
The Sequestration of Oxy-Polybrominated Diphenyl Ethers in the Nudibranchs Miamira magnifica and Miamira miamirana

 ,
,
Abstract
:1. Introduction
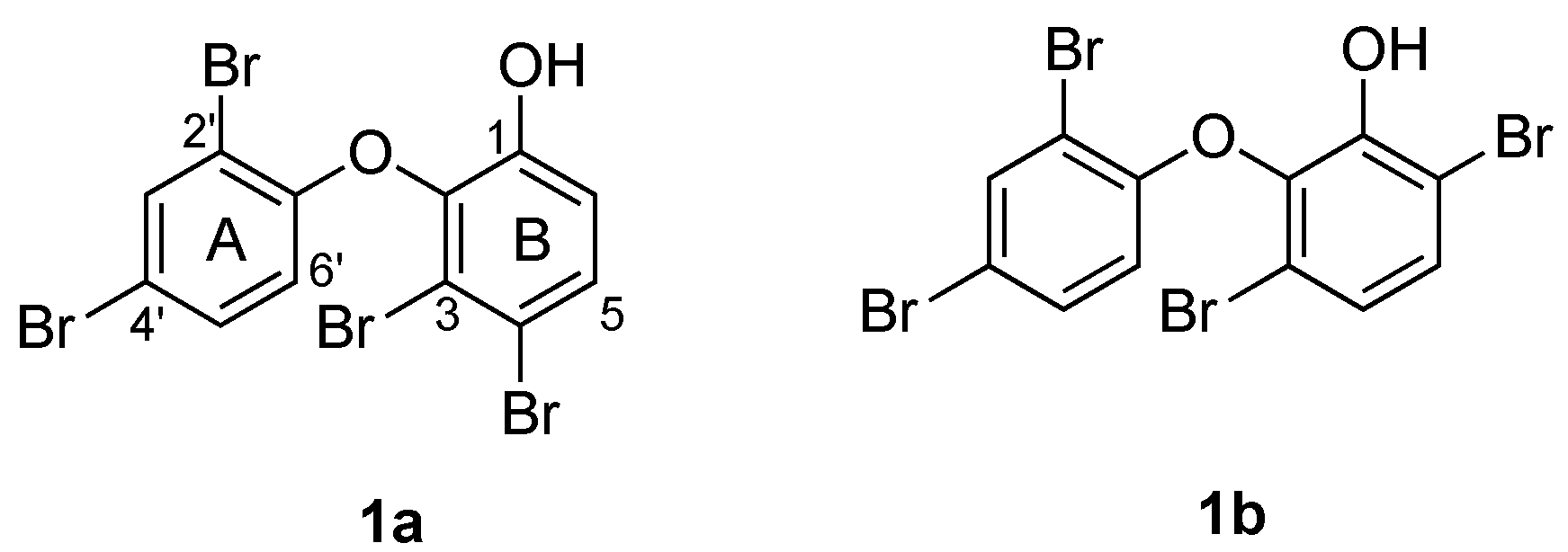
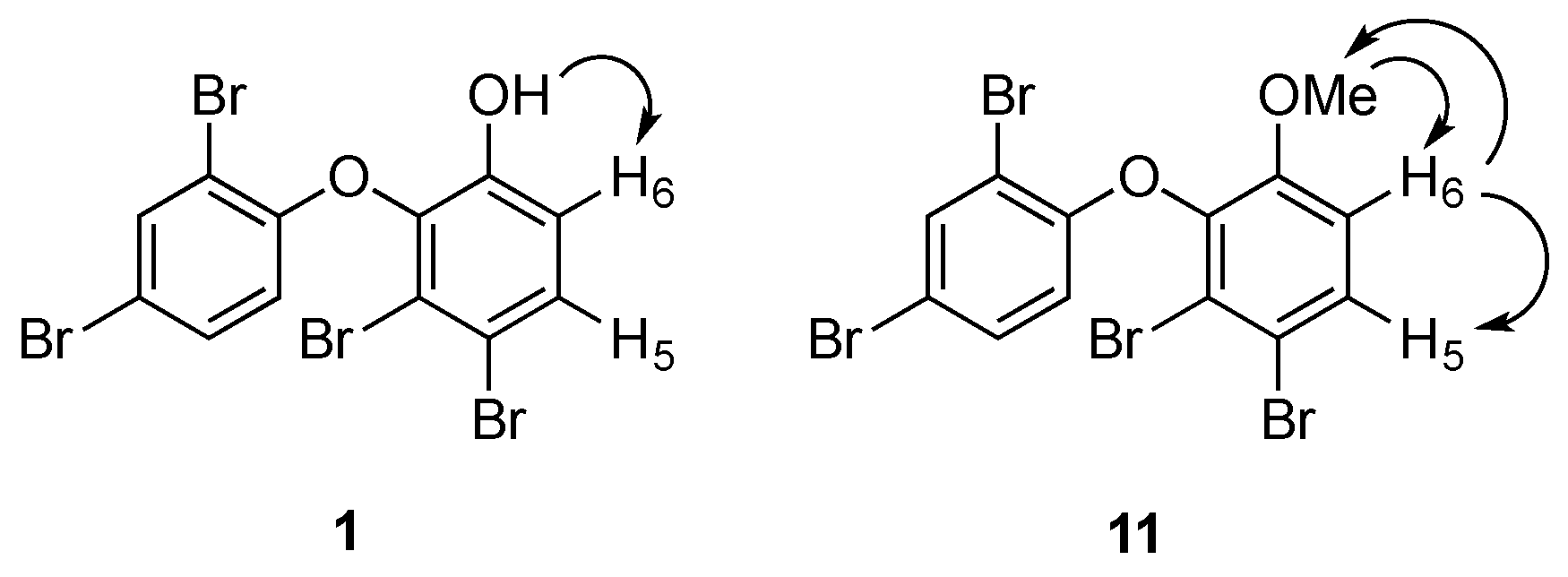
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedure
3.2. Biological Material
3.3. Extraction and Isolation of O-PBDEs
3.4. Methylation of 1
3.5. Brine Shrimp Lethality Assay
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hanif, N.; Tanaka, J.; Setiawan, A.; Trianto, A.; de Voogd, N.J.; Murni, A.; Tanaka, C.; Higa, T. Polybrominated diphenyl ethers from the Indonesian sponge Lamellodysidea herbacea. J. Nat. Prod. 2007, 70, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Utkina, N.K.; Kazantseva, M.V.; Denisenko, V.A. Brominated diphenyl ethers from the marine sponge Dysidea fragilis. Chem. Nat. Compd. 1987, 23, 508–509. [Google Scholar] [CrossRef]
- Il’in, S.G.; Utkina, N.K.; Veselova, M.V.; Struchkov, Y.T. Crystal structure of the complex of brominated diphenyl ethers from the marine sponge Dysidea fragilis. Russ. Chem. Bull. 1996, 756–758. [Google Scholar]
- Carté, B.; Faulkner, D.J. Polybrominated diphenyl ethers from Dysidea herbacea, Dysidea chlorea and Phyllospongia foliascens. Tetrahedron 1981, 37, 2335–2339. [Google Scholar] [CrossRef]
