Construction of Monophosphoryl Lipid A Producing Escherichia coli Mutants and Comparison of Immuno-Stimulatory Activities of Their Lipopolysaccharides

Abstract
:1. Introduction
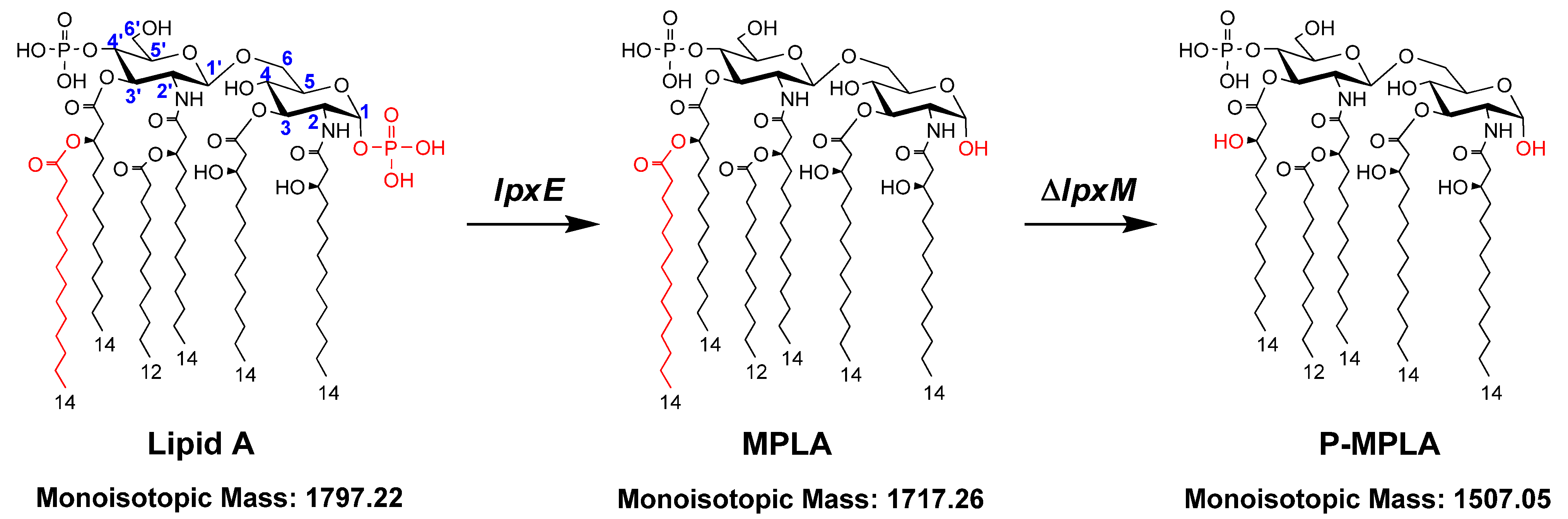
2. Results and Discussion
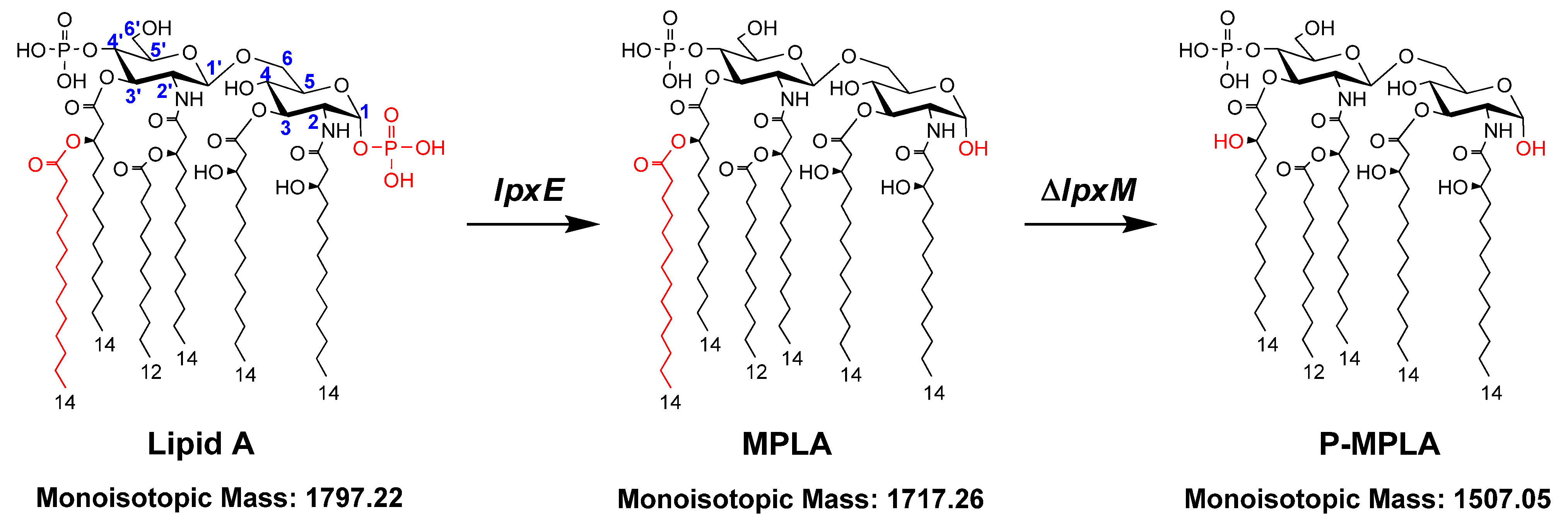
2.1. Construction of E. coli Strains that Can Synthesize MPLA or P-MPLA by Chromosomal Gene Integration and/or Gene Deletion
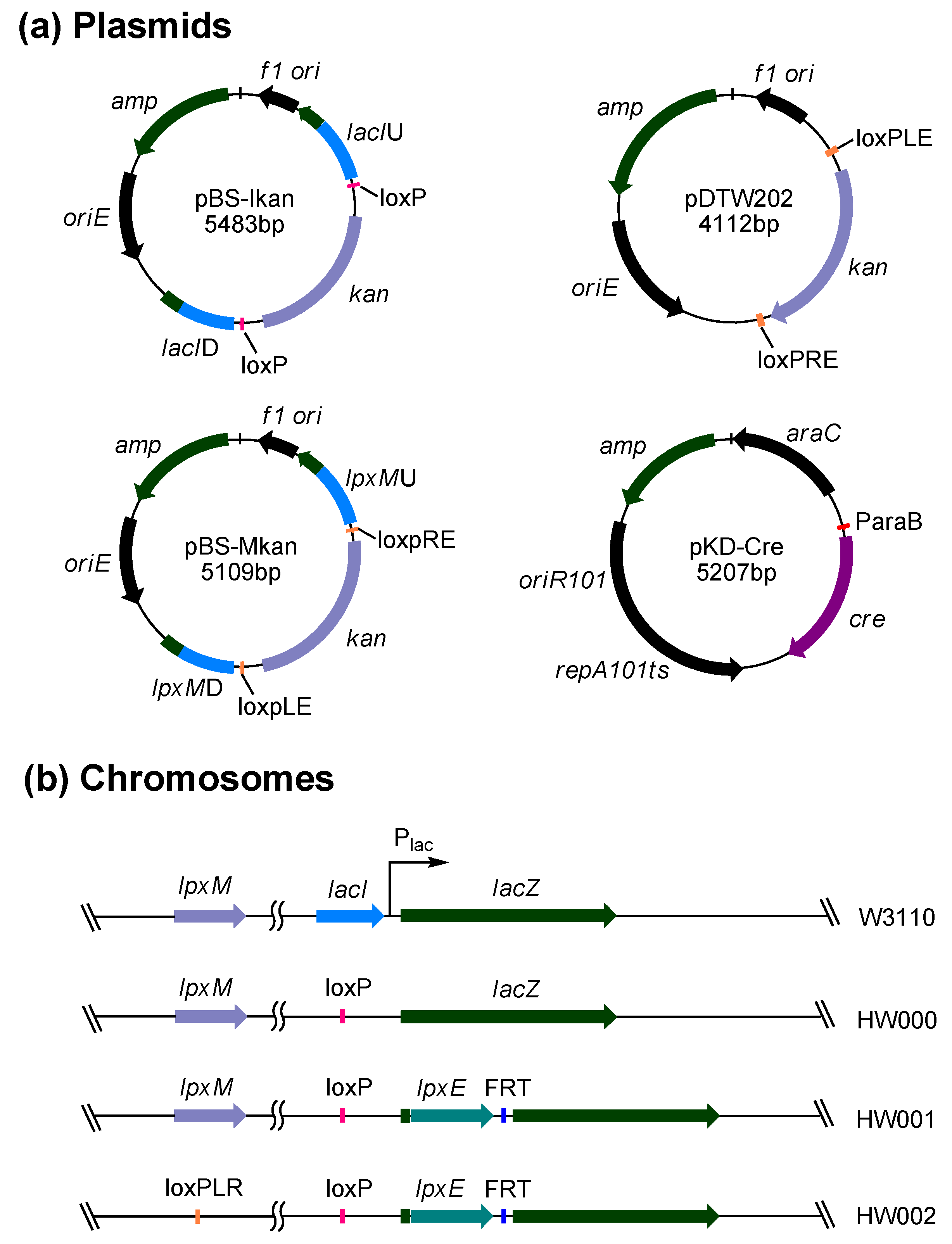
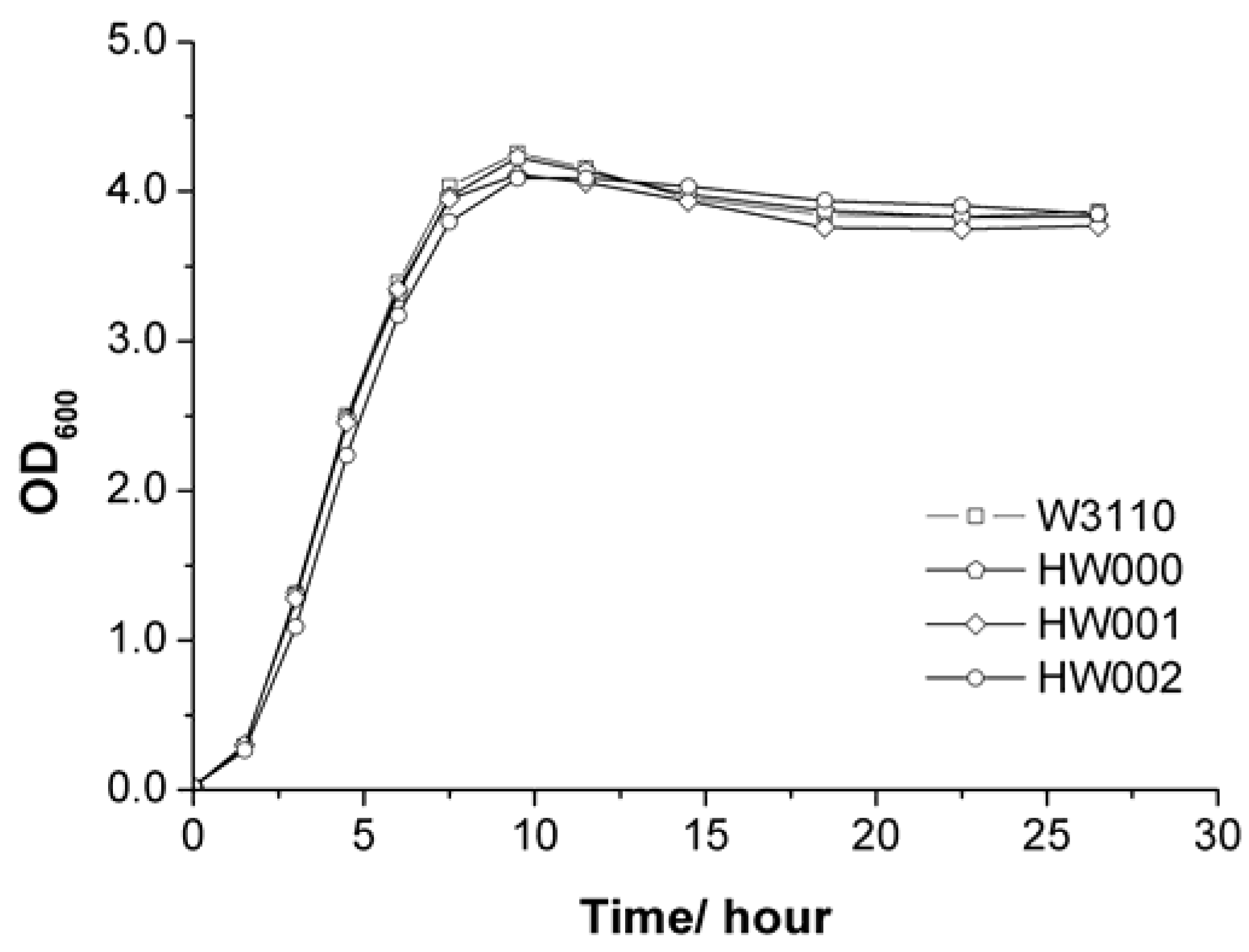
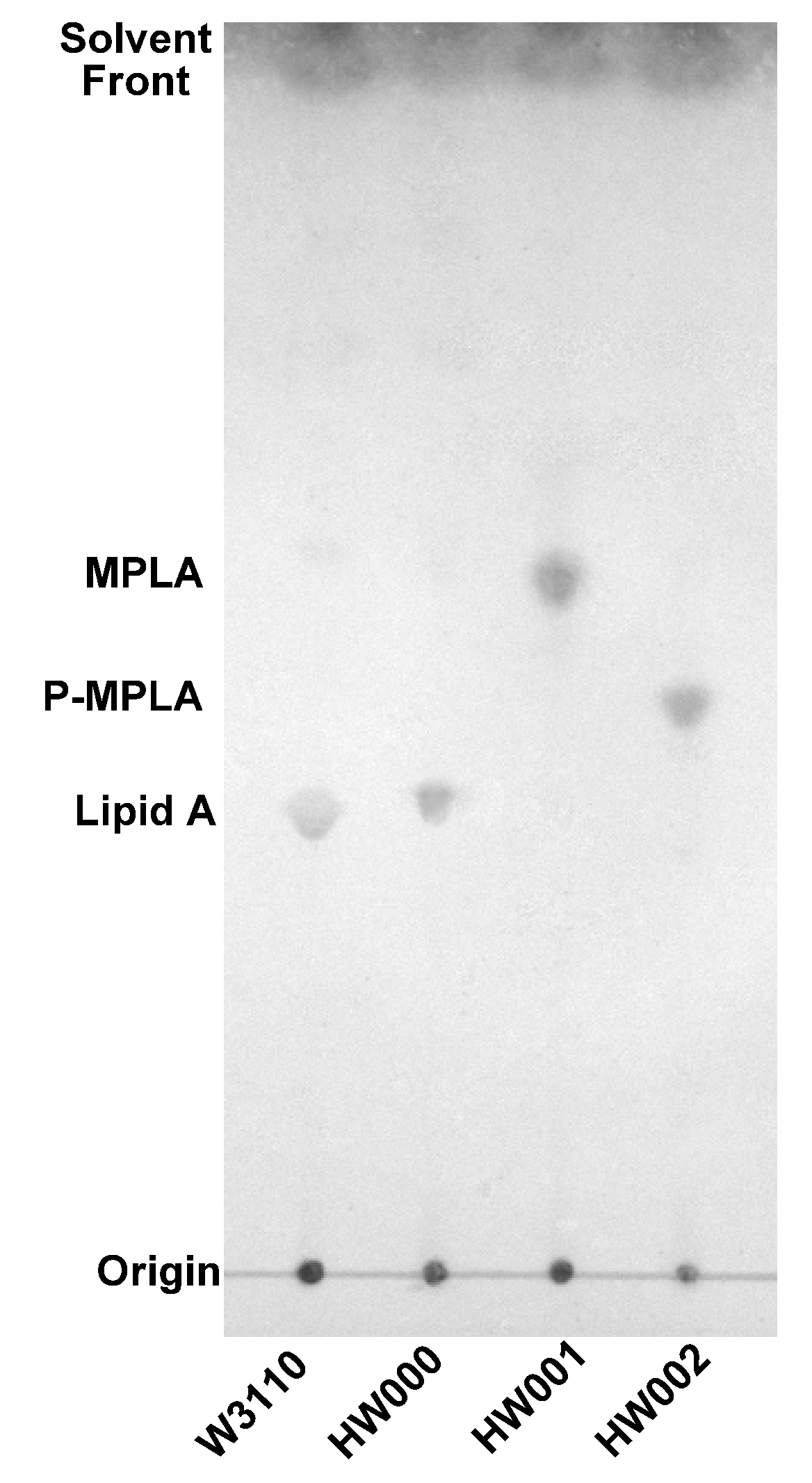
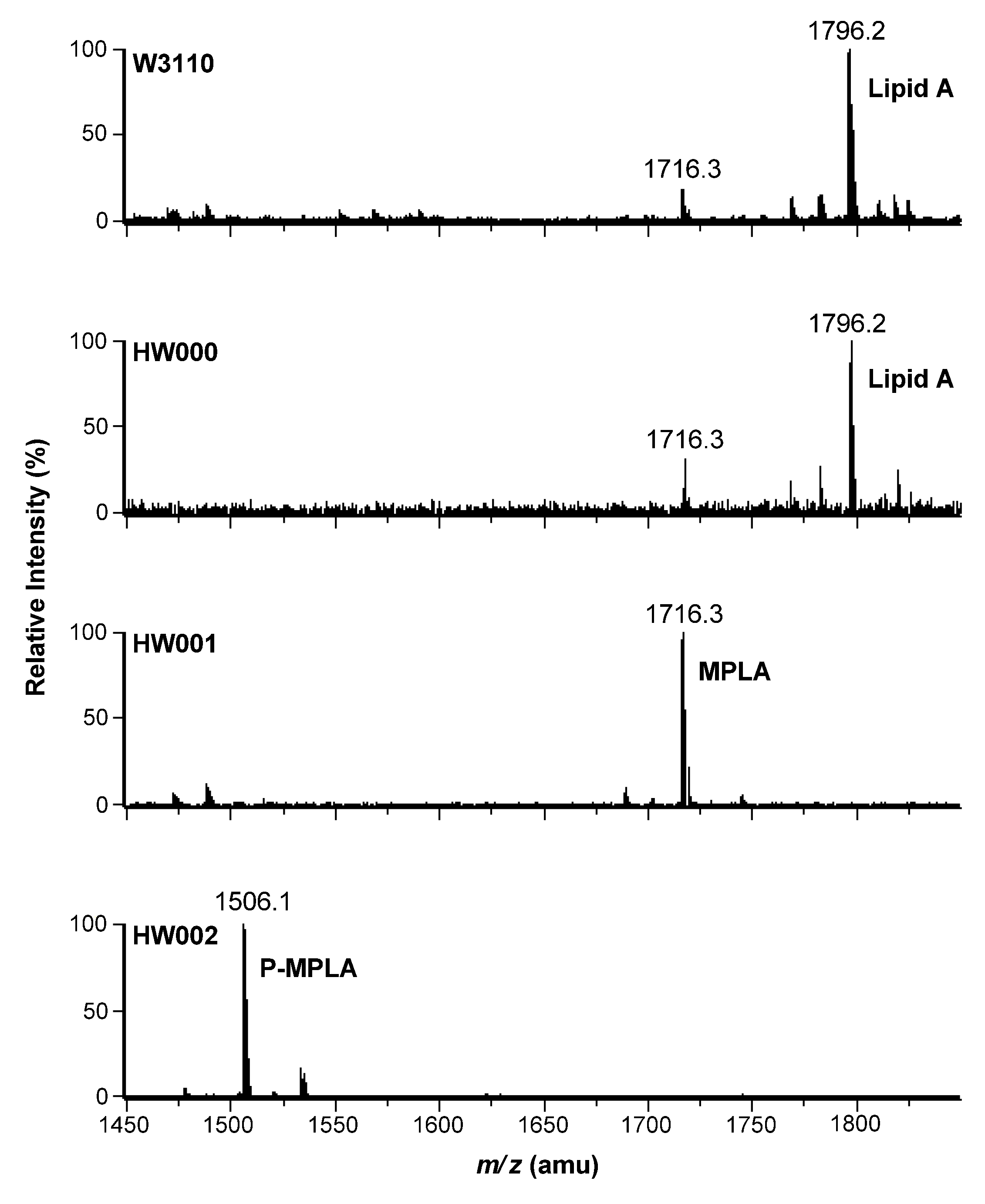
2.2. Structure Analysis of Lipid A Extracted from Three E. coli Mutants

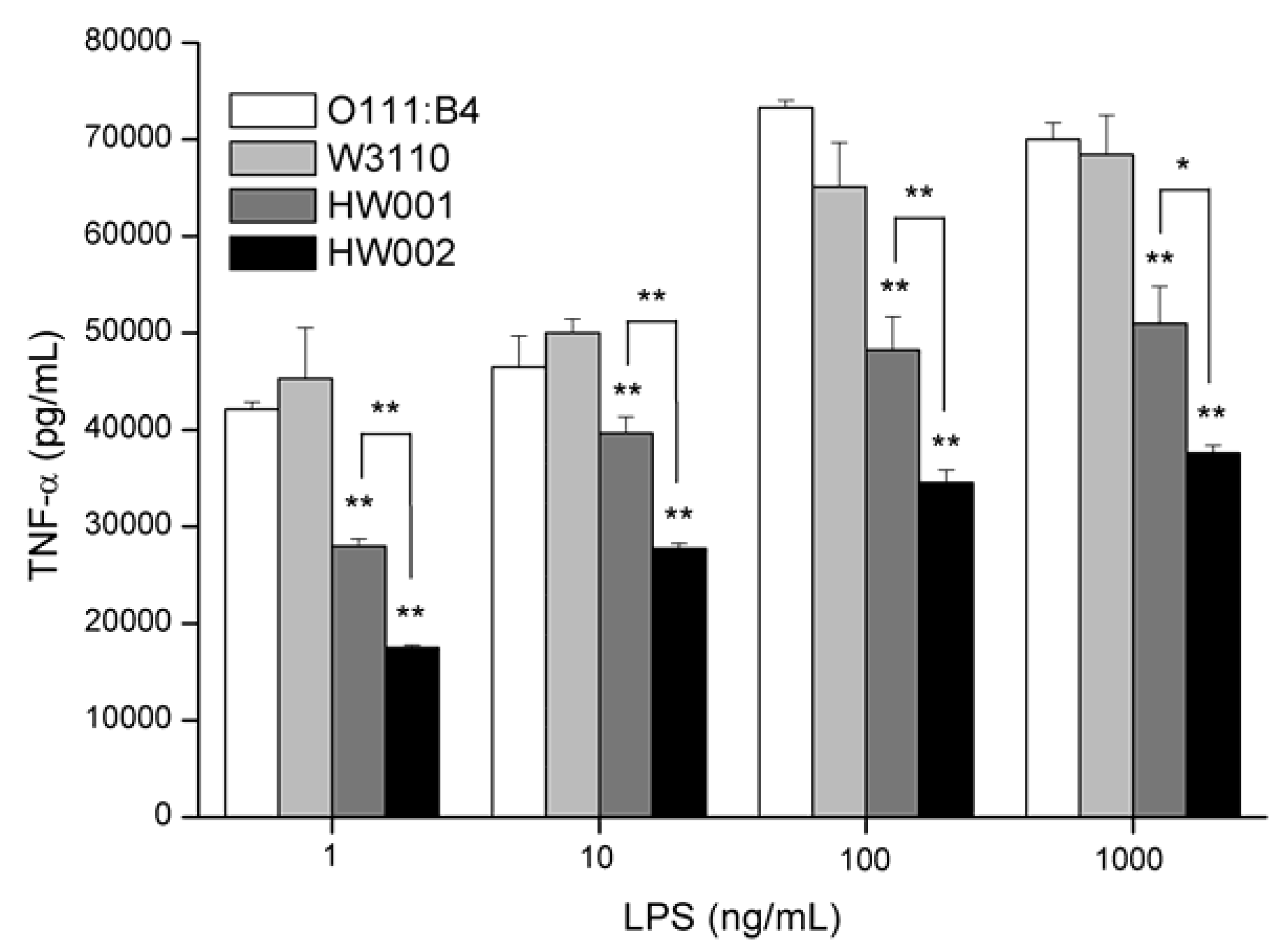
2.3. Comparison of the Stimulation Activities of LPS Isolated from Different E. coli Mutants
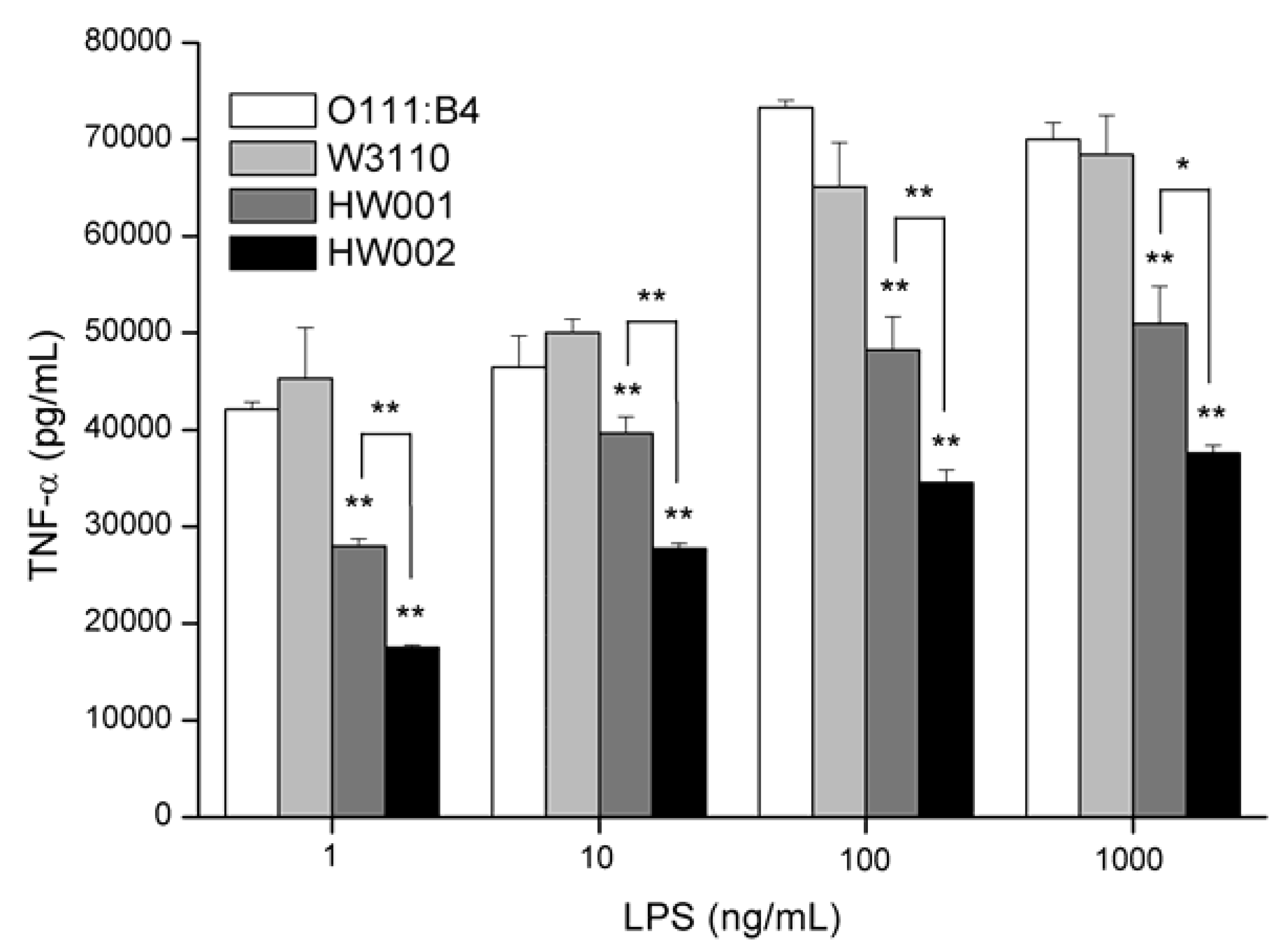
3. Experimental Section
3.1. DNA Preparation and PCR Techniques

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′→3′) | Restriction site |
|---|---|---|
| lacIU-F | CCGCTCGAGACAGTATCGGCCTCAGGAAGAT | XhoI |
| lacIU-R | CGGAATTCTTAATGCAGCTGGCACGACAGG | EcoRI |
| lacID-F | GAAGATCTGCGACATCGTATAACGTTACTG | BglII |
