Abstract
The term metabolic/cardiometabolic/insulin resistance syndrome could generally be defined as the co-occurrence of several risk factors inclusive of systemic arterial hypertension. Not only that organizations, such as the world health organization (WHO) have identified high blood pressure as one of the main risk factors of the cardiometabolic syndrome, but there is also a link between the occurrence of insulin resistance/impaired glucose tolerance and hypertension that would consequently lead to type-2 diabetes (T2D). Hypertension is medicated by various classes of synthetic drugs; however, severe or mild adverse effects have been repeatedly reported. To avoid and reduce these adverse effects, natural alternatives, such as bioactive peptides derived from different sources have drawn the attention of researchers. Among all types of biologically active peptides inclusive of marine-derived ones, this paper’s focus would solely be on fish and fishery by-processes’ extracted peptides and products. Isolation and fractionation processes of these products alongside their structural, compositional and digestion stability characteristics have likewise been briefly discussed to better address the structure-activity relationship, expanding the reader’s knowledge on research and discovery trend of fish antihypertensive biopeptides. Furthermore, drug-likeness of selected biopeptides was predicted by Lipinski’s rules to differentiate a drug-like biopeptide from nondrug-like one.
1. Introduction
Cardiometabolic syndrome (CMS) is defined as a cluster of several risk factors by various health organizations which marginally varies among all (Table 1) [1]. These risk factors according to WHO manifest in subjects with insulin-resistance of normal glucose tolerance, 10%, subjects of impaired fasting glucose/impaired glucose tolerance, 50%, and subjects of T2D, 80% [2]. These risk factors are undoubtedly interconnected with almost identical underlying mediators, mechanisms and pathways (evident prothrombotic and pro-inflammatory state) that subsequently lead to the development of cardiovascular diseases (CVD) and diabetes [3]. Hence, global mortality rates associated with CVD, as the leading cause of death, were as high as 17 million in 2012 [4]. Along with CVD, diabetes-associated complications as the fourth/fifth causes of death worldwide claimed the lives of 3.2 million in 2003 [4].

Table 1.
Definition and criteria of metabolic syndrome by different organizations.
Adapted from Grundy et al. [1] Subjects diagnosed with metabolic syndrome are two-times as likely to face death, five-times as likely to develop T2D, and three-times as likely to experience a heart attack or stroke in comparison to subjects without the syndrome [3]. Statistics estimate 20–25% of metabolic syndrome cases among the world’s adult population and the same trend is accordingly observed among North Americans [4]. During the past two decades number of T2D cases have nearly doubled due to inappropriate dietary habits (e.g., a diet high in saturated fat and cholesterol) and lifestyle (with 1.1 billion overweight and 312 million obese adults) [5]. In the year 2015, 415 million cases of T2D have been reported by International Diabetes Federation that is predicted to rise to 642 million by 2040 with the estimated cost of 500 billion dollars annually for the health care system, worldwide [6]. These estimates have been confirmed by non-communicable diseases (NCD) risk factor collaboration and WHO. Not only that metabolic syndrome greatly distresses the life expectancy of individuals, but also enforces significant economic burden on societies (Table 2) [7]. According to Lind et al., 25–47% of the subjects with hypertension happen to be diagnosed with insulin resistance/impaired glucose tolerance [8]. High blood pressure alone, as a remediable factor, had been responsible for 6% of the deaths in 1997 worldwide [8,9,10]. Hypertension has been recognized by the National Cholesterol Education Program Adult Treatment Panel III (NCEP-ATP III) and WHO as one of the main risk factors leading to CMS (Table 1) [1].
Table 2.
The cost of care for hypertensive patients with metabolic syndrome components in the European Community.
Hypertension has a broad definition not only outlining blood pressure, but also its association with functional and structural cardiac and vascular abnormalities which consequently damage the target organs (heart, kidneys, brain, vasculature and other organs) causing premature morbidity and mortality [11]. Blood pressure affecting 25–30% of developed countries’ population, as the conventional biomarker, now is announced as an element of a subject’s total cardiovascular risk rather than as a sole risk factor [11,12]. According to 1998 USA statistics, high blood pressure distresses 24–31% of its population enforcing $108 billion on the health care system [13]. Worldwide, in 2000, hypertension affected approximately 972 million and is forecasted to reach 1.56 billion by 2025 [14]. Furthermore, pre-hypertension, an inflammatory associated condition initiating a more severe hypertensive state, is affected by alterations of the immune system [15]. Cytokines, interleukin 6 (IL-6) and tumor necrosis factor alpha (TNF-α), which regulate the production of high-sensitivity C-reactive protein (hsCRP) correspondingly are linked to high blood pressure and obesity [12]. TNF-α and IL-1β levels, counter regulated by IL-6 and soluble TNF-α receptor/soluble TNF-α receptor 2 (sTNFR1/sTNFR2) by TNF-α, correlate with impaired endothelium-dependent dilatation and induction of insulin resistance followed by high blood pressure, respectively [16]. Systemic and chronic mild (low grade) inflammation impairs vasodilation mechanism leading to endothelial dysfunction which is a predecessor factor in the co-occurrence of dyslipidemia, impaired fibrinolysis, and hypertension that are ascribed to insulin resistance and subsequently obesity, as well as diabetes [13,15].
In addition to obesity and diabetes, hypertension is potentially a variable known risk factor for atrial fibrillation which is the major component of stroke and associated mortality; nevertheless, its pathophysiological mechanism is not yet well understood [12]. Inflammation, on the one hand, is a catalyst for the structural changes of the left ventricle and the left atrium caused by atrial fibrillation; on the other hand, it is important in inducing the production of angiotensin II, as well as enhanced activity of renin-angiotensin-aldosterone system (RAAS) (Figure 1) [12,17]. Blood pressure, high blood pressure and left ventricle hypertrophy incidences are directly related to high amounts of hsCRP which are produced in chronically inflamed tissues [12]. Inflammatory infiltrates, and oxidative damage upregulate the expression of angiotensin-II receptors causing increased atrial cell death and leukocyte infiltration that are clearly found within the arterial tissue in atrial fibrillation cases [12,18]. Albeit, there is more evidence on the association of atrial fibrillation with heart’s inflammatory and systemic inflammatory diseases (myocarditis, pericarditis, psoriasis).
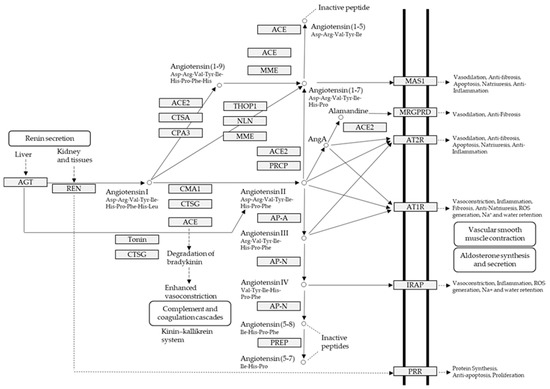
Figure 1.
Human renin-angiotensin system pathway map. AGT: Angiotensinogen, ACE2: Angiotensin I converting enzyme 2, AngA: Angiotensin A, AP-A: Glutamyl aminopeptidase, AP-N: Alanyl aminopeptidase (membrane), AT1R: Angiotensin II receptor type 1, AT2R: Angiotensin II receptor type 2, CMA1: Chymase 1, CPA3: Carboxypeptidase A3, CTSA: Cathepsin A, CTSG: Cathepsin G, IRAP: Leucyl and cystinyl aminopeptidase, MAS1: MAS1 proto-oncogene, G protein-coupled receptor, MME: Membrane metalloendopeptidase, MRGPRD: MAS related GPR family member D, NLN: Neurolysin, PRCP: Prolylcarboxypeptidase, PREP: Prolyl endopeptidase, PRR: ATPase H+ transporting accessory protein 2, REN: Renin, THOP1: Thimet oligopeptidase. Figure 1 has been adapted and modified from KEGG Pathway Maps.
Several classes of drugs; direct inhibitors of the renin-angiotensin system (angiotensin-converting enzyme inhibitors, angiotensin-receptor blockers), adrenergic beta-antagonists, diuretics, sodium chloride symport inhibitors, and calcium channel blockers, are prescribed for the treatment of hypertension, though, accompanying side-effects limit their practice in medicine. For instance, upon ingestion of angiotensin-I-converting enzyme (ACE) inhibitors patient may encounter severe or mild adverse effects, such as cough, headache, diarrhea, dizziness, fatigue, angioedema, hyperkalemia or, in rare cases, renal and cardiac failure [19]. Different classes of drugs are suggested, corresponding to the indication of hypertension. For example, with an indication of heath failure diuretics, beta-blockers, ACE inhibitors, angiotensin receptor blockers, calcium channel blockers and aldosterone antagonists are the first-line medications recommended for the effective management of hypertension [20]. For chronic kidney disease indication, ACE inhibitors, angiotensin receptor blockers, and calcium channel blockers are suggested to bring the blood pressure under control [20]. New pharmaceutical ingredients are designed in a way to target the inflammatory processes and modulations of its pathways (RAAS-blockers and statins) for the treatment of hypertension and related disorders [21]. In agreement cholesterol level is not any longer recognized as dyslipidemia treatment goal for reduction of atherosclerotic cardiovascular risk in 2013 published guidelines of the American Heart Association and American College of Cardiology. Correspondingly, CVD risk prediction algorithms are now based on hypertension, and not on low-density lipoprotein anymore, increasing the sum of combination therapy (e.g., RAAS-blockers and statins) prescribed patients [22].