- Hattori, T.; Konno, A.; Adachi, K.; Shizuri, Y. Four new bioactive bromophenols from the Palauan sponge Phyllospongia dendyi. Fish. Sci. 2001, 67, 899–903. [Google Scholar] [CrossRef]
- Liu, H.; Namikoshi, M.; Meguro, S.; Nagai, H.; Kobayashi, H.; Yao, X. Isolation and characterization of polybrominated diphenyl ethers as inhibitors of microtubule assembly from the marine sponge Phyllospongia dendyi collected at Palau. J. Nat. Prod. 2004, 67, 472–474. [Google Scholar] [CrossRef] [PubMed]
- Carte, B.; Kernan, M.R.; Barrabee, E.B.; Faulkner, D.J.; Matsumoto, G.K.; Clardy, J. Metabolites of the nudibranch Chromodoris funerea and the singlet oxygen oxidation products of furodysin and furodysinin. J. Org. Chem. 1986, 51, 3528–3532. [Google Scholar] [CrossRef]
- Fahey, S.J.; Garson, M.J. Geographic variation of natural products of tropical nudibranch Asteronotus cespitosus. J. Chem. Ecol. 2002, 28, 1773–1785. [Google Scholar] [CrossRef] [PubMed]
- Becerro, M.A.; Starmer, J.A.; Paul, V.J. Chemical defenses of cryptic and aposematic gastropterid molluscs feeding on their host sponge Dysidea granulosa. J. Chem. Ecol. 2006, 32, 1491–1500. [Google Scholar] [CrossRef] [PubMed]