| lacID-R | AACTGCAGCAAGACCGCAAGGAATTAATCG | PstI |
| lpxMU-F | CCCAAGCTTGCGTTACGGCGATCATCACCAT | HindIII |
| lpxMU-R | CCCGGCTAGCAAAGATCTTTATCCCATCAAAT | NheI |
| lpxMD-F | GTGGATCCATCAAACTCAGGAATGTATTCGC | BamHI |
| lpxMD-R | ATGAGCTCATGGTCGCAGCTACACCACGCG | SacI |
| kan-loxP-F | ATGAATTCGCGGCCGCATAACTTCGTATAA | EcoRI |
| kan-loxP-R | AGAGATCTGCGGCCGCATAACTTCGTATAA | BglII |
| loxpLE-F | TTACCGTTCGTATAATGTATGCTATACGAAGTTATA | / |
| loxpLE-R | TCGATATAACTTCGTATAGCATACATTATACGAACGGTAAGGCC | / |
| loxpRE-F | CTAGTATAACTTCGTATAATGTATGCTATACGAACGGTATAGC | / |
| loxpRE-R | TATACCGTTCGTATAGCATACATTATACGAAGTTATA | / |
| kan-F | TAACTCGAGATTCACGCTGCCGCAAGCACTCAGG | XhoI |
| kan-R | AAGTCTAGACGAACCCCAGAGTCCCGCTCAGA | XbaI |
| kan-loxpLE-F | ATGGATCCAATACGACTCACTATAGGGCG | BamHI |
| kan-loxpRE-R | ACCTCTAGAGCGCAATTAACCCTCACTAAAG | XbaI |
| cre-F | CGAGCTCAGGAGGTTATAAAGGATGTCCAATTTACTG | SacI |
| ACCGTACACCA | ||
| cre-R | TACCATGGTCTAATCGCCATCTTCCAGCAG | NcoI |
| lacZ-F | CCATTCAGGCTGCGCAACTGTT | / |
| lacZ-R | TTAATGCAGCTGGCACGACAGG | / |
3.2. Construction of Plasmids and Bacterial Strains
| Strains or Plasmids | Description | Source |
|---|---|---|