In the concept of prevention and treatment of CMS associated risk factors inclusive of hypertension, natural products, such as peptides have shown promising effects. Many studies have shown that bioactive peptides from animals (terrestrial, marine), and plant sources, in addition to their many health effects, could also be utilized in the effective management of hypertension, a key factor of cardiometabolic diseases occurrences [23,24,25,26,27,28,29]. ACE inhibition activity of food-derived peptides, inclusive of fish-extracted ones, are primarily by competitive inhibition mode rather than non- and or uncompetitive modes [28,30,31]. Food-originated anti-ACE biopeptides bind to the enzyme and hinder its activity, bringing the blood pressure under control. For example, in a study by Fang et al. interactions of three food-isolated oligopeptides with ACE have thoroughly been studied, by computational modeling, elucidating different inhibition mechanisms of antihypertensive biopeptides (Figure 2) [32]. The majority of anti-ACE peptides are not specific to one family of an animal or a plant and can certainly be found in many other species, e.g., antihypertensive salmon di- and tri-peptides with comparable animal-, plant-, marine- and or insect-origin sequences (Table 3). For instance, fractions and subfractions of jellyfish enzymatic hydrolysate have hindered the ACE activity at IC50 as low as 1.28 mg mL−1 and 0.16 mg mL−1 respectively in an in-vitro experimental model [33]. The most potent penta-peptides of the same hydrolysate, isolated by gel and high-performance liquid chromatography (HPLC) with polar amino acids at the C-terminal, also had blood pressure (BP) decreasing effect in spontaneously hypertensive rats (SHR) when administered orally at the concentration of 10 mg kg−1 body weight (BW) [33]. The above is only an example of many marine animals including many fish types with anti-hypertension activities [34]. Varying fish-derived biopeptides, in terms of MW, length and composition, have inhibited the activity of ACE and controlled blood pressure successfully in the in-vitro and or in the in-vivo experimental settings, as well as human studies. Anti-ACE fish biopeptides, similar to other bioactives, mostly exert their effect on the subject non-cytotoxically, negligible and or zero cytotoxicity, making them superior to the synthetic drugs for treatment and prevention of hypertension. These biopeptides can be produced by digestion, isolation, and purification processes, affecting the structural, compositional and characteristics of the final product, discussed further in the following sections. Authors herein have solitary tried to bring together more than twenty studies directed on fish-extracted peptides or fish-originated products demonstrating their antihypertensive and or ACE inhibitory effects in different experimental settings and models.
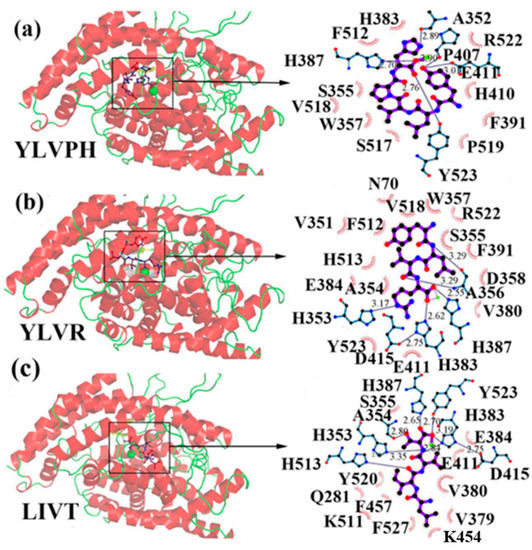
Figure 2.
3D plots of the frontier molecular orbital of three food-extracted anti-ACE (angiotensin-I-converting enzyme) biopeptides. Per docking results Tyr-Leu-Val-Pro-His occupied an active groove surrounded by Ala352, Glu411, Tyr523, and His387 (His387, His512, Ser355, Val518, Trp357, Ser517, Pro519, Pro391, His410, and Arg522 having van der Waals contacts with the penta-peptide YLAPH), Tyr-Leu-Val-Arg occupied an active groove surrounded by His353, Ala356, His383, and His387 (Asn70, Val518, Trp357, Arg522, Val351, Phe512, and Ser355 having van der Waals contacts with the tetra-peptide, YLVR), Leu-Ile-Val-Thr occupied an active groove surrounded by Ala356, His353, His383, His387, His513, Tyr523 (Ser355, Glu411, Tyr520, Gln281, Phe457, Lys511, Phe527, Lys454, Val379, and Val380 having van der Waals contacts with the tetra-peptide, LIVT). Adapted and modified from Fang et al. [32].
Table 3.
Salmon di- and tri- peptides with identical counterpart sequences found in sources other than fish.
2. Biological Activity of Fish, Fish By-Product Protein and Peptides on Hypertension
ACE (with alternative names of dipeptidyl carboxypeptidase I and kininase II), a carboxydipeptidase, deactivates bradykinin (a vasodilator) and controls blood pressure through converting angiotensin-I (an inactive decapeptide prohormone, Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu) to angiotensin-II (a potent octapeptide vasoconstrictor, Asp-Arg-Val-Tyr-Ile-His-Pro-Phe) stimulating the release of aldosterone, and thus, increasing blood pressure (Figure 1) [13]. Physiochemical properties of human ACE (predicted by ProtParam), its active sites and sequence are listed, and shown in Table 4 and Figure 3 [37]. ACE inhibitors, e.g., captopril, are regularly used to treat hypertension, and other cardio-related diseases as an influencer of blood pressure [21]. Proline moiety of captopril interacts with ACE active site, primarily through two histidine residues, inhibiting its activity [38]. Binding sites of captopril on ACE have been observed at Trp67, Asn68, Thr71, Asn72, Met340 and Arg348 residues [39]. Generally, the proline residue, in most of the captopril alike anti-ACE drugs, e.g., enalaprilat, zofenoprilat, fosinoprilat, etc., impedes the enzyme’s activity [38].
Table 4.
Physiochemical properties of ACE.

Figure 3.
Sequence and secondary structure of chain A human ACE (PDB code: 1O8A). Active sites, according to Attique et al. are; His317, Ala318, Ser319, His347, Glu348, His351, Glu375, Phe421, Lys475, Phe476, His477, Val482, Tyr484, Tyr487.
Synthetic ACE inhibitors impose rather significant side effects, such as a dry cough and/or contradiction with other medications on the subjects, therefore, natural plant, animal, and marine-based alternatives have attracted great attention recently [35,40]. Sulfhydryl-containing, dicarboxylate-containing, and phosphonate-containing agents are the main groups of ACE inhibitors [41]. Among many types of bioactive compounds, peptides from various sources, such as bovine, egg, milk and plants have been the subject of many studies, and shown biological activity against hypertension, e.g., di- and tri-peptides of different sources with matching salmon-extracted sequences (Table 3) [35]. Kumar et al. have extensively reviewed natural ACE inhibitors [35]. Among marine-based peptides whether plants or animals, fish bioactive peptides derived using various extraction, isolation and fractionation techniques have also been investigated for their anti-hypertensive activities, such as ACE inhibition (Table 5 and Table 6). Of all the farmed and wild fish species, many carp, salmon, tilapia, sardine, and tuna have been analyzed for their biological activities (Table 6). These species’ proteinaceous portion contains not only the essential amino acids possessing all the parameters of a good food, but also their associated hydrolysates have countless health benefits inclusive of antihypertension effects (Table 6).
Table 5.
Fractionation, purification and characterization methods of fish anti-ACE and antihypertensive peptides.
Table 6.
Biological activity of fish and fish by-products’ derived peptides on hypertension.
In an in-vivo experimental setting, after two hours (h) treatment, tested grass carp material showed significant antihypertensive activity (43 mmHg systolic blood pressure (SBP) maximal drop at the dosage of 100 mg kg−1 body weight (BW)) in SHR compared to the control group [42]. Grass carp peptides of size 725–1228 Da, rich in leucine, aspartic acid, phenylalanine and glycine amino acids, at 100 mg kg−1 BW in SHR 6 h post-administration significantly reduced SBP of approximately 30 mm Hg, in comparison to the control group (captopril), the medication of choice, with similar effect at dosage of 10 mg kg−1 BW) and the effect persisted for several weeks after the initial administration [43]. In an in-vitro study, hydrolysates of salmon pectoral fin by-products (digested by alcalase, flavourzyme, neutrase, pepsin, protamex and trypsin) that were isolated and fractionated to various peptide fractions by consecutive chromatography, demonstrated competitive, non-competitive and mixed inhibition modes against ACE in a dose-dependent manner with IC50 values of 7.72–10.77 μM [44]. For the extraction and hydrolysis of intended peptides, selection of enzymes and the subsequent fractionation and purification techniques are vital considerations, since these factors govern the structure of the final product and its mode of action in the target site (Table 5).
3. Impact of Enzymatic Digestion and Purification Processes on Antihypertension and ACE Inhibition Activity
The review of Lee et al., focusing on animal-, marine-, and plant-origin ACE inhibitors, have concluded that optimal hydrolysis treatment depends on the type of the protein source [23]. Optimal hydrolysis treatment for different food sources have been postulated as; animal-extracted enzymes (e.g., pepsin, trypsin, chymotrypsin) for animal products, and microbe-extracted enzymes (e.g., alcalase, neutrase, thermolysin) for marine organisms and plants [23]. In addition to enzymatic hydrolysis, solvent extraction has also efficaciously been unitized for production of plant-based ACE-inhibitors [24]. The proteolytic enzymes from different sources (from animals’ digestive tract, microbes and other natural sources), including autolysis in which naturally occurring enzymes are actively engaged in the fermentation process, have overall been used for the extraction of ACE-inhibiting peptides from marine-organisms [34]. In accordance, of the above-mentioned digestion processes, enzymatic hydrolysis has commonly been used in the preparation of fish-origin antihypertensive and anti-ACE hydrolysates, with the exception using water-extraction procedure which has shown to be an effective method for isolation of mushroom-based ACE-inhibitors (Table 5) [24,49].