- Vetter, W.; Stoll, E.; Garson, M.J.; Fahey, S.J.; Gaus, C.; Müller, J.F. Sponge halogenated natural products found at parts-per-million levels in marine mammals. Environ. Toxicol. Chem. 2002, 21, 2014–2019. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wiseman, S.; Chang, H.; Zhang, X.; Jones, P.D.; Hecker, M.; Kannan, K.; Tanabe, S.; Hu, J.; Lam, M.H.W. Origin of hydroxylated brominated diphenyl ethers: Natural compounds or man-made flame retardants? Environ. Sci. Technol. 2009, 43, 7536–7542. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Xu, L.; Reddy, C.M. Two abundant bioaccumulated halogenated compounds are natural products. Science 2005, 307, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Guitart, C.; Slattery, M.; Ankisetty, S.; Radwan, M.; Ross, S.J.; Letcher, R.J.; Reddy, C.M. Contemporary 14C radiocarbon levels of oxygenated polybrominated diphenyl ethers (O-PBDEs) isolated in sponge-cyanobacteria associations. Mar. Pollut. Bull. 2011, 62, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Unson, M.D.; Holland, N.D.; Faulkner, D.J. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of the crystalline metabolite in the sponge tissue. Mar. Biol. 1994, 119, 1–11. [Google Scholar] [CrossRef]