| Strains | ||
| W3110 | Wild-type E. coli, F−, λ− | Laboratory strain |
| W3110/pKD46 | W3110 transformed by pKD46 | This work |
| HW000 | W3110 ΔlacI | This work |
| HW001 | W3110 ΔlacIlacZ::lpxE | This work |
| HW002 | W3110 ΔlacIΔlpxMlacZ::lpxE | This work |
| Plasmids | ||
| pKD46 | ParaBγβ exo, Repts,AmpR | [23] |
| pMG2-kanr | loxP-kan-loxP, AmpR, KanR | [20] |
| pK18mobsacB | Mob+, sacB, KanR | [22] |
| pDTW202 | loxPLE-kan-loxPRE, AmpR, KanR | This work |
| pCP20 | FLP+, λ cI857+, λpRRepts, CamR, AmpR | [23] |
| pSH47 | GAL1p-Cre-CYC1T, AmpR | [24] |
| pKD-Cre | ParaB cre, Repts, AmpR | This work |
| pBlueScript II SK+ | Cloning vector, ColE1, lacZ, AmpR | Stratagene |
| pBS-Ikan | Plasmid for deleting lacI in E. coli | This work |
| pBS-Mkan | Plasmid for deleting lpxM in E. coli | This work |
| pWSK29-FnlpxE-Fkan | Plasmid for containing lpxE | [19] |
3.3. Isolation and Analysis of Lipid A from E. coli Strains
3.4. Extraction and Purification of LPS
3.5. LPS Stimulation of the Murine Macrophage Cell Line RAW264.7
4. Conclusions
Acknowledgments
References
