
Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life

 ,
, 

Abstract
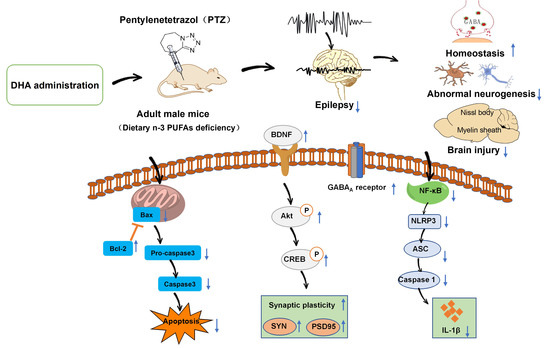
1. Introduction
2. Results
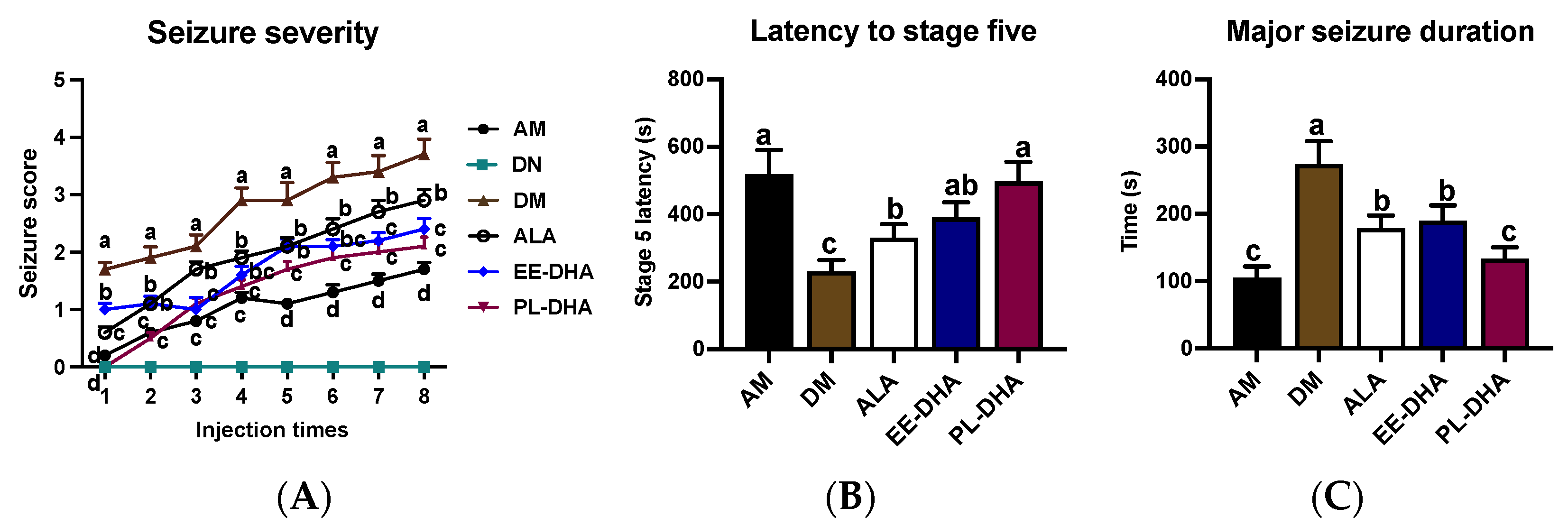
2.1. Effects of Dietary n-3 PUFAs in Adulthood on PTZ-Induced Epileptic Seizure of Mice Feeding n-3 PUFAs Deficiency in Early Life
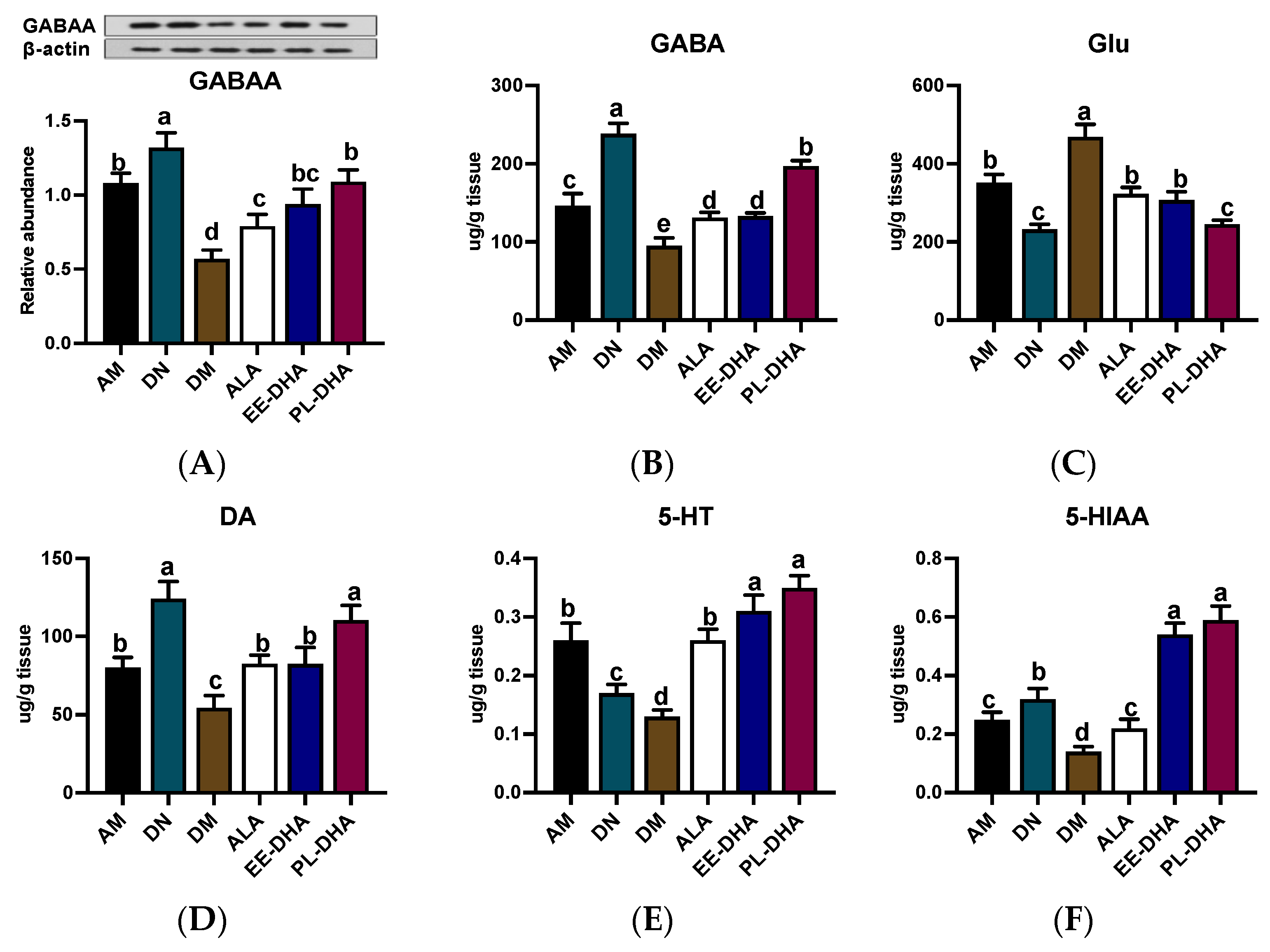
2.2. Effects of Dietary n-3 PUFAs in Adulthood on Neurotransmitter Disturbances Associated with Epilepsy in Mice with Early-Life n-3 PUFAs Deficiency
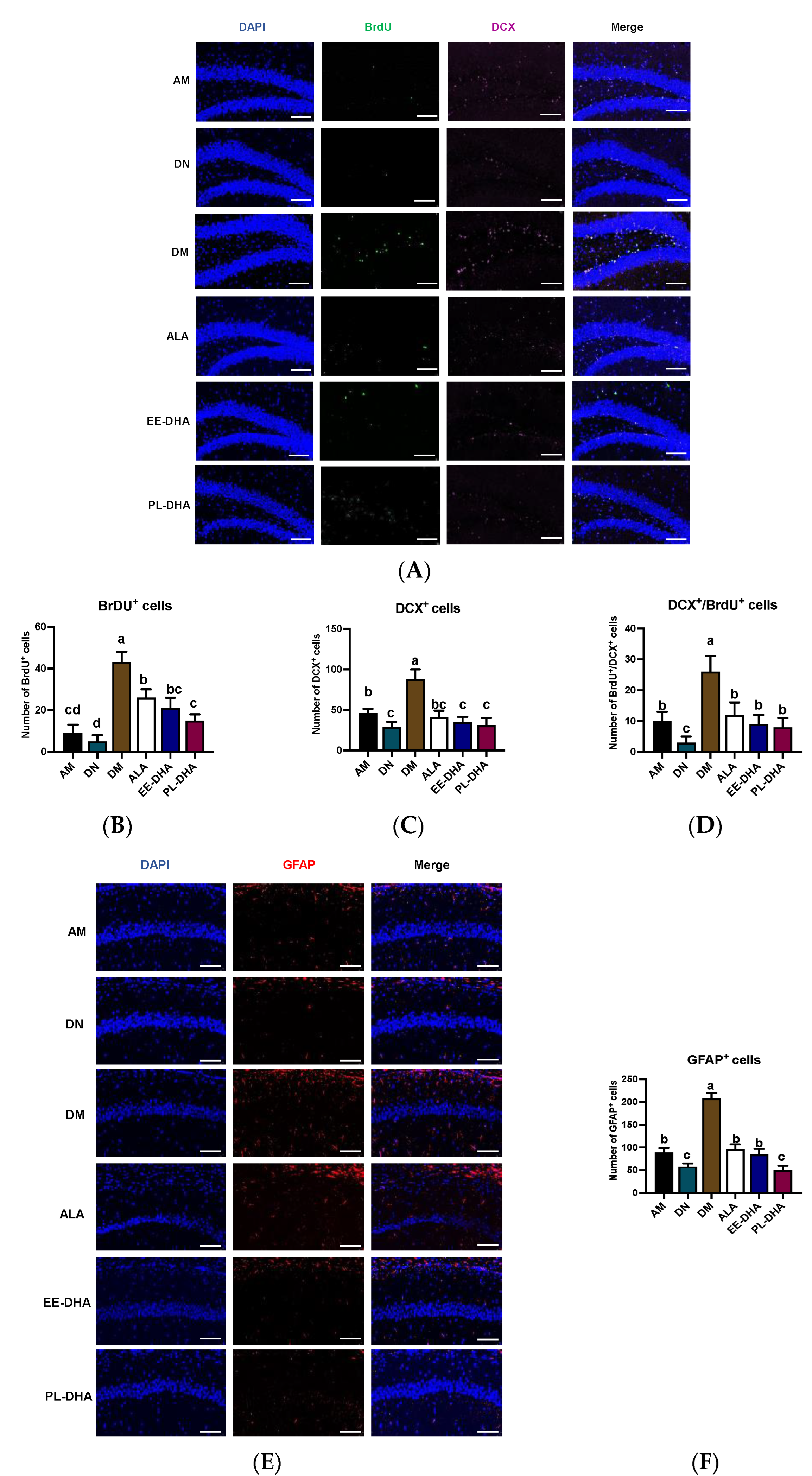
2.3. Effects of Dietary n-3 PUFAs in Adulthood on the Central Nervous System Injury and Neurogenesis in Mice with Early-Life n-3 PUFAs Deficiency and PTZ-Induced Epilepsy
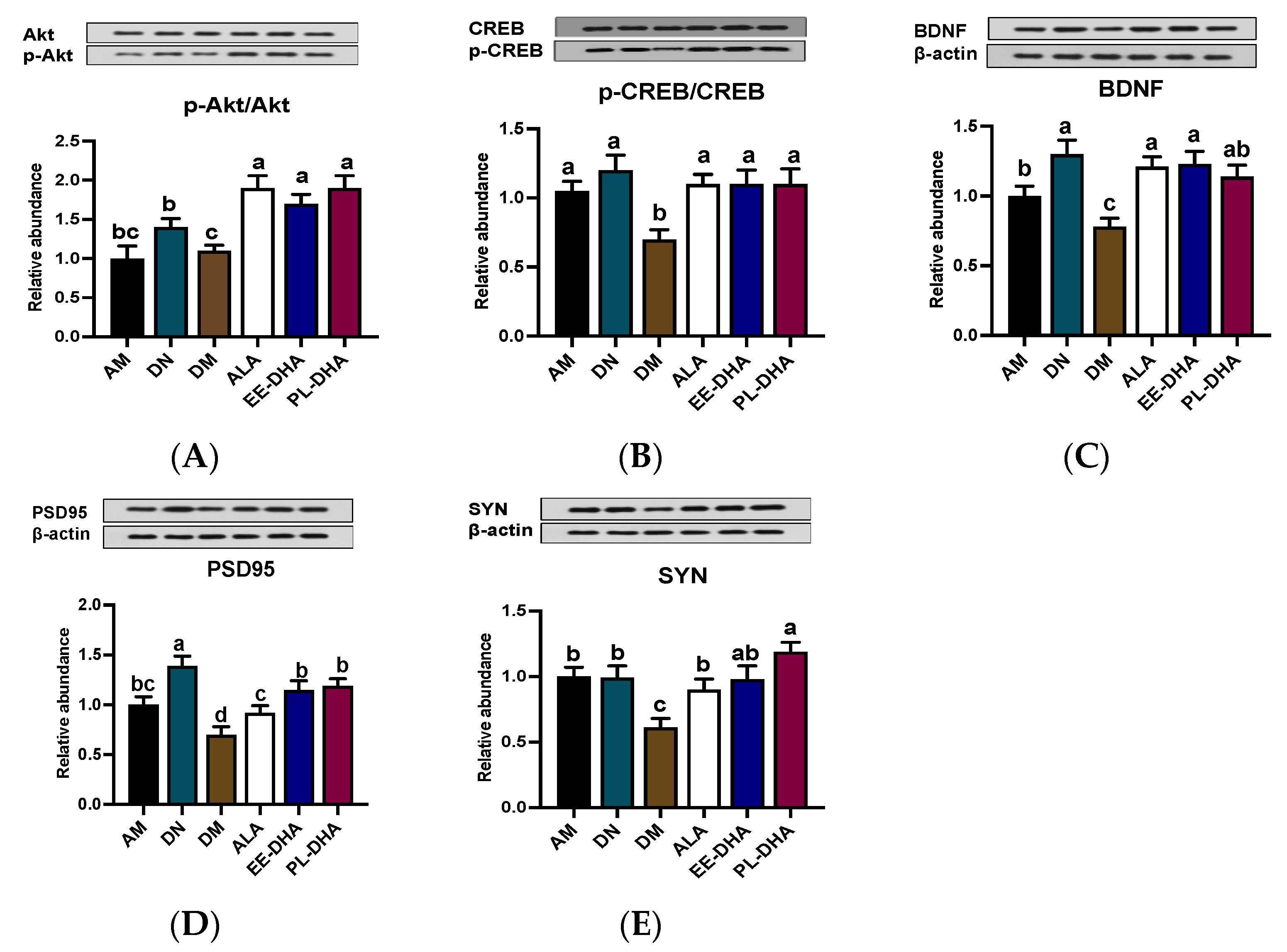
2.4. Effects of Dietary n-3 PUFAs in Adulthood on AKT/CREB/BDNF Pathway and Synaptic Plasticity in Mice with Early-Life n-3 PUFAs Deficiency and PTZ-Induced Epilepsy
2.5. Effects of Dietary n-3 PUFAs in Adulthood on Mitochondria-Dependent Apoptosis in the Hippocampus of PTZ-Induced Epileptic Mice with Early-Life n-3 PUFAs Deficiency
2.6. Effects of Dietary n-3 PUFAs in Adulthood on Hippocampal Neuroinflammation in PTZ-Induced Epileptic Mice with Early-Life n-3 PUFAs Deficiency
2.7. Effects of Dietary n-3 PUFAs in Adulthood on Hippocampal Fatty Acid Composition of PTZ-Induced Epileptic Mice with Early-Life n-3 PUFAs Deficiency
3. Discussion
4. Materials and Methods
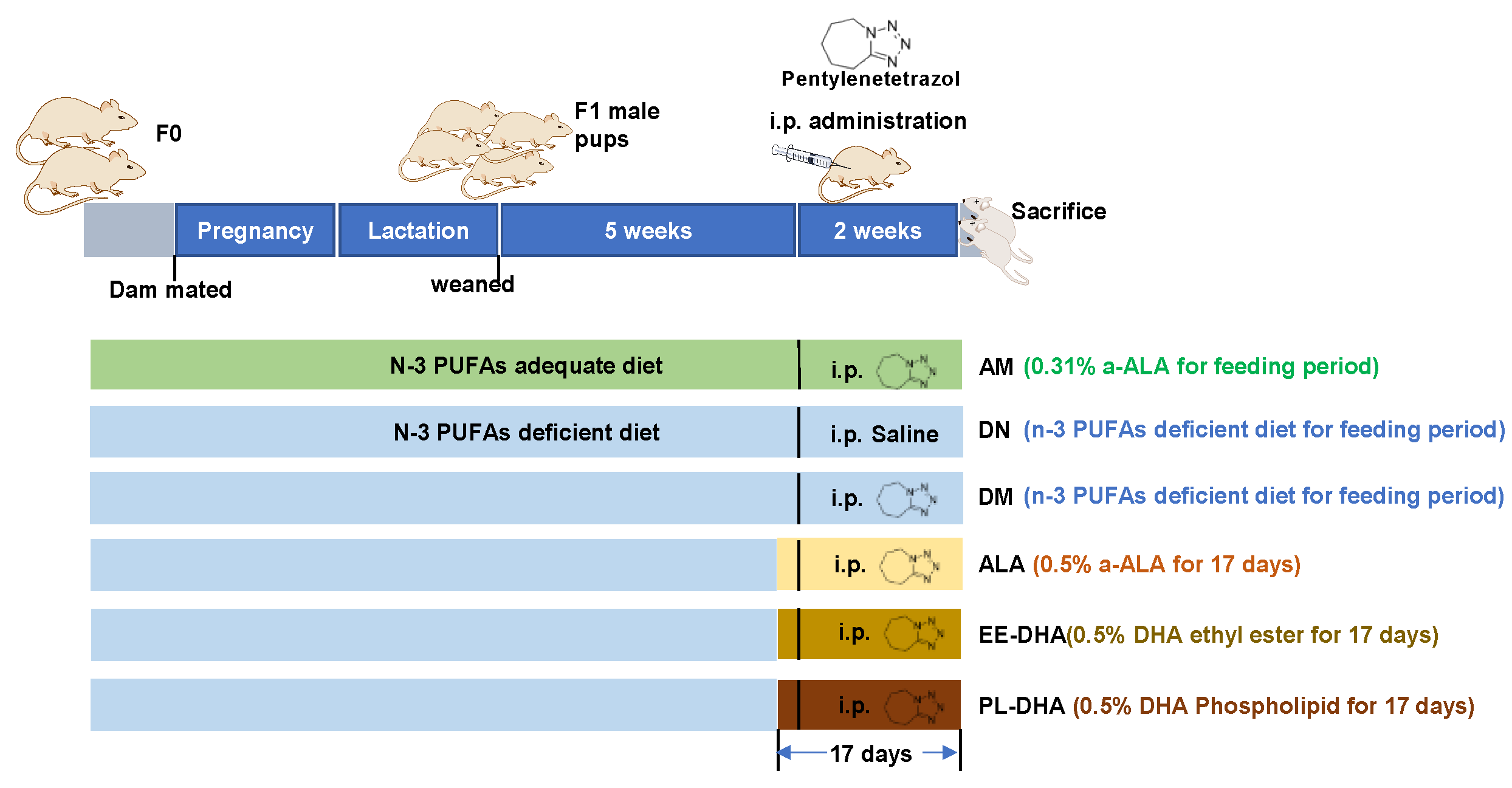
4.1. Animals and Administration
4.2. LC-MS/MS Analysis for Neurotransmitter
4.3. Histological Examination
4.4. Immunohistochemistry
4.5. Immunofluorescence Detection of Brain
4.6. Western Blot and ELISA Analysis