- Agarwal, V.; El Gamal, A.A.; Yamanaka, K.; Poth, D.; Kersten, R.D.; Schorn, M.; Allen, E.E.; Moore, B.S. Biosynthesis of polybrominated aromatic organic compounds by marine bacteria. Nat. Chem. Biol. 2014, 10, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Yong, K.W.L.; Mudianta, I.W.; Cheney, K.L.; Mollo, E.; Blanchfield, J.T.; Garson, M.J. Isolation of norsesterterpenes and spongian diterpenes from Dorisprismatica (=Glossodoris) atromarginata. J. Nat. Prod. 2015, 78, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Katavic, P.L.; Jumaryatno, P.; Hooper, J.N.A.; Blanchfield, J.T.; Garson, M.J. Oxygenated terpenoids from the Australian sponges Coscinoderma matthewsi and Dysidea sp., and the nudibranch Chromodoris albopunctata. Aust. J. Chem. 2012, 65, 531–538. [Google Scholar] [CrossRef]
- White, A.M.; Pierens, G.K.; Forster, L.C.; Winters, A.E.; Cheney, K.L.; Garson, M.J. Rearranged diterpenes and norditerpenes from three Australian Goniobranchus mollusks. J. Nat. Prod. 2016, 79, 477–483. [Google Scholar] [CrossRef] [PubMed]
- White, A.M.; Dewi, A.S.; Cheney, K.L.; Winters, A.E.; Blanchfield, J.T.; Garson, M.J. Oxygenated diterpenes from the Indo-Pacific nudibranchs Goniobranchus splendidus and Ardeadoris egretta. Nat. Prod. Commun. 2016, 11, 921–924. [Google Scholar]
- Capon, R.; Ghisalberti, E.L.; Jefferies, P.R.; Skelton, B.W.; White, A.H. Structural studies of halogenated diphenyl ethers from a marine sponge. J. Chem. Soc. Perk. Tran. 1981, 39, 2464–2467. [Google Scholar] [CrossRef]
