
Genus Acanthella—A Wealthy Treasure: Secondary Metabolites, Synthesis, Biosynthesis, and Bioactivities

 ,
, 
Abstract
1. Introduction
2. Methodology
3. Secondary Metabolites of Genus Acanthella
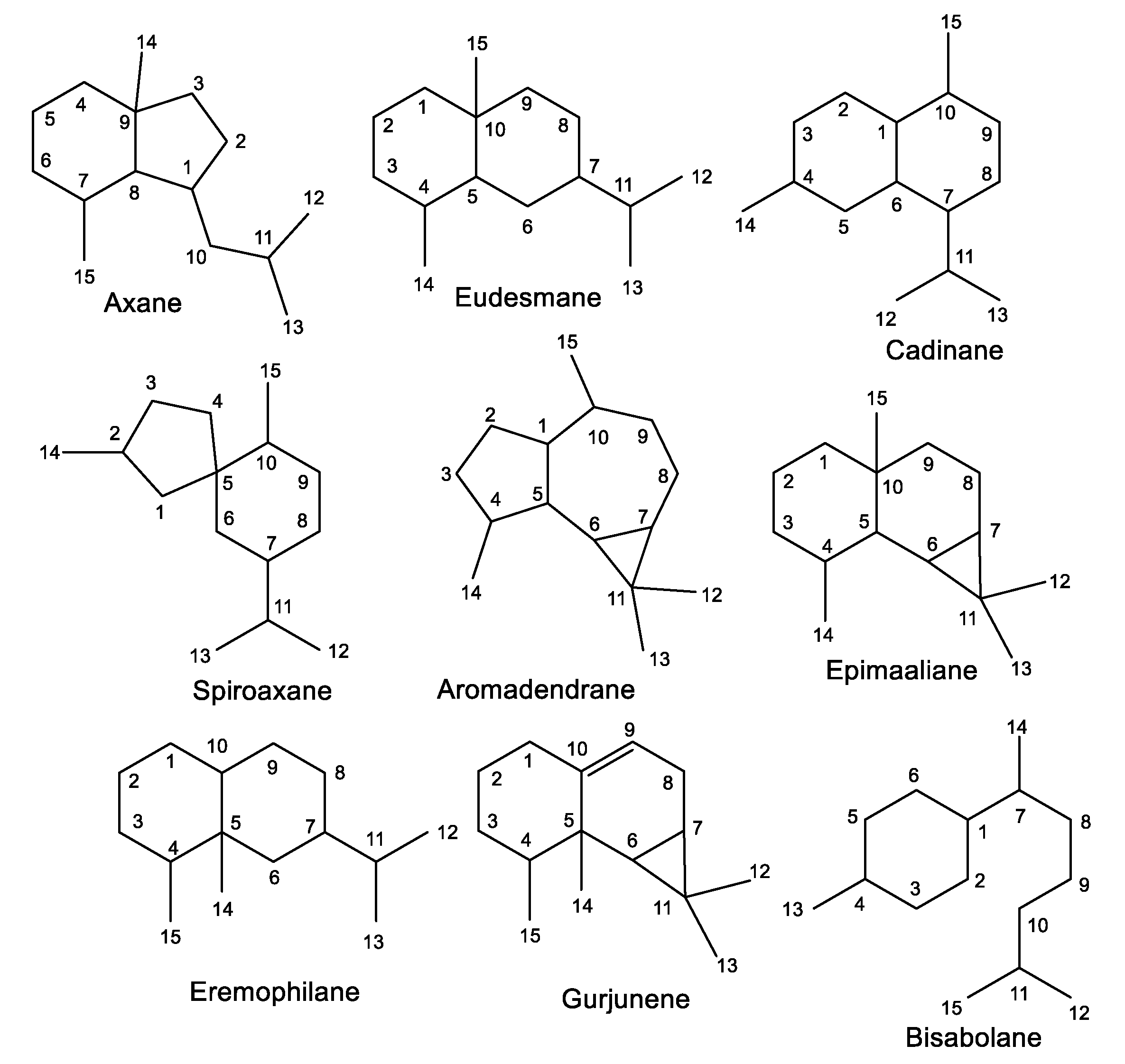
3.1. Sesquiterpenes
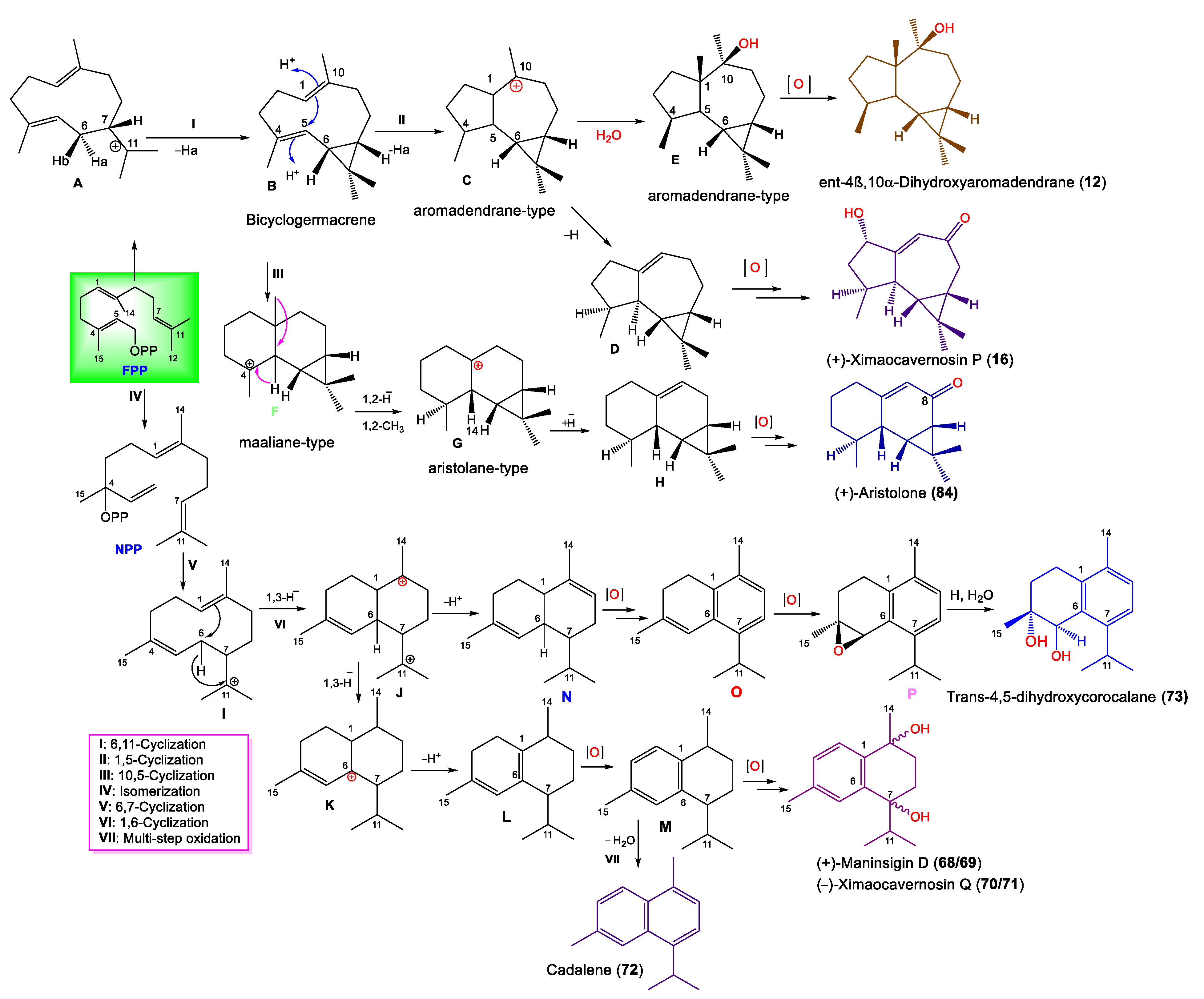
3.1.1. Aromadendrane-Type Sesquiterpenes
3.1.2. Spiroaxane-Type Sesquiterpenes
3.1.3. Eudesmane-Type Sesquiterpenes
3.1.4. Cadinene-Type Sesquiterpenes
3.1.5. Other Sesquiterpenes
3.2. Diterpenoids
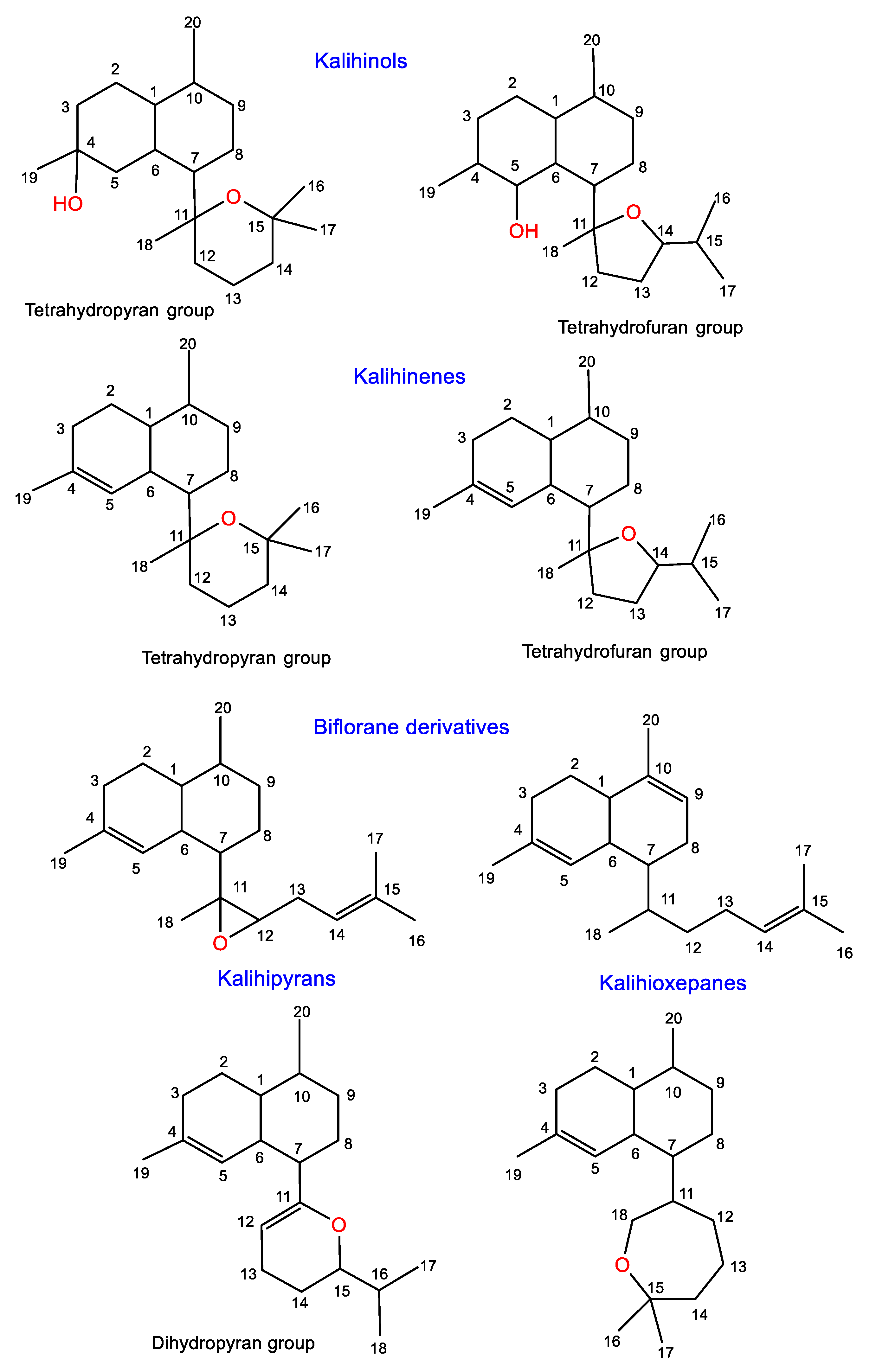
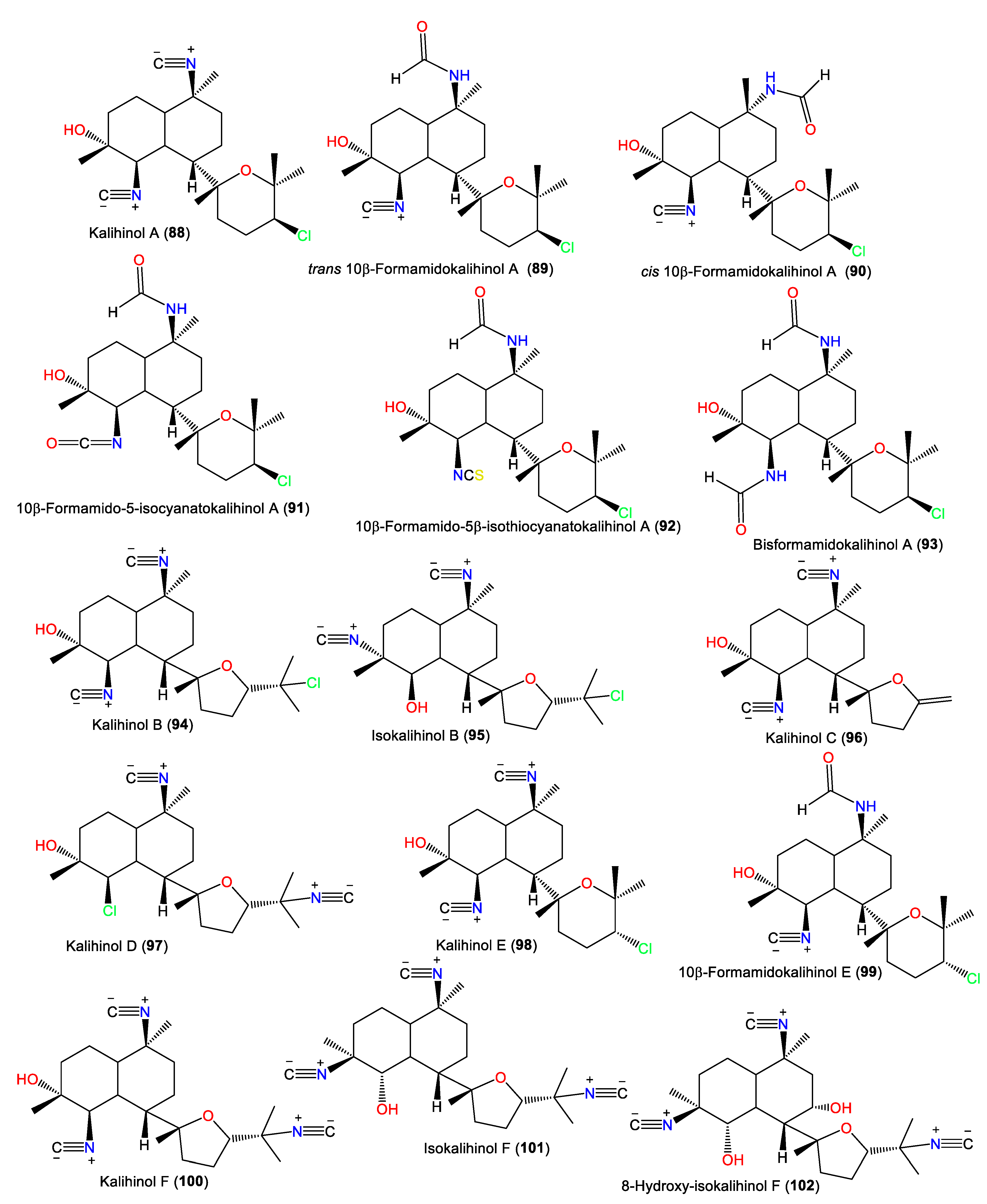
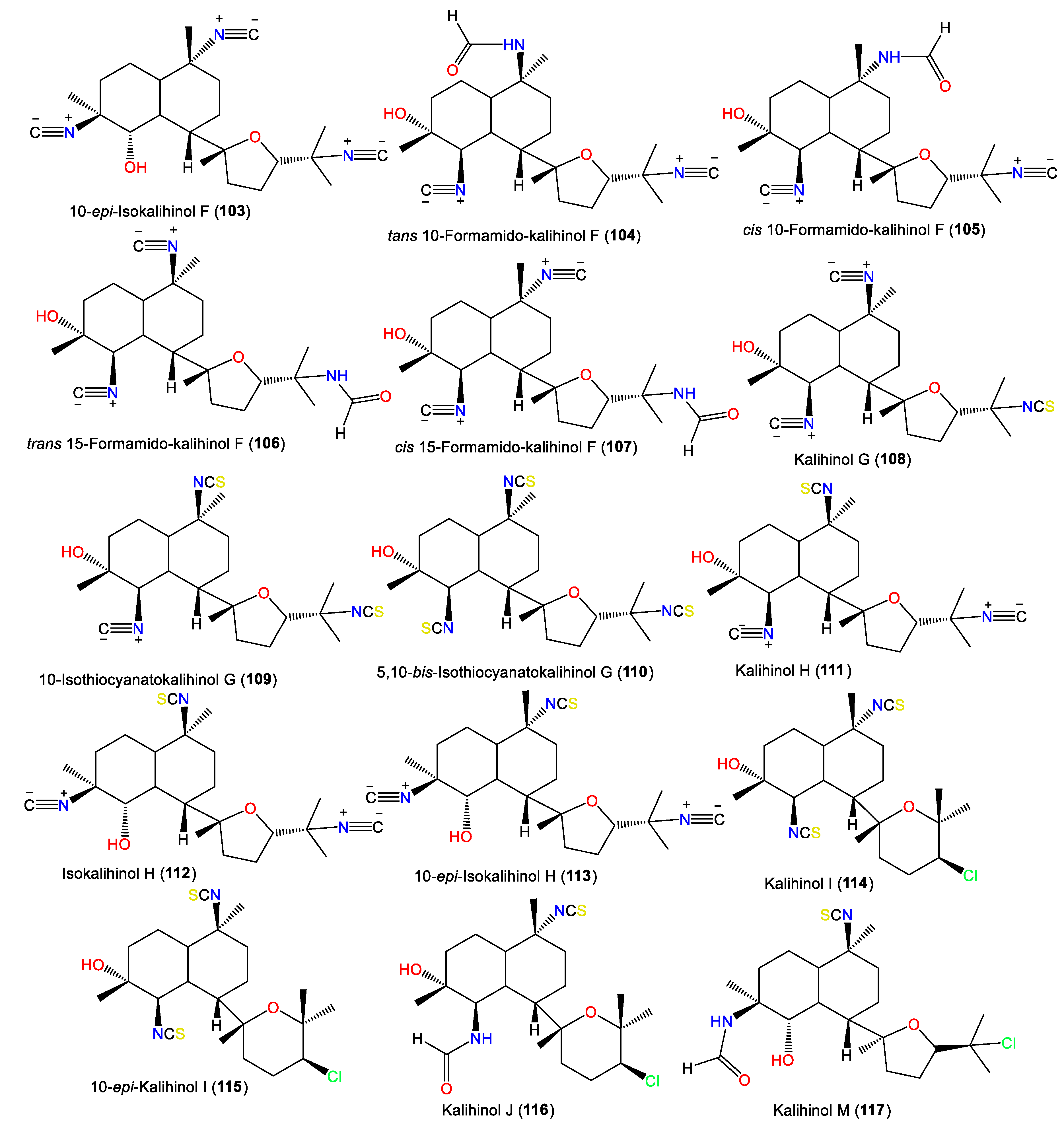
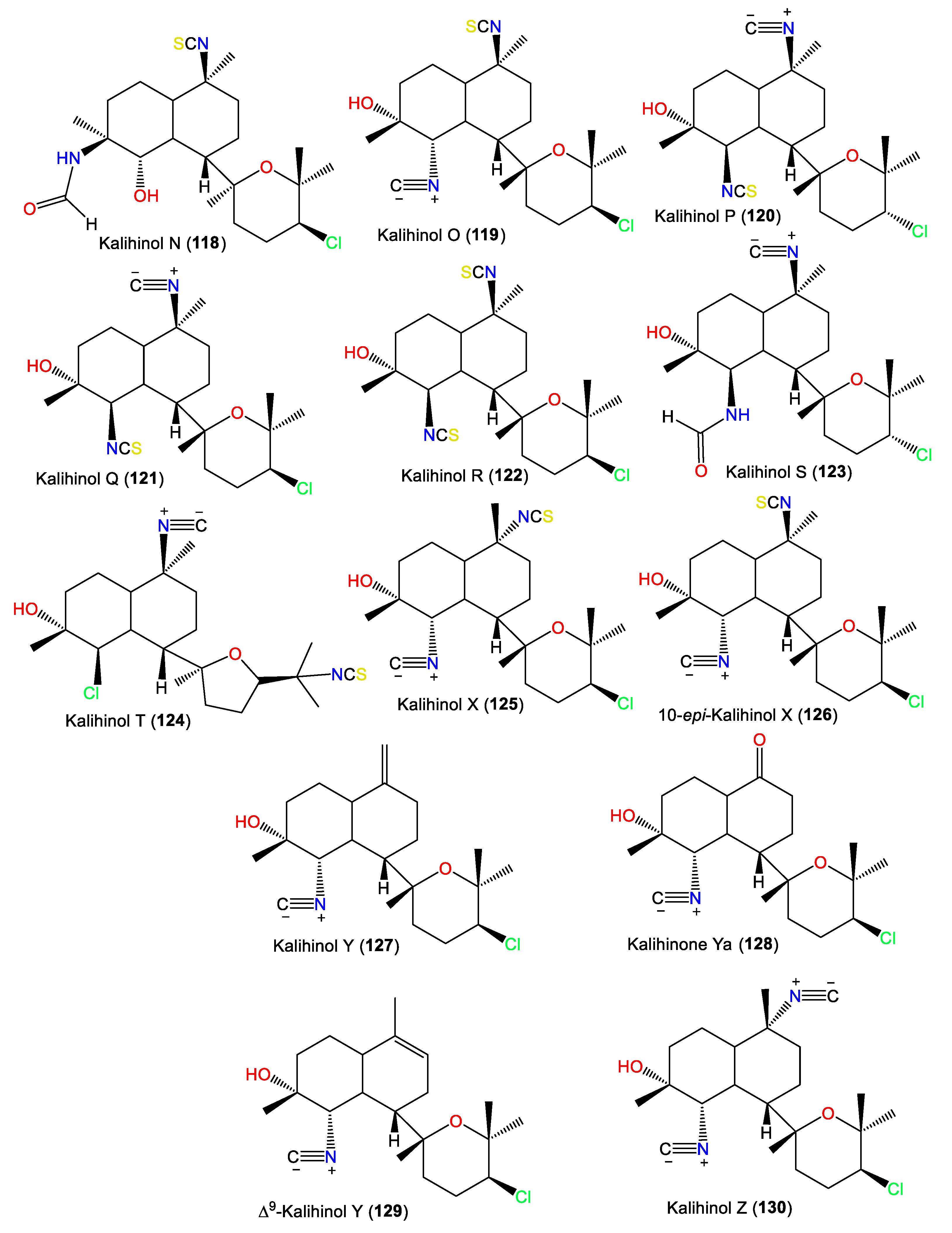
3.2.1. Kalihinols
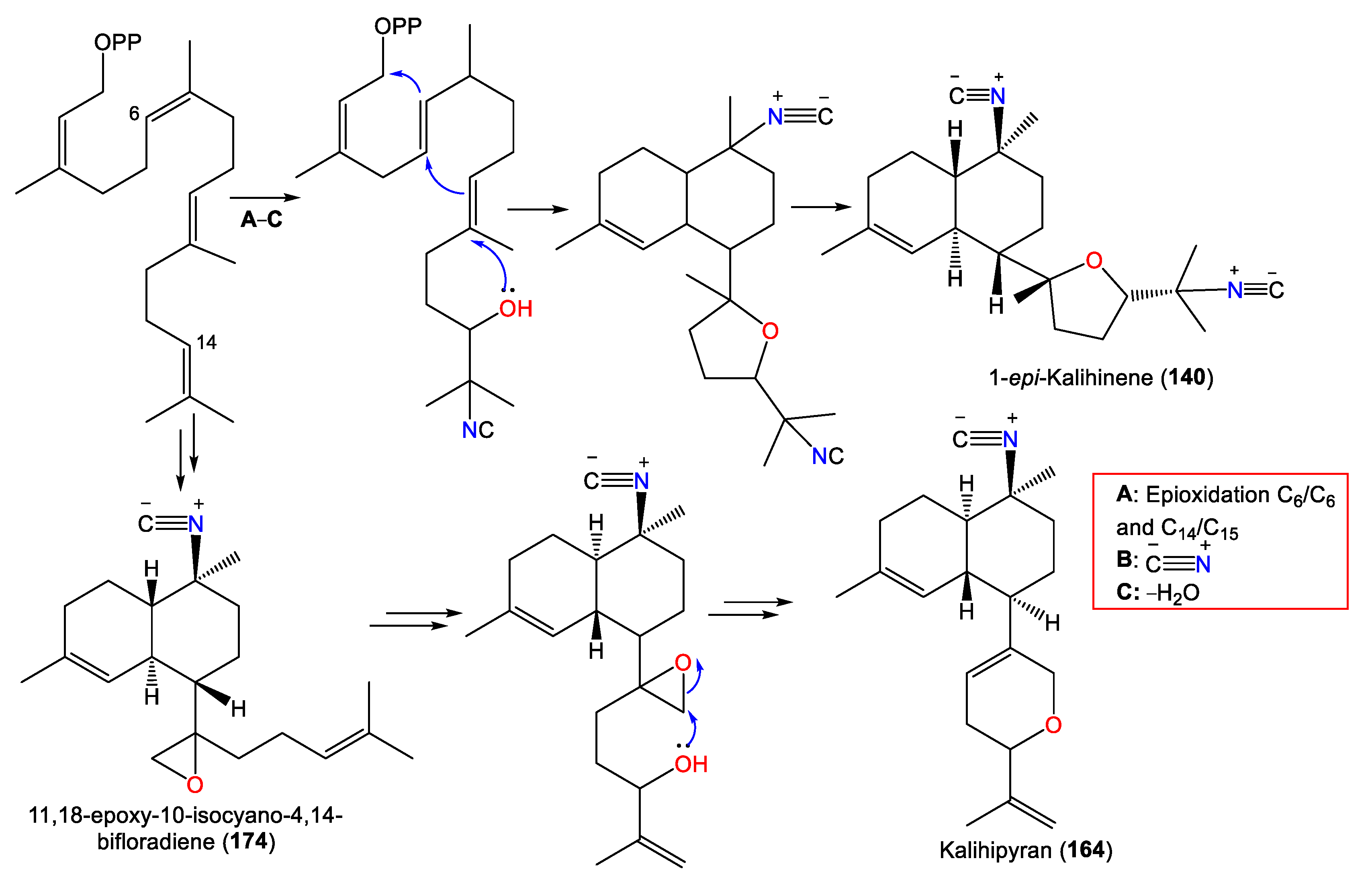
3.2.2. Kalihinenes
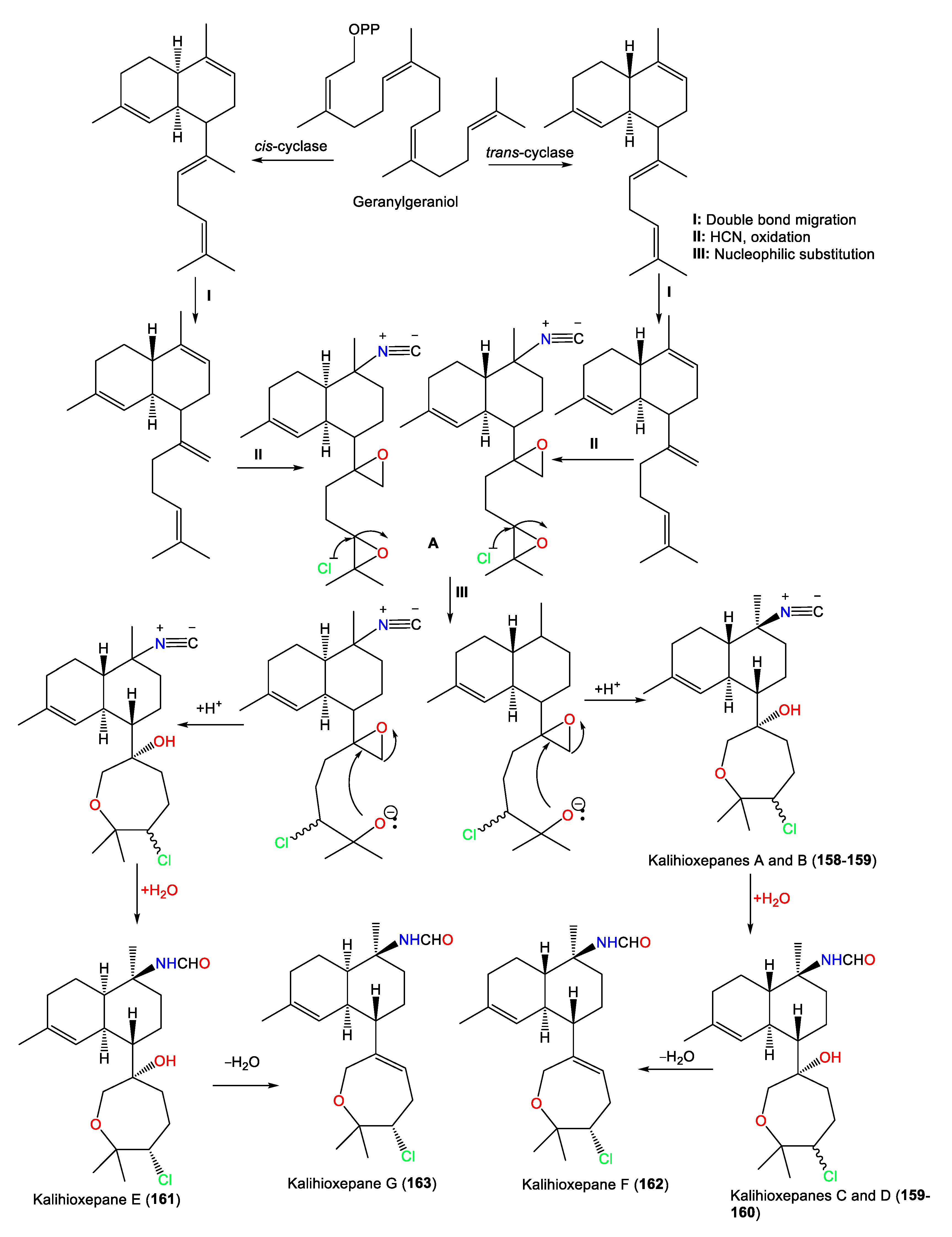
3.2.3. Kalihipyrans and Kalihioxepanes
3.2.4. Biflorane Diterpenes
3.3. Alkaloids
3.4. Steroid Compounds
3.5. Other Metabolites
4. Biological Activities of Acanthella Species and Their Metabolites
4.1. Antimicrobial and Antifouling Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
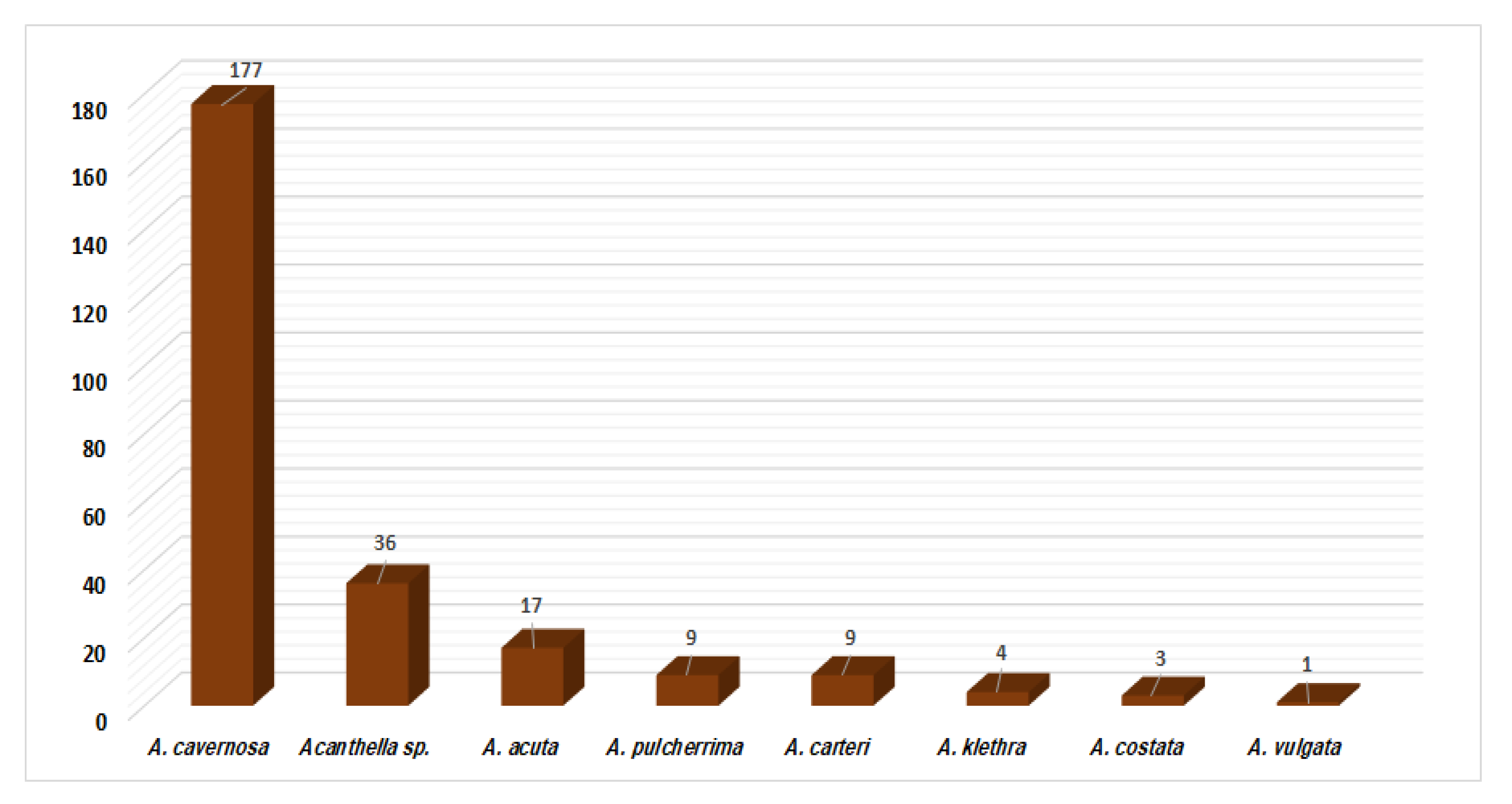{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Biological Activity | Assay, Organism or Cell Line | Biological Results | Ref. | |
|---|---|---|---|---|---|
| Compound | Positive Control | ||||
| (+)-10(R)-Isothiocyanatoalloaromadendrane (9) | Cytotoxicity | MTT/A549 | 1.98 µg/mL (IC50) | - | [42] |
| Axisonitrile-3 (17) | Antimalarial | Plasmodium falciparum-D6/Microculture radioisotope | 142.0 ng/mL (IC50) | Chloroquine 1.95 ng/mL (IC50) | [19] |
| Plasmodium falciparum-W2/Microculture radioisotope | 16.5 ng/mL (IC50) | Chloroquine 22.8 ng/mL (IC50) | [19] | ||
| Cytotoxicity | MTT/A549 | 2.44 µg/mL (IC50) | - | [42] | |
| 11-Isothiocyano-7βH-eudesm-5-ene (42) | Antimalarial | Plasmodium falciparum-D6/Microculture radioisotope | 2240.0 ng/mL (IC50) | Chloroquine 1.95 ng/mL (IC50) | [19] |
| Plasmodium falciparum-W2/Microculture radioisotope | 610.0 ng/mL (IC50) | Chloroquine 22.8 ng/mL (IC50) | [19] | ||
| (lR,5R,6R,8S)-Dec[4.4.0]ane-1,5-dimethyl-8-(1′-methylethenyl)-5-isothiocyanate (45) | Antimalarial | Plasmodium falciparum-D6/Microculture radioisotope | 4000.0 ng/mL (IC50) | Chloroquine 1.95 ng/mL (IC50) | [19] |
| Plasmodium falciparum-W2/Microculture radioisotope | 550.0 ng/mL (IC50) | Chloroquine 22.8 ng/mL (IC50) | [19] | ||
| Kalihinol A (88) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 50.0 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| Disc-diffusion/Ruegeria CtaxMed-2 | 0.67 mm (IZD) at 10 μg/Disc | Streptomycin 0.83 mm (IZD) at 10 μg/Disc | [58] | ||
| Disc-diffusion/Vibrio sp. (NAP-4) | 1.0 mm (IZD) at 10 μg/Disc | Streptomycin 1.83 mm (IZD) at 10 μg/Disc | [58] | ||
| Disc-diffusion/Vibrio furnissii | 6.33 mm (IZD) at 10 μg/Disc | Streptomycin 2.50 mm (IZD) at 10 μg/Disc | [58] | ||
| Antifouling | Settlement inhibition/Balanus amphitrite | 0.087 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55,56] | |
| Settlement inhibition/Balanus amphitrite | 0.92 µM (EC50) | Seawater + DMSO | [16] | ||
| Cytotoxicity | MTT/HCT-116 | 17.40 µM (IC50) | - | [16] | |
| trans and cis 10β-Formamidokalihinol A (89/90) | Antibacterial | Disc-diffusion/Vibrio sp. (NAP-4) | 1.0 mm (IZD) at 10 μg/Disc | Streptomycin 1.83 mm (IZD) at 10 μg/Disc | [58] |
| Disc-diffusion/Vibrio furnissii | 5.67 mm (IZD) at 10 μg/Disc | Streptomycin 2.50 mm (IZD) at 10 μg/Disc | [58] | ||
| Antifouling | Settlement inhibition/Balanus amphitrite | 1.37 µM (EC50) | Seawater + DMSO | [16] | |
| Cytotoxicity | MTT/CT-26 | 28.82 µM (IC50) | - | [16] | |
| 10β-Formamido-5β-isothiocyanatokalihinol A (92) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.41 µM (EC50) | Seawater + DMSO | [16] |
| Isokalihinol B (95) | Cytotoxicity | MTT/P388 | 0.8 μg/mL | - | [23] |
| Kalihinol D (97) | Cytotoxicity | MTT/A549 | 3.17 μg/mL (IC50) | - | [42] |
| Kalihinol E (98) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.85 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 18.31 µM (IC50) | - | [16] | |
| Kalihinol F (100) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 12.5 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| Trans 10-Formamido-kalihinol F (104/105) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 50.0 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| 15-Formamido-kalihinol F (106/107) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 50.0 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| Kalihinol G (108) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 3.12 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| 10-epi-Kalihinol I (115) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.27 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 28.67 µM (IC50) | - | [16] | |
| Kalihinol J (116) | Antibacterial | Microbroth dilution/B. subtilis PY79 | 50.0 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] |
| Kalihinol O (119) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.43 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 5.97 µM (IC50) | - | [16] | |
| Kalihinol P (120) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.72 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 10.68 µM (IC50) | - | [16] | |
| MTT/H1299 | 26.21 µM (IC50) | - | [16] | ||
| Kalihinol Q (121) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.48 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 20.55 µM (IC50) | - | [16] | |
| Kalihinol R (122) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.16 µM (EC50) | Seawater + DMSO | [16] |
| Cytotoxicity | MTT/HCT-116 | 13.44 µM (IC50) | - | [16] | |
| Kalihinol S (123) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.53 µM (EC50) | Seawater + DMSO | [16] |
| Kalihinol T (124) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.74 µM (EC50) | Seawater + DMSO | [16] |
| 10-epi-Kalihinol X (126) | Cytotoxicity | MTT/A549 | 9.30 µg/mL (IC50) | - | [42] |
| Antifouling | Settlement inhibition/Balanus amphitrite | 0.69 µM (EC50) | Seawater + DMSO | [16] | |
| Cytotoxicity | MTT/HCT-116 | 8.21 µM (IC50) | - | [16] | |
| Kalihiacyloxyamide C (133) | Cytotoxicity | SRB/L-02 | 14.6 μM (IC50) | Doxorubicin not detected | [62] |
| MTT/K562 | 6.4 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| SRB/ASPC-1 | 19.0 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| SRB/H69AR | 12.0 μM (IC50) | Doxorubicin 15.1 μM (IC50) | [62] | ||
| SRB/MDA-MB-231 | 12.5 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| Kalihiacyloxyamide D (134) | Cytotoxicity | SRB/L-02 | 8.0 μM (IC50) | Doxorubicin not detected | [62] |
| MTT/K562 | 6.3 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| SRB/MDA-MB-231 | 7.3 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| Kalihiacyloxyamide G (137) | Cytotoxicity | SRB/L-02 | 19.2 μM (IC50) | Doxorubicin not detected | [62] |
| MTT/K562 | 15.0 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| SRB/MDA-MB-231 | 13.4 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| Kalihiacyloxyamide H (138) | Cytotoxicity | SRB/H69AR | 16.8 μM (IC50) | Doxorubicin 15.1 μM (IC50) | [62] |
| SRB/MDA-MB-231 | 12.5 μM (IC50) | Doxorubicin ˂ 1.04 μM (IC50) | [62] | ||
| Kalihinene (139) | Cytotoxicity | MTT/P388 | 1.2 μg/mL | - | [23] |
| Antibacterial | Microbroth dilution/B. subtilis PY79 | 6.25 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] | |
| Kalihinene E (145) | Cytotoxicity | MTT/HCT-116 | 14.36 μM (IC50) | Camptothecin 9.25 μM (IC50) | [63] |
| MTT/HeLa | 13.36 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 17.78 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| MTT/MDA-MB-231 | 12.84 μM (IC50) | Camptothecin 0.50 μM (IC50) | [63] | ||
| Kalihinene X (147) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.49 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55,56] |
| Antibacterial | Microbroth dilution/B. subtilis PY79 | 1.56 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] | |
| Cytotoxicity | MTT/HCT-116 | 12.25 μM (IC50) | Camptothecin 9.25 μM (IC50) | [63] | |
| MTT/A549 | 8.55 μM (IC50) | Camptothecin 2.32 μM (IC50) | [63] | ||
| MTT/HeLa | 10.59 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 13.02 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| MTT/MDA-MB-231 | 7.46 μM (IC50) | Camptothecin 0.50 μM (IC50) | [63] | ||
| Kalihinene Y (148) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.45 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55,56] |
| Antibacterial | Microbroth dilution/B. subtilis PY79 | 1.56 μg/mL (MIC) | -Trimethoprim 1.0 μg/mL (MIC) -Ciprofloxacin 1.0 μg/mL (MIC) -Rifampin 1.0 μg/mL (MIC) -Sulfamethoxazole 4.0 μg/mL (MIC) -Chloramphenicol 16.0 μg/mL (MIC) -Polymyxin B 16.0 μg/mL (MIC) | [21] | |
| Cytotoxicity | MTT/A549 | 16.12 μM (IC50) | Camptothecin 2.32 μM (IC50) | [63] | |