4.7. Fatty Acid Profile Analysis of Hippocampus
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huberfeld, G.; Blauwblomme, T.; Miles, R. Hippocampus and epilepsy: Findings from human tissues. Rev. Neurol. 2015, 171, 236–251. [Google Scholar] [CrossRef] [PubMed]
- Seifert, G.; Steinhäuser, C. Neuron-astrocyte signaling and epilepsy. Exp. Neurol. 2013, 244, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Fonseca, E.; Ettcheto, M.; Sánchez-López, E.; de Rojas, I.; Alonso-Lana, S.; Morató, X.; Souto, E.B.; Toledo, M.; Boada, M.; et al. Epilepsy in Neurodegenerative Diseases: Related Drugs and Molecular Pathways. Pharmaceuticals 2021, 14, 1057. [Google Scholar] [CrossRef] [PubMed]
- Beghi, E. The Epidemiology of Epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Leyrolle, Q.; Decoeur, F.; Dejean, C.; Brière, G.; Leon, S.; Bakoyiannis, I.; Baroux, E.; Sterley, T.L.; Bosch-Bouju, C.; Morel, L.; et al. N-3 PUFA deficiency disrupts oligodendrocyte maturation and myelin integrity during brain development. Glia 2022, 70, 50–70. [Google Scholar] [CrossRef]
- Wu, F.; Wang, D.D.; Shi, H.H.; Wang, C.C.; Xue, C.H.; Wang, Y.M.; Zhang, T.T. N-3 PUFA-Deficiency in Early Life Exhibits Aggravated MPTP-Induced Neurotoxicity in Old Age while Supplementation with DHA/EPA-Enriched Phospholipids Exerts a Neuroprotective Effect. Mol. Nutr. Food Res. 2021, 65, e2100339. [Google Scholar] [CrossRef]
- Wang, D.D.; Wu, F.; Ding, L.; Shi, H.H.; Xue, C.H.; Wang, Y.M.; Zhang, T.T. Dietary n-3 PUFA Deficiency Increases Vulnerability to Scopolamine-Induced Cognitive Impairment in Male C57BL/6 Mice. J. Nutr. 2021, 151, 2206–2214. [Google Scholar] [CrossRef]