Following digestion, in general, hydrolysates are further processed to obtain a purer, and likely, a more potent product. Next, products are subjected to filtration. This commonly is practiced for every type of food source. Subsequently, a chromatographic technique, or combination of few, would be used for separation of the peptides based on their many structural and compositional characteristics (Table 5). A clear example is that ACE-inhibition activity of jellyfish hydrolysate was enhanced by 8-fold (from IC50 of 1.28 mg mL−1 to the low end of 0.16 mg mL−1) by desalting and gel column chromatographing [33]. Ultra-filtering of grass carp lysate with 10 kDa molecular weight MWCO ultrafiltration (UF) membrane improved IC50 by about 3-fold (from 0.692 mg mL−1 to 0.272 mg mL−1) and further desalting by about 7-fold (from 0.692 mg mL−1 to 0.105 mg mL−1) [42]. Even so, precipitating and desalting, frequently applied to plant- and marine-based hydrolysates, is not so common for fish-hydrolysates [24,33,42]. Separation and purification of these molecules are well necessary for the characterization and identification of the biopeptides, in terms of amino acid sequences and molecular mass (Table 5).
3.1. Enzymatic Digestion
Depending on the amino acid composition and the peptide bonds of the selected sample, food source, many different enzymes could be used to produce the most preferred potent bioactive molecule(s). An enzymatic reaction, depending on the type of enzyme and the substrate, results in final products with diverse functionalities (Table 5 and Table 6).
3.1.1. Effect of Enzymes on Different Peptide Bonds
Alcalase and neutrase, both, produce peptides with hydrophobic amino acids at the C-terminus. Alcalase, an esterase, explicitly produces short peptides (usually < 10 kDa) with hydrophobic amino acids at the C-terminal, as well as heterocyclic amino esters, whereas, neutrase, neutral zinc metallo endo-protease, is mainly specific to leucine, tyrosine, tryptophan and phenylalanine [54,78,79,80]. On the other hand, papain with broad specificity cleaves and catalyzes peptide bonds of basic amino acids, leucine, methionine, tryptophan, glycine, tyrosine, phenylalanine and amino acid ethyl ester residues with a preference for hydrophobic amino acid large side-chains contrary to alcalase, which is evident in numerous studies [33,54,78,79,80,81]. The chymotrypsin, like papain, cleaves bulky side chains, rather non-polar amino acids, with valine, alanine, leucine, proline, tyrosine, phenylalanine, histidine, and tryptophan at C-terminal [57,60,63]. Flavourzyme, an exopeptidase with exocatalytic action, generates a great deal of amino acids, as well as long chain peptides with higher MW (>19 kDa in the hydrolysis of poultry meals and flaxseed) [82,83]. Proteinase K, a serine protease, cuts at the peptide bond site next to the carboxylic group of aliphatic, aromatic amino acids with a preference for hydrophobic amino acids comparable to alcalase and neutrase [54,84].
Due to improved enzymatic endo-actions and specific hydrolysis of carboxyl-terminal hydrophobic amino acids, alcalase has been preferred over other common commercial and non-commercial enzymes for the extraction of ACE inhibitory peptides specifically from grass carp and other fish types [42,44,45,85]. According to Lee et al. alcalase may not be suitable for the digestion of every fish type [57]. For the extraction of ACE inhibitors from dried bonito, over a broad range of enzymes, thermolysin was the most superior [60]. Consecutive digestion of the peptic hydrolysate of dried bonito by digestive proteases affected the effectivity of peptides, nonetheless, the same method had a non-significant effect on the additional digestion of thermolysin hydrolysate [60]. The trypsin/chymotrypsin and chymotrypsin increased the potency of the peptic hydrolysate by 19% (IC50 38 µg mL−1) and 13% (IC50 41 µg mL−1), respectively, nevertheless trypsin alone decreased this effect by 38% (IC50 65 µg mL−1) in comparison to the pepsin hydrolysate (IC50 47 µg mL−1) [60].
3.1.2. Effect of Enzymes on the Separation of Blood Pressure Lowering Peptides
In the study of Ghassem et al., the biological activity of the enzymatic hydrolysates, from snakehead fish, in decreasing order were; alcalase > proteinase k > flavourzyme > neutrase > papain in comparison to the crude sarcoplasmic snakehead fish muscle protein with the least and alcalase hydrolysate with highest activities [54]. A similar approach was followed by Wijesekara et al. where muscle protein of seaweed pipefish was digested through enzymatic hydrolysis by various commercial enzymes, and all the resultant hydrolysates exhibited anti-ACE effect, ~70–86%, in decreasing order of alcalase > trypsin > papain, pepsin > neutrase > pronase [55]. Bonito hydrolysates all affected the ACE activity to some degree, IC50 ranging from 29 µg mL−1 to 175 µg mL−1, in decreasing order of thermolysin > trypsin/chymotrypsin digested peptic hydrolysate > chymotrypsin digested peptic hydrolysate > pepsin > trypsin digested peptic hydrolysate > chymotrypsin > trypsin > trypsin/chymotrypsin [60]. Tuna fish frame proteins, when processed by specific and non-specific commercial enzymes, demonstrated ACE inhibition activity to varying degrees, ~45–88%, and the effects in decreasing order of activity were; pepsin > α-chymotrypsin > neutrase > alcalase > trypsin > papain [57].
3.2. Isolation and Purification
Moreover, of the above-discussed procedures and techniques, mainly membrane filtration and chromatographic techniques are in use for isolation and purification of anti-hypertensive and anti-ACE biopeptides from fish lysates (Table 5). While smaller MWCO ranges are applied for plant-based hydrolysates (1–10 kDa), a wider range of 1 kDa up to 30 kDa has been adapted for fish hydrolysates [24]. Comparable chromatographic techniques (e.g., size exclusion chromatography (SEC), ion (cat- and an-ion) exchange chromatography (IEC), solid phase extraction (SPE), reversed phase high performance liquid chromatography (RP-HPLC) are practiced for fish- and plant-based hydrolysates (Table 5) [24]. Derived products significantly vary from one another in terms of MW, chain length, structure and composition, which would be influential on the properties of the peptide and subsequently its bioactivity (Table 5, Table 6 and Table 7).
Table 7.
Structures of selected fish-extracted antihypertensive and ACE inhibitory oligopeptides.
Effect of Different Fractionation Processes on the Separation of Blood Pressure Lowering Peptides
In a study by Chen et al. fresh grass carp was primarily hydrolyzed by two commonly used enzymes, alcalase and neutrase. Subsequently, peptides were isolated by UF process, MWCO 10 kDa, and the resultant fraction was further desalted using mixed ion exchange resins [42]. Desalted permeate had significantly higher ACE inhibition activity (~2–6 times higher) than the permeate of UF, the initial hydrolysate and the reference protein, soya peptide [42]. Molecular weight distribution analyses of the desalted UF permeate fraction revealed that > 90% of the peptides were of sizes 190–1000 Da [42]. Amino acid composition analysis results showed that glutamic- and aspartic-acid (negatively charged hydrophobic amino acids) are the most dominant amino acids in the fraction and the abundance of amino acids follow similar trends in both the hydrolysate and the desalted protein [42].
In accordance, fish scale peptides of the same species that were hydrolyzed using neutral protease and isolated by macroporous resins resulted in hydrophobic ACE inhibitory di- to penta-peptides of sizes 145 Da and 650 Da [46]. Another ACE inhibitory tri-peptide from whole fresh grass carp with MW of < 3 kDa was hydrolyzed by alcalase, afterwards isolated and purified sequentially using UF, macroporous adsorption resin, and two steps of RP-HPLC [45]. Alcalase and protamex digested salmon hydrolysate at 1 mg mL−1 inhibited 56% and 50% of the ACE activity, respectively, whereas, others’ effect was below 23% [44]. Molecular mass distribution of salmon trimming hydrolysate showed that higher percentage of corolase-derived peptides, about 44%, were < 5 kDa in comparison to different enzyme preparations, and evidence has confirmed the improved bioactivity of smaller size peptides [50]. In general, alcalase and corolase release more of short proline-rich hydrophobic amino acids, also observed in the study of Neves et al., are well known for their anti-ACE attributes [50,86,87]. In agreement, salmon alcalase-digested hydrolysate was about 97% rich in peptides with MW of <10 kDa [50,86,87].
The fractionation of tilapia by chromatographic techniques, four-step purification, significantly affected the ACE inhibition activity of the different fractions obtained throughout the process [53]. The most potent ACE inhibitory peptide of the tilapia, IC50 of 0.15 mg mL−1, was obtained by SEC in a successive fractionation process [53]. Successive purification of tuna frame protein peptic hydrolysate by chromatographic techniques resulted in a semi-pure product (PHII-F3-3) which was ~20-times more potent in comparison to the crude hydrolysate [57]. One of the three fractions (F3) from alcalase protein hydrolysate of the seaweed pipefish using FPLC on DEAE FF anion exchange column was significantly active with IC50 of 0.068 mg mL−1 [55]. F3 was sub-fractionated by RP-HPLC on C18 column to three fractions with the IC50 values of 0.62 mg mL−1 and 1.44 mg mL−1 for F3-II and F3-III, respectively [55]. Evidently, the peptides in the hydrolysate have a synergistic effect as F3 is 9-times and 21-times more potent than its sub-fractions, the F3-II and F3-III, correspondingly [55]. Ultrafiltration, MWCO 5 kDa, non-significantly improved anti-ACE effect by 3% only (88% ACE inhibition with 0.14 mg peptide concentration) in comparison to the 85% inhibition of crude hydrolysate of tilapia (Oreochromis niloticus) by 0.51 mg of peptides, followed by DEAE-Sephacel anion exchange chromatography fraction (59% at 0.07 mg of peptides), CM-Sepharose cation exchange chromatography fraction (38% at 0.02 mg of peptides), and Superdex peptide 10/300 GL size exclusion chromatography fractions with least effectivity (35% at 0.01 mg of peptides) [53]. Use of diverse lysing, separating and purifying procedures lead to different cleavage sites, length and amino acid types of a peptide chain, consequently affecting the bioactivity of the final product (Table 5 and Table 6).
