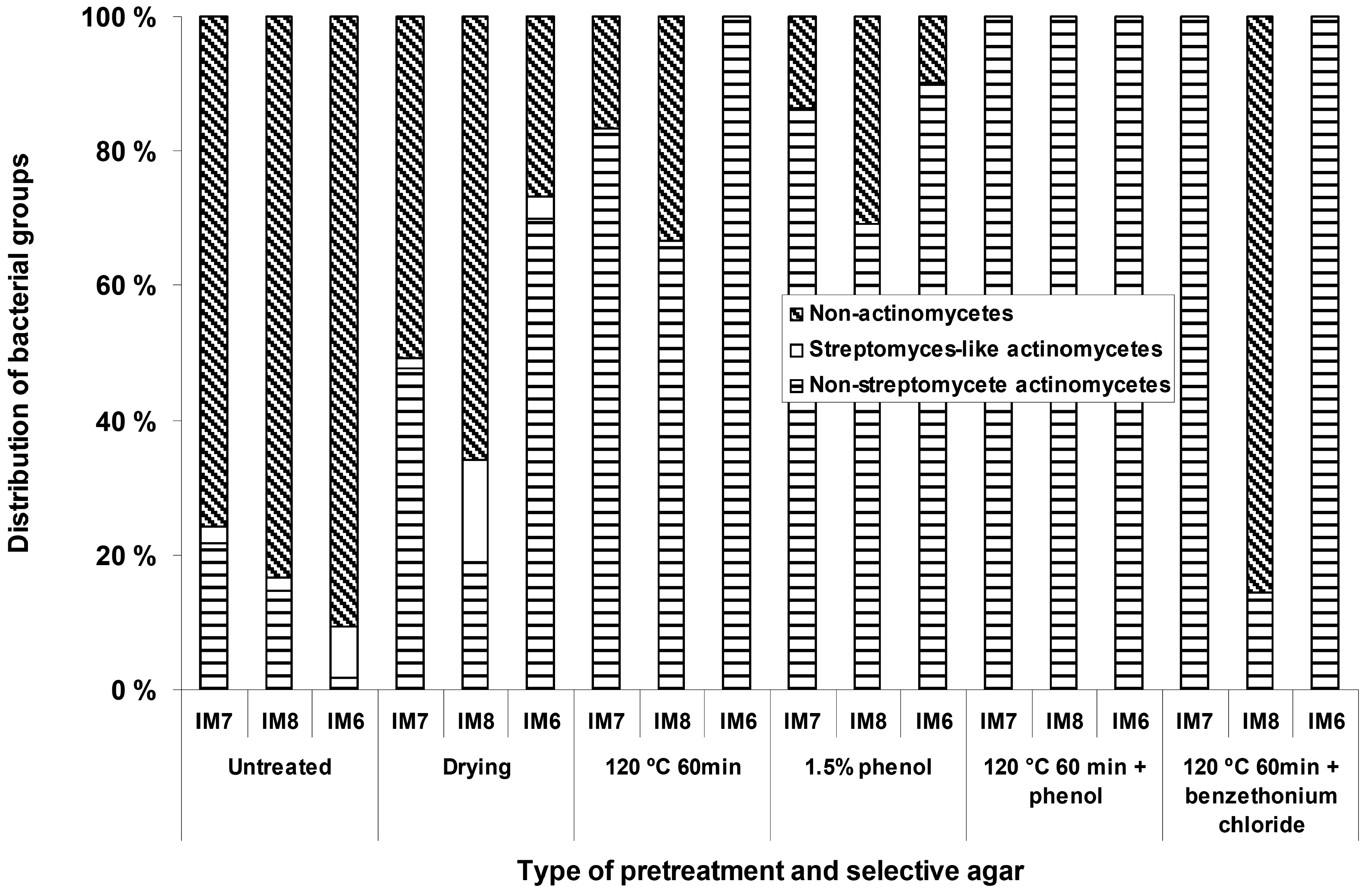{kind=link}
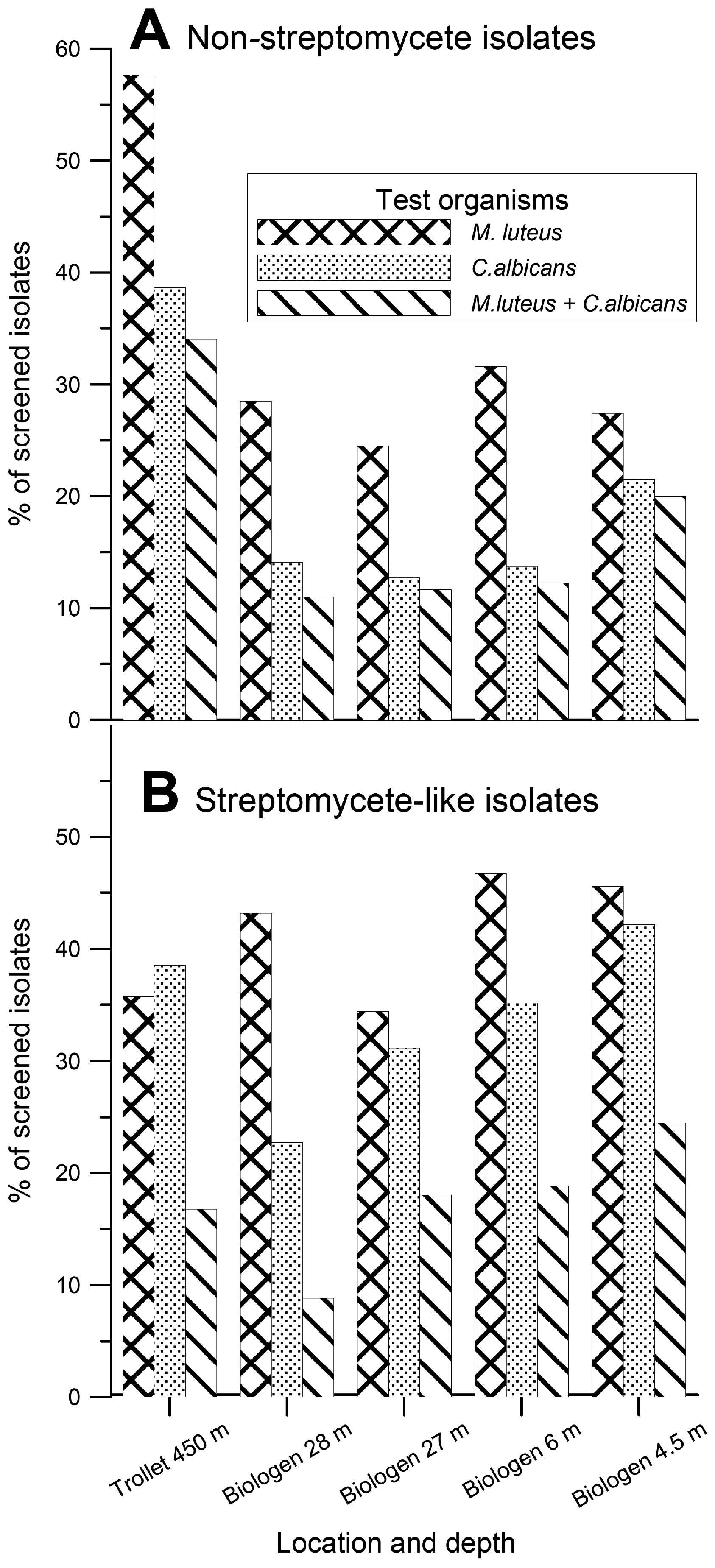{kind=link}
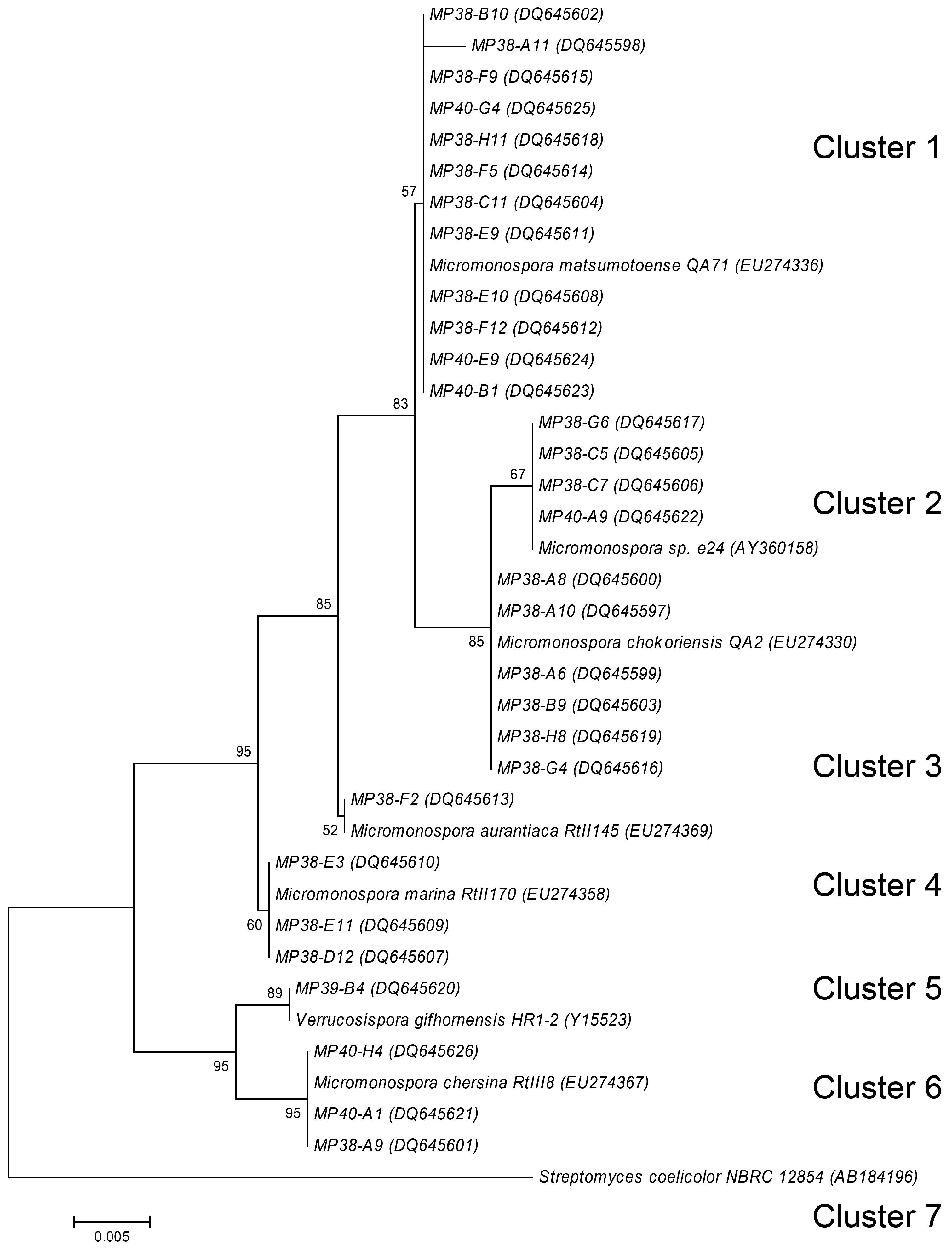{kind=link}
| Site | Sample ID | Depth (m) | Sediment type | Vegetation | Org C(%) | Org N(%) |
|---|---|---|---|---|---|---|
| Biologen | B1 | 4.5 | Fine mud with small stones | + | 0.7 | 0.04 |
| Biologen | B2 | 6.0 | Clay and stones | ++ | 1.1 | 0.06 |
| Biologen | B3 | 27 | Fine muddy sand | – | 0.6 | 0.05 |
| Biologen | B4 | 28 | Clay | – | 0.7 | 0.04 |
| Trollet | T1 | 450 | Clay with fine organic material | – | 1.8 | 0.15 |
Isolation of actinomycete bacteria
| Sample ID | Medium | CFU/mL wet sediment | ||
|---|---|---|---|---|
| Bacteria | Streptomyces-like | Non-Streptomyces actinomycetes | ||
| N.D. - Not detected, no colonies on the agar plates. Plates were incubated at 20 oC for up to 4 weeks. | ||||
| B1 | IM5b | 1.2 x 106 | 2.5 x 102 | 5.0 x 102 |
| IM6b | 9.5 x 104 | 5.5 x 102 | 2.0 x 103 | |
| IM7b | 8.4 x 104 | 1.5 x 103 | 2.4 x 104 | |