- Madore, C.; Leyrolle, Q.; Morel, L.; Rossitto, M.; Greenhalgh, A.D.; Delpech, J.C.; Martinat, M.; Bosch-Bouju, C.; Bourel, J.; Rani, B.; et al. Essential omega-3 fatty acids tune microglial phagocytosis of synaptic elements in the mouse developing brain. Nat. Commun. 2020, 11, 6133. [Google Scholar] [CrossRef]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef]
- Vreugdenhil, M.; Bruehl, C.; Voskuyl, R.A.; Kang, J.X.; Leaf, A.; Wadman, W.J. Polyunsaturated fatty acids modulate sodium and calcium currents in CA1 neurons. Proc. Natl. Acad. Sci. USA 1996, 93, 12559–12563. [Google Scholar] [CrossRef]
- Rogers, L.K.; Valentine, C.J.; Keim, S.A. DHA supplementation: Current implications in pregnancy and childhood. Pharmacol. Res. 2013, 70, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Nabekura, J.; Noguchi, K.; Witt, M.R.; Nielsen, M.; Akaike, N. Functional modulation of human recombinant gamma-aminobutyric acid type A receptor by docosahexaenoic acid. J. Biol. Chem. 1998, 273, 11056–11061. [Google Scholar] [CrossRef] [PubMed]
- DeGiorgio, C.M.; Taha, A.Y. Omega-3 fatty acids (ῳ-3 fatty acids) in epilepsy: Animal models and human clinical trials. Expert Rev. Neurother. 2016, 16, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Kawano, S.; Itoh, K.; Ishihara, Y. Maternal intake of docosahexaenoic acid decreased febrile seizure sensitivity by increasing estrogen synthesis in offspring. Epilepsy Behav. 2021, 121, 108038. [Google Scholar] [CrossRef]
- Basak, S.; Vilasagaram, S.; Duttaroy, A.K. Maternal dietary deficiency of n-3 fatty acids affects metabolic and epigenetic phenotypes of the developing fetus. Prostaglandins Leukot. Essent. Fat. Acids 2020, 158, 102109. [Google Scholar] [CrossRef]