| MTT/HeLa | 10.05 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 14.41 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| MTT/MDA-MB-231 | 15.23 μM (IC50) | Camptothecin 0.50 μM (IC50) | [63] | ||
| Kalihinene Z (149) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.1 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55,56] |
| 10-Formamidokalihinene (150) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.095 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55,56] |
| Cytotoxicity | MTT/A549 | 6.98 μM (IC50) | Camptothecin 2.32 μM (IC50) | [63] | |
| MTT/HeLa | 13.30 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 14.53 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| MTT/MDA-MB-231 | 6.84 μM (IC50) | Camptothecin 0.50 μM (IC50) | [63] | ||
| 15-Formamidokalihinene (151) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.14 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55] |
| MTT/A549 | 17.53 μM (IC50) | Camptothecin 2.32 μM (IC50) | [63] | ||
| MTT/HeLa | 14.74 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 16.39 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| Kalihipyran A (165) | Antifouling | Settlement inhibition/Balanus amphitrite | 1.3 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55] |
| Cytotoxicity | MTT/A549 | 13.09 μM (IC50) | Camptothecin 2.32 μM (IC50) | [63] | |
| MTT/HeLa | 11.19 μM (IC50) | Camptothecin 6.98 μM (IC50) | [63] | ||
| MTT/QGY-7701 | 13.53 μM (IC50) | Camptothecin 4.05 μM (IC50) | [63] | ||
| Kalihipyran B (166) | Antifouling | Settlement inhibition/Balanus amphitrite | 0.85 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55] |
| Cavernene A (169) | Cytotoxicity | MTT/HCT-116 | 6.31 μM (IC50) | Camptothecin 9.25 μM (IC50) | [63] |
| Cytotoxicity | MTT/HCT-116 | 8.99 μM (IC50) | Camptothecin 9.25 μM (IC50) | [63] | |
| Biflora-4,9,15-triene (168) | Antifouling | Settlement inhibition/Balanus amphitrite | 9.7 µg/mL (IC50) | CuSO4 0.15 µg/mL (IC50) | [55] |
| Hymenialdisine (185) | Cytotoxicity | AlamarBlue/A2780S | 146.8 μM (IC50) | Cisplatin 31.4 μM (IC50) | [82] |
| 2β-Hydroxy-4,7-diketo-A-norcholest-5-en-2-oic acid (208) | Antifouling | Settlement inhibition/Balanus albicostatus | 8.2 μg/mL EC50) | Capsaicin 1.32 μg/mL (EC50) | [74] |
| 24S-Ethyl-2β-hydroxy-4,7-diketo-A-norcholest-5-en-2-oic acid (209) | Antifouling | Settlement inhibition/Balanus albicostatus | 23.5 μg/mL (EC50) | Capsaicin 1.32 μg/mL (EC50) | [74] |
| 2β-Hydroxy-4,7-diketo-24R-methyl-A-norcholest-5,22(E)-diene-2-oic acid (210) | Antifouling | Settlement inhibition/Balanus albicostatus | 31.6 μg/mL (EC50) | Capsaicin 1.32 μg/mL (EC50) | [74] |
4.2. Cytotoxic Activity
4.3. Larvicidal and Antimalarial Activities
4.4. Cyclooxygenase Inhibitory and α2B Adrenoceptor Agonistic Activities
4.5. Insecticidal and Anthelmintic Activities
4.6. Antioxidant Activity
5. Nanoparticles
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| A549 | Human lung adenocarcinoma epithelial cell line |
| A2780s | Human ovarian cancer cell line |
| A2780CP | Human ovarian cancer cell line |
| ASPC-1 | Human pancreatic cancer cell line |
| CCL2 | C–C motif chemokine ligand 2 |
| CD | Circular dichroism |
| CH2Cl2 | Dichloromethane |
| CHCl3 | Chloroform |
| COX-2 | Cyclooxygenase-2 |
| CT-26 | Murine colorectal carcinoma cell line |
| DKPs | Diketopiperazines |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| EC50 | Half maximal effective concentration |
| EtOAc | Ethyl acetate |
| H69 | Chemo-sensitive human small cell lung cancer cell line |
| H69AR | Chemo-resistant human small cell lung cancer cell line |
| H1299 | Human non-small cell lung carcinoma cell line |
| HCT-116 | Human colon cancer cell line |
| HeLa | Human cervical epithelioid carcinoma cell line |
| HPLC | High-performance liquid chromatography |
| HRESIMS | High-resolution electrospray ionization mass spectroscopy |
| HT-29 | Human colon cancer cell line |
| IZD | Inhibition zone diameter |
| K562 | Human erythroleukemic cell line |
| KB | Human oral epidermoid carcinoma cell line |
| L02 | Human liver cell line |
| LC50 | Lethal concentration that kills 50% |
| LD50 | Half maximal lethal concentration |
| LD100 | Maximal lethal concentration |
| IR | Infrared |
| LPS | Lipopolysaccharide |
| Ls174T | Human colorectal cancer cell line |
| MDA-MB-231 | Human breast cancer cell line |
| MeOH | Methanol |
| MRC-5 | Human lung fibroblasts |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| n-BuOH | n-butanol |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NMR | Nuclear magnetic resonance |
| P 388 | Human leukemia cell line |
| PC-3 | Human prostatic testosterone-independent cell line |
| PDCD4 | Programmed cell death 4 |
| QGY-7701 | Human hepatocellular carcinoma cell line |
| QM-NMR | Quantum mechanical nuclear magnetic resonance |
| RP-18 | Reversed phase-18 |
| SiO2 CC | Silica gel column chromatography |
| T47D | Human hormone-dependent breast cancer cell line |
| TDDFT-ECD | Time-dependent density functional theory/electronic circular dichroism |
| TNF-α | Tumor necrosis factor alpha |
| TPA | Tetradecanoylphorbol-13-acetate |
References
- Ibrahim, S.R.M.; Fadil, S.A.; Fadil, H.A.; Eshmawi, B.A.; Mohamed, S.G.A.; Mohamed, G.A. Fungal Naphthalenones; Promising Metabolites for Drug Discovery: Structures, Biosynthesis, Sources, and Pharmacological Potential. Toxins 2022, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Mohamed, G.A.; Ibrahim, S.R. Lansium domesticum—A Fruit with Multi-Benefits: Traditional Uses, Phytochemicals, Nutritional Value, and Bioactivities. Nutrients 2022, 14, 1531. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Choudhry, H.; Asseri, A.H.; Elfaky, M.A.; Mohamed, S.G.; Mohamed, G.A. Stachybotrys chartarum—A Hidden Treasure: Secondary Metabolites, Bioactivities, and Biotechnological Relevance. J. Fungi 2022, 8, 504. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.; Mohamed, G.A. Bright Side of Fusarium oxysporum: Secondary Metabolites Bioactivities and Industrial Relevance in Biotechnology and Nanotechnology. J. Fungi 2021, 7, 943. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Fadil, S.A.; Fadil, H.A.; Hareeri, R.H.; Abdallah, H.M.; Mohamed, G.A. Ethnobotanical Uses, Phytochemical Composition, Biosynthesis, and Pharmacological Activities of Carpesium abrotanoides L. (Asteraceae). Plants 2022, 11, 1598. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khedr, A.I.M.; Zayed, M.F.; El-Kholy, A.A.S. Genus Hylocereus: Beneficial Phytochemicals, Nutritional Importance, and Biological Relevance—A Review. J. Food Biochem. 2018, 42, e12491. [Google Scholar] [CrossRef]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine Natural Products as Source of New Drugs: A Patent Review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef]
- Banerjee, P.; Mandhare, A.; Bagalkote, V. Marine Natural Products as Source of New Drugs: An Updated Patent Review (July 2018–July 2021). Expert Opin. Ther. Pat. 2022, 32, 317–363. [Google Scholar] [CrossRef] [PubMed]
- Kollár, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine Natural Products: Bryostatins in Preclinical and Clinical Studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef]
- Rangel, M.; Falkenberg, M. An Overview of the Marine Natural Products in Clinical Trials and on the Market. J. Coast. Life Med. 2015, 3, 421–428. [Google Scholar]
- Haque, N.; Parveen, S.; Tang, T.; Wei, J.; Huang, Z. Marine Natural Products in Clinical Use. Mar. Drugs 2022, 20, 528. [Google Scholar] [CrossRef]
- Khan, S.; Al-Fadhli, A.A.; Tilvi, S. Discovery of Cytotoxic Natural Products from Red Sea Sponges: Structure and Synthesis. Eur. J. Med. Chem. 2021, 220, 113491. [Google Scholar] [CrossRef]
- Maslin, M.; Gaertner-Mazouni, N.; Debitus, C.; Joy, N.; Ho, R. Marine Sponge Aquaculture Towards Drug Development: An Ongoing History of Technical, Ecological, Chemical Considerations and Challenges. Aquacult. Rep. 2021, 21, 100813. [Google Scholar] [CrossRef]
- de Voogd, N.J.; Alvarez, B.; Boury-Esnault, N.; Carballo, J.L.; Cárdenas, P.; Díaz, M.-C.; Dohrmann, M.; Downey, R.; Goodwin, C.; Hajdu, E.; et al. World Porifera Database. Acanthella Schmidt, 1862. 2023. Available online: https://www.marinespecies.org/porifera/porifera.php?p=taxdetails&id=165648 (accessed on 8 April 2023).
- Emsermann, J.; Kauhl, U.; Opatz, T. Marine Isonitriles and their Related Compounds. Mar. Drugs 2016, 14, 16. [Google Scholar] [CrossRef]
- Xu, Y.; Li, N.; Jiao, W.; Wang, R.; Peng, Y.; Qi, S.; Song, S.; Chen, W.; Lin, H. Antifouling and Cytotoxic Constituents from the South China Sea Sponge Acanthella cavernosa. Tetrahedron 2012, 68, 2876–2883. [Google Scholar] [CrossRef]
- Macabeo, A.P.G.; Guce, F.d. Bromopyrrole-Imidazole Alkaloids from Acanthella carteri Dendy (Axinellidae). Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 720–723. [Google Scholar]