| IM8b | 8.0 x 104 | 2.0 x 103 | 1.8 x 104 | |
| B2 | IM5b | 5.4 x 105 | 2.5 x 103 | 5.0 x 103 |
| IM6b | 8.1 x 104 | 6.0 x 103 | 1.5 x 103 | |
| IM7b | 7.4 x 104 | 2.0 x 103 | 1.6 x 104 | |
| IM8b | 1.1 x 105 | 2.3 x 103 | 1.6 x 104 | |
| B3 | IM5b | 5.1 x 105 | 1.5 x 102 | N.D |
| IM6b | 1.0 x 105 | 2.0 x 102 | 3.5 x 103 | |
| IM7b | 7.7 x 104 | 2.0 x 102 | 1.3 x 104 | |
| IM8b | 2.6 x105 | 5.0 x 102 | 1.1 x 104 | |
| B4 | IM5b | 3.6 x 105 | 5.0 x 101 | 5.0 x 102 |
| IM6b | 1.2 x 105 | 4.0 x 102 | 3.5 x 102 | |
| IM7b | 9.4 x 104 | 7.0 x 102 | 1.6 x 104 | |
| IM8b | 1.4 x 105 | 9.0 x 102 | 1.7 x 104 | |
| T1 | IM5b | 2.5 x 104 | N.D. | 1.3 x 103 |
| IM6b | 7.2 x 103 | 7.5 x 101 | 6.7 x 103 | |
| IM7b | 8.0 x 103 | N.D. | 2.3 x 103 | |
| IM8b | 6.3 x 103 | 2.5 x 101 | 2.5 x 103 | |
Sea water requirement
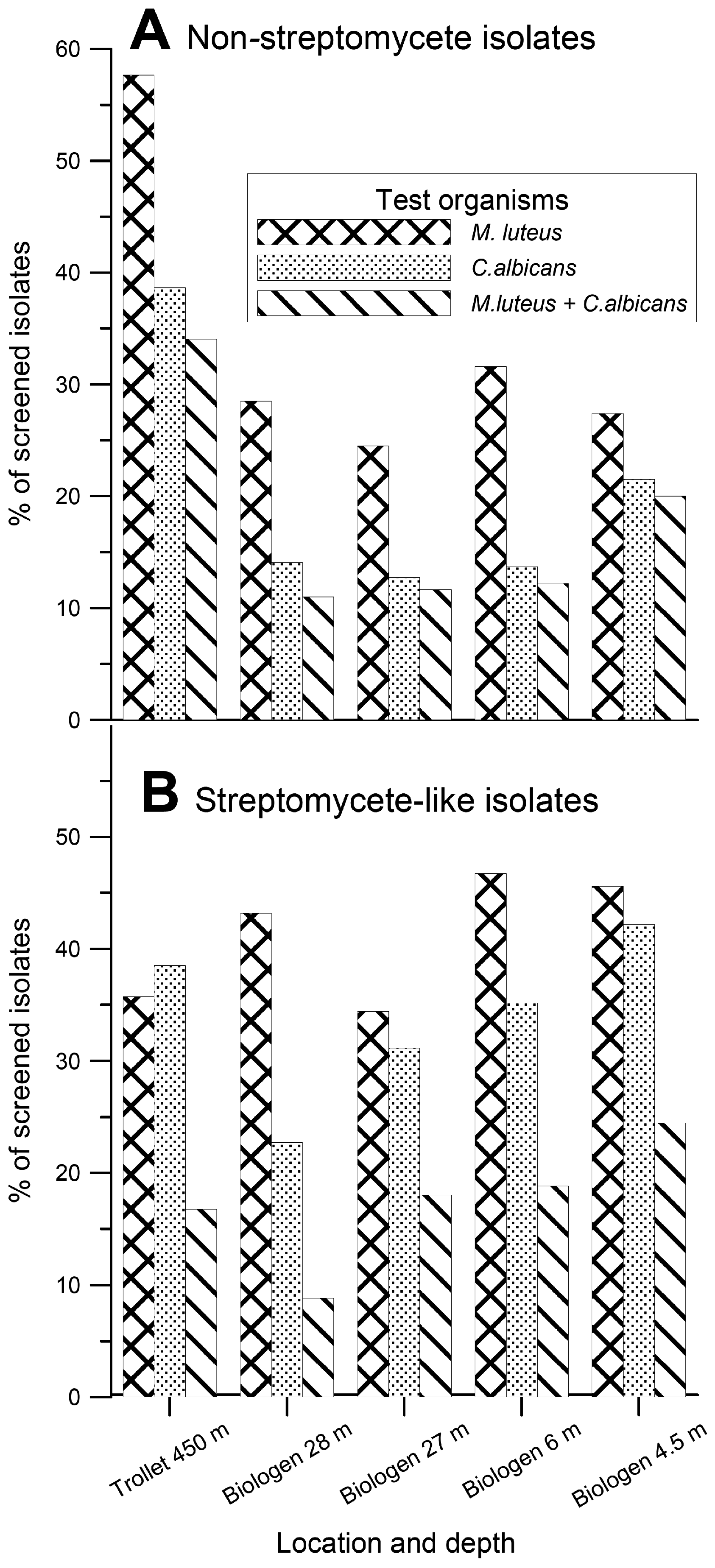
Biological activities
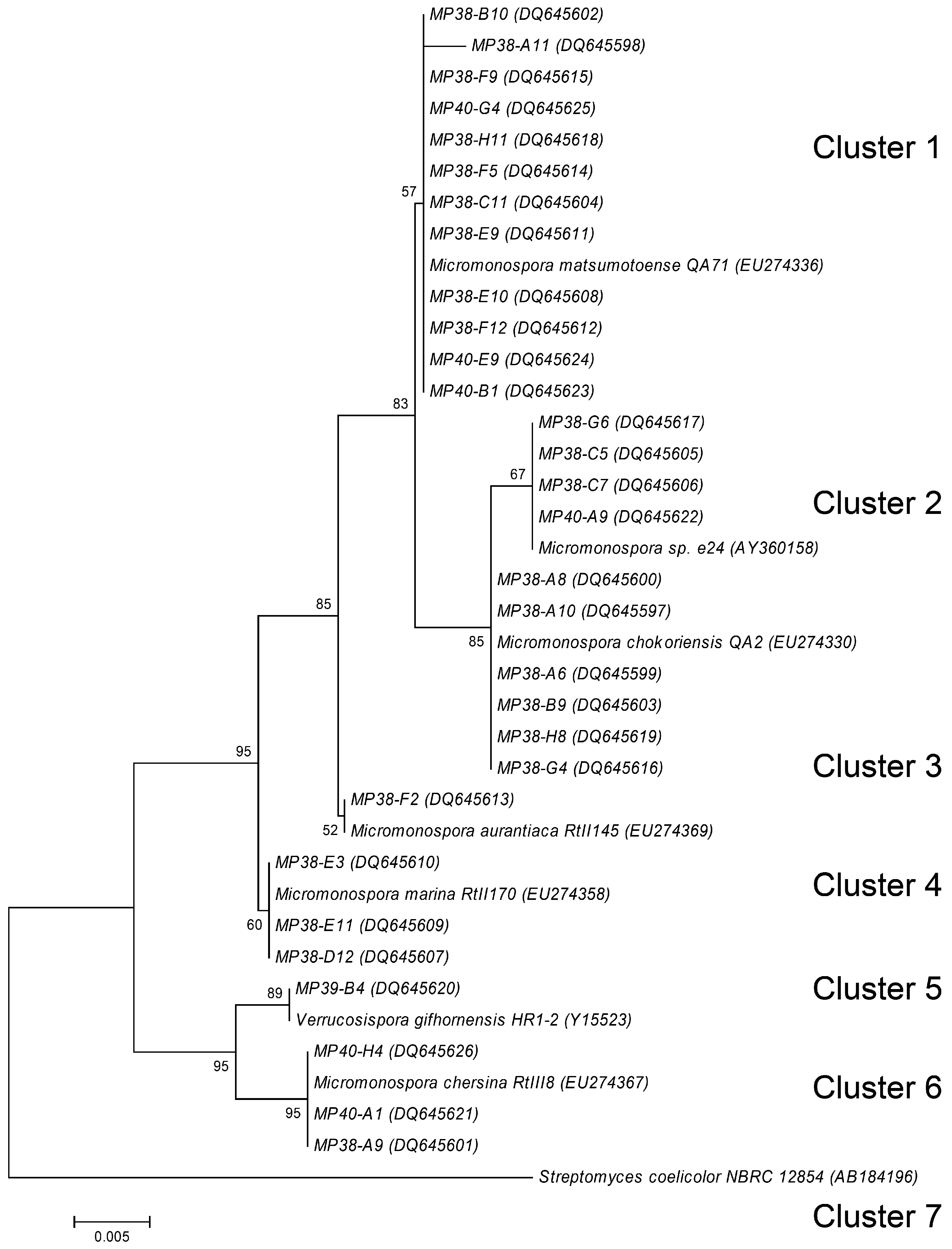
Molecular taxonomy
Discussion
Experimental Section
Sample collection and isolation of bacteria