- Salvá, J.; Faulkner, D.J. A new brominated diphenyl ether from a Philippine Dysidea species. J. Nat. Prod. 1990, 53, 757–760. [Google Scholar] [CrossRef]
- Bowden, B.F.; Towerzey, L.; Junk, P.C. A new brominated diphenyl ether from the marine sponge Dysidea herbacea. Aust. J. Chem. 2000, 53, 299–301. [Google Scholar] [CrossRef]
- Agrawal, M.S.; Bowden, B.F. Marine sponge Dysidea herbacea revisited: Another brominated diphenyl ether. Mar. Drugs 2005, 3, 9–14. [Google Scholar] [CrossRef]
- Calcul, L.; Chow, R.; Oliver, A.G.; Tenney, K.; White, K.N.; Wood, A.W.; Fiorilla, C.; Crews, P. NMR Strategy for unraveling structures of bioactive sponge-derived oxy-polyhalogenated diphenyl ethers. J. Nat. Prod. 2009, 72, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Pretsch, E.; Bühlmann, P.; Affolter, C.; Pretsch, E.; Bhuhlmann, P.; Affolter, C. Structure Determination of Organic Compounds; Springer: Berlin, Germany, 2009; Volume 13. [Google Scholar]
- Duffy, J.E.; Paul, V.J. Prey nutritional quality and the effectiveness of chemical defenses against tropical reef fishes. Oecologia 1992, 90, 333–339. [Google Scholar] [CrossRef]