- Wang, X.; Quinn, P.J. Lipopolysaccharide: Biosynthetic pathway and structure modification. Prog. Lipid Res. 2010, 49, 97–107. [Google Scholar]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar]
- Wang, X.; Quinn, P.J. Endotoxins: Lipopolysaccharides of Gram-Negative Bacteria. In Endotoxins: Structure, Function and Recognition; Wang, X., Quinn, P.J., Eds.; Springer: London, UK, 2010; Volume 53, pp. 3–26. [Google Scholar]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar]
- Silipo, A.; Molinaro, A. The Diversity of the Core Oligosaccharide in Lipopolysaccharides. In Endotoxins: Structure, Function and Recognition; Wang, X., Quinn, P.J., Eds.; Springer: London, UK, 2010; Volume 53, pp. 69–100. [Google Scholar]
- Baker, P.J.; Hraba, T.; Taylor, C.E.; Myers, K.R.; Takayama, K.; Qureshi, N.; Stuetz, P.; Kusumoto, S.; Hasegawa, A. Structural features that influence the ability of lipid A and its analogs to abolish expression of suppressor T cell activity. Infect. Immun. 1992, 60, 2694–2701. [Google Scholar]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar]
- Moingeon, P.; Haensler, J.; Lindberg, A. Towards the rational design of Th1 adjuvants. Vaccine 2001, 19, 4363–4372. [Google Scholar]
