
Indole Diterpenes from Mangrove Sediment-Derived Fungus Penicillium sp. UJNMF0740 Protect PC12 Cells against 6-OHDA-Induced Neurotoxicity via Regulating the PI3K/Akt Pathway


Abstract
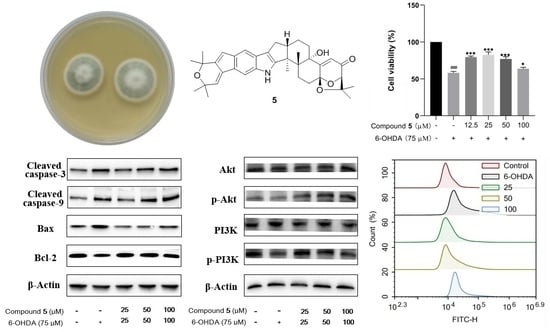
:
1. Introduction
2. Results
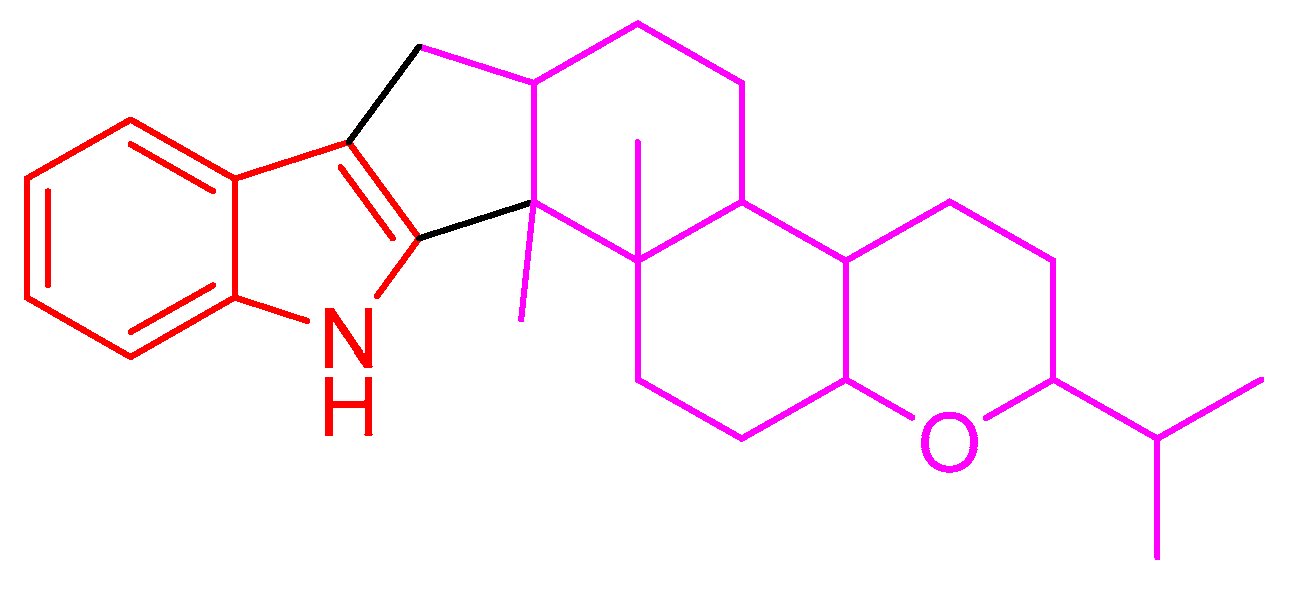
2.1. Structural Elucidation
2.2. Antimicrobial Activities
2.3. Neuroprotective Effects
3. Materials and Methods
3.1. General Experimental Procedure
3.2. Fungal Material
3.3. Fermentation, Extraction, and Isolation
3.4. Assay of Antimicrobial Activity
3.5. Cell Line and Culture
3.6. Cell Viability Assay
3.7. Hoechst 33258-Staining Assay
3.8. Flow Cytometric Analysis
3.9. Western Blot Assay
3.10. Intracellular ROS Assay
3.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reddy, P.; Guthridge, K.; Vassiliadis, S.; Hemsworth, J.; Hettiarachchige, I.; Spangenberg, G.; Rochfort, S. Tremorgenic Mycotoxins: Structure Diversity and Biological Activity. Toxins 2019, 11, 302. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Chen, M.; Sun, Z.; Ma, G.; Chen, D.; Wu, H.; Yang, J.; Li, Y.; Xu, X. The Biosynthesis Related Enzyme, Structure Diversity and Bioactivity Abundance of Indole-Diterpenes: A Review. Molecules 2022, 27, 6870. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Qi, J.; Wang, P.; Liu, C.; Gao, J.-M. The chemical structures and biological activities of indole diterpenoids. Nat. Prod. Bioprospect. 2023, 13, 3. [Google Scholar] [CrossRef]
- Imlach, W.L.; Finch, S.C.; Dunlop, J.; Dalziel, J.E. Structural determinants of lolitrems for inhibition of BK large conductance Ca2+-activated K+ channels. Eur. J. Pharmacol. 2009, 605, 36–45. [Google Scholar] [CrossRef]
- Xu, M.; Gessner, G.; Groth, I.; Lange, C.; Christner, A.; Bruhn, T.; Deng, Z.; Li, X.; Heinemann, S.H.; Grabley, S.; et al. Shearinines D-K, new indole triterpenoids from an endophytic Penicillium sp. (strain HKI0459) with blocking activity on large-conductance calcium-activated potassium channels. Tetrahedron 2007, 63, 435–444. [Google Scholar] [CrossRef]
- Tomoda, H.; Tabata, N.; Yang, D.J.; Takayanagi, H.; Omura, S. Terpendoles, novel ACAT inhibitors produced by Albophoma yamanashiensis. III. Production, isolation and structure elucidation of new components. J. Antibiot. 1995, 48, 793–804. [Google Scholar] [CrossRef]
- Nakazawa, J.; Yajima, J.; Usui, T.; Ueki, M.; Takatsuki, A.; Imoto, M.; Toyoshima, Y.Y.; Osada, H. A novel action of terpendole E on the motor activity of mitotic Kinesin Eg5. Chem. Biol. 2003, 10, 131–137. [Google Scholar] [CrossRef]