- Alvi, K.A.; Tenenbaum, L.; Crews, P. Anthelmintic Polyfunctional Nitrogen-Containing Terpenoids from Marine Sponges. J. Nat. Prod. 1991, 54, 71–78. [Google Scholar] [CrossRef]
- Angerhofer, C.K.; Pezzuto, J.M.; König, G.M.; Wright, A.D.; Sticher, O. Antimalarial Activity of Sesquiterpenes from the Marine Sponge Acanthella klethra. J. Nat. Prod. 1992, 55, 1787–1789. [Google Scholar] [CrossRef] [PubMed]
- Annie Selva Sonia, G.; Lipton, A.P.; Paul Raj, R. Antibacterial Activity of Marine Sponge Extracts Against Fish Pathogenic Bacteria. Isr. J. Aquac.—Bamidgeh 2008, 60, 172–176. [Google Scholar] [CrossRef]
- Bugni, T.S.; Singh, M.P.; Chen, L.; Arias, D.A.; Harper, M.K.; Greenstein, M.; Maiese, W.M.; Concepción, G.P.; Mangalindan, G.C.; Ireland, C.M. Kalihinols from Two Acanthella cavernosa Sponges: Inhibitors of Bacterial Folate Biosynthesis. Tetrahedron 2004, 60, 6981–6988. [Google Scholar] [CrossRef]
- Chang, C.W.; Patra, A.; Baker, J.A.; Scheuer, P.J. Kalihinols, Multifunctional Diterpenoid Antibiotics from Marine Sponges Acanthella Spp. J. Am. Chem. Soc. 1987, 109, 6119–6123. [Google Scholar] [CrossRef]
- Fusetani, N.; Yasumuro, K.; Kawai, H.; Natori, T.; Brinen, L.; Clardy, J. Kalihinene and Isokalihinol B, Cytotoxic Diterpene Isonitriles from the Marine Sponge Acanthella klethra. Tetrahedron Lett. 1990, 31, 3599–3602. [Google Scholar] [CrossRef]
- Grkovic, T.; Blees, J.S.; Bayer, M.M.; Colburn, N.H.; Thomas, C.L.; Henrich, C.J.; Peach, M.L.; McMahon, J.B.; Schmid, T.; Gustafson, K.R. Tricyclic Guanidine Alkaloids from the Marine Sponge Acanthella cavernosa that Stabilize the Tumor Suppressor PDCD4. Mar. Drugs 2014, 12, 4593–4601. [Google Scholar] [CrossRef]
- Hammami, S.; Bergaoui, A.; Boughalleb, N.; Romdhane, A.; Khoja, I.; Kamel, M.B.H.; Mighri, Z. Antifungal Effects of Secondary Metabolites Isolated from Marine Organisms Collected from the Tunisian Coast. C. R. Chim. 2010, 13, 1397–1400. [Google Scholar] [CrossRef]
- Nishikawa, K.; Umezawa, T.; Garson, M.J.; Matsuda, F. Confirmation of the Configuration of 10-Isothiocyanato-4-Cadinene Diastereomers through Synthesis. J. Nat. Prod. 2012, 75, 2232–2235. [Google Scholar] [CrossRef] [PubMed]
- White, R.D.; Keaney, G.F.; Slown, C.D.; Wood, J.L. Total Synthesis of (±)-Kalihinol C. Org. Lett. 2004, 6, 1123–1126. [Google Scholar] [CrossRef] [PubMed]
- Miyaoka, H.; Abe, Y.; Sekiya, N.; Mitome, H.; Kawashima, E. Total Synthesis of Antimalarial Diterpenoid (+)-Kalihinol A. Chem. Comm. 2012, 48, 901–903. [Google Scholar] [CrossRef] [PubMed]
- White, R.D.; Wood, J.L. Progress Toward the Total Synthesis of Kalihinane Diterpenoids. Org. Lett. 2001, 3, 1825–1827. [Google Scholar] [CrossRef]
- Minale, L.; Riccio, R.; Sodano, G. Acanthellin-1, an Unique Isonitrile Sesquiterpene from the Sponge Acanthella acuta. Tetrahedron 1974, 30, 1341–1343. [Google Scholar] [CrossRef]
- Dumdei, E.J.; Flowers, A.E.; Garson, M.J.; Moore, C.J. The Biosynthesis of Sesquiterpene Isocyanides and Isothiocyanates in the Marine Sponge Acanthella cavernosa (Dendy); Evidence for Dietary Transfer to the Dorid Nudibranch Phyllidiella pustulosa. Comp. Biochem. Physiol. Part A Physiol. 1997, 118, 1385–1392. [Google Scholar] [CrossRef]
- Fusetani, N.; Wolstenholme, H.J.; Shinoda, K.; Asai, N.; Matsunaga, S.; Onuki, H.; Hirota, H. Two Sesquiterpene Isocyanides and a Sesquiterpene Thiocyanate from the Marine Sponge Acanthella Cf. cavernosa and the Nudibranch Phyllidia ocellata. Tetrahedron Lett. 1992, 33, 6823–6826. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, Z.; Yao, L.; Wang, J.; Guo, Y.; Li, X. Nitrogenous Sesquiterpenoids from the South China Sea Nudibranch Hexabranchus sanguineus and its Possible Sponge-Prey Acanthella Cavernosa: Chiral Separation, Stereochemistry and Chemical Ecology. Chin. J. Chem. 2022, 40, 235–246. [Google Scholar] [CrossRef]
- Braekman, J.C.; Daloze, D.; Deneubourg, F.; Huysecom, J.; Vandevyver, G. I-Isocyanoaromadendrane, A New Isonitrile Sesquiterpene from the Sponge Acanthella acuta. Bull. Sociétés Chim. Belg. 1987, 96, 539–543. [Google Scholar] [CrossRef]
- Capon, R.J.; MacLeod, J.K. New Isothiocyanate Sesquiterpenes from the Australian Marine Sponge Acanthella pulcherrima. Aust. J. Chem. 1988, 41, 979–983. [Google Scholar] [CrossRef]
- Braekman, J.C.; Daloze, D.; Moussiaux, B.; Stoller, C.; Deneubourg, F. Sponge Secondary Metabolites: New Results. Pure Appl. Chem. 1989, 61, 509–512. [Google Scholar] [CrossRef]
- Jumaryatno, P.; Rands-Trevor, K.; Blanchfield, J.T.; Garson, M.J. Isocyanates in Marine Sponges: Axisocyanate-3, a New Sesquiterpene from Acanthella cavernosa. ARKIVOC 2007, vii, 157–166. [Google Scholar] [CrossRef]
- Jumaryatno, P.; Stapleton, B.L.; Hooper, J.N.; Brecknell, D.J.; Blanchfield, J.T.; Garson, M.J. A Comparison of Sesquiterpene Scaffolds Across Different Populations of the Tropical Marine Sponge Acanthella cavernosa. J. Nat. Prod. 2007, 70, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Mayol, L.; Piccialli, V.; Sica, D. Nitrogenous Sesquiterpenes from the Marine Sponge Acanthella acuta: Three New Isocyanide-Isothiocyanate Pairs. Tetrahedron 1987, 43, 5381–5388. [Google Scholar] [CrossRef]
- Hirota, H.; Tomono, Y.; Fusetani, N. Terpenoids with Antifouling Activity Against Barnacle Larvae from the Marine Sponge Acanthella cavernosa. Tetrahedron 1996, 52, 2359–2368. [Google Scholar] [CrossRef]
- Yan, X.; Zhu, X.; Yu, J.; Jin, D.; Guo, Y.; Mollo, E.; Cimino, G. 3-Oxo-Axisonitrile-3, a New Sesquiterpene Isocyanide from the Chinese Marine Sponge Acanthella Sp. J. Asian Nat. Prod. Res. 2006, 8, 579–584. [Google Scholar] [CrossRef]
- Sun, J.; Chen, K.; Yao, L.; Liu, H.; Guo, Y. A New Kalihinol Diterpene from the Hainan Sponge Acanthella Sp. Arch. Pharm. Res. 2009, 32, 1581–1584. [Google Scholar] [CrossRef]
- Shen, S.; Yang, Q.; Zang, Y.; Li, J.; Liu, X.; Guo, Y. Anti-Inflammatory Aromadendrane-and Cadinane-Type Sesquiterpenoids from the South China Sea Sponge Acanthella cavernosa. Beilstein J. Org. Chem. 2022, 18, 916–925. [Google Scholar] [CrossRef] [PubMed]
- König, G.M.; Wright, A.D.; Sticher, O.; Fronczek, F.R. Two New Sesquiterpene Isothiocyanates from the Marine Sponge Acanthella klethra. J. Nat. Prod. 1992, 55, 633–638. [Google Scholar] [CrossRef]
- Clark, R.J.; Stapleton, B.L.; Garson, M.J. New Isocyano and Isothiocyanato Terpene Metabolites from the Tropical Marine Sponge Acanthella cavernosa. Tetrahedron 2000, 56, 3071–3076. [Google Scholar] [CrossRef]
- Burgoyne, D.L.; Dumdei, E.J.; Andersen, R.J. Acanthenes A to C: A Chloro, Isothiocyanate, Formamide Sesquiterpene Triad Isolated from the Northeastern Pacific Marine Sponge Acanthella Sp. and the Dorid Nudibranch Cadlina luteomarginata. Tetrahedron 1993, 49, 4503–4510. [Google Scholar] [CrossRef]
- Ciminiello, P.; Magno, S.; Mayol, L.; Piccialli, V. Cis-Eudesmane Nitrogenous Metabolites from the Marine Sponges Axinella Cannabina and Acanthella acuta. J. Nat. Prod. 1987, 50, 217–220. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, W.; Li, S.; Ye, J.; Huan, X.; Gavagnin, M.; Yao, L.; Wang, H.; Miao, Z.; Li, X. Cytotoxic Nitrogenous Terpenoids from Two South China Sea Nudibranchs Phyllidiella pustulosa, Phyllidia Coelestis, and their Sponge-Prey Acanthella cavernosa. Mar. Drugs 2019, 17, 56. [Google Scholar] [CrossRef]
- Nogata, Y.; Yoshimura, E.; Shinshima, K.; Kitano, Y.; Sakaguchi, I. Antifouling Substances Against Larvae of the Barnacle Balanus Amphitrite from the Marine Sponge, Acanthella cavernosa. Biofouling 2003, 19, 193–196. [Google Scholar] [CrossRef]
- Fan, W.; Wang, X.; Cai, H.; Sun, L.; Yang, L.; Nie, S. Chemical Analysis of the South China Sea Spine Body Sponge Acanthella cavernosa. J. Pharm. Pract. 2016, 34, 138–141, 166. [Google Scholar]
- Rodríguez, J.; Nieto, R.M.; Hunter, L.M.; Diaz, M.C.; Crews, P.; Lobkovsky, E.; Clardy, J. Variation among Known Kalihinol and New Kalihinene Diterpenes from the Sponge Acanthella cavernosa. Tetrahedron 1994, 50, 11079–11090. [Google Scholar] [CrossRef]
- Chang, C.W.; Patra, A.; Roll, D.M.; Scheuer, P.J.; Matsumoto, G.K.; Clardy, J. Kalihinol-A, a Highly Functionalized Diisocyano Diterpenoid Antibiotic from a Sponge. J. Am. Chem. Soc. 1984, 106, 4644–4646. [Google Scholar] [CrossRef]
- Omar, S.; Albert, C.; Fanni, T.; Crews, P. Polyfunctional Diterpene Isonitriles from Marine Sponge Acanthella carvenosa. J. Org. Chem. 1988, 53, 5971–5972. [Google Scholar] [CrossRef]
- Braekman, J.C.; Daloze, D.; Gregoire, F.; Popov, S.; van Soest, R. Two New Kalihinenes from the Marine Sponge Acanthella cavernosa. Bull. Soc. Chim. Belg. 1994, 103, 187–191. [Google Scholar] [CrossRef]
- Okino, T.; Yoshimura, E.; Hirota, H.; Fusetani, N. New Antifouling Kalihipyrans from the Marine Sponge Acanthella vavernosa. J. Nat. Prod. 1996, 59, 1081–1083. [Google Scholar] [CrossRef]