4. Structure-Activity Relationship
Scientists are in general agreement of a strong association between the ACE-inhibition and or anti-hypertensive effect of a biopeptide with its structure; size, chain length, type and order of amino acids in the sequence. Conclusively decades ago, Cheung et al. declared the existence of a hydrophobic amino acid at either of the peptides’ terminals vital for ACE-inhibition activity of a peptide, namely, phenylalanine, proline at C-terminal and isoleucine, valine at N-terminal of the biopeptide [88]. In agreement with Cheung et al., phenylalanine exists in a great deal of fish blood pressure controlling peptides, either at C- or N- terminal (18 sequences in total), of varying masses and lengths (Table 6 and Table 7). Even so surprisingly years later, Daskaya-Dikmen reviewing plant-derived ACE-inhibitors, though agreeing on the importance and relationship between ACE-inhibition activity and peptide structure, could not specifically conclude on the structural and compositional feature of a potent blood pressure lowering biopeptide [24].
Cushman et al. back in 1971, suggested the possible interaction of ACE active sites with tyrosine, tryptophan, proline, phenylalanine and one hydrophobic amino acid at the C-terminal residue to be responsible for the ACE inhibition activity of the potent peptides [87]. In agreement with the proposition of Cushman et al. amino acid sequences of the potent fractions of salmon by-product hydrolysate with hydrophobic amino acids at the C-terminal residue were Val-Trp-Asp-Pro-Pro-Lys-Phe-Asp, Phe-Glu-Asp-Tyr-Val-Pro-Leu-Ser-Cys-Phe, and Phe-Asn-Val-Pro-Leu-Tyr-Glu [44,87]. In general, salmon muscle dipeptides with phenylalanine in their structure exhibited the significant activity of which Ile-Phe, at 50 µM with 86% inhibition was the most compelling [51]. The dipeptide Ile-Phe has repeatedly been reported as an anti-hypertensive peptide of animal and plant source (e.g., chicken, pork, bovine, egg, soybean, etc.) (Table 3). Presence of same Ile-Phe in animal-, plant- and marine-origin oligopeptide sequences also have been shown, per AHTPDB database, to effectively prevent hypertension (available at http://crdd.osdd.net/raghava/ahtpdb/index.php). The dipeptides Val-Phe and Leu-Phe were the most abundant active peptides in the salmon muscle hydrolysate constituting 0.12% and 0.06% of the total content, respectively [51]. Comparable peptides have been frequently identified in sources other than fish with potent blood pressure lowing effects (Table 3).
The two isolated and identified penta- and hexa-peptides sequence of snakehead muscle sarcoplasmic protein alcalase hydrolysate, Leu-Tyr-Pro-Pro-Pro and Tyr-Ser-Met-Tyr-Pro-Pro, contained proline and one hydrophobic amino acid at the C-terminal which has earlier been reported to interact with ACE active sites hindering its activity [54,87]. Interaction of skate originated oligopeptides (Met-Val-Gly-Ser-Ala-Pro-Gly-Val-Leu and Leu-Gly-Pro-Leu-Gly-His-Gln) with ACE has been observed at Trp67, Asn68, Thr71, Asn72, Thr74, Glu76, Thr77, Lys338, Asp346 and Arg348 [67]. Looking closely at the pattern of snakehead-extracted anti-ACE pentapeptide (Leu-Tyr-Pro-Pro-Pro; IC50 1.3 μM) and cuttlefish-extracted anti-ACE pentapeptide (Val-Glu-Leu-Tyr-Pro; IC50 5.22 μM) presence of Leu-Tyr-Pro order of amino acids in both sequences is conspicuous (Table 7) [54,66]. Interestingly sequencing of the selected salmon trimmings corolase hydrolysate fractions revealed Tyr-Pro as the most potent ACE inhibitory dipeptide (IC50 5.21 μM) which is in accordance with the scheme of Cushman et al. [50,87]. A further up-close look at snakehead- and cuttlefish-extracted pentapeptides one could observe the occurrence of Tyr-Pro, the most active salmon-extracted anti-ACE dipeptide, in both of the anti-ACE biopeptides [50,54,66]. The sequences of sea broom hydrolysate based on IC50 values, 0.003–0.16 mg mL−1, in decreasing order were Val-Ile-Tyr > Val-Tyr > Gly-Tyr > Gly-Phe [56].
Presence of proline in the sequence of fish-extracted biopeptides may play more vital than MW, chain length and hydrophobicity. Cuttlefish-origin di-peptide, Gly-Ser (163.0 Da, hydrophobicity of +9.51 Kcal mol−1), was respectively ~ 220- and 60-times less potent than cuttlefish-origin Val-Glu-Leu-Tyr-Pro penta-peptide and Ser-Thr-His-Gly-Val-Trp hexa-peptide with comparable hydrophobicities (+9.25 and +9.54 Kcal mol−1) (Table 7) [66]. In accordance with proline richness and its number of occurrences in a sequence could greatly increase the potency of a biomolecule. Snakehead Leu-Tyr-Pro-Pro-Pro penta-peptide (IC50 1.3 μM), containing three prolines, was ~2-times more active than Tyr-Ser-Met-Tyr-Pro-Pro hexa-peptide (IC50 2.8 μM), containing two prolines, with similar properties (pI, charge and hydrophobicity) (Table 7). It shall be emphasized that scientists agree on the importance of proline moiety in the activity of synthetic ACE inhibitors [38]. However, comparing the two deca-peptides with relatively comparable properties, the occurrence of two phenylalanine (Phe-Glu-Asp-Tyr-Val-Pro-Leu-Ser-Cys-Phe), one at C- and another at N-terminus, appears to exert more effective anti-ACE effect (~ 3-times) than two successive prolines at C-terminus (Leu-Leu-Met-Leu-Asp-Asn-Asp-Leu-Pro-Pro) of a bioactive sequence (Table 7) [44,69].
Existence of three sequential proline residues in a sequence results in a polyproline structure of which the left-handed-polyproline-II helices are more common than their right-handed counterparts. Proline-rich polyproline peptides, especially left-handed structures, have been documented as indispensable for the presentation of antimicrobial, immunomodulation, antioxidant and other bioactivities [89]. Tri-proline peptides engage in protein-protein signaling interactions thus playing important in various cell signal transduction pathways [90]. Vitali et al. have reviewed proline-rich peptides as potential leads for research and development of pharmaceutical compounds [89]. Leu-Tyr-Pro-Pro-Pro may, in that sense, owe its potency to the polyproline structure. Factually, structure-wise, proline and hydroxyproline amino acids, are derivatives of pyrrolidine. Pyrrolidine moiety is common in drug discovery, and its ring structure can be found in many. More than a dozen FDA approved drugs are merely substituted pyrrolidine compounds [91,92]. Interestingly of all the fish anti-ACE and antihypertensive sequences, reviewed herein, there are sixteen peptides with proline at their C-terminal (none at N-terminal) and of those four of the sequences contain 2-3 successive prolines [50,54,69]. Of all, Leu-Tyr-Pro-Pro-Pro (1.3 μM, 585 Da) and Asp-Tyr-Gly-Leu-Tyr-Pro (62 mΜ, 349 Da) were the most and the least potent sequences, based on reported IC50 values, with more or less similar properties except MW and number of proline residues (Table 6 and Table 7) [54,60].
Presence of tyrosine (a polar amino acid) and phenylalanine at the C-terminus along with hydrophobic amino acids (valine and glycine) at the N-terminus may contribute to the activity of sea broom-origin di- and tri-peptides [56,87]. The most effective peptide, Val-Ile-Tyr, was 190-times more potent in comparison to the crude sea broom hydrolysate [56]. Existence of tyrosine among fish-isolated antihypertensive fractions is rather common, and fifteen of the isolated peptides happen to contain this amino acid at either of their extremities (mainly at C- and equally at N-terminal or penultimate C-terminal position). Among those fifteen peptides, Tyr-Ser-Met-Tyr-Pro-Pro (IC50 2.8 μM, 756 Da) and Gly-Tyr (IC50 265 μM, 238 Da) were the most and the least active ones [54,56].
Dipeptides containing tryptophan at the C-terminal are relatively more potent than their reversed sequence counterparts with tryptophan at the N-terminal [30,51]. Consistently dipeptides (Ile-Trp, Leu-Trp) with tryptophan at C-terminal could withstand the sequential digestive enzymes reaction (pepsin, trypsin, chymotrypsin) of 70% and 61%, respectively, in comparison to the crude salmon peptide with the stability of only 47% [51]. Dipeptides Val-Trp, Ile-Trp, Gly-Trp, Met-Trp, Ile-Trp, Leu-Trp (tryptophan at C-terminal position), Trp-Ala, Trp-Met (tryptophan at the N-terminal position), Phe-Leu, Leu-Phe, Val-Leu, Ile-Leu, Leu-Ile, Val-Phe, Ile-Phe, Tyr-Phe (hydrophobic amino acid at C-terminal position) Phe-Tyr (tyrosine at C-terminal position) along with tripeptides Val-Ile-Phe, Ile-Val-Phe, Phe-Val-Leu, Ala-Phe-Leu, Leu-Val-Leu, Val-Ile-Leu, Ile-Val-Leu, Phe-Ile-Ala, Tyr-Leu-Val, and Ile-Val-Trp (hydrophobic amino acid at C-terminal position) extracted and isolated from salmon and its by-products have been found to be active ACE inhibitors [51]. Most effective salmon muscle peptides were Ile-Val-Trp and Ile-Trp which exhibited 35% and 49% ACE inhibition, at a concentration of 1.0 mM and 1.0 μM respectively in comparison to the crude hydrolysate with IC50 of 79 µg mL−1 [51].
