
No Association between Single Nucleotide Polymorphisms (SNPs) of the Interferon-Induced Transmembrane Protein 3 (IFITM3) Gene and the Susceptibility of Alzheimer’s Disease (AD)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Genomic DNA Extraction
2.3. Polymerase Chain Reaction (PCR)
2.4. Amplicon Sequencing and Genotyping
2.5. Statistical Analysis
3. Results
3.1. Subject Description
3.2. Evaluation of the Association of the Susceptibility of AD with the Genotype, Allele and Haplotype Frequencies of the IFITM3 Gene in the Korean Population
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, B.H. In Silico Evaluation of Acetylation Mimics in the 27 Lysine Residues of Human Tau Protein. Curr. Alzheimer Res. 2019, 16, 379–387. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, B.H. Identification of Somatic Mutations in Dementia-related Genes in Cancer Patients. Curr. Alzheimer Res. 2020, 17, 835–844. [Google Scholar] [CrossRef]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gu, B.J.; Masters, C.L.; Wang, Y.J. A systemic view of Alzheimer disease-insights from amyloid-beta metabolism beyond the brain. Nat. Rev. Neurol. 2017, 13, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Liu, F.; Gong, C.X.; Grundke-Iqbal, I. Tau in Alzheimer disease and related tauopathies. Curr. Alzheimer Res. 2010, 7, 656–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, H.; Jia, L.; Jia, J. Promoter polymorphisms which modulate APP expression may increase susceptibility to Alzheimer’s disease. Neurobiol. Aging 2008, 29, 194–202. [Google Scholar] [CrossRef]
- Bird, T.D. Early-Onset Familial Alzheimer Disease. In GeneReviews((R)); Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A., Eds.; NCBI Bookshelf Site: Seattle, WA, USA, 1993. [Google Scholar]
- Beck, J.A.; Poulter, M.; Campbell, T.A.; Uphill, J.B.; Adamson, G.; Geddes, J.F.; Revesz, T.; Davis, M.B.; Wood, N.W.; Collinge, J.; et al. Somatic and germline mosaicism in sporadic early-onset Alzheimer’s disease. Hum. Mol. Genet. 2004, 13, 1219–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanoiselee, H.M.; Nicolas, G.; Wallon, D.; Rovelet-Lecrux, A.; Lacour, M.; Rousseau, S.; Richard, A.C.; Pasquier, F.; Rollin-Sillaire, A.; Martinaud, O.; et al. APP, PSEN1, and PSEN2 mutations in early-onset Alzheimer disease: A genetic screening study of familial and sporadic cases. PLoS Med. 2017, 14, e1002270. [Google Scholar] [CrossRef] [Green Version]
- Chavez-Gutierrez, L.; Bammens, L.; Benilova, I.; Vandersteen, A.; Benurwar, M.; Borgers, M.; Lismont, S.; Zhou, L.; Van Cleynenbreugel, S.; Esselmann, H.; et al. The mechanism of gamma-Secretase dysfunction in familial Alzheimer disease. EMBO J. 2012, 31, 2261–2274. [Google Scholar] [CrossRef]