- Okino, T.; Yoshimura, E.; Hirota, H.; Fusetani, N. Antifouling Kalihinenes from the Marine Sponge Acanthella cavernosa. Tetrahedron Lett. 1995, 36, 8637–8640. [Google Scholar] [CrossRef]
- Miyaoka, H.; Shimomura, M.; Kimura, H.; Yamada, Y.; Kim, H.; Yusuke, W. Antimalarial Activity of Kalihinol A and New Relative Diterpenoids from the Okinawan Sponge, Acanthella Sp. Tetrahedron 1998, 54, 13467–13474. [Google Scholar] [CrossRef]
- Yang, L.H.; Lee, O.O.; Jin, T.; Li, X.C.; Qian, P.Y. Antifouling Properties of 10β-Formamidokalihinol-A and Kalihinol A Isolated from the Marine Sponge Acanthella cavernosa. Biofouling 2006, 22, 23–32. [Google Scholar] [CrossRef]
- Trimurtulu, G.; Faulkner, D.J. Six New Diterpene Isonitriles from the Sponge Acanthella cavernosa. J. Nat. Prod. 1994, 57, 501–506. [Google Scholar] [CrossRef]
- Karuso, P.; Scheuer, P.J. Biosynthesis of Isocyanoterpenes in Sponges. J. Org. Chem. 1989, 54, 2092–2095. [Google Scholar] [CrossRef]
- Ohta, E.; Ohta, S.; Hongo, T.; Hamaguchi, Y.; Andoh, T.; Shioda, M.; Ikegami, S. Inhibition of Chromosome Separation in Fertilized Starfish Eggs by Kalihinol F, a Topoisomerase I Inhibitor obtained from a Marine Sponge. Biosci. Biotechnol. Biochem. 2003, 67, 2365–2372. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Han, X.; Zhang, D.; Hou, H.; Xiao, L.; Li, G. Kalihiacyloxyamides AH, A-Acyloxy Amide Substituted Kalihinane Diterpenes Isolated from the Sponge Acanthella cavernosa Collected in the South China Sea. Phytochemistry 2023, 206, 113512. [Google Scholar] [CrossRef]
- Xu, Y.; Lang, J.; Jiao, W.; Wang, R.; Peng, Y.; Song, S.; Zhang, B.; Lin, H. Formamido-Diterpenes from the South China Sea Sponge Acanthella cavernosa. Mar. Drugs 2012, 10, 1445–1458. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Han, X.; Liu, G.; Zhang, D.; Hou, H.; Xiao, L.; de Voogd, N.J.; Tang, X.; Li, P.; Li, G. Kalihioxepanes A—G: Seven Kalihinene Diterpenoids from Marine Sponge Acanthella cavernosa Collected Off the South China Sea. Chin. J. Chem. 2022, 40, 1785–1792. [Google Scholar] [CrossRef]
- Shimomura, M.; Miyaoka, H.; Yamada, Y. Absolute Configuration of Marine Diterpenoid Kalihinol A. Tetrahedron Lett. 1999, 40, 8015–8017. [Google Scholar] [CrossRef]
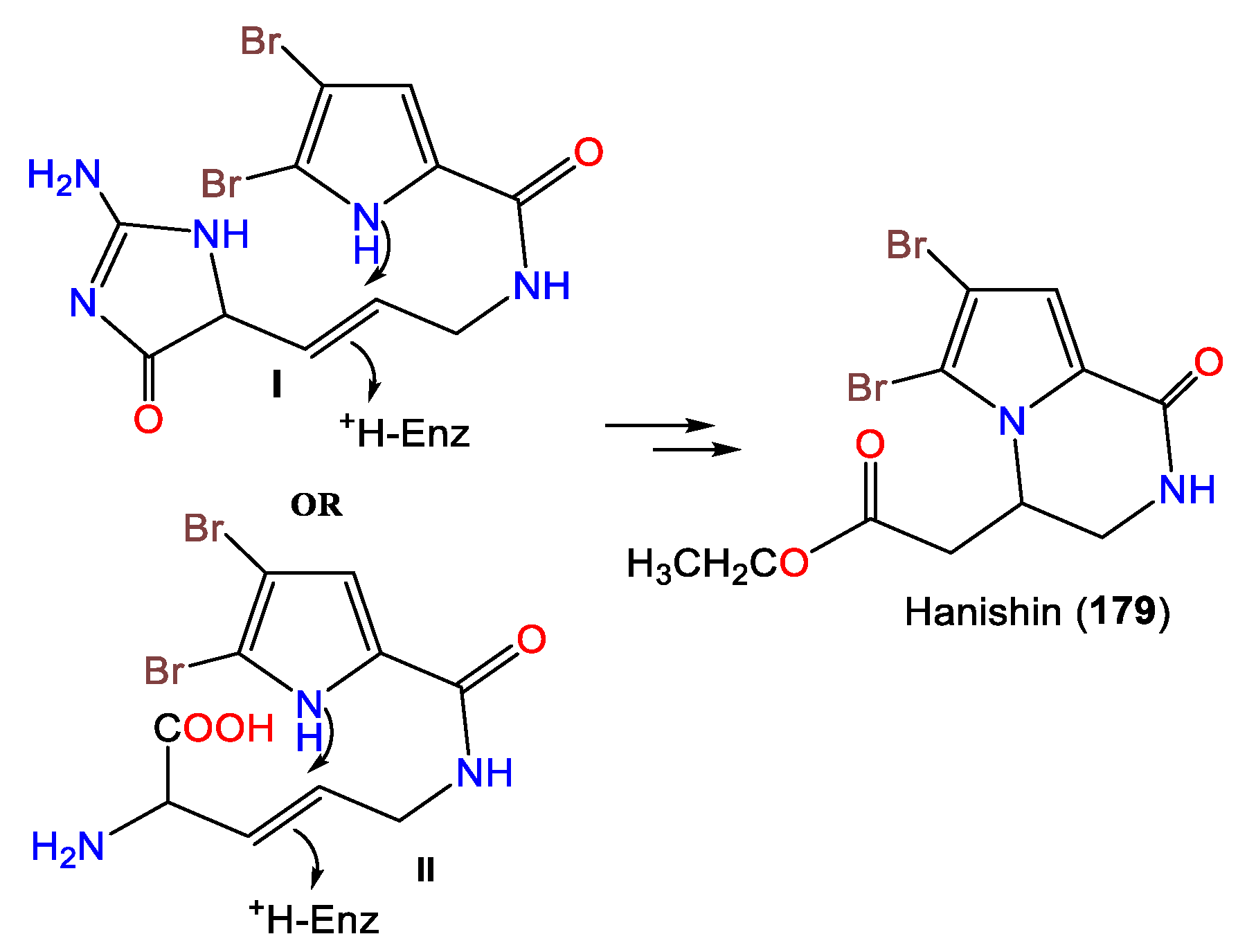
- Mancini, I.; Guella, G.; Amade, P.; Roussakis, C.; Pietra, F. Hanishin, a Semiracemic, Bioactive C9 Alkaloid of the Axinellid Sponge Acanthella ccarteri from the Hanish Islands. A Shunt Metabolite? Tetrahedron Lett. 1997, 38, 6271–6274. [Google Scholar] [CrossRef]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Mazzarella, L.; Puliti, R.; Sodano, G. Isolation and X-Ray Crystal Structure of a Novel Bromo-Compound from Two Marine Sponges. Tetrahedron Lett. 1982, 23, 767–768. [Google Scholar] [CrossRef]
- Mattia, C.A.; Mazzarella, L.; Puliti, R. 4-(2-Amino-4-Oxo-2-Imidazolin-5-Ylidene)-2-Bromo-4, 5, 6, 7-Tetrahydropyrrolo [2, 3-C] Azepin-8-One Methanol Solvate: A New Bromo Compound from the Sponge Acanthella aurantiaca. Acta Crystallogr. B Struct. Sci. Cryst. Eng. Mater. 1982, 38, 2513–2515. [Google Scholar] [CrossRef]
- qing Feng, D.; Qiu, Y.; Wang, W.; Wang, X.; gang Ouyang, P.; huan Ke, C. Antifouling Activities of Hymenialdisine and Debromohymenialdisine from the Sponge Axinella Sp. Int. Biodeterior. Biodegrad. 2013, 85, 359–364. [Google Scholar] [CrossRef]
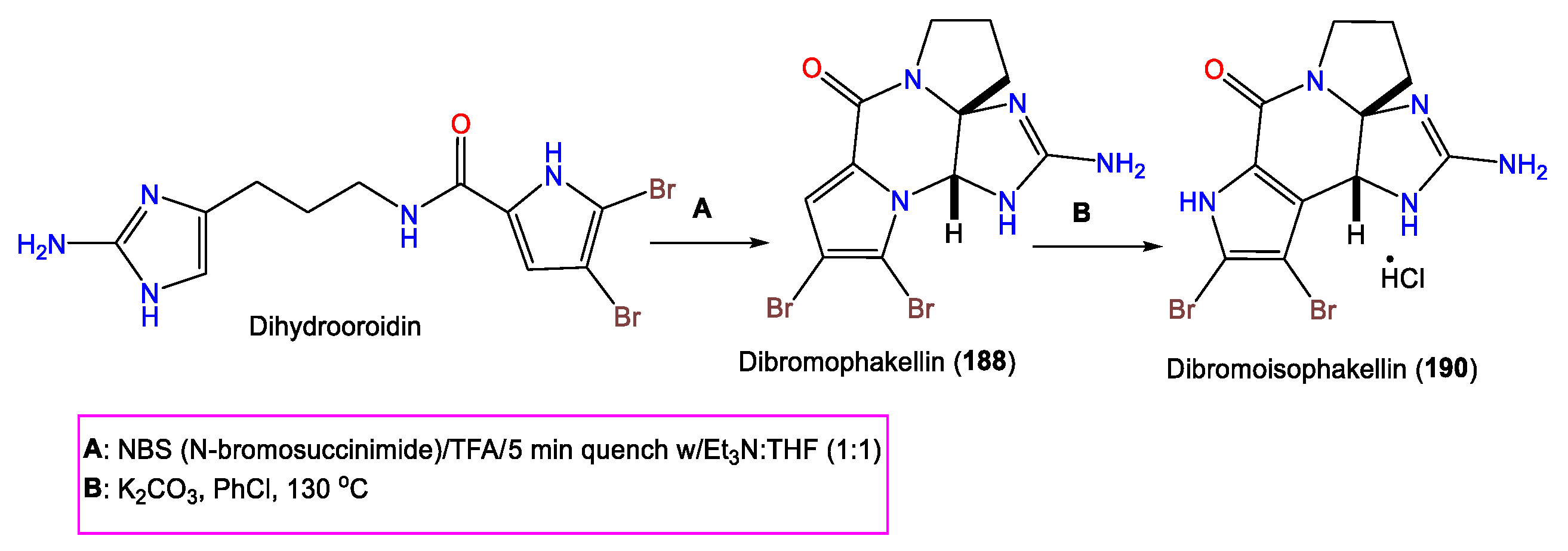
- Davis, R.A.; Fechner, G.A.; Sykes, M.; Garavelas, A.; Pass, D.M.; Carroll, A.R.; Addepalli, R.; Avery, V.M.; Hooper, J.N.; Quinn, R.J. (−)-Dibromophakellin: An α2B Adrenoceptor Agonist Isolated from the Australian Marine Sponge, Acanthella costata. Bioorg. Med. Chem. 2009, 17, 2497–2500. [Google Scholar] [CrossRef] [PubMed]
- Fedoreyev, S.A.; Utkina, N.K.; Ilyin, S.G.; Reshetnyak, M.V.; Maximov, O.B. The Structure of Dibromoisophakellin from the Marine Sponge Acanthella carteri. Tetrahedron Lett. 1986, 27, 3177–3180. [Google Scholar] [CrossRef]
- Laville, R.; Nguyen, T.B.; Moriou, C.; Petek, S.; Debitus, C.; Al-Mourabit, A. Marine Natural Occurring 2, 5-Diketopiperazines: Isolation, Synthesis and Optical Properties. Heterocycles 2015, 90, 1351–1366. [Google Scholar]
- Wiese, K.J.; Yakushijin, K.; Horne, D.A. Synthesis of Dibromophakellstatin and Dibromoisophakellin. Tetrahedron Lett. 2002, 43, 5135–5136. [Google Scholar] [CrossRef]
- Qiu, Y.; Deng, Z.W.; Xu, M.; Li, Q.; Lin, W.H. New A-nor Steroids and their Antifouling Activity from the Chinese Marine Sponge Acanthella cavernosa. Steroids 2008, 73, 1500–1504. [Google Scholar] [CrossRef]
- Tanaka Yoshito; Ito Yoshihito; Katayama Teruhisa. The Structure of Isoagelaxanthin a in Sea Sponge Acanthella vulgata. Bull. Jpn. Soc. Sci. Fish. 1982, 48, 1169–1171. [Google Scholar] [CrossRef]
- Mancini, I.; Guella, G.; Pietra, F.; Amade, P. Hanishenols A-B, Novel Linear or Methyl-Branched Glycerol Enol Ethers of the Axinellid Sponge Acanthella carteri (= Acanthella aurantiaca) from the Hanish Islands, Southern Red Sea. Tetrahedron 1997, 53, 2625–2628. [Google Scholar] [CrossRef]
- McCaffrey, E.J.; Endean, R. Antimicrobial Activity of Tropical and Subtropical Sponges. Mar. Biol. 1985, 89, 1–8. [Google Scholar] [CrossRef]
- Gab-AUa, A.A.A.; Kilada, R.W.; Shalaby, I.M.; Helmy, T. Antimicrobial Activity of some Sponges from the Gulf of Aqaba. Egypt. J. Biol. 2000, 2, 28–33. [Google Scholar]
- Xue, S.; Zhang, H.T.; Wu, P.C.; Zhang, W.; Yuan, Q. Study on Bioactivity of Extracts from Marine Sponges in Chinese Sea. J. Exp. Mar. Biol. Ecol. 2004, 298, 71–78. [Google Scholar] [CrossRef]
- Choudhury, S.; Pattnaik, P.; Sree, A.; Bapuji, M.; Mukherjee, S.C. Antibacterial Activity of Sponge Extracts Against Fish Pathogens. Aquacult. Res. 2003, 34, 1075–1077. [Google Scholar] [CrossRef]
- Rajendran, I.; Sobhana, K.S.; Annie Selva Sonia, G.; Chakraborty, K.; Vijayan, K.K.; Vijayagopal, P. Antibacterial and Antifungal Properties of Southeast Indian Coastal Sponges. J. Mar. Biol. Assoc. India 2011, 53, 272–274. [Google Scholar]
- Abdullah, N.; Al Balushi, N.; Hasan, S.I.; Al Bahlani, S.; Dobretsov, S.; Tamimi, Y.; Burney, I.A. Hymenialdisine is Cytotoxic Against Cisplatin-Sensitive but Not Against Cisplatin-Resistant Cell Lines. Sultan Qaboos Univ. Med. J. 2021, 21, 632. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Chang, C.W.; Scheuer, P.J.; Van Duyne, G.D.; Matsumoto, G.K.; Clardy, J. An Unprecedented Triisocyano Diterpenoid Antibiotic from a Sponge. J. Am. Chem. Soc. 1984, 106, 7981–7983. [Google Scholar] [CrossRef]
- Annie Selva Sonia, G.; Lipton, A.P.; Paulraj, R. Lethal Concentration of Methanol Extract of Sponges to the Brine Shrimp, Artemia salina. J. Mar. Biol. Assoc. India 2009, 51, 122–125. [Google Scholar]
- Abdillah, S.; Nurhayati, A.P.D.; Nurhatika, S.; Setiawan, E.; Heffen, W.L. Cytotoxic and Antioxidant Activities of Marine Sponge Diversity at Pecaron Bay Pasir Putih Situbondo East Java. Indonesia J. Pharm. Res. 2013, 6, 685–689. [Google Scholar] [CrossRef]
- Stanojković, T.; Milović, S.; Matić, I.; Grozdanić, N.; Kljajić, Z. In Vitro Ispitivanje Antitumorske Aktivnosti Ekstrakta Sunđera Acanthella acuta. Lek. Sirovine 2015, 89–101. [Google Scholar] [CrossRef]
- Annie Selva Sonia, G.; Lipton, A.P. Mosquito Larvicidal Activity of Marine Sponge Metabolites. Glob. J. Pharmacol. 2012, 6, 1–3. [Google Scholar]
- Putra, M.Y.; Murniasih, T.; Wibowo, J.T.; Hadi, T.A.; Untari, F.; Nisa, A.C.; Swasono, R.T. Phenolic Content, Anti-Oxidant, Anti-Plasmodium and Cytotoxic Properties of the Sponge Acanthella cavernosa. Asian Pac. J. Trop. Dis. 2016, 6, 811–815. [Google Scholar] [CrossRef]
- Rónavári, A.; Igaz, N.; Adamecz, D.I.; Szerencsés, B.; Molnar, C.; Kónya, Z.; Pfeiffer, I.; Kiricsi, M. Green Silver and Gold Nanoparticles: Biological Synthesis Approaches and Potentials for Biomedical Applications. Molecules 2021, 26, 844. [Google Scholar] [CrossRef]
- Inbakandan, D.; Sivaleela, G.; Peter, D.M.; Kiurbagaran, R.; Venkatesan, R.; Khan, S.A. Marine Sponge Extract Assisted Biosynthesis of Silver Nanoparticles. Mater. Lett. 2012, 87, 66–68. [Google Scholar] [CrossRef]
- Inbakandan, D.; Venkatesan, R.; Khan, S.A. Biosynthesis of Gold Nanoparticles Utilizing Marine Sponge Acanthella Elongata (Dendy, 1905). Colloids Surf. B Biointerfaces 2010, 81, 634–639. [Google Scholar] [CrossRef]




























| Species | Accepted as | Type Locality |
|---|---|---|
| Acanthella aculeata (Thiele, 1898) | - | Central Kuroshio Japan |
| Acanthella annulata (Sarà, 1958) | - | Western Mediterranean # |
| Acanthella acuta (Schmidt, 1862) | - | Adriatic Sea # |
| Acanthella aurantiaca (Keller, 1889) * | Stylissa carteri (Dendy, 1889) | Southern Red Sea |
| Acanthella branchia Sim (Kim and Byeon, 1990) | East China Sea South Korea | |
| Acanthella cactiformis (Carter, 1885) ** | Rhaphoxya cactiformis (Carter, 1885) | Bassian |
| Acanthella calyx (Dendy, 1922) | Cargados Carajos/Tromelin Island # | |