- Pennings, S.C.; Paul, V.J. Secondary chemistry does not limit dietary range of the specialist sea hare Stylocheilus longicauda (Quoy et Gaimard 1824). J. Exp. Mar. Biol. Ecol. 1993, 174, 97–113. [Google Scholar] [CrossRef]
- Pennings, S.C.; Pablo, S.R.; Paul, V.J.; Duffy, J.E. Effects of sponge secondary metabolites in different diets on feeding by three groups of consumers. J. Exp. Mar. Biol. Ecol. 1994, 180, 137–149. [Google Scholar] [CrossRef]
- Handayani, D.; Edrada, R.A.; Proksch, P.; Wray, V.; Witte, L.; Soest, R.W.M.V.; Kunzmann, A.; Soedarsono. Four new bioactive polybrominated diphenyl ethers of the sponge Dysidea herbacea from West Sumatra, Indonesia. J. Nat. Prod. 1997, 60, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Cheney, K.L.; White, A.; Mudianta, I.W.; Winters, A.E.; Quezada, M.; Capon, R.J.; Mollo, E.; Garson, M.J. Choose your weaponry: Selective storage of a single toxic compound, Latrunculin A, by closely related nudibranch molluscs. PLoS ONE 2016, 11, e0145134. [Google Scholar] [CrossRef] [PubMed]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Guo, Y.W.; Cimino, G. Chemical studies on Indopacific Ceratosoma nudibranchs illuminate the protective role of their dorsal horn. Chemoecology 2005, 15, 31–36. [Google Scholar] [CrossRef]
- Dexter, A.F.; Garson, M.J.; Hemling, M.E. Isolation of a novel bastadin from the temperate marine sponge Ianthella sp. J. Nat. Prod. 1993, 56, 782–786. [Google Scholar] [CrossRef]