Two of the tested synthetic tilapia-origin heptapeptides, Met-Ile-Leu-Leu-Leu-Phe-Arg and Leu-Asn-Leu-Gln-Asp-Phe-Arg contained Phe-Arg, which is a sequence previously reported for its prominent ACE inhibitory effect by BIOPEP database (available at http://www.uwm.edu.pl/biochemia/index.php/en/biopep) [53]. Tilapia-extracted hepta-peptides with C-terminal Phe-Arg clusters and parallel MW were effective at relatively low doses; however, Met-Ile-Leu-Leu-Leu-Phe-Arg with a net charge of +1 was ~ 7-times more potent than Leu-Asn-Leu-Gln-Asp-Phe-Arg with a net charge of 0. In addition, their hydrophobicity significantly differed from one another. Met-Ile-Leu-Leu-Leu-Phe-Arg was less hydrophobic (+2.46 Kcal mol−1) than Leu-Asn-Leu-Gln-Asp-Phe-Arg (+10.76 Kcal mol−1) (Table 7). Looking through AHTPDB database, regardless of peptide extraction source, the occurrence of Phe-Arg cluster is more common at N-terminal of bioactive sequences than C-terminal and or penultimate C-terminal position.
According to Cheung et al., the presence of hydrophobic amino acids, methionine and leucine, at their N-terminal may have correspondingly affected their anti-ACE activity [53,88]. Commonly, positively charged amino acids could contribute to the prevention of ACE activity, hence, l-arginine alone or in combination with lisinopril, an ACE inhibiting drug, has shown antihypertensive effects in addition to its anti-inflammatory attributes [93,94,95,96]. In accordance, no activity was detected when arginine was cut off from the active ACE inhibitor sequence of Met-Ile-Leu-Leu-Leu-Phe-Arg [53]. Slashing off arginine from synthesized peptide sequence (Leu-Asn-Leu-Gln-Asp-Phe-Arg; IC50 0.85 µM, Leu-Asn-Leu-Gln-Asp-Phe; IC50 0.51 µM) improves the anti-ACE activity by 67% as phenylalanine, a hydrophobic amino acid, is then situated at the C-terminal position, which has been repeatedly reported as one of the criteria for an ACE inhibitor peptide [44,53,87]. Arginine was positioned at C-terminal of 70% of the ten identified amino acid sequences of a selected tilapia ACE inhibitory fraction (resulted from SEC in a succeeding fractionation method with masses ranging from 791 Da to 1526 Da) with in-vitro thermo-, pH- and digestive proteases- stability [53]. Arginine is a positively charged amino acid, however, its role in stability, when situated at C-terminus, to digestive enzymes have not thoroughly been elucidated. Quirós et al. modified a hexapeptide of mammal-origin β-casein, Leu-His-Leu-Pro-Leu-Pro, for better understanding of the structure activity relationship and found out that substitution of proline at C-terminus by arginine and leucine in the penultimate position by glycine not only improved its antihypertensive activity by double, but also enhanced its stability to gastrointestinal enzymes, respectively [97]. To the author’s knowledge, a comparable study on a fish-origin peptide has not yet been directed. In this review, there are eight C-terminal and one N-terminal arginine fish-derived peptides (Table 6 and Table 7).
The MW of the two hepta-peptides isolated and identified from tilapia hydrolysate are insignificantly different from one another, however, Met-Ile-Leu-Leu-Leu-Phe-Arg (905.83 Da) with Leu-Phe, Leu-Leu-Phe and Phe-Arg ACE inhibitory peptide sequences in its chain was 7-times more effective than the other (Leu-Asn-Leu-Gln-Asp-Phe-Arg; 905.66 Da) indicative of the fact that a combination of factors (size, sequence and digestive stability) are required for the bioactivity and potency of a peptide [53]. Another pentapeptide, Leu-Lys-Pro-Asn-Met, from dried bonito with proline situated inside the sequence could only minimally resist the same digestion, 11%, while a sardine dipeptide which has also been isolated from scales of sea bream, Val-Tyr, without proline in its sequence exhibited higher stability, 32%, due to its small size [51,56]. ACE inhibition, however, may be of a synergetic type effect as with tilapia hydrolysate derived fractions that of all only few of the synthesized peptide sequences hindered the enzyme’s activity [53].
MW is not always indicative of biological activity, since purified PHII-F3-3 is a relatively heavy peptide, 21- amino acids chain and MW > 2 kDa, thus, has shown perceptible in-vitro and in-vivo anti-hypertensive effects against high blood pressure, non-competitively binding to non-active sites of ACE [57]. One of the final four resultant fractions of SEC, the final stage of successive fractionation, that was the most biologically active ACE inhibitory peptide with IC50 of 0.15 mg mL−1 was subjected to amino acid sequence analysis [53]. The MW of the sequences varied from 677 Da up to 1526 Da [53]. Lowest MW did not necessarily denote the highest potency of the peptide, since most potent sequence was Met-Ile-Leu-Leu-Leu-Phe-Arg with MW of 905 Da and IC50 of 0.12 µM [53]. Contrary to the study of Toopcham et al., MW of the identified hexapeptides, from alcalase hydrolysate of the seaweed pipefish, was characteristic of their biological activity, thus, the smallest peptide, F3-II with MW of 744 Da, was non-cytotoxically more potent than F3-III with MW of 917 Da [53,55]. Identified hexapeptides of alcalase peptidic hydrolysate of the seaweed pipefish have MW of < 1000 Da with hydrophobic (Thr-Phe-Pro-His-Gly-Pro) and positively charged (His-Trp-Thr-Thr-Gln-Arg) amino acids at C-terminal of the sequence which is in line with previous studies [53,55,93,94,95,96].
5. Stability of ACE-Inhibiting Fish-Derived Peptides to Gastrointestinal Digestion
Small size peptides (mainly with two and up to six amino acids in the chain) cannot be substrates to any digestive proteases; therefore, they remain intact throughout gastrointestinal digestion. ACE inhibitory peptides must be absorbed from the intestine and reach the cardiovascular system in their original shape to hinder the activity of ACE [36]. Digestion stability of anti-ACE peptides to digestive enzymes were 95% for Ile-Trp, 100% for Leu-Trp, and 87% for salmon peptidic hydrolysate [51]. The effectivity of the two identified penta- and hexa-peptides of alcalase snakehead fish muscle sarcoplasmic protein hydrolysate (Leu-Tyr-Pro-Pro-Pro and Tyr-Ser-Met-Tyr-Pro-Pro) was tested in an in-vitro gastrointestinal (pepsin, pancreatin) and gastrointestinal + mucosal (lower intestinal peptidase) digestion model [54]. ACE inhibition, due to the presence of proline in the sequences was not affected by gastrointestinal digestion; however, gastrointestinal + mucosal digestion had non-significant negative effect [54,66]. Comparable effects were exhibited by a pH- and heat-stable penta-peptide (Val-Glu-Leu-Tyr-Pro) of cuttlefish hydrolysate showing resistance to the digestion of pepsin, trypsin, chymotrypsin or combination of all (IC50 5.25–5.56 μM) [66].
Purified PHII-F3-3 of tuna-origin has not been tested for its resistance to digestive proteases, however, since Pro is situated at the C-terminus of the chain, therefore, this polypeptide might be well stable to the gastrointestinal digestion similar to the isolated penta- and hexa-peptide of snakehead’s muscle sarcoplasmic protein, Leu-Tyr-Pro-Pro-Pro, Tyr-Ser-Met-Tyr-Pro-Pro, etc. [54,57,66]. Val-Ala-Pro sequence of grass carp protein hydrolysate, with proline at the C-terminal, was also another example of a sequence that could resist pepsin and chymotrypsin, in an in-vitro digestive system model [45]. To improve the structure-activity relationship knowledge, identified peptides of dried bonito thermolysin hydrolysate and their fragments were synthesized slashing off amino acids from the C-terminal/N-terminal and or split in two fragmented products with the final fragment of the di- and tri-peptides by Yokoyama et al. [60]. The majority of the fragments were few to few hundred times less effective in comparison to the parent ACE inhibiting peptides [60]. Generally, fragmenting and splitting the synthesized parent peptide to di- and or tri-peptide improved the activity (Table 8) [60].
Table 8.
ACE inhibition activity of synthesized dried bonito thermolysin hydrolysate peptides and their fragments.
Gastrointestinal proteases’ (pepsin and pancreatin) digestion of cobia head protein hydrolysate surprisingly improved the activity of the fraction, MW < 3000 Da (67% abundant with peptides of molecular size 173–1749 Da), when hydrolyzed with pepsin and sequential pepsin/pancreatin digestion by 60% and 41%, respectively, in comparison to the pre-digestion condition (IC50 0.24 mg mL−1), respectively [65]. The same fraction also resisted ACE hydrolysis (based on non-significant deviations of IC50 before and after pre-incubation with ACE) proving to be of an inhibitor type peptide [65]. Val-Ala-Pro sequence resisted ACE hydrolysis and proved to be of another inhibitor type drug rather than pro-drug type one [45]. Bioactive ACE inhibitor/antihypertensive dried bonito thermolysin hydrolysate peptides could be of inhibitor type or prodrug type, since when preincubated with ACE, IC50 values were significantly affected (Table 9) [77].