- Matsui, C.; Ikeda, Y.; Iinuma, H.; Kushida, N.; Kunisada, T.; Simizu, S.; Umezawa, K. Isolation of a novel paxilline analog pyrapaxilline from fungus that inhibits LPS-induced NO production. J. Antibiot. 2014, 67, 787–790. [Google Scholar] [CrossRef]
- Ogata, M.; Ueda, J.-y.; Hoshi, M.; Hashimoto, J.; Nakashima, T.; Anzai, K.; Takagi, M.; Shin-ya, K. A novel indole-diterpenoid, JBIR-03 with anti-MRSA activity from Dichotomomyces cejpii var. cejpii NBRC 103559. J. Antibiot. 2007, 60, 645–648. [Google Scholar] [CrossRef]
- Nakanishi, K.; Doi, M.; Usami, Y.; Amagata, T.; Minoura, K.; Tanaka, R.; Numata, A.; Yamada, T. Anthcolorins A-F, novel cytotoxic metabolites from a sea urchin-derived Aspergillus versicolor. Tetrahedron 2013, 69, 4617–4623. [Google Scholar] [CrossRef]
- Sallam, A.A.; Houssen, W.E.; Gissendanner, C.R.; Orabi, K.Y.; Foudah, A.I.; El Sayed, K.A. Bioguided discovery and pharmacophore modeling of the mycotoxic indole diterpene alkaloids penitrems as breast cancer proliferation, migration, and invasion inhibitors. Medchemcomm 2013, 4, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and Structure Elucidation of Janthitrems A and D from Penicillium janthinellum and Determination of the Tremorgenic and Anti-Insect Activity of Janthitrems A and B. J. Agric. Food Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wang, Y.; Liu, P.; Fu, P.; Zhu, T.; Wang, W.; Zhu, W. Indole-Diterpenoids with Anti-H1N1 Activity from the Aciduric Fungus Penicillium camemberti OUCMDZ-1492. J. Nat. Prod. 2013, 76, 1328–1336. [Google Scholar] [CrossRef]
- Ozaki, T.; Minami, A.; Oikawa, H. Biosynthesis of indole diterpenes: A reconstitution approach in a heterologous host. Nat. Prod. Rep. 2023, 40, 202–213. [Google Scholar] [CrossRef]
- Chen, Z.L. Secondary Metabolites of Two Mangrove Sediment Fungi and Their Biological Activities. Master’s Thesis, University of Jinan, Jinan, China, 2022. [Google Scholar]
- Hu, X.-Y.; Meng, L.-H.; Li, X.; Yang, S.-Q.; Li, X.-M.; Wang, B.-G. Three New Indole Diterpenoids from the Sea-Anemone-Derived Fungus Penicillium sp. AS-79. Mar. Drugs 2017, 15, 137. [Google Scholar] [CrossRef]
- Ariantari, N.P.; Ancheeva, E.; Wang, C.; Mandi, A.; Knedel, T.-O.; Kurtan, T.; Chaidir, C.; Mueller, W.E.G.; Kassack, M.U.; Janiak, C.; et al. Indole Diterpenoids from an Endophytic Penicillium sp. J. Nat. Prod. 2019, 82, 1412–1423. [Google Scholar] [CrossRef]