Carbon and nitrogen content analysis
Isolation and production media
Cultivation, extraction and bioactivity testing
Nucleic acid extraction, 16S rDNA amplification, sequencing and analysis
Acknowledgments
References
- Baltz, R.H. Marcel Faber Roundtable: Is our antibiotic pipeline unproductive because of starvation, constipation or lack of inspiration? J. Ind. Microbiol. Biotechnol. 2006, 33, 507–513. [Google Scholar] [CrossRef]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug. Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Pelaez, F. The historical delivery of antibiotics from microbial natural products - Can history repeat? Biochem. Pharmacol. 2006, 71, 981–990. [Google Scholar]
- Weyland, H. Actinomycetes in North Sea and Atlantic Ocean sediments. Nature 1969, 223, 858. [Google Scholar] [CrossRef] [PubMed]
- Jensen, P.R.; Gontang, E.; Mafnas, C.; Mincer, T.J.; Fenical, W. Culturable marine actinomycete diversity from tropical Pacific Ocean sediments. Environ. Microbiol. 2005, 7, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.J.; Fenchel, T. Cosmopolitan metapopulations of free-living microbial eukaryotes. Protist 2004, 155, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Dolan, J.R. Microbial biogeography? J. Biogeogr. 2006, 33, 199–200. [Google Scholar] [CrossRef]
- Staley, J.T. In Microbial diversity and bioprospecting. ASM Press: Washington, DC, 2004; p. p. 41. [Google Scholar]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M; Krumins, J.A.; Kuske, C.R.; Morin, P.J.; Naeem, S.; Ovreas, L.; Reysenbach, A.L.; Smith, V.H.; Staley, J.T. Microbial biogeography: putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [PubMed]
- Sakshaug, E.; Killingtveit, Å. In Trondheimsfjorden. Tapir Forlag: Trondheim, Norway, 2000; p. p. 65. [Google Scholar]
- Fenchel, T.; King, G.M.; Blackburn, T.H. Bacterial Biogeochemistry: the ecophysiology of mineral cycling. Academic Press: San Diego, CA, USA, 1998; p. p. 117. [Google Scholar]