Table 9.
ACE inhibition and antihypertensive effects of dried bonito thermolysin hydrolysate peptides.
6. Conclusions and Possible Future Trends
According to the results of studies, reviewed herein, the effectivity of the final biopeptide product is greatly affected by the extraction processes. The enzymatic hydrolysis (selection of enzymes, substrate: Enzyme ratio, degree of hydrolysis and process time), as well as isolation/purification techniques, are equally important for the production of the fish biopeptides with anti-ACE and anti-hypertensive potency. In this context, though practically not applicable to all fish types, such as bonito or tuna, alcalase commonly have been the enzyme of choice for many. Other types of enzymes are as well in common use for the efficient digestion of fish and fish-related products. For example, more potent biopeptides resulted from thermolysin and pepsin digestion of certain types of fish than alcalase digestion.
Fish derived biopeptides whether from whole, byproducts and or even processed catch could be a natural source of ACE inhibitors/pro-drug type and antihypertensive peptides with no reported cytotoxicity. Accordingly, there is amply amount of data supporting the fact that these peptides could replace conventional synthetic drugs with similar potency and little to no adverse effects. These peptides are of various sizes and lengths; however, generally smaller MW peptides with only a few amino acids in their sequence have shown higher bioefficacy and bioavailability in comparison to the heavier MW polypeptides. Small biopeptides, in terms of MW and chain length, could better resist the digestive proteases and reach the target tissue intact to exhibit their health effect. In addition to size and chain length, certain factors in the structure, especially C-terminal, of a fish peptide are critical for ACE inhibition and antihypertensive effects. Presence of tyrosine, tryptophan, proline, phenylalanine and or positively charged amino acids next to hydrophobic amino acids would make the interaction with active sites of ACE (competitive mode of inhibition) and non-active sites of ACE (non-competitive mode of inhibition) possible. Another important feature could be the presence of proline at C-terminal so that the peptide could resist the gastrointestinal enzymes, pepsin, trypsin, and chymotrypsin for the reasons mentioned earlier. Many of the herein reviewed fish antihypertensive oligopeptides, especially di- and tri-peptides, may be suitable drug candidates considering their adsorption, distribution, metabolism, and excretion (ADME) characteristics. Drug-likeness of selected tri-, tetra- and penta-peptides, listed in Table 10, was predicted according to Lipinski’s rule of five using SwissADME web tool. However, it is noteworthy that Lipinski’s rules, mainly appropriate for oral dosage forms, are only prediction measures and many successful inhibitor drugs have shown to violate at least two of the rules. This may be the case for Leu-Lys-Pro-Asn-Met, a Bonito-extracted penta-peptide, violating two of the Lipinski’s rules yet being an effective inhibitor, marketed for controlling high blood pressure in tablet and capsule dosage forms (Table 10).
Table 10.
Drug-likeness and ADME (adsorption, distribution, metabolism, and excretion) analysis of selected anti-ACE and antihypertensive fish biopeptides by Lipinski’s rules.
In the light of this evidence, high blood pressure could be managed and treated by fish biopeptides effectively. Aforementioned health effects could not be solitary specific to grass carp, salmon, sardine or tilapia; hence, many other fish types may have similar effects, yet more research is warranted for a concrete supposition. However, successful discovery and design of natural ACE inhibitors necessitate more structure-function analysis of the blood pressure lowering fish-biopeptides. Examining the feasibility of industrial scale-up production ought to go hand in hand with the biopeptides’ incorporation into commercial products. Interestingly some of the anti-ACE fish-biopeptides have made their way through the nutraceutical and pharmaceutical industries and are the active ingredients of numerous natural health products (Table 11). These products are already marketed in North America, Europe and Asia in various forms (Table 11). To conclude, bioactive peptides of many fish sources could be good candidates for the development of natural health foodstuffs, nutraceutical and pharmaceutical products, however, more human studies and clinical trials are needed to better evaluate their bioefficacy and bioavailability in the target site and the subject. It is also of note that only about one third (12 out of 37) of the herein reviewed research articles utilized fishery byproducts, mainly frame, skin and scale, as substrates and starting raw material for the extraction of biologically active peptides. Nevertheless, it is expected that these and similar research projects make efficient use of the process byproducts and underutilized species in order to partially contribute to the targets of food security and greenhouse gas emission strategies.
Table 11.
Antihypertensive fish peptides as active ingredients in commercial products.
Funding
This work received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef]
- Isomaa, B.; Almgren, P.; Tuomi, T.; Forsén, B.; Lahti, K.; Nissén, M.; Taskinen, M.-R.; Groop, L. Cardiovascular morbidity and mortality associated with the metabolic syndrome. Diabetes Care 2001, 24, 683–689. [Google Scholar] [CrossRef]
- Gomes, M.D.B. Impact of diabetes on cardiovascular disease: An update. Int. J. Hypertens. 2013, 2013, 653789. [Google Scholar]
- Kelli, H.M.; Kassas, I.; Lattouf, O.M. Cardio metabolic syndrome: A global epidemic. J. Diabetes Metab. 2015, 6, 2. [Google Scholar] [CrossRef]
- Deitel, M. Overweight and obesity worldwide now estimated to involve 1.7 billion people. Obes. Surg. 2003, 13, 329–330. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N. Engl. J. Med. 2014, 371, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Scholze, J.; Alegria, E.; Ferri, C.; Langham, S.; Stevens, W.; Jeffries, D.; Uhl-Hochgraeber, K. Epidemiological and economic burden of metabolic syndrome and its consequences in patients with hypertension in Germany, Spain and Italy; a prevalence-based model. BMC Public Health 2010, 10, 529. [Google Scholar] [CrossRef]
- Lind, L.; Berne, C.; Lithell, H. Prevalence of insulin resistance in essential hypertension. J. Hypertens. 1995, 13, 1457–1462. [Google Scholar] [CrossRef]
- Castro, J.P.; El-Atat, F.A.; McFarlane, S.I.; Aneja, A.; Sowers, J.R. Cardiometabolic syndrome: Pathophysiology and treatment. Curr. Hypertens. Rep. 2003, 5, 393–401. [Google Scholar] [CrossRef]
- Murray, C.J.; Lopez, A.D. Global mortality, disability, and the contribution of risk factors: Global Burden of Disease Study. Lancet 1997, 349, 1436–1442. [Google Scholar] [CrossRef]
- Giles, T.D.; Berk, B.C.; Black, H.R.; Cohn, J.N.; Kostis, J.B.; Izzo, J.L.; Weber, M.A. Expanding the definition and classification of hypertension. J. Clin. Hypertens. 2005, 7, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Angeli, F.; Reboldi, G.; Verdecchia, P. Hypertension, inflammation and atrial fibrillation. J. Hypertens. 2014, 32, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Bautista, L. Inflammation, endothelial dysfunction, and the risk of high blood pressure: Epidemiologic and biological evidence. J. Hum. Hypertens. 2003, 17, 223. [Google Scholar] [CrossRef] [PubMed]
- Chockalingam, A.; Campbell, N.R.; Fodor, J.G. Worldwide epidemic of hypertension. Can. J. Cardiol. 2006, 22, 553–555. [Google Scholar] [CrossRef]
- Harrison, D.G.; Guzik, T.J.; Lob, H.E.; Madhur, M.S.; Marvar, P.J.; Thabet, S.R.; Vinh, A.; Weyand, C.M. Inflammation, immunity, and hypertension. Hypertension 2011, 57, 132–140. [Google Scholar] [CrossRef]
- Berry, C.; Clark, A. Catabolism in chronic heart failure. Eur. Heart J. 2000, 21, 521–532. [Google Scholar] [CrossRef]
- Maksimowicz-McKinnon, K.; Bhatt, D.L.; Calabrese, L.H. Recent advances in vascular inflammation: C-reactive protein and other inflammatory biomarkers. Curr. Opin. Rheumatol. 2004, 16, 18–24. [Google Scholar] [CrossRef]
- Cardin, S.; Li, D.; Thorin-Trescases, N.; Leung, T.-K.; Thorin, E.; Nattel, S. Evolution of the atrial fibrillation substrate in experimental congestive heart failure: Angiotensin-dependent and-independent pathways. Cardiovasc. Res. 2003, 60, 315–325. [Google Scholar] [CrossRef]
- Nguyen, Q.; Dominguez, J.; Nguyen, L.; Gullapalli, N. Hypertension management: An update. Am. Health Drug Benefits 2010, 3, 47. [Google Scholar]
- Chobanian, A.V.; Bakris, G.L.; Black, H.R.; Cushman, W.C.; Green, L.A.; Izzo, J.L., Jr.; Jones, D.W.; Materson, B.J.; Oparil, S.; Wright, J.T., Jr. The seventh report of the joint national committee on prevention, detection, evaluation, and treatment of high blood pressure: The JNC 7 report. JAMA J. Am. Med Assoc. 2003, 289, 2560–2571. [Google Scholar] [CrossRef]