- Houlden, H.; Crook, R.; Dolan, R.J.; McLaughlin, J.; Revesz, T.; Hardy, J. A novel presenilin mutation (M233V) causing very early onset Alzheimer’s disease with Lewy bodies. Neurosci. Lett. 2001, 313, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, R.J., 3rd; Shen, J. Presenilin-1 mutations and Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2017, 114, 629–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zani, A.; Yount, J.S. Antiviral Protection by IFITM3 In Vivo. Curr. Clin. Microbiol. Rep. 2018, 5, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Jeong, B.H. Ethnic variation in risk genotypes based on single nucleotide polymorphisms (SNPs) of the interferon-inducible transmembrane 3 (IFITM3) gene, a susceptibility factor for pandemic 2009 H1N1 influenza A virus. Immunogenetics 2020, 72, 447–453. [Google Scholar] [CrossRef]
- Everitt, A.R.; Clare, S.; Pertel, T.; John, S.P.; Wash, R.S.; Smith, S.E.; Chin, C.R.; Feeley, E.M.; Sims, J.S.; Adams, D.J.; et al. IFITM3 restricts the morbidity and mortality associated with influenza. Nature 2012, 484, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Allen, E.K.; Randolph, A.G.; Bhangale, T.; Dogra, P.; Ohlson, M.; Oshansky, C.M.; Zamora, A.E.; Shannon, J.P.; Finkelstein, D.; Dressen, A.; et al. SNP-mediated disruption of CTCF binding at the IFITM3 promoter is associated with risk of severe influenza in humans. Nat. Med. 2017, 23, 975–983. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, M.J.; Jeong, B.H. Strong association of regulatory single nucleotide polymorphisms (SNPs) of the IFITM3 gene with influenza H1N1 2009 pandemic virus infection. Cell. Mol. Immunol. 2020, 17, 662–664. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, B.H. No Correlation of the Disease Severity of Influenza A Virus Infection with the rs12252 Polymorphism of the Interferon-Induced Transmembrane Protein 3 Gene. Intervirology 2017, 60, 69–74. [Google Scholar] [CrossRef]
- Kim, Y.C.; Jeong, M.J.; Jeong, B.H. Genetic association between the rs12252 SNP of the Interferon-Induced transmembrane protein gene and Influenza A Virus Infection in the Korean population. Mol. Cell. Toxicol. 2021, 17, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.Y.; Frost, G.R.; Wu, X.; Crump, C.; Pan, S.J.; Wong, E.; Barros, M.; Li, T.; Nie, P.; Zhai, Y.; et al. The innate immunity protein IFITM3 modulates gamma-secretase in Alzheimer’s disease. Nature 2020, 586, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Kruglyak, L.; Nickerson, D.A. Variation is the spice of life. Nat. Genet. 2001, 27, 234–236. [Google Scholar] [CrossRef]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef]
- Correani, V.; Di Francesco, L.; Mignogna, G.; Fabrizi, C.; Leone, S.; Giorgi, A.; Passeri, A.; Casata, R.; Fumagalli, L.; Maras, B.; et al. Plasma Membrane Protein Profiling in Beta-Amyloid-Treated Microglia Cell Line. Proteomics 2017, 17, 1600439. [Google Scholar] [CrossRef]