- Vetter, W.; Kirres, J.; Bendig, P. Bromination of 2-methoxydiphenyl ether to an average of tetrabrominated 2-methoxydiphenyl ethers. Chemosphere 2011, 84, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
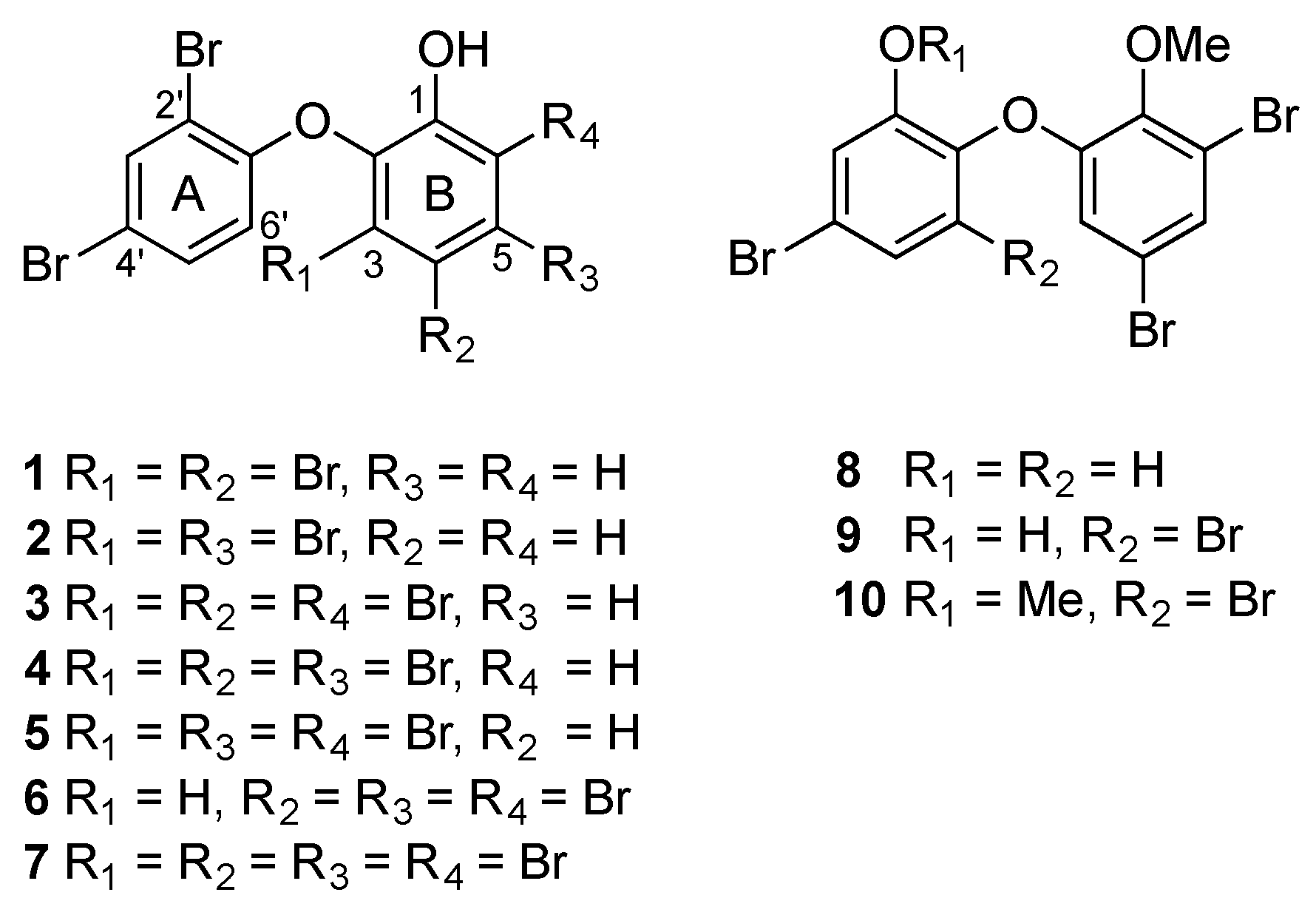
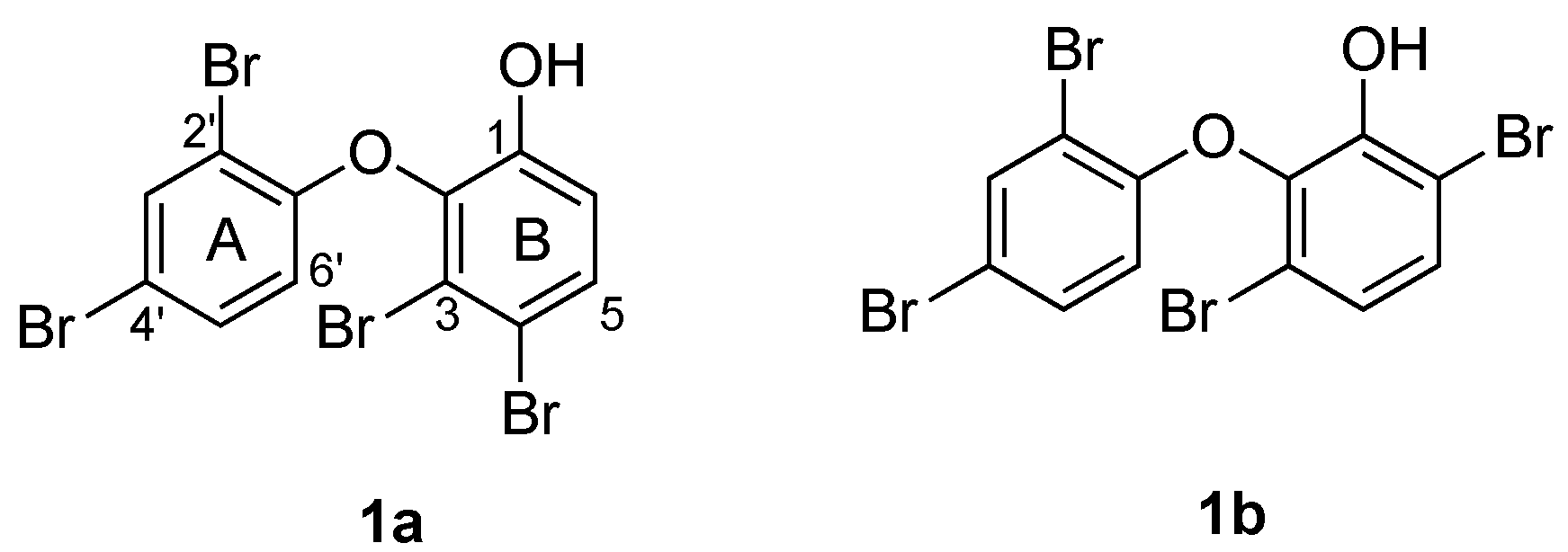
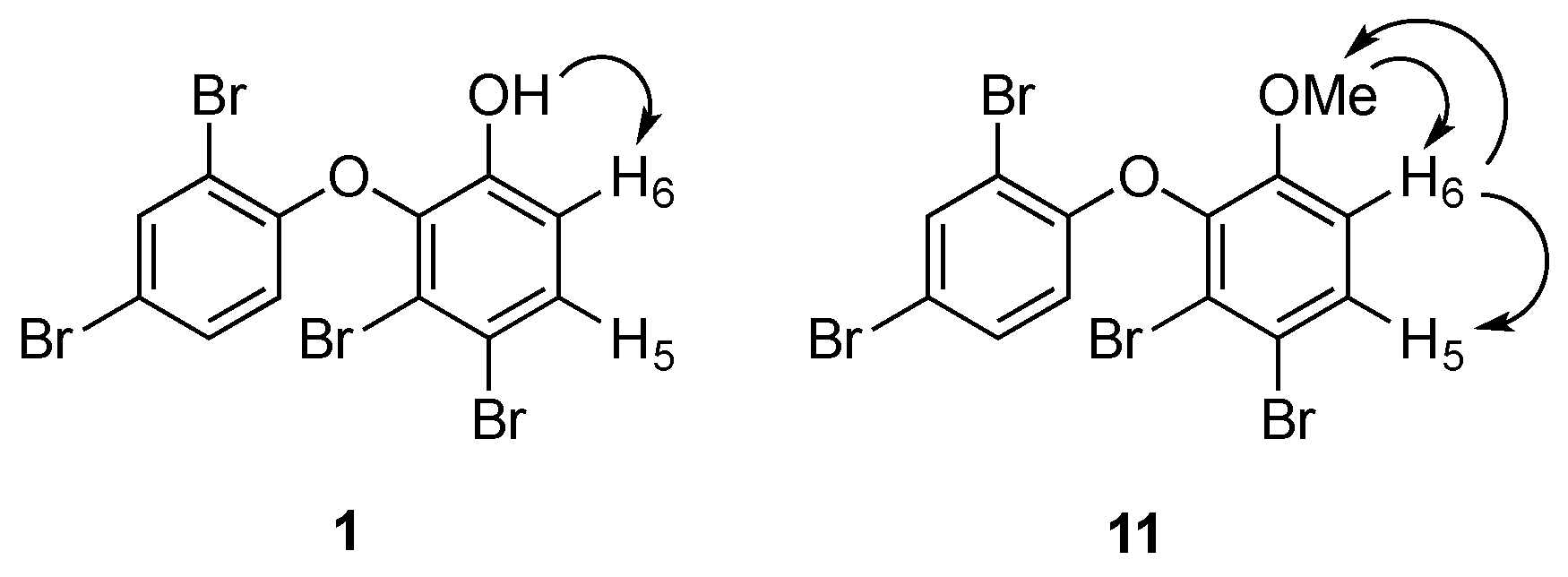

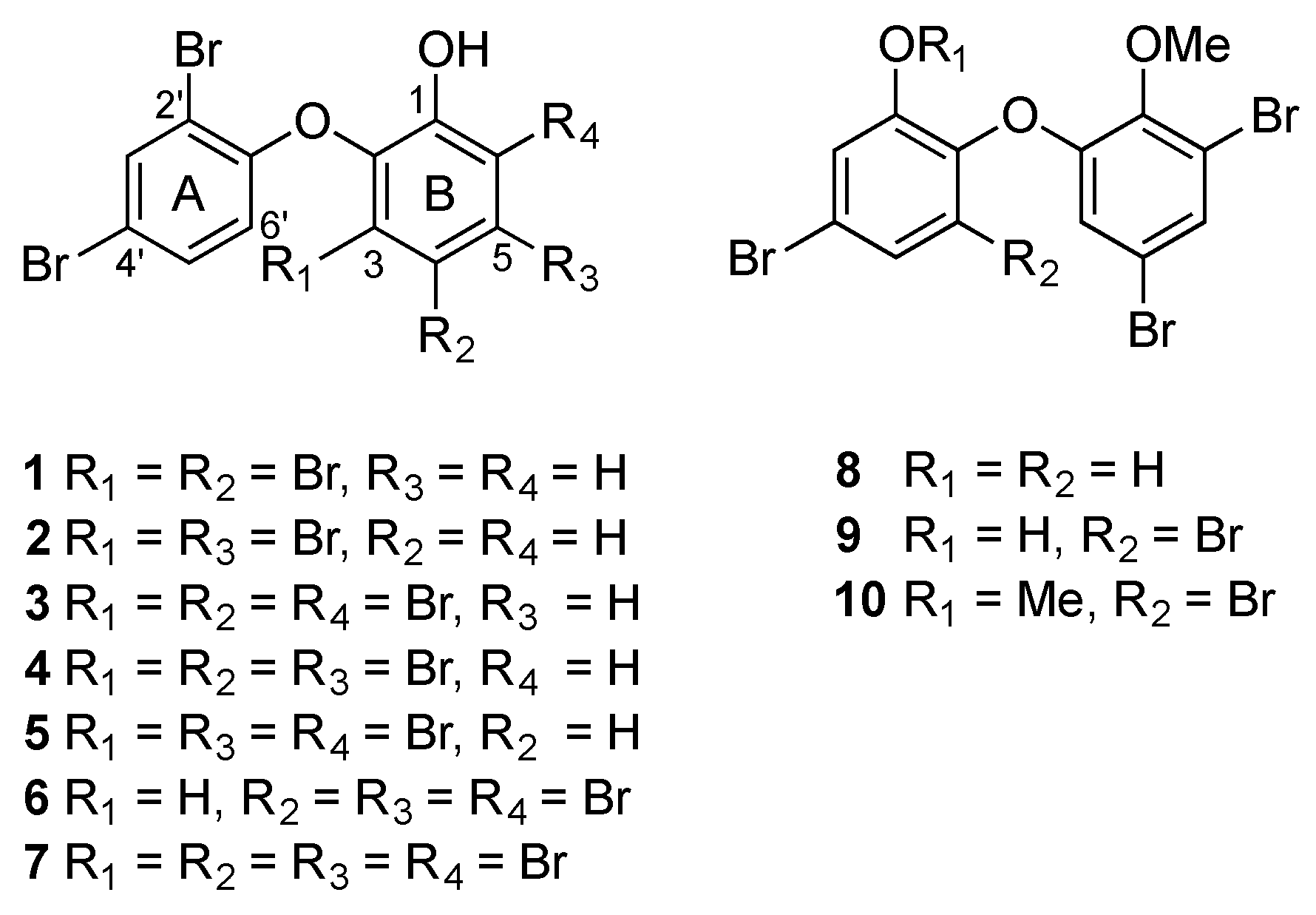{kind=link}
{kind=link}
{kind=link}
| C | 1 a (exp) | 1a b | 1b b |
|---|---|---|---|
| 1 | 149.1 | 147.3 | 151.6 |
| 2 | 140.3 | 148.8 c | 148.8 c |
| 3 | 120.0 | 116.6 | 112.3 c |
| 4 | 116.0 | 118.7 | 126.3 c |
| 5 | 131.2 | 128.5 | 128.5 |
| 6 | 117.1 | 116.6 | 109.0 c |
| O-PBDEs | M. magnifica | M. miamirana | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mantle | % w/w a | Gut | % w/w a | Horn | % w/w a,b | Mantle | % w/w a,b | Gut | % w/w a,b | ||
| O-triBDE | 8 | - | - | - | - | - | - | X | 70% | X | 16% |
| O-tetraBDE | 1 | X | 31% | X | 6% | X | 18% | - | - | - | - |
| 2 | X | 45% | X | 60% | X | 64% | - | - | - | - | |
| 9 | - | - | - | - | - | - | X | 25% | X | 42% | |
| 10 | - | - | - | - | - | - | X | 5% | X | 42% | |
| O-pentaBDE | 3 | X | 22% | X | 18% | X | 18% | - | - | - | - |
| 4 | - | - | X | 5% | - | - | - | - | - | - | |