- Werner, F.-M.; Coveñas, R. Classical neurotransmitters and neuropeptides involved in generalized epilepsy in a multi-neurotransmitter system: How to improve the antiepileptic effect? Epilepsy Behav. 2017, 71, 124–129. [Google Scholar] [CrossRef]
- Layé, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-Inflammatory Effects of Omega-3 Fatty Acids in the Brain: Physiological Mechanisms and Relevance to Pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef]
- von Schacky, C. Omega-3 fatty acids and cardiovascular disease. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 129–135. [Google Scholar] [CrossRef]
- Trépanier, M.O.; Lim, J.; Lai, T.K.; Cho, H.J.; Domenichiello, A.F.; Chen, C.T.; Taha, A.Y.; Bazinet, R.P.; Burnham, W.M. Intraperitoneal administration of docosahexaenoic acid for 14days increases serum unesterified DHA and seizure latency in the maximal pentylenetetrazol model. Epilepsy Behav. 2014, 33, 138–143. [Google Scholar] [CrossRef]
- Gavzan, H.; Hashemi, F.; Babaei, J.; Sayyah, M. A role for peroxisome proliferator-activated receptor α in anticonvulsant activity of docosahexaenoic acid against seizures induced by pentylenetetrazole. Neurosci. Lett. 2018, 681, 83–86. [Google Scholar] [CrossRef]
- Moezifar, M.; Sayyah, M.; Zendehdel, M.; Gavzan, H. Docosahexaenoic acid prevents resistance to antiepileptic drugs in two animal models of drug-resistant epilepsy. Nutr. Neurosci. 2019, 22, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Barceló-Coblijn, G.; Murphy, E.J. Alpha-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res. 2009, 48, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.T.; Xu, J.; Wang, Y.M.; Xue, C.H. Health benefits of dietary marine DHA/EPA-enriched glycerophospholipids. Prog. Lipid Res. 2019, 75, 100997. [Google Scholar] [CrossRef] [PubMed]