- Xu, Y.; Jin, M.Z.; Yang, Z.Y.; Jin, W.L. Microglia in neurodegenerative diseases. Neural Regen. Res. 2021, 16, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Zhao, Y.; Li, N.; Peng, Y.C.; Giannoulatou, E.; Jin, R.H.; Yan, H.P.; Wu, H.; Liu, J.H.; Liu, N.; et al. Interferon-induced transmembrane protein-3 genetic variant rs12252-C is associated with severe influenza in Chinese individuals. Nat. Commun. 2013, 4, 1418. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Xiao, M.; Yang, J.; Chen, Y.K.; Bai, T.; Tang, X.J.; Shu, Y.L. Association between rs12252 and influenza susceptibility and severity: An updated meta-analysis. Epidemiol. Infect. 2019, 147, E39. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Cao, B.; Ke, C.; Lu, H.; Hu, Y.; Tam, C.H.T.; Ma, R.C.W.; Guan, D.; Zhu, Z.; Li, H.; et al. IFITM3, TLR3, and CD55 Gene SNPs and Cumulative Genetic Risks for Severe Outcomes in Chinese Patients with H7N9/H1N1pdm09 Influenza. J. Infect. Dis. 2017, 216, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Mills, T.C.; Rautanen, A.; Elliott, K.S.; Parks, T.; Naranbhai, V.; Ieven, M.M.; Butler, C.C.; Little, P.; Verheij, T.; Garrard, C.S.; et al. IFITM3 and susceptibility to respiratory viral infections in the community. J. Infect. Dis. 2014, 209, 1028–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Characteristics | Cases | Controls | |
|---|---|---|---|
| Number | 177 | 233 | |
| Age | 73.19 ± 7.20 | 63.75 ± 9.74 | |
| Sex (n, %) | Male | 49 | 103 |
| Female | 128 | 129 | |
| APOE | E2/E3 | 15 (8.5) | 16 (7.1) |
| E2/E4 | 7 (4) | 1 (0.4) | |
| E3/E3 | 107 (60.5) | 83 (37.2) | |
| E3/E4 | 39 (22) | 12 (5.3) | |
| No data | 9 (5) | 111 (50) |
| Variants | Genotype Frequency, n (%) | p-Value | Allele Frequency, n (%) | p-Value | HWE | ||||
|---|---|---|---|---|---|---|---|---|---|
| CC | CG | GG | C | G | |||||
| c.-223C > G | Controls | 232 (1) | 1 (0) | 0 (0) | 465 (1) | 1 (0) | 0.9738 | ||
| Rs1297064916 | Patients | 177 (1) | 0 (0) | 0 (0) | 1.0 | 354 (1) | 0 (0) | 1.0 | - |
| GG | GT | TT | G | T | |||||
| c.-204G > T | Controls | 91 (0.39) | 109 (0.47) | 33 (0.14) | 291 (0.62) | 175 (0.38) | 0.9686 | ||
| Rs3888188 | Patients | 78 (0.44) | 78 (0.44) | 21 (0.12) | 0.5546 | 234 (0.66) | 120 (0.34) | 0.2800 | 0.8245 |
| TT | TC | CC | T | C | |||||
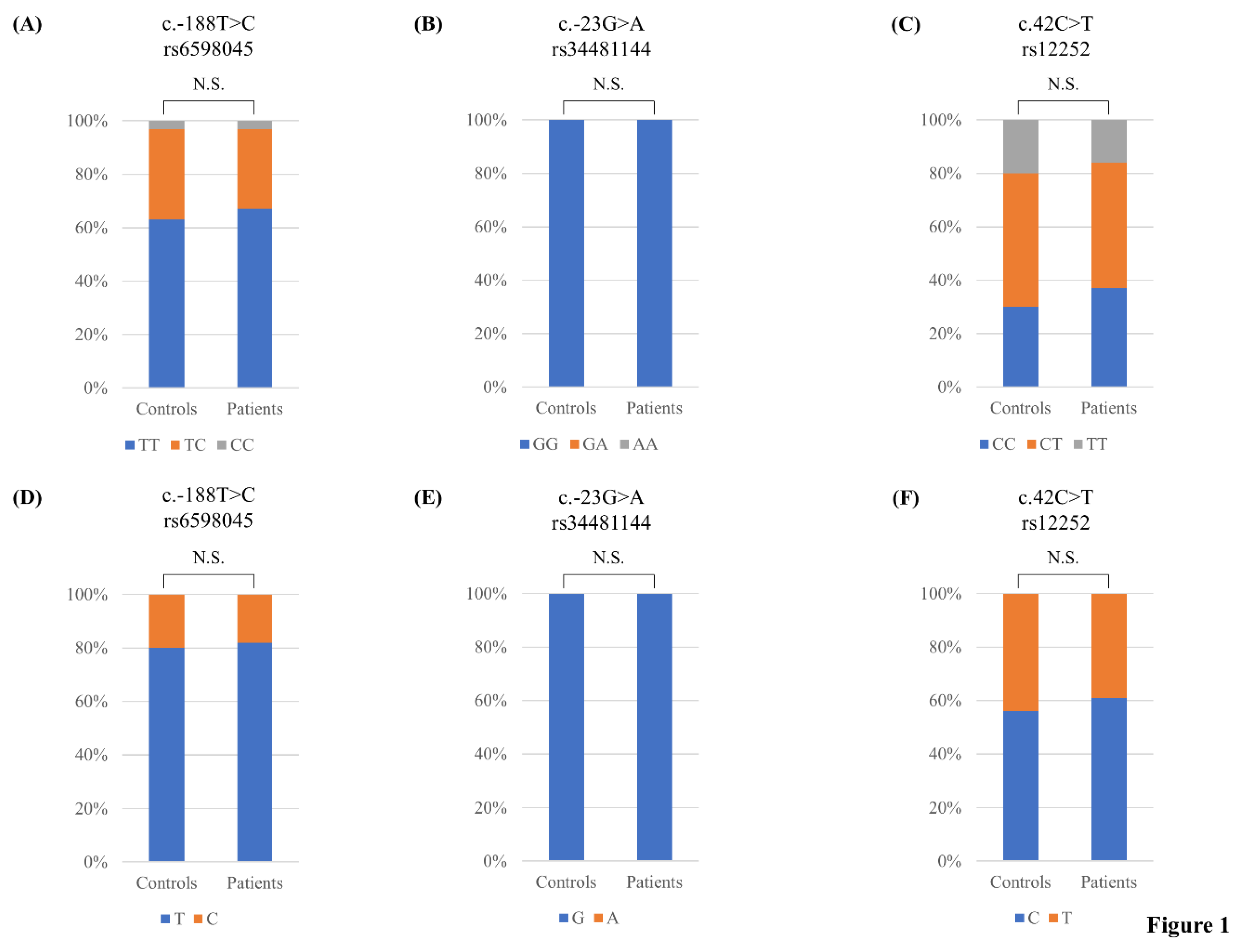
| c.-188T > C | Controls | 146 (0.63) | 80 (0.34) | 7 (0.03) | 372 (0.80) | 94 (0.2) | 0.3129 | ||
| Rs6598045 | Patients | 118 (0.67) | 53 (0.3) | 6 (0.03) | 0.6390 | 289 (0.82) | 65 (0.18) | 0.5161 | 0.9870 |
| TT | TC | CC | T | C | |||||
| c.-181T > C | Controls | 135 (0.58) | 84 (0.36) | 14 (0.06) | 354 (0.76) | 112 (0.24) | 0.8461 | ||
| Rs7478728 | Patients | 118 (0.67) | 48 (0.27) | 11 (0.6) | 0.1540 | 284 (0.8) | 70 (0.2) | 0.1459 | 0.0532 |
| AA | AC | CC | A | C | |||||
| c.-178A > C | Controls | 136 (0.58) | 83 (0.36) | 14 (0.06) | 355 (0.76) | 111 (0.24) | 0.7782 | ||
| Rs71452596 | Patients | 118 (0.67) | 48 (0.27) | 11 (0.06) | 0.1826 | 284 (0.8) | 70 (0.2) | 0.1665 | 0.0532 |
| TT | TC | CC | T | C | |||||