- Jensen, P.R.; Dwight, R.; Fenical, W. Distribution of actinomycetes in near-shore tropical marine sediments. Appl. Environ. Microbiol. 1991, 57, 1102–1108. [Google Scholar] [PubMed]
- Hayakawa, M. In Selective Isolation of Rare Actinomycetes. Queensland Complete Printing Services: Nambour, Australia, 2003; pp. pp. 56–81. [Google Scholar]
- Hayakawa, M.; Nonomura, H. Humic-acid vitamin agar, a new medium for the selective isolation of soil actinomycetes. J. Ferment. Technol. 1987, 65, 501–509. [Google Scholar] [CrossRef]
- Hayakawa, M.; Sadakata, T.; Kajiura, T.; Nonomura, H. New methods for the highly selective isolation of Micromonospora and Microbispora from soil. J. Ferment. Bioeng. 1991, 72, 320–326. [Google Scholar] [CrossRef]
- Maldonado, L.A.; Stach, J.E.M.; Pathom-aree, W.; Ward, A. C.; Bull, A.T.; Goodfellow, M. Diversity of cultivable actinobacteria in geographically widespread marine sediments. A. Van Leeuw. 2005, 87, 11–18. [Google Scholar] [CrossRef]
- Stach, J.E.M.; Maldonado, L.A.; Masson, D.G.; Ward, A.C.; Goodfellow, M.; Bull, A.T. Statistical approaches for estimating actinobacterial diverity in marine sediments. Appl. Environ. Microbiol. 2003, 69, 6189–6200. [Google Scholar] [CrossRef] [PubMed]
- Stach, J.E.M.; Maldonado, L.A.; Ward, A.C.; Goodfellow, M.; Bull, A.T. New primers for the class Actinobacteria: application to marine and terrestrial environments. Environ. Microbiol. 2003, 5, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, M.; Tamura, T.; Iino, H.; Nonomura, H. Pollen-baiting and drying method for the highly selective isolation of Actinoplanes spp. from soil. J. Ferment. Bioeng. 1991, 72, 433–438. [Google Scholar] [CrossRef]
- Palleroni, N.J. A chemotactic method for the isolation of Actinoplanaceae. Arch. Microbiol. 1980, 128, 53–55. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered chitin agar as a selective medium for enumeration of actinomycetes in water and soil. Appl. Microbiol. 1975, 29, 422–426. [Google Scholar] [PubMed]
- Terekhova, L. In Selective Isolation of Rare Actinomycetes. Queensland Complete Printing Services: Nambour, Australia, 2003; pp. pp. 82–101. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [PubMed]
Share and Cite
Bredholt, H.; Fjærvik, E.; Johnsen, G.; Zotchev, S.B. Actinomycetes from Sediments in the Trondheim Fjord, Norway: Diversity and Biological Activity. Mar. Drugs 2008, 6, 12-24. https://doi.org/10.3390/md6010012
Bredholt H, Fjærvik E, Johnsen G, Zotchev SB. Actinomycetes from Sediments in the Trondheim Fjord, Norway: Diversity and Biological Activity. Marine Drugs. 2008; 6(1):12-24. https://doi.org/10.3390/md6010012
Chicago/Turabian StyleBredholt, Harald, Espen Fjærvik, Geir Johnsen, and Sergey B. Zotchev. 2008. "Actinomycetes from Sediments in the Trondheim Fjord, Norway: Diversity and Biological Activity" Marine Drugs 6, no. 1: 12-24. https://doi.org/10.3390/md6010012
APA StyleBredholt, H., Fjærvik, E., Johnsen, G., & Zotchev, S. B. (2008). Actinomycetes from Sediments in the Trondheim Fjord, Norway: Diversity and Biological Activity. Marine Drugs, 6(1), 12-24. https://doi.org/10.3390/md6010012