| Acanthella carduus (Lamarck, 1814) ** | Phakellia carduus (Lamarck, 1814) | - |
| Acanthella carteri (Dendy, 1889) | Stylissa carteri (Dendy, 1889) | South India Sri Lanka |
| Acanthella cavernosa (Dendy, 1922) | - | Seychelles # |
| Acanthella columnata (Burton, 1928) | Phakellia columnata (Burton, 1928) | - |
| Acanthella corrugata (George and Wilson, 1919) | Axinella corrugata (George and Wilson, 1919) | Virginian |
| Acanthella costata (Kieschnick, 1898) | Banda Sea Ambon | |
| Acanthella cristagalli (Dendy, 1924) ** | Tedania (Tedaniopsis) cristagalli Dendy, 1924) | - |
| Acanthella cubensis (Alcolado, 1984) | - | Greater Antilles |
| Acanthella danerii Costa (Bavestrello, Pansini, and Bertolino, 2020) | - | Chilean Exclusive Economic Zone # Chiloense |
| Acanthella dendyi (Bergquist, 1970) | - | Northeastern New Zealand |
| Acanthella ehrenbergi (Keller, 1889) ** | Biemna ehrenbergi (Keller, 1889) | Southern Red Sea |
| Acanthella elongata (Dendy, 1905) | Auletta elongata (Dendy, 1905) | - |
| Acanthella erecta (Carter, 1876) | - | Celtic Seas |
| Acanthella euctimena (Hentschel, 1912) ** | Phakettia euctimena (Hentschel, 1912) | Arafura Sea |
| Acanthella flabellata (Tanita, 1961) | - | Central Kuroshio Current |
| Acanthella flabelliformis (Keller, 1889) ** | Echinodictyum flabelliforme (Keller, 1889) | Southern Red Sea |
| Acanthella flagelliformis (van Soest and Stentoft, 1988) | Eastern Caribbean # | |
| Acanthella gorgonoides (Thomas, 1984) | - | Eastern India |
| Acanthella hirciniopsis (Carter, 1885) * | Rhaphoxya cactiformis (Carter, 1885) | Bassian |
| Acanthella hispida (Pulitzer-Finali, 1982) | Southern China # | |
| Acanthella inflexa (Pulitzer-Finali, 1982) ** | Stylissa inflexa (Pulitzer-Finali, 1982) | Central and Southern Great Barrier Reef # |
| Acanthella insignis (Thiele, 1898) | - | Central Kuroshio Current Japan |
| Acanthella klethra (Pulitzer-Finali, 1982) | - | Central and Southern Great Barrier Reef # |
| Acanthella ligulata (Burton, 1928) | - | Andaman Sea Coral Coast |
| Acanthella lyrata (Esper, 1794) ** | Auletta lyrata (Esper, 1794) | - |
| Acanthella mastophora (Schmidt, 1870) | - | Floridian # |
| Acanthella megaspicula (Thomas, 1984) | - | Eastern India |
| Acanthella minuta (Tanita, 1968) | - | Central Kuroshio Current |
| Acanthella multiformis (Vosmaer, 1885) | - | Northern Norway and Finnmark |
| Acanthella obtusa (Schmidt, 1862) ** | Dictyonella obtusa (Schmidt, 1862) | Adriatic Sea |
| Acanthella oviforma (Tanita and Hoshino, 1989) | - | Central Kuroshio Current |
| Acanthella pulcherrima (Ridley and Dendy, 1886) | - | Central Kuroshio Current |
| Acanthella ramosa (Kumar, 1925) | - | Eastern India # |
| Acanthella ramus (Sim, Kim, and Byeon, 1990) | - | East China Sea # |
| Acanthella saladinorum (Sim-Smith, Hickman Jr, and Kelly, 2021) | - | Eastern Galapagos Islands # |
| Acanthella simplex (Thiele, 1898) | - | Central Kuroshio Current |
| Acanthella stanleei (Nascimento, Cavalcanti, and Pinheiro, 2019) | - | Northeastern Brazil # |
| Acanthella stipitata (Carter, 1881) ** | Phakellia stipitata (Carter, 1881) | Bassian |
| Acanthella styloida (Tanita and Hoshino, 1989) | - | Central Kuroshio Current |
| Acanthella tenuispiculata (Dendy, 1897) | - | Great Australian Bight, Australia |
| Acanthella vaceleti (van Soest and Stentoft, 1988) | - | Eastern Caribbean # Barbados Exclusive Economic Zone |
| Acanthella vulgata (Thiele, 1898) | - | Central Kuroshio Current |
| Acanthella xutha (de Laubenfels, 1954) | - | East Caroline Islands |
| Compound Name | Mol. Wt. | Mol. Formula | Species | Sampling Locations | Ref. |
|---|---|---|---|---|---|
| Aromadendrane-type sesquiterpenes | |||||
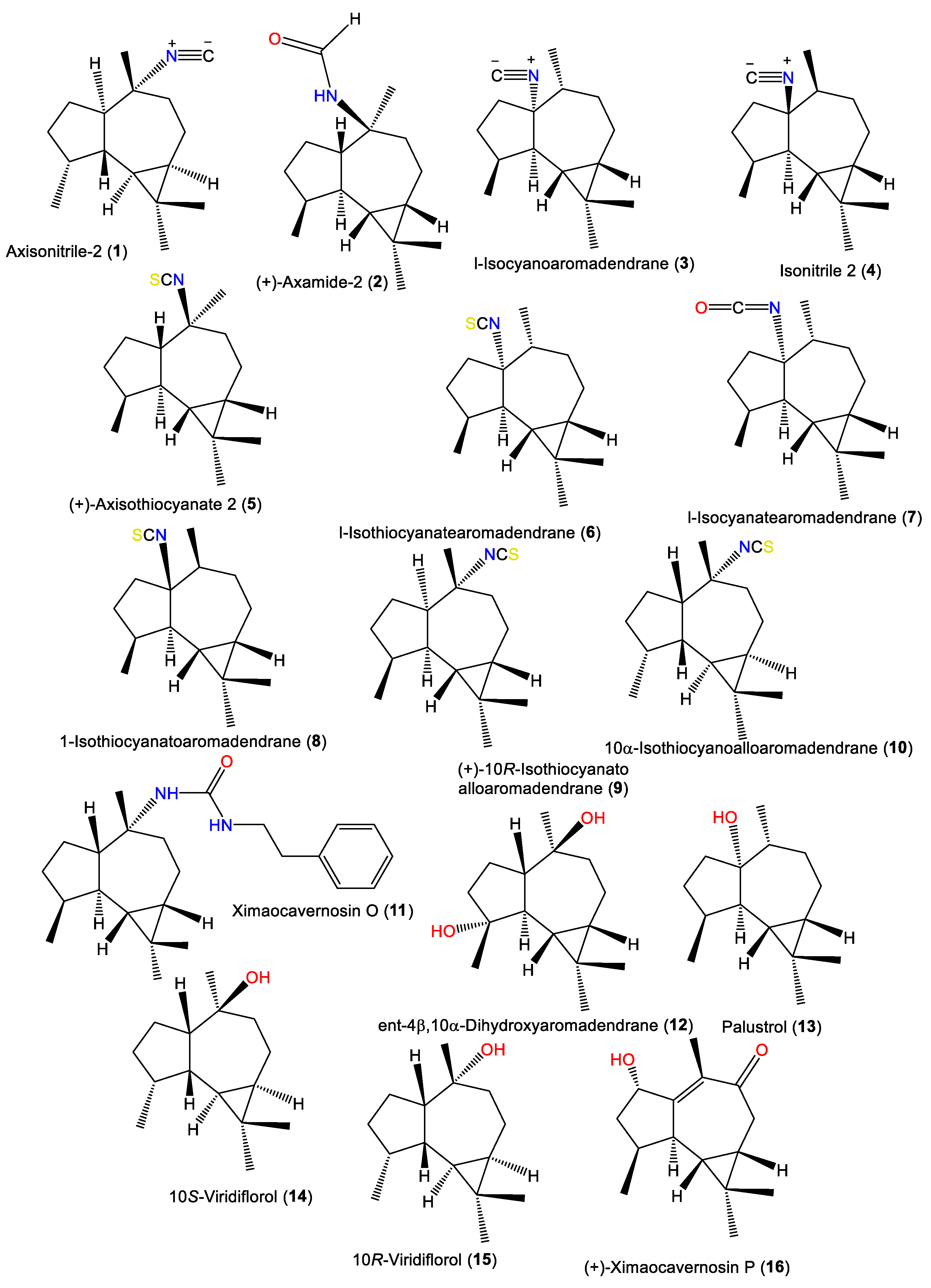
| Axisonitrile-2 (1) | 231 | C16H25N | A. cavernosa | Hachijo-Jima Island, Japan | [32] |
| (+)-Axamide 2 (2) | 249 | C16H27NO | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| l-Isocyanoaromadendrane (3) | 231 | C16H25N | A. acuta | Near Banyuls, France | [34,35] |
| - | - | A. pulcherrima | Weed Reef, Darwin, Australia | [35] | |
| - | - | A. acuta | Near Banyuls, France | [36] | |
| - | - | A. cavernosa | Tani’s Reef, Mooloolaba, Australia | [37] | |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| Isonitrile 2 (4) | 231 | C16H25N | A. acuta | Bay of Naples, southern Italy | [39] |
| (+)-Axisothiocyanate 2 (5) | 263 | C16H25NS | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| l-Isothiocyanatearomadendrane (6) | 263 | C16H25NS | A. acuta | Near Banyuls, France | [36] |
| - | - | A. cavernosa | Tani’s Reef, Mooloolaba, Australia | [37] | |
| 1-Isocyanatearomadendrane (7) | 247 | C16H25NO | A. acuta | Near Banyuls, France | [36] |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| 1-Isothiocyanatoaromadendrane (8) | 263 | C16H25NS | A. acuta | Bay of Naples, southern Italy | [39] |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| (+)-10R-Isothiocyanatoalloaromadendrane (9) | 263 | C16H25NS | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | Acanthella sp. | Yalong Bay, Hainan, China | [42] | |
| - | - | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] | |
| 10α-Isothiocyanoalloaromadendrane (10) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | A. cavernosa | Tani’s Reef, Mooloolaba, Australia | [37] | |
| Ximaocavernosin O (11) | 368 | C24H36N2O | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| ent-4β,10α-Dihydroxyaromadendrane (12) | 238 | C15H26O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| Palustrol (13) | 222 | C15H26O | A. acuta | Near Banyuls, France | [34] |
| - | - | A. acuta | Near Banyuls, France | [36] | |
| 10S-Viridiflorol (14) | 222 | C15H26O | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| 10R-Viridiflorol (15) | 222 | C15H26O | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| (+)-Ximaocavernosin P (16) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| Spiroaxane-type sesquiterpenes | |||||
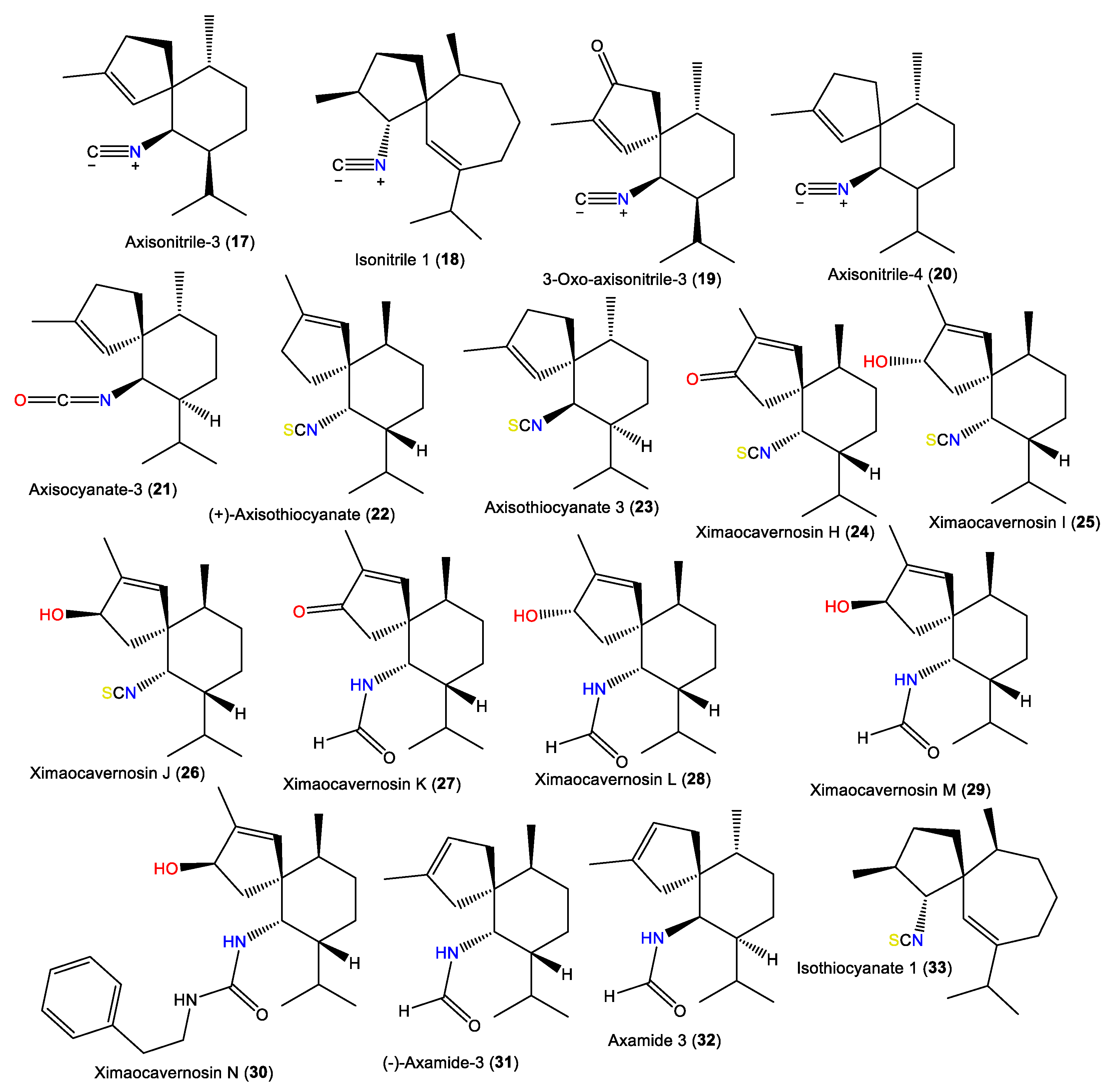
| Axisonitrile-3 (17) | 231 | C16H25N | A. acuta | Near Banyuls, France | [34] |
| - | - | A. cavernosa | Thailand | [18] | |
| - | - | A. klethra | Pelorus Island, Queensland, Australia | [19] | |
| - | - | A. klethra | Vicinities of Phantom and Pelom Islands, Queensland, Australia | [44] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [31] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | A. cavernosa | Tani’s Reef, Mooloolaba, Australia | [37] | |
| Isonitrile 1 (18) | 245 | C17H27N | A. acuta | Bay of Naples, southern Italy | [39] |
| 3-Oxoaxisonitrile-3 (19) | 245 | C16H23NO | Acanthella sp. | Ximao Sea, Hainan, China | [41] |
| Axisonitrile-4 (20) | 231 | C16H25N | A. acuta | Sidi Elghdamssi island, Monastir region, Tunisia | [25] |
| Axisocyanate-3 (21) | 247 | C16H25NO | A. cavernosa | Tani’s Reef, Mooloolaba, Australia | [37] |
| (+)-Axisothiocyanate (22) | 263 | C16H25NS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Axisothiocyanate 3 (23) | 263 | C16H25NS | A. klethra | Pelorus Island, Queensland, Australia | [19] |
| - | - | A. klethra | Vicinities of Phantom and Pelom Islands, Queensland, Australia | [44] | |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| - | - | A. cavernosa | Hachijo-jima Island, Japan | [40] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [31] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| Ximaocavernosin H (24) | 277 | C16H22NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin I (25) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin J (26) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin K (27) | 263 | C16H25NO2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin L (28) | 265 | C16H27NO2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin M (29) | 265 | C16H27NO2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Ximaocavernosin N (30) | 368 | C24H36N2O | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (–)-Axamide 3 (31) | 249 | C16H27NO | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Axamide 3 (32) | 249 | C16H27NO | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| Isothiocyanate 1 (33) | 277 | C17H27NS | A. acuta | Bay of Naples, southern Italy | [39] |
| Eudesmane-type sesquiterpenes | |||||
| Acanthellin-1 (34) | 231 | C16H25N | A. acuta | Bay of Naples, southern Italy | [30] |
| - | - | A. acuta | Near Banyuls, France | [34] | |
| - | - | A. acuta | Bay of Taranto near Porto Cesareo, Southern Italy | [47] | |
| - | - | A. acuta | Sidi Elghdamssi Island, Monastir region, Tunisia | [25] | |
| Acanthene B (35) | 263 | C16H25NS | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| Acanthine B (36) | 263 | C16H25NS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Acanthene C (37) | 249 | C16H27NO | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| Axiriabiline A (38) | 249 | C16H27NO | A. cavernosa | Xidao Island, Hainan, China | [48] |
