
Donor Variability Alters the Characteristics of Human Brain Microvascular Endothelial Cells
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Immunocytochemistry
2.3. Western Blotting
2.4. Assessment of In Vitro Blood–Brain Barrier Integrity and Function
2.5. Tube Formation Assay
2.6. Senescence-Associated β-Galactosidase Activity
2.7. Statistical Analyses
3. Results
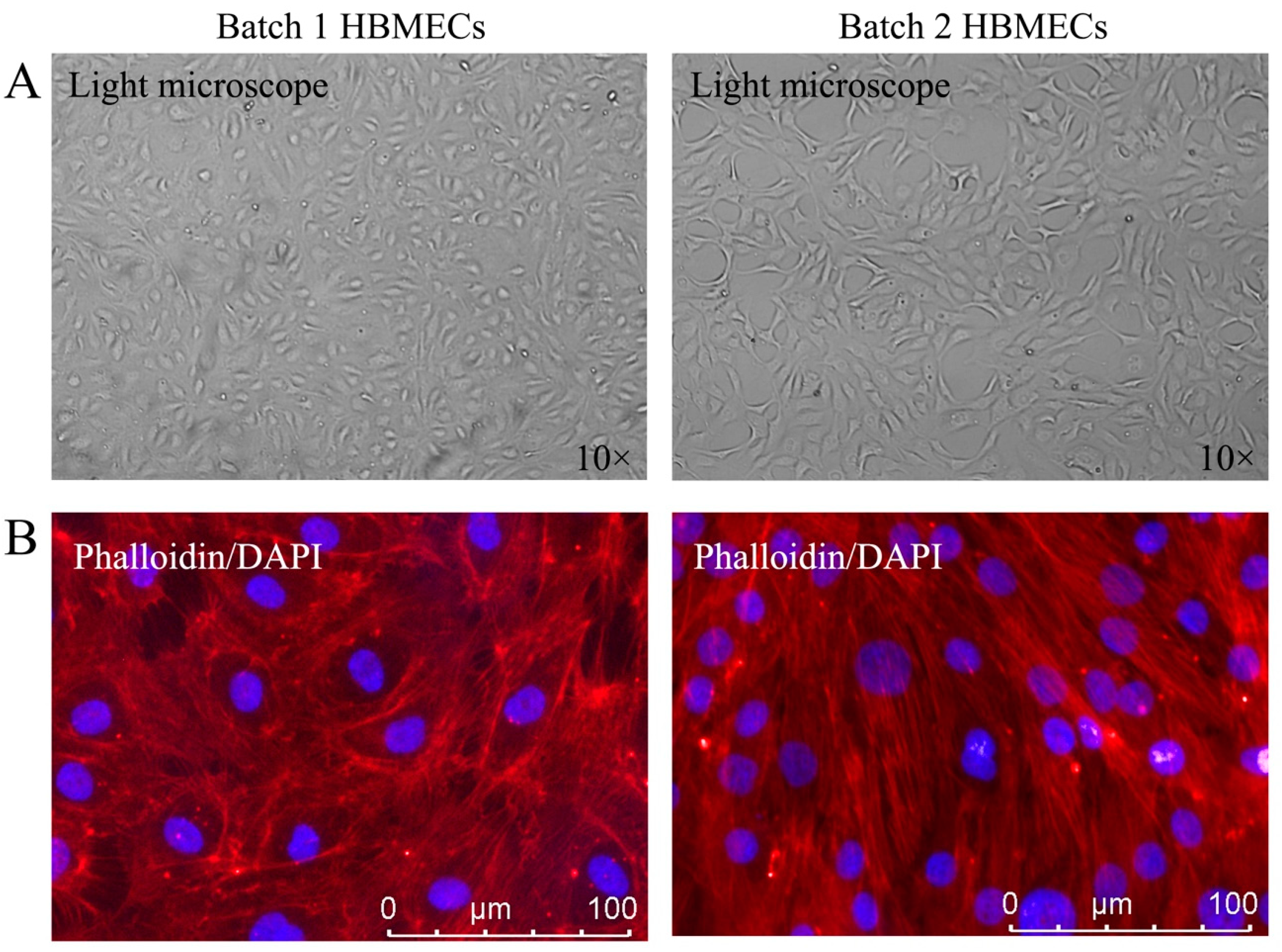
3.1. HBMECs Display Different Phenotypes
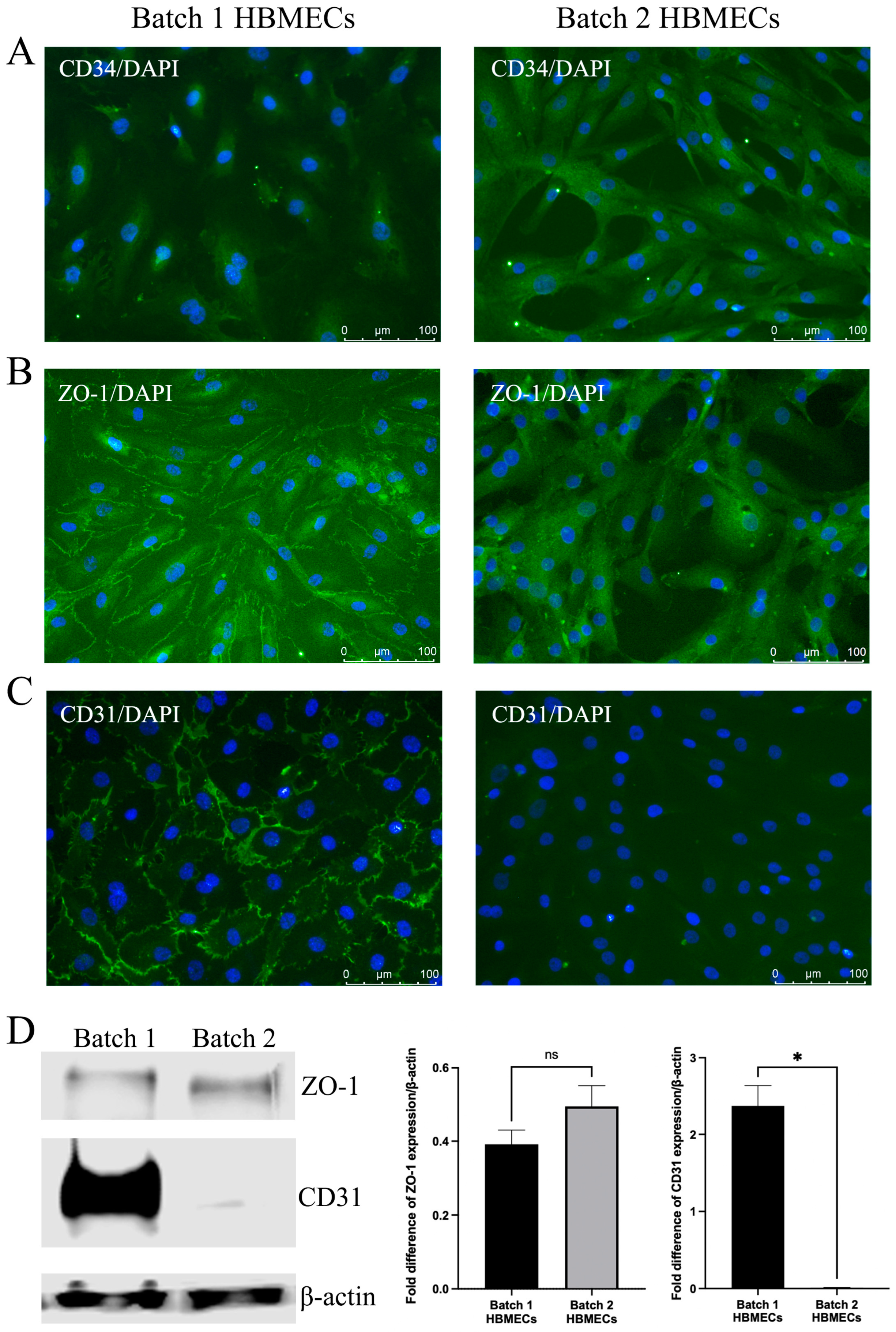
3.2. HBMECs Express Endothelial Markers Differently
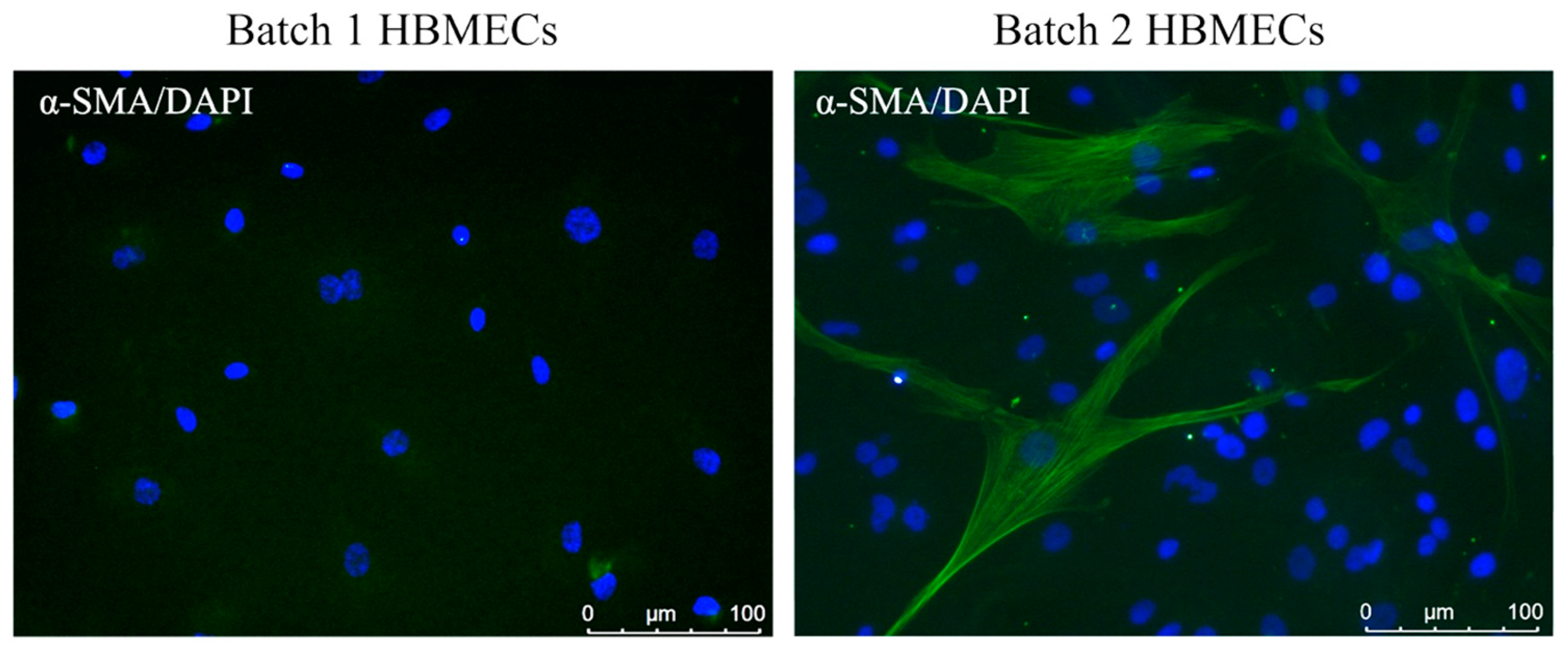