- Smetanina, O.F.; Kalinovsky, A.I.; Khudyakova, Y.V.; Pivkin, M.V.; Dmitrenok, P.S.; Fedorov, S.N.; Ji, H.; Kwak, J.-Y.; Kuznetsova, T.A. Indole alkaloids produced by a marine fungus isolate of Penicillium janthinellum Biourge. J. Nat. Prod. 2007, 70, 906–909. [Google Scholar] [CrossRef]
- You, J.; Du, L.; King, J.B.; Hall, B.E.; Cichewicz, R.H. Small-Molecule Suppressors of Candida albicans Biofilm Formation Synergistically Enhance the Antifungal Activity of Amphotericin B against Clinical Candida Isolates. ACS Chem. Biol. 2013, 8, 840–848. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Antiinsectan Alkaloids: Shearinines A-C and a New Paxilline Derivative from the Ascostromata of Eupenicillium shearii. Tetrahedron 1995, 51, 3959–3968. [Google Scholar] [CrossRef]
- Liang, Z.-Y.; Shen, N.-X.; Zhou, X.-J.; Zheng, Y.-Y.; Chen, M.; Wang, C.-Y. Bioactive indole diterpenoids and polyketides from the marine-derived fungus Penicillium javanicum. Chem. Nat. Compd. 2020, 56, 379–382. [Google Scholar] [CrossRef]
- Nozawa, K.; Nakajima, S.; Kawai, K.I.; Udagawa, S.I. Isolation and structures of indoloditerpenes, possible biosynthetic intermediates to the tremorgenic mycotoxin, paxilline, from Emericella striata. J. Chem. Soc. Perkin. Trans. I 1988, 9, 2607–2610. [Google Scholar] [CrossRef]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell. Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Kang, M.; Chang, C.; Wei, H.; Zhang, C.; Chen, Y. Multiple forms of cell death: A focus on the PI3K/AKT pathway. J. Cell. Physiol. 2023, 238, 2206–2238. [Google Scholar] [CrossRef] [PubMed]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, B.; Xu, L.; Yu, S.; Fu, J.; Wang, J.; Yan, X.; Su, J. ROS-Induced mtDNA Release: The Emerging Messenger for Communication between Neurons and Innate Immune Cells during Neurodegenerative Disorder Progression. Antioxidants 2021, 10, 1917. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Liu, X.; Lian, M.; Mao, Y. Citronellol Prevents 6-OHDA-Induced Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis in Parkinson Disease Model of SH-SY5Y Cells via Modulating ROS-NO, MAPK/ERK, and PI3K/Akt Signaling Pathways. Neurotoxic. Res. 2022, 40, 2221–2237. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhai, H.; Zhu, K.; Yu, J.-H.; Zhang, Y.; Wang, Y.; Jiang, C.-S.; Zhang, X.; Zhang, Y.; Zhang, H. Bioactive Pyridone Alkaloids from a Deep-Sea-Derived Fungus Arthrinium sp. UJNMF0008. Mar. Drugs 2018, 16, 174. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 | 4 | ||||