- Kanegasaki, S.; Kojima, Y.; Matsuura, M.; Homma, J.Y.; Yamamoto, A.; Kumazawa, Y.; Tanamoto, K.; Yasuda, T.; Tsumita, T.; Imoto, M.; et al. Biological activities of analogues of lipid A based chemically on the revised structural model. Eur. J. Biochem. 1984, 143, 237–242. [Google Scholar] [CrossRef]
- Opal, S.M. Endotoxins and Other Sepsis Triggers. In Endotoxemia and Endotoxin Shock: Disease, Diagnosis and Therapy; Ronco, C., Piccinni, P., Rosner, M., Eds.; Karger: Basel, Switzerland, 2010; Volume 167, pp. 14–24. [Google Scholar]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar]
- Gaekwad, J.; Zhang, Y.; Zhang, W.; Reeves, J.; Wolfert, M.A.; Boons, G.J. Differential induction of innate immune responses by synthetic lipid A derivatives. J. Biol. Chem. 2010, 285, 29375–29386. [Google Scholar]
- Mata-Haro, V.; Cekic, C.; Martin, M.; Chilton, P.M.; Casella, C.R.; Mitchell, T.C. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science 2007, 316, 1628–1632. [Google Scholar]
- Khan, S.A.; Everest, P.; Servos, S.; Foxwell, N.; Zähringer, U.; Brade, H.; Rietschel, E.T.; Dougan, G.; Charles, I.G.; Maskell, D.J. A lethal role for lipid A in Salmonella infections. Mol. Microbiol. 2002, 29, 571–579. [Google Scholar]