- De Miguel, C.; Rudemiller, N.P.; Abais, J.M.; Mattson, D.L. Inflammation and hypertension: New understandings and potential therapeutic targets. Curr. Hypertens. Rep. 2015, 17, 507. [Google Scholar] [CrossRef] [PubMed]
- Drapala, A.; Sikora, M.; Ufnal, M. Statins, the renin–angiotensin–aldosterone system and hypertension–a tale of another beneficial effect of statins. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hur, S.J. Antihypertensive peptides from animal products, marine organisms, and plants. Food Chem. 2017, 228, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-converting enzyme (ACE)-inhibitory peptides from plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef]
- Norris, R.; FitzGerald, R.J. Antihypertensive peptides from food proteins. In Bioactive Food Peptides in Health and Disease; InTech Publishers: London, UK, 2013; pp. 45–72. [Google Scholar]
- Aluko, R.E. Antihypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef]
- Martínez-Maqueda, D.; Miralles, B.; Recio, I.; Hernández-Ledesma, B. Antihypertensive peptides from food proteins: A review. Food Funct. 2012, 3, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, J.; Malini, M.; Joshy, V.A. A critical review on food protein derived antihypertensive peptides. Drug Invent. Today 2019, 12, 474–479. [Google Scholar]
- Hernández-Ledesma, B.; del Mar Contreras, M.; Recio, I. Antihypertensive peptides: Production, bioavailability and incorporation into foods. Adv. Colloid Interface Sci. 2011, 165, 23–35. [Google Scholar] [CrossRef]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Inhibition properties of dipeptides from salmon muscle hydrolysate on angiotensin I-converting enzyme. Int. J. Food Sci. Technol. 2006, 41, 383–386. [Google Scholar] [CrossRef]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Isolation of Peptides with Angiotensin I-converting Enzyme Inhibitory Effect Derived from Hydrolysate of Upstream Chum Salmon Muscle. J. Food Sci. 2003, 68, 1611–1614. [Google Scholar] [CrossRef]
- Fang, L.; Geng, M.; Liu, C.; Wang, J.; Min, W.; Liu, J. Structural and molecular basis of angiotensin-converting enzyme by computational modeling: Insights into the mechanisms of different inhibitors. PLoS ONE 2019, 14, e0215609. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Jia, A.; Zhang, Y.; Zhu, H.; Zhang, C.; Sun, Z.; Liu, C. Purification and characterization of angiotensin I converting enzyme inhibitory peptides from jellyfish Rhopilema esculentum. Food Res. Int. 2013, 50, 339–343. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, A.; Sharma, R.; Baruwa, A. Pharmacological review on natural ACE inhibitors. Der. Pharm. Lett. 2010, 2, 273–293. [Google Scholar]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-originating ACE inhibitors, including antihypertensive peptides, as preventive food components in blood pressure reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Attique, S.A.; Hassan, M.; Usman, M.; Atif, R.M.; Mahboob, S.; Al-Ghanim, K.A.; Bilal, M.; Nawaz, M.Z. A Molecular Docking Approach to Evaluate the Pharmacological Properties of Natural and Synthetic Treatment Candidates for Use against Hypertension. Int. J. Environ. Res. Public Health 2019, 16, 923. [Google Scholar] [CrossRef]
- Bhuyan, B.J.; Mugesh, G. Synthesis, characterization and antioxidant activity of angiotensin converting enzyme inhibitors. Org. Biomol. Chem. 2011, 9, 1356–1365. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Kang, K.-H.; Ryu, B.; Vo, T.-S.; Jung, W.-K.; Byun, H.-G.; Kim, S.-K. Angiotensin-I converting enzyme inhibitory peptides from antihypertensive skate (Okamejei kenojei) skin gelatin hydrolysate in spontaneously hypertensive rats. Food Chem. 2015, 174, 37–43. [Google Scholar] [CrossRef]
- Izzo, J.; Joseph, L.; Weir, M.R. Angiotensin-converting enzyme inhibitors. J. Clin. Hypertens. 2011, 13, 667–675. [Google Scholar] [CrossRef]
- Elliott, W.J.; Meyer, P.M. Incident diabetes in clinical trials of antihypertensive drugs: A network meta-analysis. Lancet 2007, 369, 201–207. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Xia, W.; Xiong, Y.L.; Ye, R.; Wang, H. Grass carp peptides hydrolysed by the combination of Alcalase and Neutrase: Angiotensin-I converting enzyme (ACE) inhibitory activity, antioxidant activities and physicochemical profiles. Int. J. Food Sci. Technol. 2016, 51, 499–508. [Google Scholar] [CrossRef]
- Wang, S.; Lin, L.-M.; Wu, Y.-N.; Fang, M.; Yu, Y.-Q.; Zhou, J.; Gong, Z.-Y. Angiotensin I Converting Enzyme (ACE) inhibitory activity and antihypertensive effects of grass carp peptides. Food Sci. Biotechnol. 2014, 23, 1661–1666. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Jeon, Y.-J.; Kim, Y.-T.; Je, J.-Y. Angiotensin I converting enzyme (ACE) inhibitory peptides from salmon byproduct protein hydrolysate by Alcalase hydrolysis. Process. Biochem. 2012, 47, 2240–2245. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Zhong, Q.; Wu, Y.; Xia, W. Purification and characterization of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide derived from enzymatic hydrolysate of grass carp protein. Peptides 2012, 33, 52–58. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Z.; Xu, S. Macroporous resin purification of grass carp fish (Ctenopharyngodon idella) scale peptides with in vitro angiotensin-I converting enzyme (ACE) inhibitory ability. Food Chem. 2009, 117, 387–392. [Google Scholar] [CrossRef]
- Borawska, J.; Darewicz, M.; Vegarud, G.; Iwaniak, A.; Minkiewicz, P. Ex vivo digestion of carp muscle tissue–ACE inhibitory and antioxidant activities of the obtained hydrolysates. Food Funct. 2015, 6, 210–217. [Google Scholar] [CrossRef]
- Nakajima, K.; Yoshie-Stark, Y.; Ogushi, M. Comparison of ACE inhibitory and DPPH radical scavenging activities of fish muscle hydrolysates. Food Chem. 2009, 114, 844–851. [Google Scholar] [CrossRef]
- Itou, K.; Nagahashi, R.; Saitou, M.; Akahane, Y. Antihypertensive effect of narezushi, a fermented mackerel product, on spontaneously hypertensive rats. Fish. Sci. 2007, 73, 1344–1352. [Google Scholar]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; FitzGerald, R.J. Bioactive peptides from Atlantic salmon (Salmo salar) with angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory, and antioxidant activities. Food Chem. 2017, 218, 396–405. [Google Scholar] [CrossRef]
- Enari, H.; Takahashi, Y.; Kawarasaki, M.; Tada, M.; Tatsuta, K. Identification of angiotensin I-converting enzyme inhibitory peptides derived from salmon muscle and their antihypertensive effect. Fish. Sci. 2008, 74, 911–920. [Google Scholar] [CrossRef]
- Kawasaki, T.; Seki, E.; Osajima, K.; Yoshida, M.; Asada, K.; Matsui, T.; Osajima, Y. Antihypertensive effect of valyl-tyrosine, a short chain peptide derived from sardine muscle hydrolyzate, on mild hypertensive subjects. J. Human Hypertens. 2000, 14, 519. [Google Scholar] [CrossRef]
- Toopcham, T.; Roytrakul, S.; Yongsawatdigul, J. Characterization and identification of angiotensin I-converting enzyme (ACE) inhibitory peptides derived from tilapia using Virgibacillus halodenitrificans SK1-3-7 proteinases. J. Funct. Foods 2015, 14, 435–444. [Google Scholar] [CrossRef]
- Ghassem, M.; Babji, A.S.; Said, M.; Mahmoodani, F.; Arihara, K. Angiotensin I–converting enzyme inhibitory peptides from snakehead fish sarcoplasmic protein hydrolysate. J. Food Biochem. 2014, 38, 140–149. [Google Scholar] [CrossRef]
- Wijesekara, I.; Qian, Z.-J.; Ryu, B.; Ngo, D.-H.; Kim, S.-K. Purification and identification of antihypertensive peptides from seaweed pipefish (Syngnathus schlegeli) muscle protein hydrolysate. Food Res. Int. 2011, 44, 703–707. [Google Scholar] [CrossRef]
- Fahmi, A.; Morimura, S.; Guo, H.-C.; Shigematsu, T.; Kida, K.; Uemura, Y. Production of angiotensin I converting enzyme inhibitory peptides from sea bream scales. Process. Biochem. 2004, 39, 1195–1200. [Google Scholar] [CrossRef]
- Lee, S.-H.; Qian, Z.-J.; Kim, S.-K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2010, 118, 96–102. [Google Scholar] [CrossRef]
- Matsui, T.; Matsufuji, H.; Seki, E.; Osajima, K.; Nakashima, M.; Osajima, Y. Inhibition of angiotensin I-converting enzyme by Bacillus licheniformis alkaline protease hydrolyzates derived from sardine muscle. Biosci. Biotechnol. Biochem. 1993, 57, 922–925. [Google Scholar] [CrossRef]
- Otani, L.; Ninomiya, T.; Murakami, M.; Osajima, K.; Kato, H.; Murakami, T. Sardine peptide with angiotensin I-converting enzyme inhibitory activity improves glucose tolerance in stroke-prone spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2009, 73, 2203–2209. [Google Scholar] [CrossRef]
- Yokoyama, K.; Chiba, H.; Yoshikawa, M. Peptide inhibitors for angiotensin i-converting enzyme from thermolysin digest of dried bonitot. Biosci. Biotechnol. Biochem. 1992, 56, 1541–1545. [Google Scholar] [CrossRef]