- Bateup, H.S.; Johnson, C.A.; Denefrio, C.L.; Saulnier, J.L.; Kornacker, K.; Sabatini, B.L. Excitatory/inhibitory synaptic imbalance leads to hippocampal hyperexcitability in mouse models of tuberous sclerosis. Neuron 2013, 78, 510–522. [Google Scholar] [CrossRef]
- Bonansco, C.; Fuenzalida, M. Plasticity of Hippocampal Excitatory-Inhibitory Balance: Missing the Synaptic Control in the Epileptic Brain. Neural Plast. 2016, 2016, 8607038. [Google Scholar] [CrossRef]
- Casillas-Espinosa, P.M.; Powell, K.L.; O’Brien, T.J. Regulators of synaptic transmission: Roles in the pathogenesis and treatment of epilepsy. Epilepsia 2012, 53 (Suppl. 9), 41–58. [Google Scholar] [CrossRef] [PubMed]
- Błaszczyk, B.; Czuczwar, S.J. Epilepsy coexisting with depression. Pharmacol. Rep. 2016, 68, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Pang, Q.; Liu, Y.; Shang, W.; Zhai, G.; Ge, M. Neuronal apoptosis in the resected sclerotic hippocampus in patients with mesial temporal lobe epilepsy. J. Clin. Neurosci. 2007, 14, 835–840. [Google Scholar] [CrossRef]
- Firląg, M.; Kamaszewski, M.; Gaca, K.; Adamek, D.; Bałasińska, B. The neuroprotective effect of long-term n-3 polyunsaturated fatty acids supplementation in the cerebral cortex and hippocampus of aging rats. Folia Neuropathol. 2013, 51, 235–242. [Google Scholar] [CrossRef]
- Jiruska, P.; Shtaya, A.B.; Bodansky, D.M.; Chang, W.C.; Gray, W.P.; Jefferys, J.G. Dentate gyrus progenitor cell proliferation after the onset of spontaneous seizures in the tetanus toxin model of temporal lobe epilepsy. Neurobiol. Dis. 2013, 54, 492–498. [Google Scholar] [CrossRef]