- Persing, D.H.; Coler, R.N.; Lacy, M.J.; Johnson, D.A.; Baldridge, J.R.; Hershberg, R.M.; Reed, S.G. Taking toll: Lipid A mimetics as adjuvants and immunomodulators. Trends Microbiol. 2002, 10, S32–S37. [Google Scholar]
- Evans, J.T.; Cluff, C.W.; Johnson, D.A.; Lacy, M.J.; Persing, D.H.; Baldridge, J.R. Enhancement of antigen-specific immunity via the TLR4 ligands MPL adjuvant and Ribi. 529. Expert Rev. Vaccines 2003, 2, 219–229. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Guan, Z.; Ingram, B.O.; Six, D.A.; Song, F.; Wang, X.; Zhao, J. Discovery of new biosynthetic pathways: The lipid A story. J. Lipid Res. 2009, 50, S103–S108. [Google Scholar]
- Wang, X.; Karbarz, M.J.; McGrath, S.C.; Cotter, R.J.; Raetz, C.R.H. MsbA transporter-dependent lipid A 1-dephosphorylation on the periplasmic surface of the inner membrane. J. Biol. Chem. 2004, 279, 49470–49478. [Google Scholar]
- Chen, J.; Tao, G.; Wang, X. Construction of an Escherichia coli mutant producing monophosphoryl lipid A. Biotechnol. Lett. 2011, 33, 1013–1019. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, L.; Ding, Z.; Wang, Z.X.; Shi, G.Y. Improving ethanol productivity by modification of glycolytic redox factor generation in glycerol-3-phosphate dehydrogenase mutants of an industrial ethanol yeast. J. Ind. Microbiol. Biotechnol. 2011, 38, 935–943. [Google Scholar]
- Arakawa, H.; Lodygin, D.; Buerstedde, J.M. Mutant loxP vectors for selectable marker recycle and conditional knock-outs. BMC Biotechnol. 2001, 1, 1–8. [Google Scholar]
- Schaefer, A.; Tauch, A.; Jaeger, W.; Kalinowski, J.; Thierbach, G.; Puehler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK 18 and pK 19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Gueldener, U.; Heinisch, J.; Koehler, G.; Voss, D.; Hegemann, J. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucl. Acids Res. 2002, 30, e23–e23. [Google Scholar]
- Wang, X.; Zhang, C.; Shi, F.; Hu, X. Purification and Characterization of Lipopolysaccharides. In Endotoxins: Structure, Function and Recognition; Wang, X., Quinn, P.J., Eds.; Springer: London, UK, 2010; Volume 53, pp. 27–52. [Google Scholar]
- MassLynx Software, version 4.1, Waters Corporation: Milford, MA, USA, 2005.