- Ktari, N.; Nasri, R.; Mnafgui, K.; Hamden, K.; Belguith, O.; Boudaouara, T.; El Feki, A.; Nasri, M. Antioxidative and ACE inhibitory activities of protein hydrolysates from zebra blenny (Salaria basilisca) in alloxan-induced diabetic rats. Process. Biochem. 2014, 49, 890–897. [Google Scholar] [CrossRef]
- Lassoued, I.; Mora, L.; Nasri, R.; Aydi, M.; Toldrá, F.; Aristoy, M.-C.; Barkia, A.; Nasri, M. Characterization, antioxidative and ACE inhibitory properties of hydrolysates obtained from thornback ray (Raja clavata) muscle. J. Proteom. 2015, 128, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.-J.; Je, J.-Y.; Kim, S.-K. Antihypertensive effect of angiotensin I converting enzyme-inhibitory peptide from hydrolysates of bigeye tuna dark muscle, Thunnus obesus. J. Agric. Food Chem. 2007, 55, 8398–8403. [Google Scholar] [CrossRef]
- Jung, W.-K.; Mendis, E.; Je, J.-Y.; Park, P.-J.; Son, B.W.; Kim, H.C.; Choi, Y.K.; Kim, S.-K. Angiotensin I-converting enzyme inhibitory peptide from yellowfin sole (Limanda aspera) frame protein and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2006, 94, 26–32. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, Y.; Hong, P.; Cao, W. Angiotensin I converting enzyme inhibitory activity and antihypertensive effect in spontaneously hypertensive rats of cobia (Rachycentron canadum) head papain hydrolysate. Food Sci. Technol. Int. 2013, 19, 209–215. [Google Scholar] [CrossRef]
- Balti, R.; Bougatef, A.; Sila, A.; Guillochon, D.; Dhulster, P.; Nedjar-Arroume, N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chem. 2015, 170, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Ryu, B.; Kim, S.-K. Active peptides from skate (Okamejei kenojei) skin gelatin diminish angiotensin-I converting enzyme activity and intracellular free radical-mediated oxidation. Food Chem. 2014, 143, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Je, J.-Y.; Park, P.-J.; Kwon, J.Y.; Kim, S.-K. A novel angiotensin I converting enzyme inhibitory peptide from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. J. Agric. Food Chem. 2004, 52, 7842–7845. [Google Scholar] [CrossRef]
- Himaya, S.; Ngo, D.-H.; Ryu, B.; Kim, S.-K. An active peptide purified from gastrointestinal enzyme hydrolysate of Pacific cod skin gelatin attenuates angiotensin-1 converting enzyme (ACE) activity and cellular oxidative stress. Food Chem. 2012, 132, 1872–1882. [Google Scholar] [CrossRef]
- Khantaphant, S.; Benjakul, S.; Kishimura, H. Antioxidative and ACE inhibitory activities of protein hydrolysates from the muscle of brownstripe red snapper prepared using pyloric caeca and commercial proteases. Process. Biochem. 2011, 46, 318–327. [Google Scholar] [CrossRef]
- Raghavan, S.; Kristinsson, H.G. ACE-inhibitory activity of tilapia protein hydrolysates. Food Chem. 2009, 117, 582–588. [Google Scholar] [CrossRef]
- Auwal, S.M.; Abidin, N.Z.; Zarei, M.; Tan, C.P.; Saari, N. Identification, structure-activity relationship and in silico molecular docking analyses of five novel angiotensin I-converting enzyme (ACE)-inhibitory peptides from stone fish (Actinopyga lecanora) hydrolysates. PLoS ONE 2019, 14, e0197644. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Vo, T.-S.; Ryu, B.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitory peptides from Pacific cod skin gelatin using ultrafiltration membranes. Process. Biochem. 2016, 51, 1622–1628. [Google Scholar] [CrossRef]
- Erkkilä, A.T.; Schwab, U.S.; de Mello, V.D.; Lappalainen, T.; Mussalo, H.; Lehto, S.; Kemi, V.; Lamberg-Allardt, C.; Uusitupa, M.I. Effects of fatty and lean fish intake on blood pressure in subjects with coronary heart disease using multiple medications. Eur. J. Nutr. 2008, 47, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Yahia, D.A.; Madani, S.; Prost, J.; Bouchenak, M.; Belleville, J. Fish protein improves blood pressure but alters HDL 2 and HDL 3 composition and tissue lipoprotein lipase activities in spontaneously hypertensive rats. Eur. J. Nutr. 2005, 44, 10–17. [Google Scholar] [CrossRef]
- Ait-Yahia, D.; Madani, S.; Savelli, J.-L.; Prost, J.; Bouchenak, M.; Belleville, J. Dietary fish protein lowers blood pressure and alters tissue polyunsaturated fatty acid composition in spontaneously hypertensive rats. Nutrition 2003, 19, 342–346. [Google Scholar] [CrossRef]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE-inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar] [CrossRef]
- Xu, W.; Kong, B.-H.; Zhao, X.-H. Optimization of some conditions of Neutrase-catalyzed plastein reaction to mediate ACE-inhibitory activity in vitro of casein hydrolysate prepared by Neutrase. J. Food Sci. Technol. 2014, 51, 276–284. [Google Scholar] [CrossRef]
- Li, G.H.; Wan, J.Z.; Le, G.W.; Shi, Y.H. Novel angiotensin I-converting enzyme inhibitory peptides isolated from Alcalase hydrolysate of mung bean protein. J. Peptide Sci. Off. Publ. Eur. Pept. Soc. 2006, 12, 509–514. [Google Scholar] [CrossRef]
- Mahmoodani, F.; Ghassem, M.; Babji, A.S.; Yusop, S.M.; Khosrokhavar, R. ACE inhibitory activity of pangasius catfish (Pangasius sutchi) skin and bone gelatin hydrolysate. J. Food Science Technol. 2014, 51, 1847–1856. [Google Scholar] [CrossRef]
- Schwab, L.W.; Kloosterman, W.M.; Konieczny, J.; Loos, K. Papain catalyzed (co) oligomerization of α-amino acids. Polymers 2012, 4, 710–740. [Google Scholar] [CrossRef]
- Nchienzia, H.; Morawicki, R.; Gadang, V. Enzymatic hydrolysis of poultry meal with endo-and exopeptidases. Poult. Sci. 2010, 89, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Karamać, M.; Kosińska-Cagnazzo, A.; Kulczyk, A. Use of different proteases to obtain flaxseed protein hydrolysates with antioxidant activity. Int. J. Mol. Sci. 2016, 17, 1027. [Google Scholar] [CrossRef] [PubMed]
- Kay, J. Specificity of Proteolysis; Wiley Online Library: Hoboken, NJ, USA, 1993. [Google Scholar]
- Li-Chan, E.C.; Hunag, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Peptides derived from Atlantic salmon skin gelatin as dipeptidyl-peptidase IV inhibitors. J. Agric. Food Chem. 2012, 60, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Mullally, M.M.; O’Callaghan, D.M.; FitzGerald, R.J.; Donnelly, W.; Dalton, J.P. Proteolytic and peptidolytic activities in commercial pancreatic protease preparations and their relationship to some whey protein hydrolyzate characteristics. J. Agric. Food Chem. 1994, 42, 2973–2981. [Google Scholar] [CrossRef]
- Cushman, D.; Cheung, H. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Cheung, H.-S.; Wang, F.-L.; Ondetti, M.A.; Sabo, E.F.; Cushman, D.W. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar]
- Vitali, A. Proline-rich peptides: Multifunctional bioactive molecules as new potential therapeutic drugs. Curr. Protein Pept. Sci. 2015, 16, 147–162. [Google Scholar] [CrossRef]
- Morgan, A.A.; Rubenstein, E. Proline: The distribution, frequency, positioning, and common functional roles of proline and polyproline sequences in the human proteome. PLoS ONE 2013, 8, e53785. [Google Scholar] [CrossRef]
- De Luca, L.; Chiminazzo, A.; Sperni, L.; Strukul, G.; Scarso, A. Pyrrolidine-Containing Bisphosphonates as Potential Anti-Resorption Bone Drugs. Chem. A Eur. J. 2017, 23, 3474–3478. [Google Scholar] [CrossRef]
- Trapero, A.; Llebaria, A. A prospect for pyrrolidine iminosugars as antidiabetic α-glucosidase inhibitors. J. Med. Chem. 2012, 55, 10345–10346. [Google Scholar] [CrossRef]
- Reyes, A.A.; Porras, B.H.; Chasalow, F.I.; Klahr, S. L-arginine decreases the infiltration of the kidney by macrophages in obstructive nephropathy and puromycin-induced nephrosis. Kidney Int. 1994, 45, 1346–1354. [Google Scholar] [CrossRef] [PubMed]
- Zoja, C.; Benigni, A.; Camozzi, D.; Corna, D.; Longaretti, L.; Todeschini, M.; Remuzzi, G. Combining lisinopril and l-arginine slows disease progression and reduces endothelin-1 in passive Heymann nephritis. Kidney Int. 2003, 64, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Guang, C.; Phillips, R.D. Plant food-derived angiotensin I converting enzyme inhibitory peptides. J. Agric. Food Chem. 2009, 57, 5113–5120. [Google Scholar] [CrossRef]
- Liu, M.; Du, M.; Zhang, Y.; Xu, W.; Wang, C.; Wang, K.; Zhang, L. Purification and identification of an ACE inhibitory peptide from walnut protein. J. Agric. Food Chem. 2013, 61, 4097–4100. [Google Scholar] [CrossRef] [PubMed]
- Quirós, A.; del Mar Contreras, M.; Ramos, M.; Amigo, L.; Recio, I. Stability to gastrointestinal enzymes and structure–activity relationship of β-casein-peptides with antihypertensive properties. Peptides 2009, 30, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Gevaert, B.; Veryser, L.; Verbeke, F.; Wynendaele, E.; De Spiegeleer, B. Fish hydrolysates: A regulatory perspective of bioactive peptides. Protein Pept. Lett. 2016, 23, 1052–1060. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Tiwari, B. Bioactive carbohydrates and peptides in foods: An overview of sources, downstream processing steps and associated bioactivities. Int. J. Mol. Sci. 2015, 16, 22485–22508. [Google Scholar] [CrossRef]








































© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).