| 6α-Isocyano-5αH,7αH,10α-eudesm-4(14)-ene) (39) | 231 | C16H25N | A. acuta | Bay of Taranto near Porto Cesareo, Southern Italy | [47] |
| - | - | A. acuta | Bay of Naples, southern Italy | [39] | |
| 11-Isocyano-7βH-eudesm-5-ene (40) | 231 | C16H25N | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| 11-Formamido-7βH-eudesm-5-ene (41) | 249 | C16H27NO | A. cavernosa | Xidao Island, Hainan, China | [48] |
| 11-Isothiocyano-7βH-eudesm-5-ene (42) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| - | - | A. klethra | Pelorus Island, Queensland, Australia | [19] | |
| - | - | A. klethra | Vicinities of Phantom and Pelom Islands, Queensland, Australia | [44] | |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [31] | |
| 6α-Isothiocyano-5αH,7αH,10α-eudesm-4(14)-ene (43) | 263 | C16H25NS | A. acuta | Bay of Taranto near Porto Cesareo, Southern Italy | [47] |
| - | - | A. acuta | Bay of Naples, southern Italy | [39] | |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| (lR,5R,6R,8R)-Dec[4.4.0]ane-1,5-dimethyl-8-(1′-methylethenyl)-5-isothiocyanate (44) | 263 | C16H25NS | A. klethra | Pelorus Island, Queensland, Australia | [19] |
| - | - | A. klethra | Vicinities of Phantom and Pelom Islands, Queensland, Australia | [44] | |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| (lR,5R,6R,8S)-Dec[4.4.0]ane-1,5-dimethyl-8-(1′-methylethenyl)-5-isothiocyanate (45) | 263 | C16H25NS | A. klethra | Pelorus Island, Queensland, Australia | [19] |
| - | - | A. klethra | Vicinities of Phantom and Pelom Islands, Queensland, Australia | [44] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| Cadinene-type sesquiterpenes | |||||
| 10-Isothiocyanato-4-cadinene (46) | 263 | C16H25NS | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| 10-Isothiocyanato-4-amorphene (47) | 263 | C16H25NS | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| - | - | A. cavernosa | Several locations off the Japanese coast | [49] | |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| Isomer-10-isothiocyanato-4-amorphene (48) | 263 | C16H25NS | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| (+)-Ximaocavernosin A (49) | 295 | C16H25NO2S | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| (±)-Ximaocavernosin A (50) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (±)-Ximaocavernosin B (51) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (±)-Ximaocavernosin C (52) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (+)-Ximaocavernosin D (53) | 293 | C17H27NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (±)-Ximaocavernosin E (54) | 295 | C16H25NO2S | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (+)-Ximaocavernosin F (55) | 309 | C17H27NO2S | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (±)-Ximaocavernosin G (56) | 277 | C16H23NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| (±)-Axinisothiocyanate J (57) | 279 | C16H25NOS | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
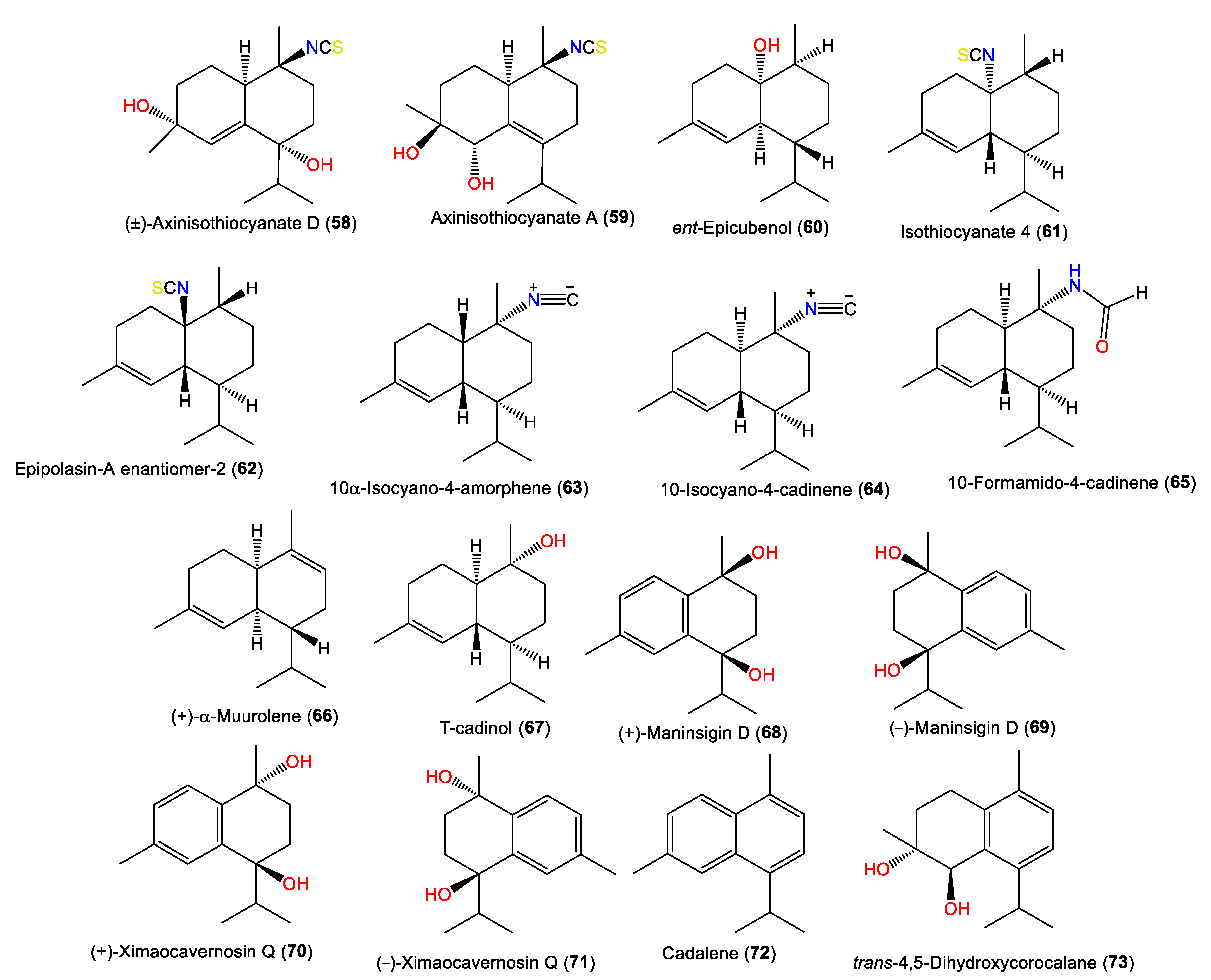
| (±)-Axinisothiocyanate D (58) | 295 | C16H25NO2S | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| Axinisothiocyanate A (59) | 295 | C16H25NO2S | A. cavernosa | Coast of Ximao Island, Hainan, China | [33] |
| ent-Epicubenol (60) | 222 | C15H26O | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| Isothiocyanate 4 (61) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| Epipolasin-A enantiomer-2 (62) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| 10α-Isocyano-4-amorphene (63) | 231 | C16H25N | A. cavernosa | Hachijo-Jima Island, Japan | [32] |
| - | - | A. cavernosa | Several locations off the Japanese coast | [49] | |
| 10-Isocyano-4-cadinene (64) | 231 | C16H25N | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| - | - | A. cavernosa | Several locations off the Japanese coast | [49] | |
| - | - | A. cavernosa | Tani’s Reef or Coral Gardens dive sites | [38] | |
| 10-Formamido-4-cadinene (65) | 249 | C16H27NO | A. cavernosa | Several locations off the Japanese coast | [49] |
| - | - | A. cavernosa | Xidao Island, Hainan, China | [48] | |
| (+)-α-Muurolene (66) | 204 | C15H24 | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| T-cadinol (67) | 222 | C15H26O | A. cavernosa | Several locations off the Japanese coast | [49] |
| (+)-Maninsigin D (68) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] | |
| (-)-Maninsigin D (69) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| (+)-Ximaocavernosin Q (70) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| (-)-Ximaocavernosin Q (71) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| Cadalene (72) | 198 | C15H18 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| trans-4,5-Dihydroxycorocalane (73) | 234 | C15H22O2 | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| Axane-type sesquiterpenes | |||||
| Cavernoisonitrile (74) | 245 | C16H23NO | A. cavernosa | Hachijo-Jima Island, Japan | [32] |
| (-)-Cavernothiocyanate (75) | 263 | C16H25NS | A. cavernosa | Hachijo-Jima Island, Japan | [32] |
| - | - | A. cavernosa | Hachijo-jima Island, Japan | [40] | |
| 1β-H,7α-methyl,8α-H,9β-methyl-Cavernothiocyanate (76) | 263 | C16H25NS | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| Bisabolene-type sesquiterpenes | |||||
| 7-Isocyano-7,8-dihydro-α-bisabolene (77) | 231 | C16H25N | A. cavernosa | Hachijo-Jima Island, Japan | [32] |
| Epimaaliane-type sesquiterpenes | |||||
| (+)-Epipolasin-A (78) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [31] | |
| Epipolasin-A enantiomer-1 (79) | 263 | C16H25NS | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| 5-Formamide-isonitrile (80) | 249 | C16H27NO | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| Isonitrile 4 (81) | 231 | C16H25N | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| Maaliol (82) | 222 | C15H26O | A. pulcherrima | Weed Reef, Darwin, Australia | [35] |
| Acanthene A (83) | 240 | C15H25Cl | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] |
| Gurjunene | |||||
| (+)-Aristolone (84) | 218 | C15H22O | A. cavernosa | Coast of Ximao Island, Hainan, China | [43] |
| - | - | A. cavernosa | South China Sea | [50] | |
| (+)-9-Aristolene (85) | 204 | C15H24 | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| Isonitrile 3 (86) | 231 | C16H25N | A. acuta | Bay of Naples, southern Italy | [39] |
| Isothiocyanate 3 (87) | 263 | C16H25NS | A. acuta | Bay of Naples, southern Italy | [39] |
| Compound Name | Mol. Wt. | Mol. Formula | Species | Sampling Locations | Ref. |
|---|---|---|---|---|---|
| Kalihinols | |||||
| Kalihinol A (88) | 392 | C22H33ClN2O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [52] |
| - | - | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] | |
| - | - | A. caruenosa | Fiji, South Pacific Ocean | [53] | |
| - | - | A. klethra | Kuchinoerabu Island of the Satsunan Archipelago, Japan | [23] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [51] | |
| - | - | A. cavernosa | Yakushima Island, southwest of Tokyo | [55,56] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japa | [57] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| - | - | A. cavernosa | Shallow water reef in Sanya Bay, Hainan Island, China | [58] | |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| - | - | Acanthella sp. | Yalong Bay, Hainan, China | [42] | |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| - | - | A. cavernosa | South China Sea | [50] | |
| trans 10β-Formamidokalihinol A (89) | 410 | C22H35ClN2O3 | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| - | - | A. cavernosa | Shallow water reef in Sanya Bay, Hainan Island, China | [58] | |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| cis 10β-Formamidokalihinol A (90) | 410 | C22H35ClN2O3 | A. cavernosa | Shallow water reef in Sanya Bay, Hainan Island, China | [58] |
| 10β-Formamido-5-isocyanatokalihinol A (91) | 426 | C22H35ClN2O4 | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| - | - | A. cavernosa | Xidao Island, Hainan, China | [48] | |
| 10β-Formamido-5β-isothiocyanatokalihinol A (92) | 442 | C22H35ClN2O3S | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| - | - | A. cavernosa | Xidao Island, Hainan, China | [48] | |
| Bisformamidokalihinol A (93) | 428 | C22H37ClN2O4 | A. cavernosa | Xidao Island, Hainan, China | [48] |
| Kalihinol B (94) | 392 | C22H33ClN2O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| - | - | A. klethra | Kuchinoerabu Island of the Satsunan Archipelago, Japan | [23] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| Isokalihinol B (95) | 392 | C22H33ClN2O2 | A. klethra | Kuchinoerabu Island of the Satsunan Archipelago, Japan | [23] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] | |
| Kalihinol C (96) | 328 | C20H28N2O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| Kalihinol D (97) | 392 | C22H33ClN2O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | Acanthella sp. | Yalong Bay, Hainan, China | [42] | |