- Hoss, F.; Rodriguez-Alcazar, J.F.; Latz, E. Assembly and regulation of ASC specks. Cell. Mol. Life Sci. 2017, 74, 1211–1229. [Google Scholar] [CrossRef] [PubMed]
- Nakagawara, A. Trk receptor tyrosine kinases: A bridge between cancer and neural development. Cancer Lett. 2001, 169, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Wong, M. Modulation of dendritic spines in epilepsy: Cellular mechanisms and functional implications. Epilepsy Behav. 2005, 7, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Santos, R.A.; Cohen-Cory, S. Impact of maternal n-3 polyunsaturated fatty acid deficiency on dendritic arbor morphology and connectivity of developing Xenopus laevis central neurons in vivo. J. Neurosci. 2015, 35, 6079–6092. [Google Scholar] [CrossRef]
- Hoffmann, K.; Lindner, M.; Gröticke, I.; Stangel, M.; Löscher, W. Epileptic seizures and hippocampal damage after cuprizone-induced demyelination in C57BL/6 mice. Exp. Neurol. 2008, 210, 308–321. [Google Scholar] [CrossRef]
- Srinivas, V.; Molangiri, A.; Mallepogu, A.; Kona, S.R.; Ibrahim, A.; Duttaroy, A.K.; Basak, S. Maternal n-3 PUFA deficiency alters uterine artery remodeling and placental epigenome in the mice. J. Nutr. Biochem. 2021, 96, 108784. [Google Scholar] [CrossRef]
- Samokhina, E.; Samokhin, A. Neuropathological profile of the pentylenetetrazol (PTZ) kindling model. Int. J. Neurosci. 2018, 128, 1086–1096. [Google Scholar] [CrossRef]
- Van Erum, J.; Van Dam, D.; De Deyn, P.P. PTZ-induced seizures in mice require a revised Racine scale. Epilepsy Behav. 2019, 95, 51–55. [Google Scholar] [CrossRef]