|---|---|---|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | δC | δH (J in Hz) | δC | δH (J in Hz) | |
| 1 | 7.95, s | |||||||
| 2 | 157.9, C | 180.0, C | 176.8, C | 155.4, C | ||||
| 3 | 58.2, C | 53.9, C | 57.5, C | 52.1, C | ||||
| 4 | 41.6, C | 40.3, C | 41.5, C | 40.0, C | ||||
| 5 | 28.2, CH2 | 2.75, m | 28.0, CH2 | 2.71, m 2.72, m | 27.5, CH2 | 2.85, dd, 15.1, 9.7 1.55, m | 27.2, CH2 | 2.70, dd, 12.4, 10.2 1.84, dd, 12.4, 9.7 |
| 6 | 28.5, CH2 | 2.67, m 1.95, dd, 12.8, 5.0 | 28.6, CH2 | 1.59, m 1.91, m | 28.5, CH2 | 2.69, dd, 10.4, 7.1 1.95, m | 28.4, CH2 | 2.84, m 2.06, m |
| 7 | 104.1, C | 104.0, C | 104.1, C | 104.4, C | ||||
| 9 | 88.0, CH | 4.29, d, 1.2 | 88.1 CH | 4.30, s | 88.0, CH | 4.28, d, 1.2 | 88.1, CH | 4.33, s |
| 10 | 196.9, C | 197.1, C | 196.9, C | 197.1, C | ||||
| 11 | 118.2, CH | 5.73, brs | 118.5, CH | 5.84, s | 118.4, CH | 5.73, s | 118.0, CH | 5.85, brs |
| 12 | 169.4, C | 169.5, C | 169.4, C | 169.4, C | ||||
| 13 | 77.3, C | 77.4, C | 77.4, C | 77.8, C | ||||
| 14 | 31.7, CH2 | 1.71, m | 30.9, CH2 | 1.74, m 1.87, m | 31.8, CH2 | 1.95, m 1.57, m | 34.0, CH2 | 1.99, m 1.92, m |
| 15 | 25.1, CH2 | 1.88, ddd, 26.1, 12.9, 7.8 1.51, m | 22.9, CH2 | 1.68, m1.74, m | 25.7, CH2 | 1.75, m 1.57, m | 21.1, CH2 | 2.09, m 1.83, m |
| 16 | 35.2, CH | 3.01, m | 33.9, CH | 3.07, m | 34.6, CH | 3.00, m | 48.7, CH | 2.84, m |
| 17 | 48.9, CH2 | 2.91, dd, 18.6, 6.2 2.32, dd, 18.6, 2.6 | 43.9, CH2 | 3.02, d, 14.4 2.64, dd, 14.9, 10.4 | 48.4, CH2 | 3.10, dd, 18.0, 5.3 2.47, dd, 18.0, 3.3 | 27.6, CH2 | 2.76, dd, 13.2, 6.4 2.47, dd, 13.2, 10.6 |
| 18 | 205.3, C | 201.2, C | 203.0, C | 119.2, C | ||||
| 19 | 122.3, C | 115.1, C | 135.2, C | 127.1, C | ||||
| 20 | 123.0, CH | 7.48, s | 125.6, CH | 7.91, s | 125.5, CH | 7.70, s | 102.8, CH | 7.86, s |
| 21 | 145.0, C | 129.9, C | 145.2, C | 130.5, C | ||||
| 22 | 36.8, CH2 | 3.33, m | 36.4, CH2 | 3.32, s | 76.3, CH | 4.91, d, 6.3 | 201.7, C | |
| 23 | 149.3, C | 151.8, C | 59.8, CH | 2.66, dd, 6.3, 3.0 | 56.0, CH | 3.03, d, 2.3 | ||
| 24 | 73.7, C | 73.7, C | 73.8, C | 74.2, C | ||||
| 26 | 71.5, C | 71.5, C | 72.6, C | 72.8, C | ||||
| 27 | 34.5, CH2 | 2.37, m | 34.3, CH2 | 2.34, brs | 127.3, CH | 6.15, d, 3.0 | 124.2, CH | 6.20, d, 2.3 |
| 28 | 131.9, C | 131.9, C | 133.0, C | 141.1, C | ||||
| 29 | 136.3, C | 151.6, C | 142.7, C | 131.5, C | ||||