| Kalihinol E (98) | 392 | C22H35ClN2O3 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Hachijo-jima Island, Japan | [40] | |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| - | - | A. cavernosa | South China Sea | [50] | |
| 10β-Formamidokalihinol E (99) | 410 | C22H33ClN2O2 | A. cavernosa | Hachijo-jima Island, Japan | [40] |
| Kalihinol F (100) | 383 | C23H33N3O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. caruenosa | Fiji, South Pacific Ocean | [53] | |
| - | - | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [60] | |
| - | - | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia. | [46] | |
| - | - | A. klethra | Kuchinoerabu Island of the Satsunan Archipelago, Japan | [23] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] | |
| - | - | Acanthella sp. | Coast of Cape Sada, Ehime Prefecture, Japan | [61] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| Isokalihinol F (101) | 383 | C23H33N3O2 | A. cavernosa | Fiji, South Pacific Ocean | [53] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] | |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [51] | |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| 8-Hydroxy-isokalihinol F (102) | 399 | C23H33N3O3 | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| 10-epi-Isokalihinol F (103) | 383 | C23H33N3O2 | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| trans 10-Formamido-kalihinol F (104) | 401 | C23H35N3O3 | A. cavernosa | Dibud, Philippines | [21] |
| cis 10-Formamido-kalihinol F (105) | 401 | C23H35N3O3 | A. cavernosa | Dibud, Philippines | [21] |
| trans 15-Formamido-kalihinol F (106) | 401 | C23H35N3O3 | A. cavernosa | Dibud, Philippines | [21] |
| cis 15-Formamido-kalihinol F (107) | 401 | C23H35N3O3 | A. cavernosa | Dibud, Philippines | [21] |
| Kalihinol G (108) | 415 | C23H33N3O2S | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| 10-Isothiocyanatokalihinol G (109) | 447 | C23H33N3O2S2 | A. cavernosa | Xisha Islets, South China Sea | [16] |
| 5,10-bis-Isothiocyanatokalihinol G (110) | 479 | C23H33N3O2S3 | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] |
| Kalihinol H (111) | 415 | C23H33N3O2S | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| Isokalihinol H (112) | 415 | C23H33N3O2S | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| 10-epi-Isokalihinol H (113) | 415 | C23H33N3O2S | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| Kalihinol I (114) | 456 | C22H33ClN2O2S2 | A. cavernosa | Thailand | [18] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| 10-epi-Kalihinol I (115) | 456 | C22H33ClN2O2S2 | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| - | - | A. cavernosa | South China Sea | [50] | |
| Kalihinol J (116) | 442 | C22H35ClN2O3S | A. cavernosa | Thailand | [18] |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| Kalihinol M (117) | 442 | C22H35ClN2O3S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol N (118) | 442 | C22H35ClN2O3S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol O (119) | 424 | C22H33ClN2O2S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol P (120) | 424 | C22H33ClN2O2S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol Q (121) | 424 | C22H33ClN2O2S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol R (122) | 456 | C22H33ClN2O2S2 | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol S (123) | 410 | C22H35ClN2O3 | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol T (124) | 424 | C22H33ClN2O2S | A. cavernosa | Xisha Islets, South China Sea | [16] |
| Kalihinol X (125) | 424 | C22H33ClN2O2S | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [53] | |
| - | - | A. cavernosa | Thailand | [18] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| - | - | Acanthella sp. | Yalong Bay, Hainan, China | [42] | |
| 10-epi-Kalihinol X (126) | 424 | C22H33ClN2O2S | Acanthella sp. | Yalong Bay, Hainan, China | [42] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [16] | |
| - | - | A. cavernosa | South China Sea | [50] | |
| Kalihinol Y (127) | 365 | C21H32ClNO2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | Acanthella caruenosa | Fiji, South Pacific Ocean | [53] | |
| - | - | A. cavernosa | Thailand | [18] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| Kalihinone Ya (128) | 367 | C20H30ClNO3 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| Δ9-Kalihinol Y (129) | 365 | C21H32ClNO2 | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] |
| Kalihinol Z (130) | 392 | C22H33ClN2O2 | Acanthella sp. | Apra Harbor, Guam, western side of the USA | [22] |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [53] | |
| - | - | A. cavernosa | Seychelles and Desnceufs Islands | [54] | |
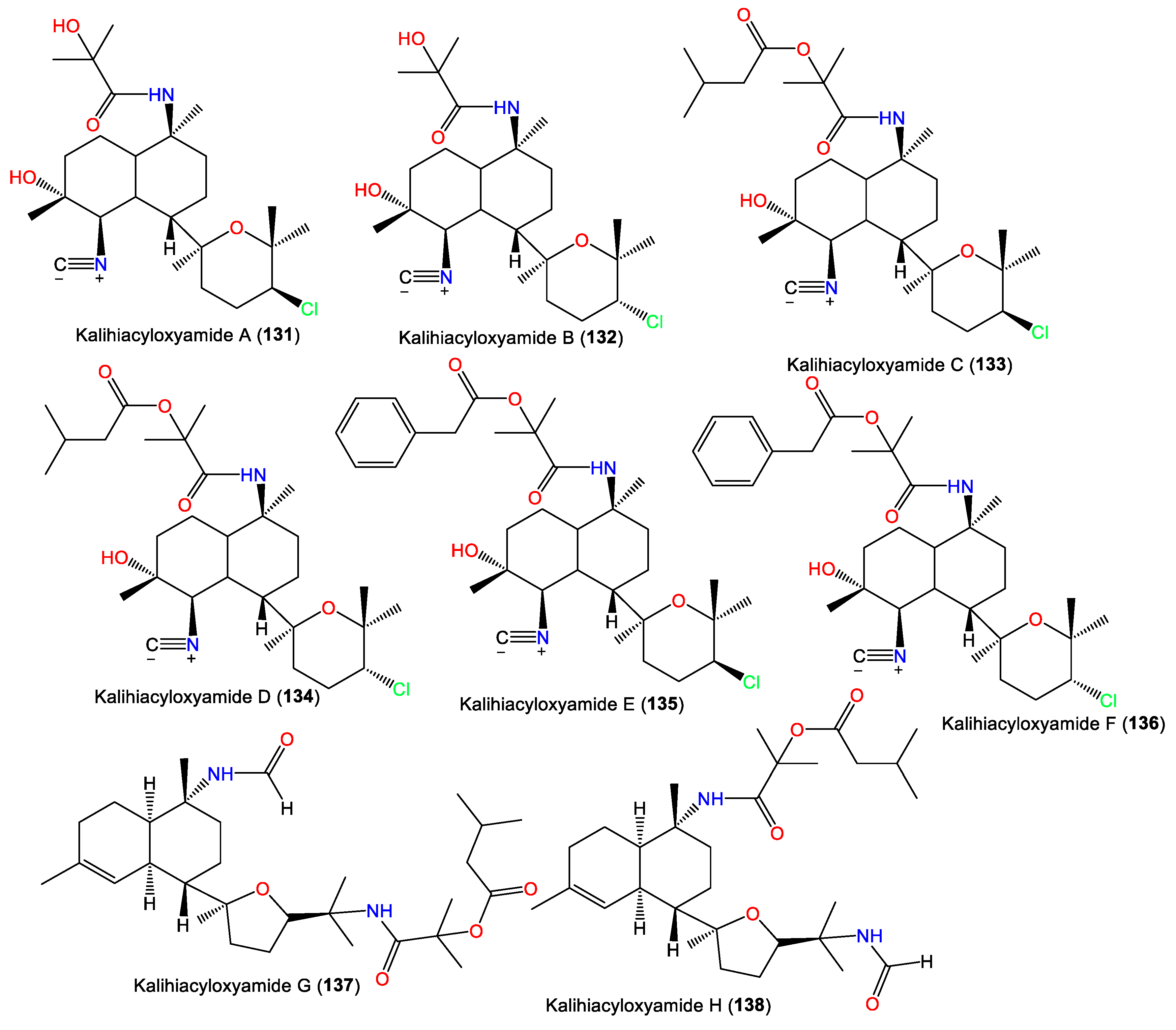
| Kalihiacyloxyamide A (131) | 468 | C25H41ClN2O4 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide B (132) | 468 | C25H41ClN2O4 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide C (133) | 552 | C30H49ClN2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide D (134) | 552 | C30H49ClN2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide E (135) | 586 | C33H47ClN2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide F (136) | 586 | C33H47ClN2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide G (137) | 518 | C30H50N2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihiacyloxyamide H (138) | 518 | C30H50N2O5 | A. cavernosa | Xisha Island, South China Sea | [62] |
| Kalihinene | |||||
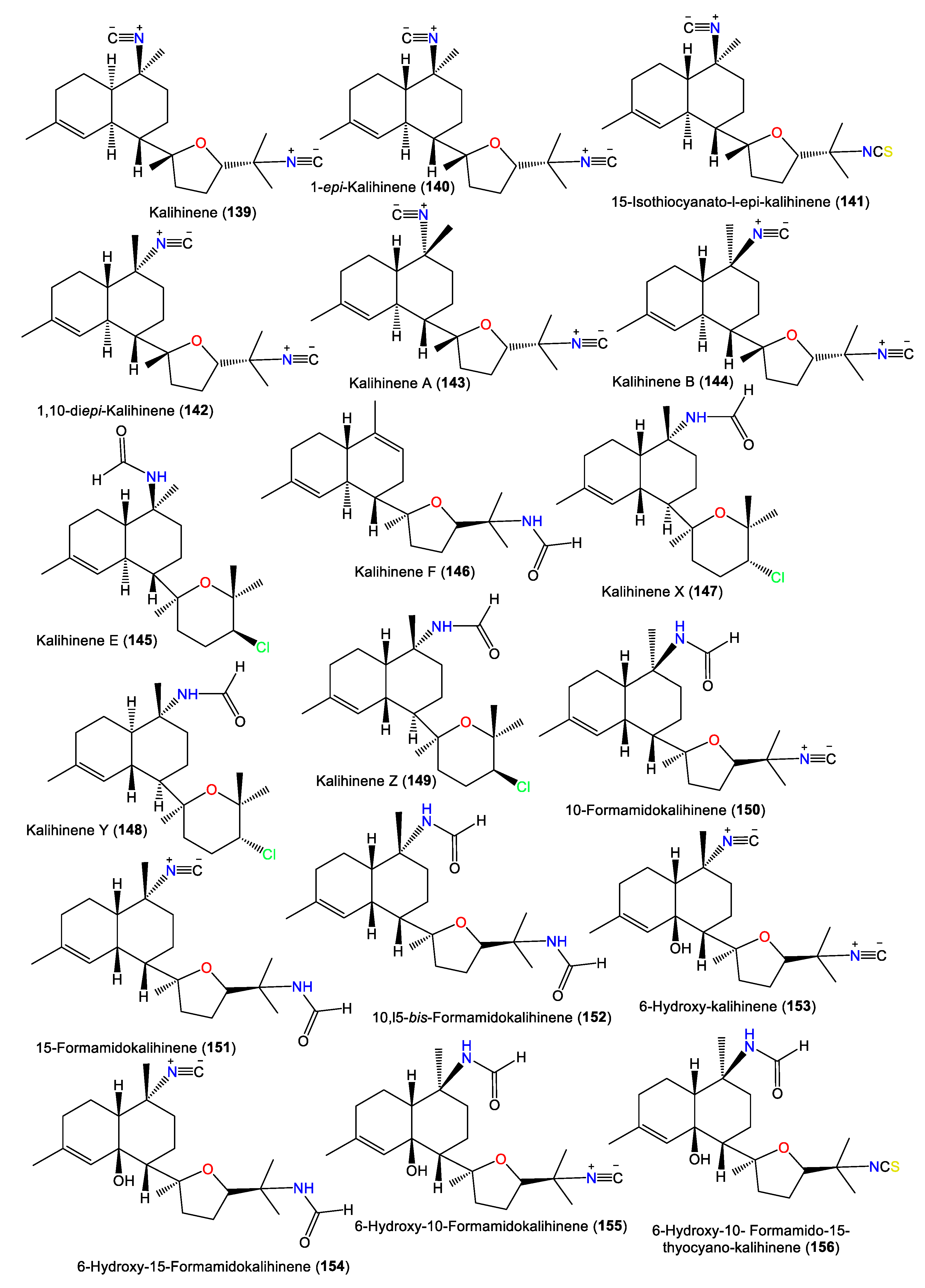
| Kalihinene (139) | 340 | C22H32N2O | A. klethra | Kuchinoerabu Island of the Satsunan Archipelago, Japan | [23] |
| - | - | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] | |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [51] | |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| - | - | A. cavernosa | Dibud, Philippines | [21] | |
| - | - | Acanthella sp. | Ximao Sea, Hainan, China | [41] | |
| 1-epi-Kalihinene (140) | 340 | C22H32N2O | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| 15-Isothiocyanato-l-epi-kalihinene (141) | 372 | C22H32N2OS | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| 1,10-diepi-Kalihinene (142) | 340 | C22H32N2O | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| Kalihinene A (143) | 340 | C22H32N2O | A. cavernosa | Seychelles and Desnceufs Islands | [54] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [63] | |
| Kalihinene B (144) | 340 | C22H32N2O | A. cavernosa | Seychelles and Desnceufs Islands | [54] |
| Kalihinene E (145) | 367 | C21H34ClNO2 | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Kalihinene F (146) | 331 | C21H33NO2 | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Kalihinene X (147) | 367 | C21H34ClNO2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55,56] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [63] | |
| Kalihinene Y (148) | 367 | C21H34ClNO2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55,56] |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [63] | |
| Kalihinene Z (149) | 367 | C21H34ClNO2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55,56] |
| 10-Formamidokalihinene (150) | 358 | C22H34N2O2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55,56] |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [51] | |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [63] | |
| 15-Formamidokalihinene (151) | 358 | C22H34N2O2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55] |
| - | - | A. cavernosa | Fiji, South Pacific Ocean | [51] | |
| - | - | A. cavernosa | Xisha Islets, South China Sea | [63] | |
| 10,l5-bis-Formamidokalihinene (152) | 376 | C22H36N2O3 | A. cavernosa | Fiji, South Pacific Ocean | [51] |
| 6-Hydroxy-kalihinene (153) | 356 | C22H32N2O2 | A. cavernosa | Fiji, South Pacific Ocean | [51] |
| - | - | Acanthella sp. | Coral reef of Ishigaki Island, Okinawa, Japan | [57] | |
| 6-Hydroxy-15-Formamidokalihinene (154) | 374 | C22H34N2O3 | A. cavernosa | Fiji, South Pacific Ocean | [51] |
| 6-Hydroxy-10-Formamidokalihinene (155) | 374 | C22H34N2O3 | A. cavernosa | Fiji, South Pacific Ocean | [51] |
| 6-Hydroxy-10-Formamido-15-thyocyano-kalihinene (156) | 406 | C22H34N2O3S | A. cavernosa | Fiji, South Pacific Ocean | [51] |
| Kalihioxepanes | |||||
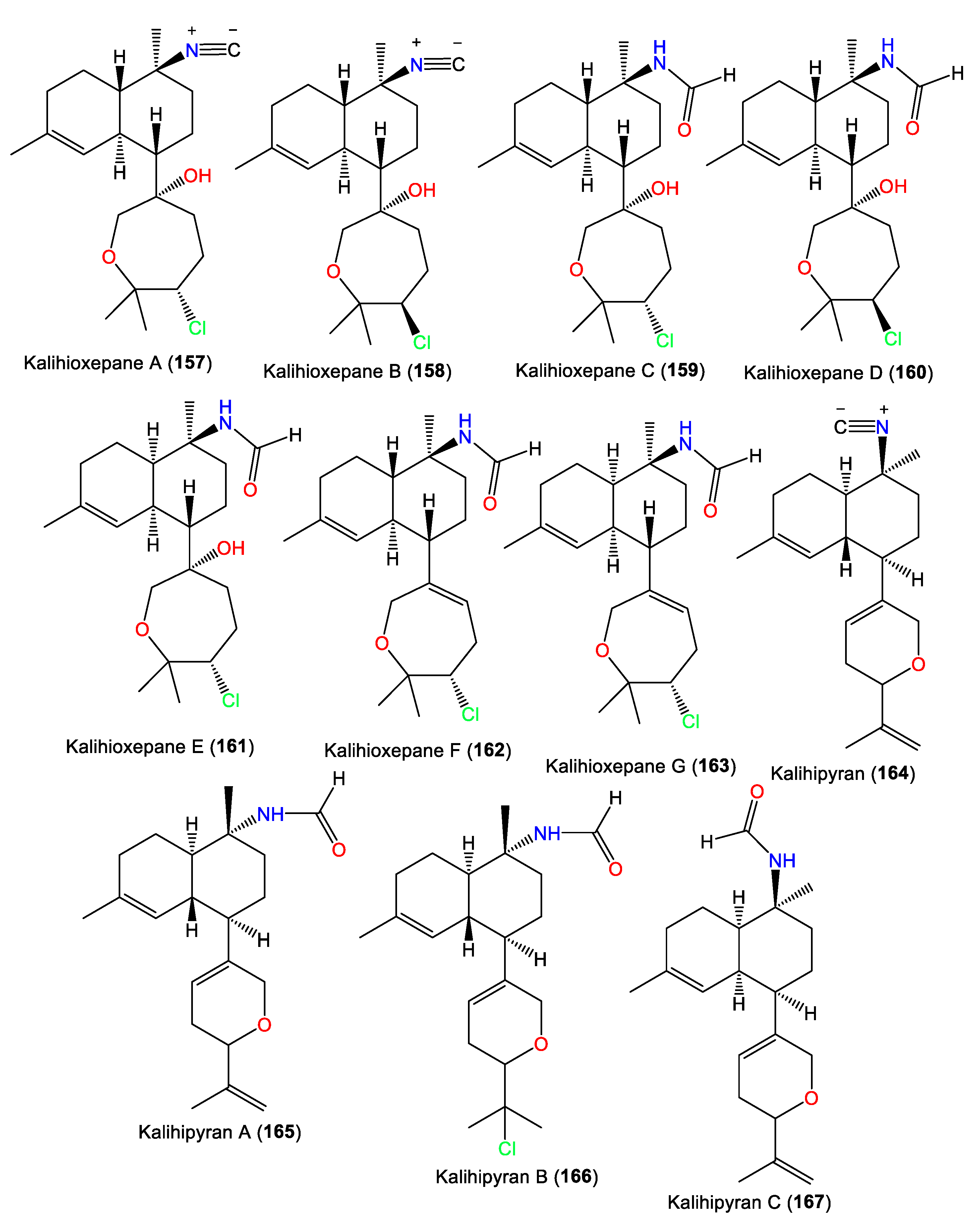
| Kalihioxepane A (157) | 365 | C21H32ClNO2 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane B (158) | 365 | C21H32ClNO2 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane C (159) | 383 | C21H34ClNO3 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane D (160) | 383 | C21H34ClNO3 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane E (161) | 383 | C21H34ClNO3 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane F (162) | 365 | C21H32ClNO2 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihioxepane G (163) | 365 | C21H32ClNO2 | A. cavernosa | Xisha Island, South China Sea | [64] |