- Huang, F.; Li, J.; Shi, H.-L.; Wang, T.-T.; Muhtar, W.; Du, M.; Zhang, B.-B.; Wu, H.; Yang, L.; Hu, Z.-B.; et al. Simultaneous quantification of seven hippocampal neurotransmitters in depression mice by LC–MS/MS. J. Neurosci. Methods 2014, 229, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Ascoli, I.; Lees, M.; Meath, J.A.; Le, B.N. Preparation of lipide extracts from brain tissue. J. Biol. Chem. 1951, 191, 833–841. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid (%) | AM | DN | DM | ALA | EE-DHA | PL-DHA |
|---|---|---|---|---|---|---|
| C14:0 | 0.21 ± 0.09 | 0.54 ± 0.10 | 0.60 ± 0.15 | 0.46 ± 0.18 | 0.23 ± 0.10 | 0.65 ± 0.23 |
| C14:1 | 2.32 ± 0.21 | 2.03 ± 0.15 | 2.01 ± 0.09 | 2.33 ± 0.42 | 2.12 ± 0.38 | 1.72 ± 0.35 |
| C16:0 | 19.76 ± 1.09 | 19.56 ± 1.19 | 21.43 ± 1.08 | 19.87 ± 1.51 | 21.47 ± 1.91 | 19.44 ± 1.47 |
| C16:1 | 0.81 ± 0.09 | 0.83 ± 0.15 | 0.95 ± 0.20 | 1.02 ± 0.10 | 0.72 ± 0.19 | 0.87 ± 0.13 |
| C18:0 | 21.13 ± 1.24 | 21.91 ± 1.51 | 21.23 ± 0.92 | 22.04 ± 1.48 | 21.62 ± 1.75 | 20.55 ± 1.37 |
| C18:1 | 20.42 ± 1.99 | 19.18 ± 2.08 | 19.83 ± 1.01 | 19.61 ± 0.91 | 20.13 ± 1.16 | 19.84 ± 1.31 |
| C18:2 (n-6) | 0.71 ± 0.21 | 0.32 ± 0.10 | 0.39 ± 0.11 | 0.43 ± 0.14 | 0.42 ± 0.21 | 0.39 ± 0.15 |
| C20:1 | 3.19 ± 0.32 a | 2.28 ± 0.22 b | 2.74 ± 0.19 ab | 2.83 ± 0.22 ab | 2.08 ± 0.29 b | 2.56 ± 0.37 b |
| C20:3 (n-3) | 1.01 ± 0.09 | 1.24 ± 0.19 | 1.02 ± 0.31 | 0.36 ± 0.13 | 0.52 ± 0.15 | 1.23 ± 0.24 |
| C20:4 (n-6) | 8.52 ± 0.91 ab | 9.20 ± 0.58 a | 8.69 ± 0.61 ab | 7.83 ± 0.39 c | 7.55 ± 0.57 bc | 8.37 ± 0. 62 ab |
| C22:4 (n-6) | 2.77 ± 0.16 b | 3.49 ± 0.23 a | 3.72 ± 0.52 a | 3.35 ± 0.61 ab | 3.28 ± 0.42 ab | 3.11 ± 0.27 ab |