| 30 | 113.3, CH | 6.55, s | 106.4, CH | 6.47, s | 118.7, CH | 7.06, s | 114.3, CH | 7.52, s |
| 31 | 149.7, C | 151.1, C | 137.7, C | 145.2, C | ||||
| 32 | 17.6, CH3 | 1.61, s | 15.5, CH3 | 1.56, s | 17.7, CH3 | 1.66, s | 16.4, CH3 | 1.42, s |
| 33 | 25.2, CH3 | 1.16, s | 24.8, CH3 | 1.23, s | 24.6, CH3 | 1.19, s | 23.9, CH3 | 1.23, s |
| 34 | 78.8, C | 78.7, C | 78.8, C | 79.0, C | ||||
| 35 | 23.2, CH3 | 1.15, s | 23.2, CH3 | 1.16, s | 23.2, CH3 | 1.15, s | 23.3, CH3 | 1.19, s |
| 36 | 29.0, CH3 | 1.432, s | 30.0, CH3 | 1.41, s | 29.0, CH3 | 1.42, s | 29.0, CH3 | 1.45, s |
| 37 | 30.9, CH3 | 1.41, s | 29.6, CH3 | 1.42, s | 23.5, CH3 | 1.15, s | 29.8, CH3 | 1.67, s |
| 38 | 30.1, CH3 | 1.427, s | 29.6, CH3 | 1.41, s | 30.2, CH3 | 1.47, s | 22.3, CH3 | 1.00, s |
| 39 | 29.2, CH3 | 1.28, s | 30.3, CH3 | 1.30, s | 31.6, CH3 | 1.32, s | 31.9, CH3 | 1.40. s |
| 40 | 30.2, CH3 | 1.33, s | 30.3, CH3 | 1.30, s | 29.5, CH3 | 1.36, s | 29.9, CH3 | 1.38, s |
| 41 | 53.3, CH3 | 3.64, s | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-X.; Chen, Z.-L.; Zhang, J.-S.; Liu, H.-S.; Ma, R.-P.; Liu, X.-P.; Li, M.-Y.; Ge, D.; Bao, J.; Zhang, H. Indole Diterpenes from Mangrove Sediment-Derived Fungus Penicillium sp. UJNMF0740 Protect PC12 Cells against 6-OHDA-Induced Neurotoxicity via Regulating the PI3K/Akt Pathway. Mar. Drugs 2023, 21, 593. https://doi.org/10.3390/md21110593
Wang X-X, Chen Z-L, Zhang J-S, Liu H-S, Ma R-P, Liu X-P, Li M-Y, Ge D, Bao J, Zhang H. Indole Diterpenes from Mangrove Sediment-Derived Fungus Penicillium sp. UJNMF0740 Protect PC12 Cells against 6-OHDA-Induced Neurotoxicity via Regulating the PI3K/Akt Pathway. Marine Drugs. 2023; 21(11):593. https://doi.org/10.3390/md21110593
Chicago/Turabian StyleWang, Xin-Xin, Ze-Long Chen, Jun-Sheng Zhang, Hai-Shan Liu, Ruo-Ping Ma, Xin-Ping Liu, Ming-Yue Li, Di Ge, Jie Bao, and Hua Zhang. 2023. "Indole Diterpenes from Mangrove Sediment-Derived Fungus Penicillium sp. UJNMF0740 Protect PC12 Cells against 6-OHDA-Induced Neurotoxicity via Regulating the PI3K/Akt Pathway" Marine Drugs 21, no. 11: 593. https://doi.org/10.3390/md21110593
APA StyleWang, X.-X., Chen, Z.-L., Zhang, J.-S., Liu, H.-S., Ma, R.-P., Liu, X.-P., Li, M.-Y., Ge, D., Bao, J., & Zhang, H. (2023). Indole Diterpenes from Mangrove Sediment-Derived Fungus Penicillium sp. UJNMF0740 Protect PC12 Cells against 6-OHDA-Induced Neurotoxicity via Regulating the PI3K/Akt Pathway. Marine Drugs, 21(11), 593. https://doi.org/10.3390/md21110593