| Kalihipyran (164) | 311 | C21H29NO | A. cavernosa | Beau Vallon Beach, Mahé, Seychelles | [59] |
| - | - | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] | |
| Kalihipyrans | |||||
| Kalihipyran A (165) | 329 | C21H31NO2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55] |
| Kalihipyran B (166) | 365 | C21H32ClNO2 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55] |
| Kalihipyran C (167) | 329 | C21H31NO2 | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Biflorane diterpenes | |||||
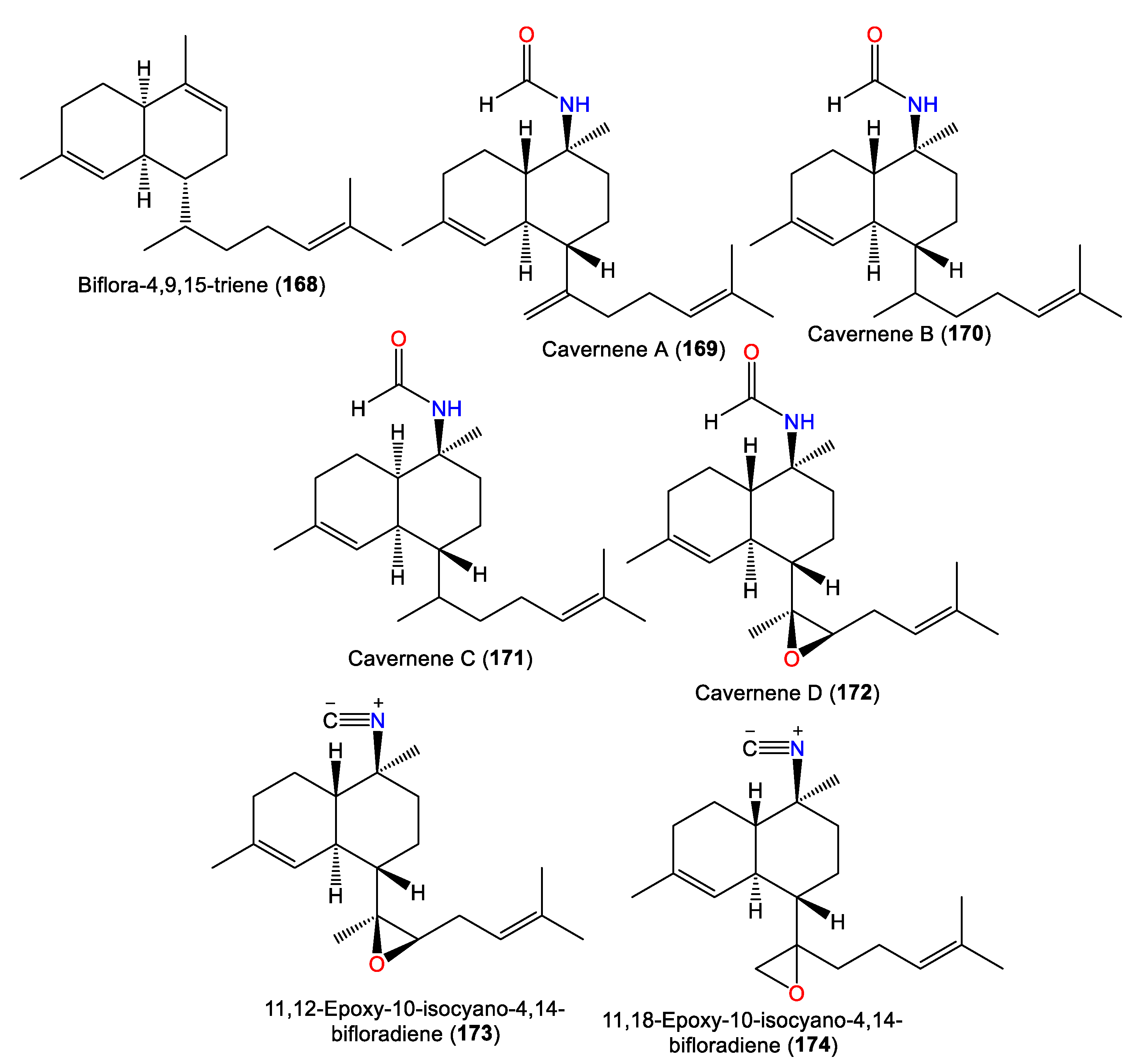
| Biflora-4,9,15-triene (168) | 272 | C20H32 | A. cavernosa | Yakushima Island, southwest of Tokyo | [55] |
| - | - | A. cavernosa | Hachijo-jima Island, Japan | [40] | |
| Cavernene A (169) | 315 | C21H33NO | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Cavernene B (170) | 317 | C21H35NO | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Cavernene C (171) | 317 | C21H35NO | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Cavernene D (172) | 331 | C21H33NO2 | A. cavernosa | Xisha Islets, South China Sea | [63] |
| Isocyanobifloradiene epoxide A (173) 11,12-epoxy-10-isocyano-4,14-bifloradiene (173) | 313 | C21H31NO | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| Isocyanobifloradiene epoxide B (174) 11,18-epoxy-10-isocyano-4,14-bifloradiene (174) | 313 | C21H31NO | A. cavernosa | Heron Island, Great Barrier Reef, Australia | [45] |
| Compound Name | Mol. Wt. | Mol. Formula | Species | Sampling Locations | Ref. |
|---|---|---|---|---|---|
| 4-Bromo-1H-pyrrole-2-carboxamide (175) | 187 | C5H5BrN2O | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [66] |
| 4,5-Dibromo-1H-pyrrole-2-carboxamide (176) | 265 | C5H4Br2N2O | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [66] |
| Oroidin (177) | 386 | C11H11Br2N5O | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [66] |
| - | - | A. acuta | Sidi Elghdamssi island, Monastir region, Tunisia | [25] | |
| Axinellizine (178) | 408 | C11H10Br2N5NaO | A. acuta | Sidi Elghdamssi island, Monastir region, Tunisia | [25] |
| Hanishin (179) | 377 | C11H12Br2N2O2 | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [66] |
| 4-(2-Amino-4-oxo-2-imidazolin-5-ylidene)-2-bromo-4,5,6,7-tetrahydropyrrolo [2,3-c] azepin-8-one (180) | 323 | C11H10BrN5O2 | A. aurantiaca | Bay of Naples, southern Italy | [67,68] |
| 4-(2-Amino-4-oxo-2-imidazolin-5-ylidene)-4,5,6,7-tetrahydropyrrolo [2,3-c] azepin-8-one (181) | 281 | C11H12ClN5O2 | A. aurantiaca | Bay of Naples, southern Italy | [67] |
| Spongiacidin D (182) | 323 | C11H9BrN4O3 | A. carteri | Dalahican Sea, near Ilaya, Lucena, Quezon, Philippines | [17] |
| 3-Bromohymenialdisin (183) | 402 | C11H11Br2N5O2 | A. carteri | Dalahican Sea, near Ilaya, Lucena, Quezon, Philippines | [17] |
| Dihydrospongiacidine (184) | 404 | C11H13Br2N5O2 | A. carteri | Dalahican Sea, near Ilaya, Lucena, Quezon, Philippines | [17] |
| Hymenialdisine (185) | 323 | C11H10BrN5O2 | Acanthella sp. | Yongxing Island, South China Sea | [69] |
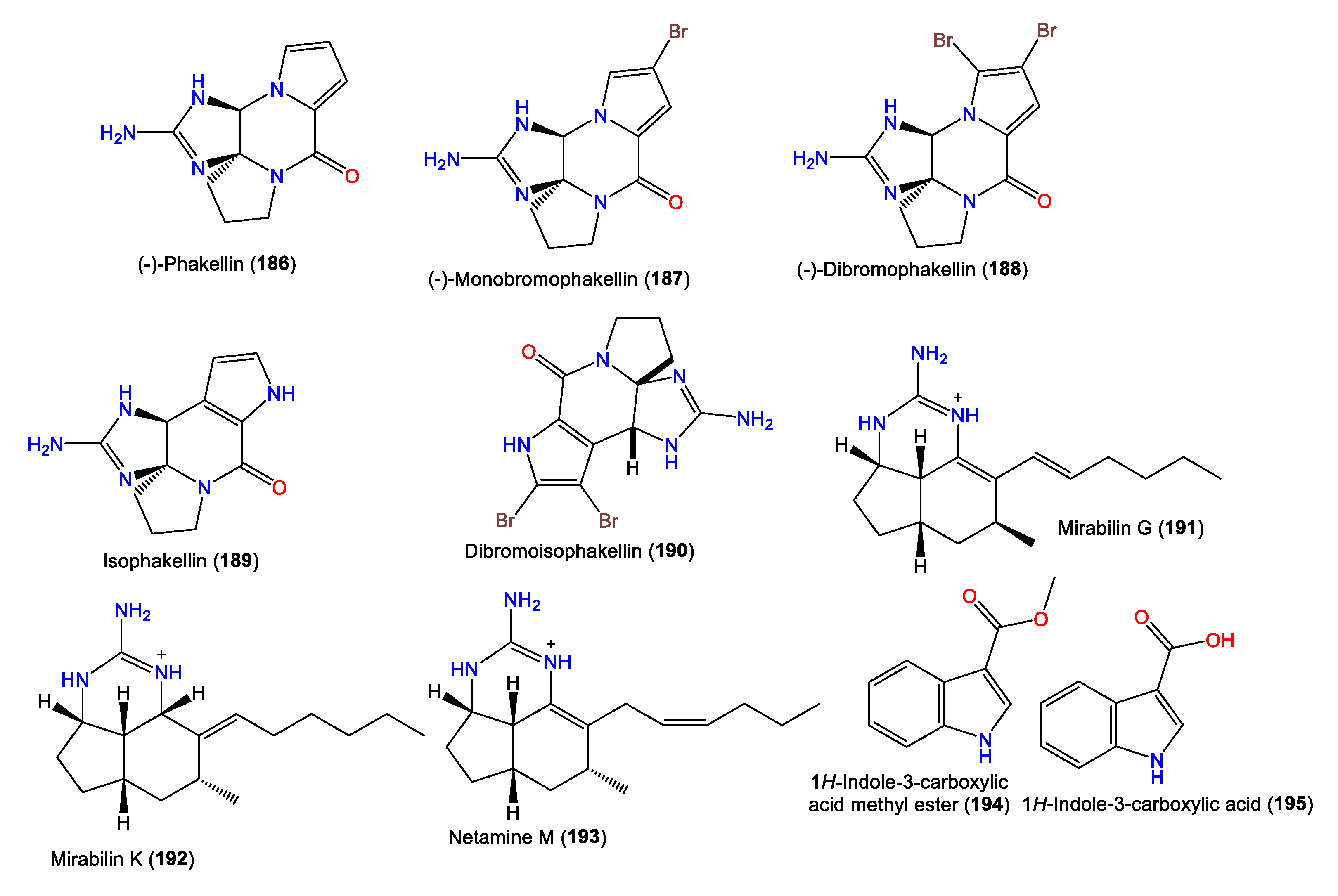
| (-)-Phakellin (186) | 231 | C11H13N5O | A. costata | Sykes Reef, Capricorn-Bunker Group Great Barrier Reef, Queensland, Australia | [70] |
| (-)-Monobromophakellin (187) | 309 | C11H12BrN5O | A. costata | Sykes Reef, Capricorn-Bunker Group Great Barrier Reef, Queensland, Australia | [70] |
| (-)-Dibromophakellin (188) | 386 | C11H11Br2N5O | A. costata | Sykes Reef, Capricorn-Bunker Group Great Barrier Reef, Queensland, Australia | [70] |
| Isophakellin (189) | 231 | C11H13N5O | A. carteri | Madagascan coast | [71] |
| Dibromoisophakellin (190) | 386 | C11H11Br2N5O | A. carteri | Madagascan coast | [71] |
| Mirabilin G (191) | 274 | C17H28N3 | A. cavernosa | Southwestern Australia | [24] |
| Mirabilin K (192) | 276 | C17H30N3 | A. cavernosa | Southwestern Australia | [24] |
| Netamine M (193) | 274 | C17H28N3 | A. cavernosa | Southwestern Australia | [24] |
| 1H-Indole-3-carboxylic acid methyl ester (194) | 175 | C10H9NO2 | A. cavernosa | South China Sea | [50] |
| 1H-Indole-3-carboxylic acid (195) | 161 | C9H7NO2 | A. cavernosa | South China Sea | [50] |
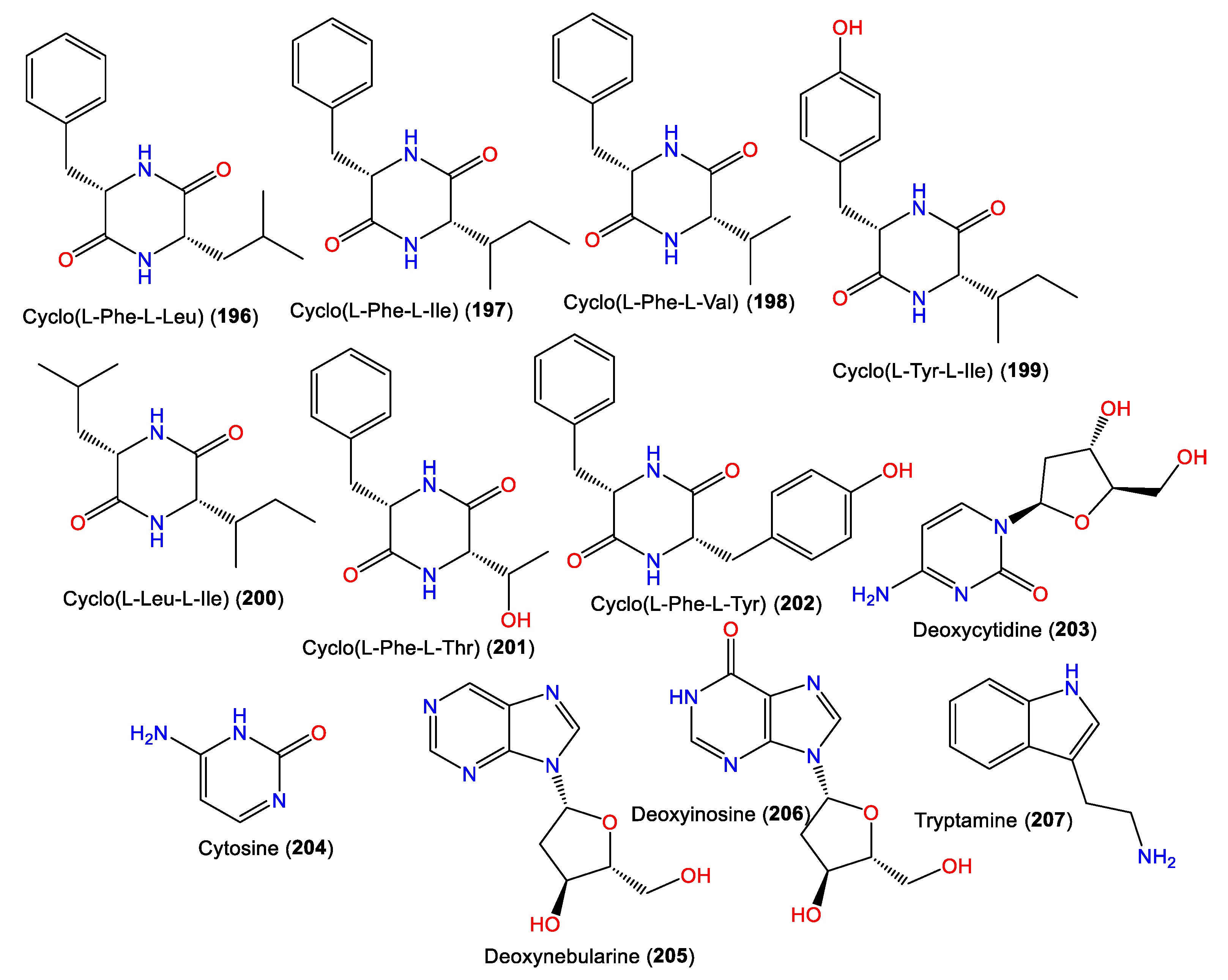
| Cyclo(L-Phe-L-Leu) (196) | 260 | C15H20N2O2 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Phe-L-Ile) (197) | 260 | C15H20N2O2 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Phe-L-Val) (198) | 246 | C14H18N2O2 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Tyr-L-Ile) (199) | 276 | C15H20N2O3 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Leu-L-Ile) (200) | 226 | C12H22N2O2 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Phe-L-Thr) (201) | 248 | C13H16N2O3 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cyclo(L-Phe-L-Tyr) (202) | 310 | C18H18N2O3 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Deoxycytidine (203) | 227 | C9H13N3O4 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Cytosine (204) | 111 | C4H5N3O | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Deoxynebularine (205) | 236 | C10H12N4O3 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Deoxyinosine (206) | 252 | C10H12N4O4 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Tryptamine (207) | 160 | C10H12N2 | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Compound Name | Mol. Wt. | Mol. Formula | Species | Sampling Locations | Ref. |
|---|---|---|---|---|---|
| Steroids | |||||
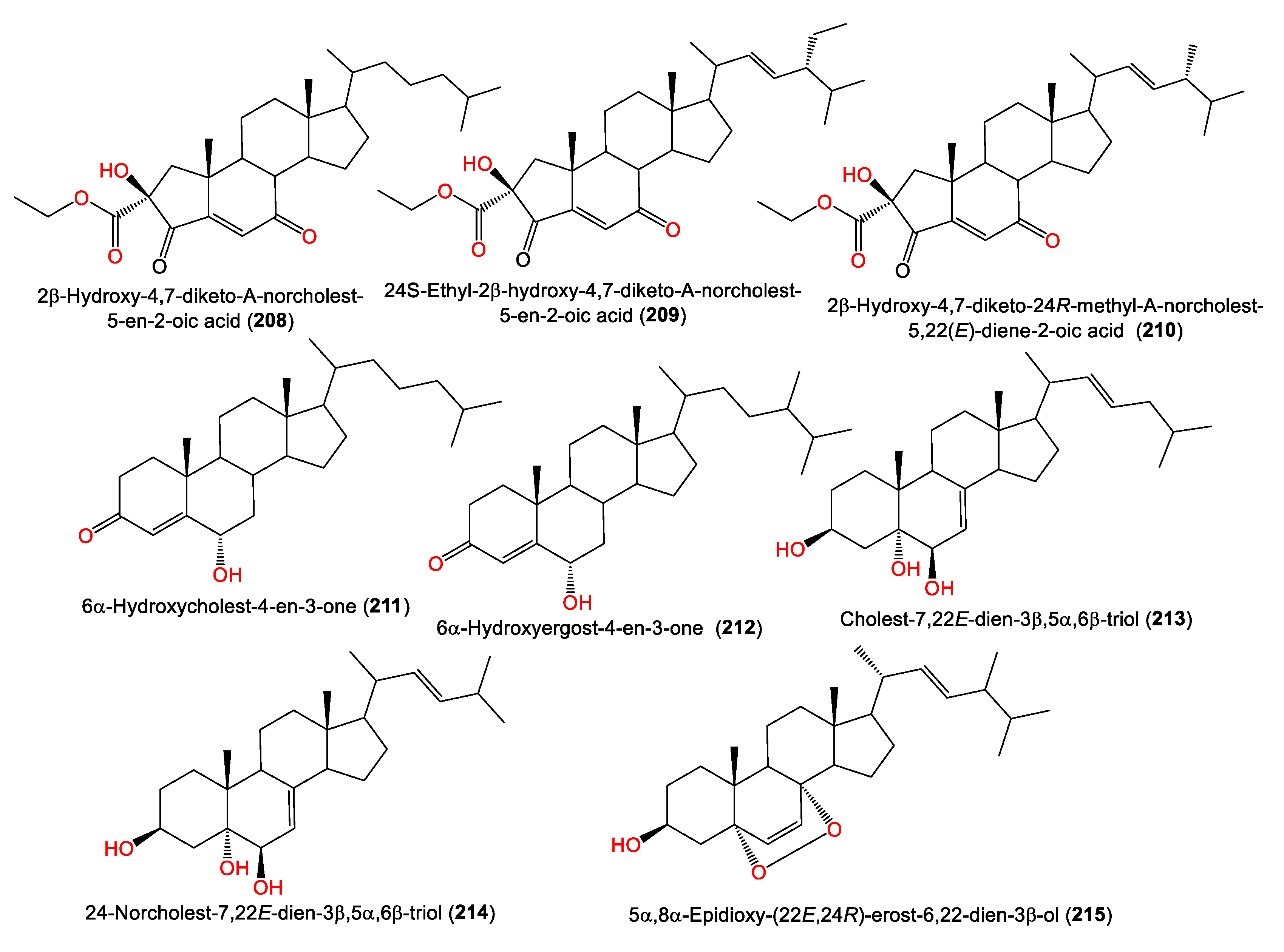
| 2β-Hydroxy-4,7-diketo-A-norcholest-5-en-2-oic acid (208) | 472 | C29H44O5 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 24S-Ethyl-2β-hydroxy-4,7-diketo-A-norcholest-5-en-2-oic acid (209) | 500 | C31H48O5 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 2β-Hydroxy-4,7-diketo-24R-methyl-A-norcholest-5,22(E)-diene-2-oic acid (210) | 484 | C30H44O5 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 6α-Hydroxycholest-4-en-3-one (211) | 400 | C27H44O2 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 6α-Hydroxyergost-4-en-3-one (212) | 414 | C28H46O2 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| Cholest-7,22E-dien-3β,5α,6β-triol (213) | 416 | C27H44O3 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 24-Norcholest-7,22E-dien-3β,5α,6β-triol (214) | 402 | C26H42O3 | A. cavernosa | Inner coral reef, Hainan Island, China | [74] |
| 5α,8α-Epidioxy-(22E,24R)-erost-6,22-dien-3β-ol (215) | 428 | C28H44O3 | A. cavernosa | South China Sea | [50] |
| Other metabolites | |||||
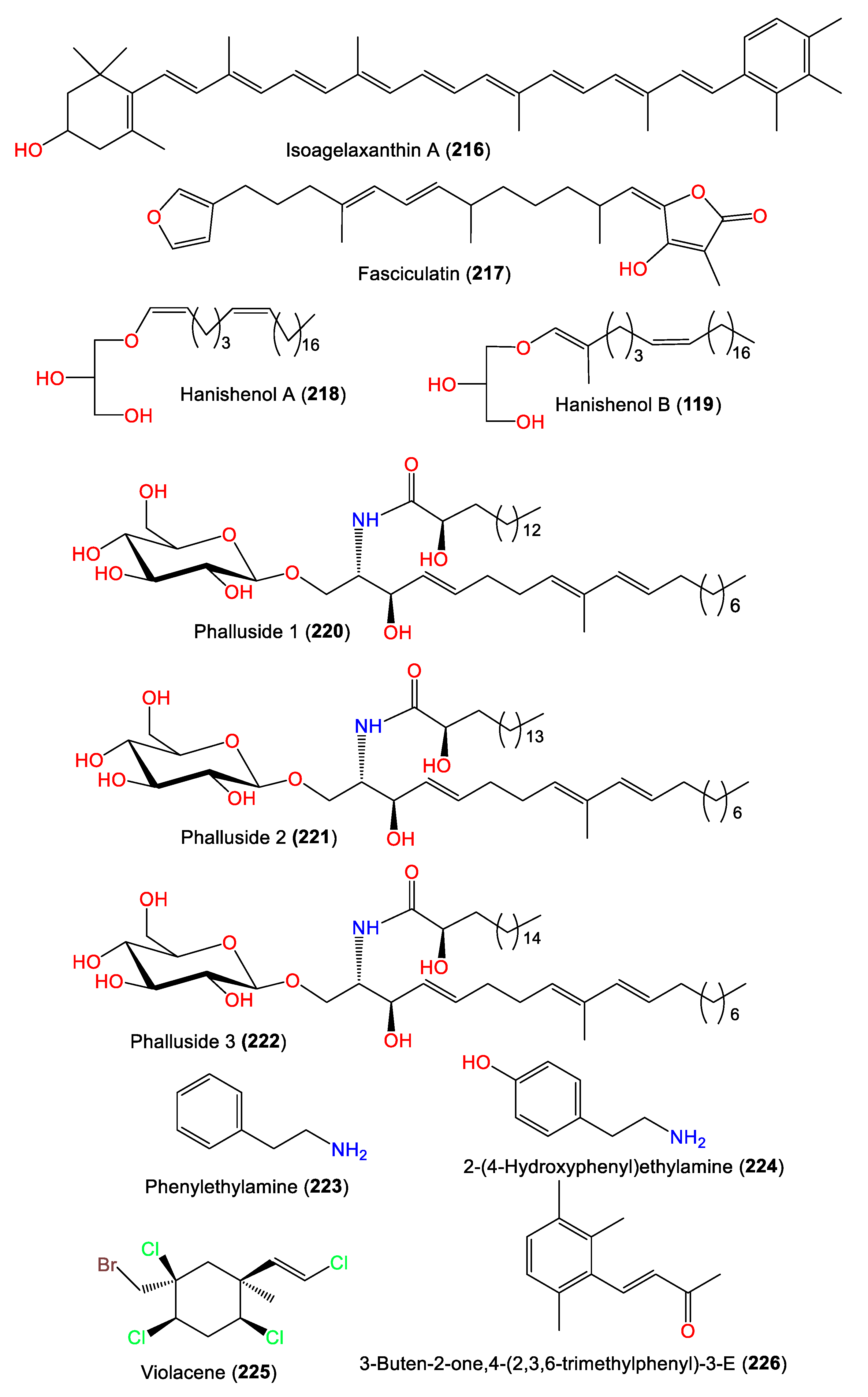
| Isoagelaxanthin A (216) | 548 | C40H52O | A. vulgata | Makura-zaki, Kagoshima, Japan | [75] |
| Fasciculatin (217) | 398 | C25H34O4 | A. acuta | Sidi Elghdamssi island, Monastir region, Tunisia | [25] |
| Hanishenol A (218) | 424 | C27H52O3 | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [76] |
| Hanishenol B (219) | 438 | C28H54O3 | A. carteri | Northern coast, Hanish Island Yemen, South Red Sea | [76] |
| Phalluside 1 (220) | 739 | C42H77NO9 | A. acuta | Sidi Elghdamssi Island, Monastir region, Tunisia | [25] |
| Phalluside 2 (221) | 753 | C43H79NO9 | A. acuta | Sidi Elghdamssi Island, Monastir region, Tunisia | [25] |
| Phalluside 3 (222) | 767 | C44H81NO9 | A. acuta | Sidi Elghdamssi Island, Monastir region, Tunisia | [25] |
| Phenylethylamine (223) | 121 | C8H11N | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| 2-(4-Hydroxyphenyl)ethylamine (224) | 137 | C8H11NO | A. cavernosa | Coast of Vanua Levu (Fiji island), South Pacific Ocean | [72] |
| Violacene (225) | 351 | C10H13BrCl4 | Acanthella sp. | Queen Charlotte Island chain off the coast of British Columbia | [46] |
| 3-Buten-2-one,4-(2,3,6-trimethylphenyl)-3-E (226) | 188 | C13H16O | A. cavernosa | South China Sea | [50] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, S.R.M.; Ghazawi, K.F.; Miski, S.F.; ALsiyud, D.F.; Mohamed, S.G.A.; Mohamed, G.A. Genus Acanthella—A Wealthy Treasure: Secondary Metabolites, Synthesis, Biosynthesis, and Bioactivities. Mar. Drugs 2023, 21, 257. https://doi.org/10.3390/md21040257
Ibrahim SRM, Ghazawi KF, Miski SF, ALsiyud DF, Mohamed SGA, Mohamed GA. Genus Acanthella—A Wealthy Treasure: Secondary Metabolites, Synthesis, Biosynthesis, and Bioactivities. Marine Drugs. 2023; 21(4):257. https://doi.org/10.3390/md21040257
Chicago/Turabian StyleIbrahim, Sabrin R. M., Kholoud F. Ghazawi, Samar F. Miski, Duaa Fahad ALsiyud, Shaimaa G. A. Mohamed, and Gamal A. Mohamed. 2023. "Genus Acanthella—A Wealthy Treasure: Secondary Metabolites, Synthesis, Biosynthesis, and Bioactivities" Marine Drugs 21, no. 4: 257. https://doi.org/10.3390/md21040257
APA StyleIbrahim, S. R. M., Ghazawi, K. F., Miski, S. F., ALsiyud, D. F., Mohamed, S. G. A., & Mohamed, G. A. (2023). Genus Acanthella—A Wealthy Treasure: Secondary Metabolites, Synthesis, Biosynthesis, and Bioactivities. Marine Drugs, 21(4), 257. https://doi.org/10.3390/md21040257