| C22:5 (n-6) | 0.44 ± 0.12 d | 7.97 ± 0.47 a | 6.75 ± 0.37 a | 4.61 ± 0.36 b | 3.84 ± 0.28 c | 3.75 ± 0.25 c |
| C24:0 | 0.93 ± 0.10 | 0.77 ± 0.19 | 0.76 ± 0.15 | 1.17 ± 0.21 | 0.38 ± 0.09 | 0.97 ± 0.12 |
| C22:6 (n-3) | 14.85 ± 1.07 a | 8.52 ± 0.87 c | 7.41 ± 0.61 c | 11.05 ± 0.82 b | 12.83 ± 1.31 a | 13.61 ± 1.09 a |
| Other fatty acid | 2.93 ± 0.21 | 2.17 ± 0.19 | 2.47 ± 0.32 | 3.04 ± 0.28 | 2.82 ± 0.16 | 2.94 ± 0.35 |
| ∑n-6 LC-PUFA/ ∑n-3 LC-PUFA | 0.78 ± 0.12 d | 2.43 ± 0.19 a | 2.04 ± 0.21 a | 1.35 ± 0.11 b | 1.06 ± 0.09 c | 1.11 ± 0.10 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.-C.; Wang, C.-C.; Li, X.-Y.; Wang, D.-D.; Wang, Y.-M.; Xue, C.-H.; Wen, M.; Zhang, T.-T. Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life. Mar. Drugs 2023, 21, 354. https://doi.org/10.3390/md21060354
Zhao Y-C, Wang C-C, Li X-Y, Wang D-D, Wang Y-M, Xue C-H, Wen M, Zhang T-T. Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life. Marine Drugs. 2023; 21(6):354. https://doi.org/10.3390/md21060354
Chicago/Turabian StyleZhao, Ying-Cai, Cheng-Cheng Wang, Xiao-Yue Li, Dan-Dan Wang, Yu-Ming Wang, Chang-Hu Xue, Min Wen, and Tian-Tian Zhang. 2023. "Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life" Marine Drugs 21, no. 6: 354. https://doi.org/10.3390/md21060354
APA StyleZhao, Y.-C., Wang, C.-C., Li, X.-Y., Wang, D.-D., Wang, Y.-M., Xue, C.-H., Wen, M., & Zhang, T.-T. (2023). Supplementation of n-3 PUFAs in Adulthood Attenuated Susceptibility to Pentylenetetrazol Induced Epilepsy in Mice Fed with n-3 PUFAs Deficient Diet in Early Life. Marine Drugs, 21(6), 354. https://doi.org/10.3390/md21060354