- Westphal, O.; Jann, K. Bacterial Lipopolysaccharide-Extraction with Phenol Water and Further Application of Procedure. In Methods in Carbohydrate Chemistry; Whistler, R.L., Wolfrom, M.L., Eds.; Academic Press: New York, NY, USA, 1965; Volume 9, pp. 83–91. [Google Scholar]
- Tirsoaga, A.; Novikov, A.; Adib-Conquy, M.; Werts, C.; Fitting, C.; Cavaillon, J.M.; Caroff, M. Simple method for repurification of endotoxins for biological use. Appl. Environ. Microbiol. 2007, 73, 1803–1808. [Google Scholar]
- Carlson, R.W.; Sanders, R.E.; Napoli, C.; Albersheim, P. Host-symbiont interactions III. Purification and partial characterization of Rhizobium lipopolysaccharides. Plant Physiol. 1978, 62, 912–917. [Google Scholar]
- Fox, C.B.; Friede, M.; Reed, S.G.; Ireton, G.C. Synthetic and Natural TLR4 Agonists as Safe and Effective Vaccine Adjuvants. In Endotoxins: Structure, Function and Recognition; Wang, X., Quinn, P.J., Eds.; Springer: London, UK, 2010; Volume 53, pp. 303–321. [Google Scholar]
- Reed, S.G.; Bertholet, S.; Coler, R.N.; Friede, M. New horizons in adjuvants for vaccine development. Trends Immunol. 2009, 30, 23–32. [Google Scholar]
- Casella, C.R.; Mitchell, T.C. Putting endotoxin to work for us: Monophosphoryl lipid A as a safe and effective vaccine adjuvant. Cell. Mol. Life Sci. 2008, 65, 3231–3240. [Google Scholar]
- Guo, L.; Lim, K.B.; Gunn, J.S.; Bainbridge, B.; Darveau, R.P.; Hackett, M.; Miller, S.I. Regulation of lipid A modifications by Salmonella typhimurium virulence genes phoP-phoQ. Science 1997, 276, 250–253. [Google Scholar]
- Kong, Q.; Six, D.A.; Roland, K.L.; Liu, Q.; Gu, L.; Reynolds, C.M.; Wang, X.; Raetz, C.R.H.; Curtiss, R. Salmonella synthesizing 1-monophosphorylated lipopolysaccharide exhibits low endotoxic activity while retaining its immunogenicity. J. Immunol. 2011, 187, 412–423. [Google Scholar]
- Somerville, J.E.; Cassiano, L.; Darveau, R.P. Escherichia coli msbB gene as a virulence factor and a therapeutic target. Infect. Immun. 1999, 67, 6583–6590. [Google Scholar]
- Lee, S.R.; Kim, S.H.; Jeong, K.J.; Kim, K.S.; Kim, Y.H.; Kim, S.J.; Kim, E.; Kim, J.W.; Chang, K.T. Multi-immunogenic outer membrane vesicles derived from an MsbB-deficient Salmonella enterica serovar typhimurium mutant. J. Microbiol. Biotechnol. 2009, 19, 1271–1279. [Google Scholar]
- Kawasaki, K.; Ernst, R.K.; Miller, S.I. Deacylation and palmitoylation of lipid A by Salmonella outer membrane enzymes modulate host signaling through Toll-like receptor 4. J. Endotoxin Res. 2004, 10, 439–444. [Google Scholar]
- Samples Availability: Available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, Y.; Li, Y.; Chen, J.; Tan, Y.; Guan, F.; Wang, X. Construction of Monophosphoryl Lipid A Producing Escherichia coli Mutants and Comparison of Immuno-Stimulatory Activities of Their Lipopolysaccharides. Mar. Drugs 2013, 11, 363-376. https://doi.org/10.3390/md11020363
Han Y, Li Y, Chen J, Tan Y, Guan F, Wang X. Construction of Monophosphoryl Lipid A Producing Escherichia coli Mutants and Comparison of Immuno-Stimulatory Activities of Their Lipopolysaccharides. Marine Drugs. 2013; 11(2):363-376. https://doi.org/10.3390/md11020363
Chicago/Turabian StyleHan, Yaning, Ye Li, Jiuzhou Chen, Yanzhen Tan, Feng Guan, and Xiaoyuan Wang. 2013. "Construction of Monophosphoryl Lipid A Producing Escherichia coli Mutants and Comparison of Immuno-Stimulatory Activities of Their Lipopolysaccharides" Marine Drugs 11, no. 2: 363-376. https://doi.org/10.3390/md11020363
APA StyleHan, Y., Li, Y., Chen, J., Tan, Y., Guan, F., & Wang, X. (2013). Construction of Monophosphoryl Lipid A Producing Escherichia coli Mutants and Comparison of Immuno-Stimulatory Activities of Their Lipopolysaccharides. Marine Drugs, 11(2), 363-376. https://doi.org/10.3390/md11020363