| c.-175T > C | Controls | 136 (0.59) | 82 (0.35) | 14 (0.06) | 354 (0.76) | 110 (0.24) | 0.7271 | ||
| Rs7479267 | Patients | 118 (0.67) | 48 (0.17) | 11 (0.06) | 0.2030 | 284 (0.8) | 70 (0.2) | 0.1785 | 0.0532 |
| CC | CT | TT | C | T | |||||
| c.-128C > T | Controls | 229 (0.98) | 4 (0.02) | 0 (0) | 462 (0.99) | 4 (0.01) | 0.8949 | ||
| Rs536077498 | Patients | 173 (0.98) | 4 (0.02) | 0 (0) | 0.7307 | 350 (0.99) | 4 (0.01) | 0.7318 | 0.8792 |
| GG | GA | AA | G | A | |||||
| c.-23G > A | Controls | 233 (1) | 0 (0) | 0 (0) | 466 (1) | 0 (0) | - | ||
| Rs34481144 | Patients | 177 (1) | 0 (0) | 0 (0) | - | 354 (1) | 0 (0) | - | - |
| Wt/Wt | Wt/Del | Del/Del | Wt | Del | |||||
| c.-13_-4Del | Controls | 232 (1) | 1 (1) | 0 (0) | 465 (1) | 1 (0) | 0.9738 | ||
| TTCGCTGGAC | Patients | 177 (1) | 0 (0) | 0 (0) | 1.0 | 354 (1) | 0 (0) | 1.0 | - |
| Rs773536963 | |||||||||
| CC | CT | TT | C | T | |||||
| c.42C > T | Controls | 73 (0.3) | 116 (0.5) | 44 (0.2) | 262 (0.56) | 204 (0.44) | 0.8621 | ||
| Rs12252 | Patients | 66 (0.37) | 83 (0.47) | 28 (0.16) | 0.4138 | 215 (0.61) | 139 (0.39) | 0.1946 | 0.8228 |
| CC | CT | TT | C | T | |||||
| c.165C > T | Controls | 231 (0.99) | 2 (0.01) | 0 (0) | 464 (1) | 2 (0) | 0.9475 | ||
| Rs11553885 | Patients | 175 (0.99) | 2 (0.01) | 0 (0) | 1.0 | 352 (1) | 2 (0) | 1.0 | 0.9397 |
| Haplotype | Frequency | p-Value | |
|---|---|---|---|
| Control (n = 354) | Patients (n = 466) | ||
| CGTTATCwtCC | 193 (0.545) | 272 (0.583) | 0.2705 |
| CTTCCCCwtTC | 82 (0.232) | 90 (0.193) | 0.1798 |
| CTCTATCwtTC | 49 (0.139) | 63 (0.135) | 0.8940 |
| CGCTATCwtTC | 20 (0.056) | 21 (0.045) | 0.4569 |
| Others | 10 (0.028) | 20 (0.044) | 0.2677 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, S.-Y.; Kim, Y.-C.; Jeong, B.-H. No Association between Single Nucleotide Polymorphisms (SNPs) of the Interferon-Induced Transmembrane Protein 3 (IFITM3) Gene and the Susceptibility of Alzheimer’s Disease (AD). Medicina 2022, 58, 55. https://doi.org/10.3390/medicina58010055
Won S-Y, Kim Y-C, Jeong B-H. No Association between Single Nucleotide Polymorphisms (SNPs) of the Interferon-Induced Transmembrane Protein 3 (IFITM3) Gene and the Susceptibility of Alzheimer’s Disease (AD). Medicina. 2022; 58(1):55. https://doi.org/10.3390/medicina58010055
Chicago/Turabian StyleWon, Sae-Young, Yong-Chan Kim, and Byung-Hoon Jeong. 2022. "No Association between Single Nucleotide Polymorphisms (SNPs) of the Interferon-Induced Transmembrane Protein 3 (IFITM3) Gene and the Susceptibility of Alzheimer’s Disease (AD)" Medicina 58, no. 1: 55. https://doi.org/10.3390/medicina58010055
APA StyleWon, S.-Y., Kim, Y.-C., & Jeong, B.-H. (2022). No Association between Single Nucleotide Polymorphisms (SNPs) of the Interferon-Induced Transmembrane Protein 3 (IFITM3) Gene and the Susceptibility of Alzheimer’s Disease (AD). Medicina, 58(1), 55. https://doi.org/10.3390/medicina58010055