| 5 | trace | 2% | X | 7% | - | - | - | - | - | - | |
| 6 | - | - | trace | 1% | - | - | - | - | - | - | |
| O-hexaBDE | 7 | - | - | trace | 3% | - | - | - | - | - | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dewi, A.S.; Cheney, K.L.; Urquhart, H.H.; Blanchfield, J.T.; Garson, M.J. The Sequestration of Oxy-Polybrominated Diphenyl Ethers in the Nudibranchs Miamira magnifica and Miamira miamirana. Mar. Drugs 2016, 14, 198. https://doi.org/10.3390/md14110198
Dewi AS, Cheney KL, Urquhart HH, Blanchfield JT, Garson MJ. The Sequestration of Oxy-Polybrominated Diphenyl Ethers in the Nudibranchs Miamira magnifica and Miamira miamirana. Marine Drugs. 2016; 14(11):198. https://doi.org/10.3390/md14110198
Chicago/Turabian StyleDewi, Ariyanti S., Karen L. Cheney, Holly H. Urquhart, Joanne T. Blanchfield, and Mary J. Garson. 2016. "The Sequestration of Oxy-Polybrominated Diphenyl Ethers in the Nudibranchs Miamira magnifica and Miamira miamirana" Marine Drugs 14, no. 11: 198. https://doi.org/10.3390/md14110198
APA StyleDewi, A. S., Cheney, K. L., Urquhart, H. H., Blanchfield, J. T., & Garson, M. J. (2016). The Sequestration of Oxy-Polybrominated Diphenyl Ethers in the Nudibranchs Miamira magnifica and Miamira miamirana. Marine Drugs, 14(11), 198. https://doi.org/10.3390/md14110198