3.3. Scrutiny of Alpha-Smooth Muscle Actin Expression in Endothelial Cells
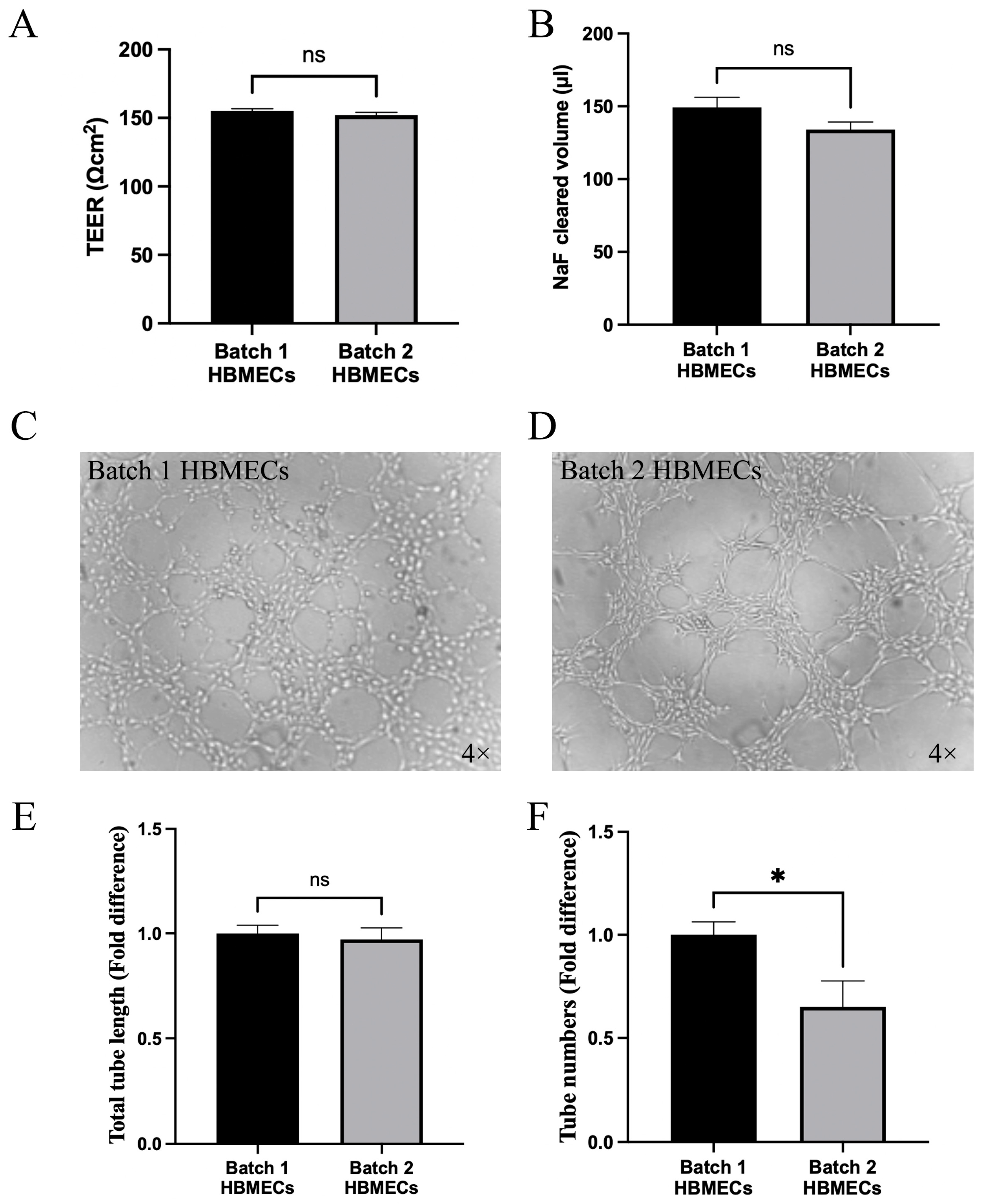
3.4. Effect of Changes in Endothelial Cell Characteristics on Functional Activity
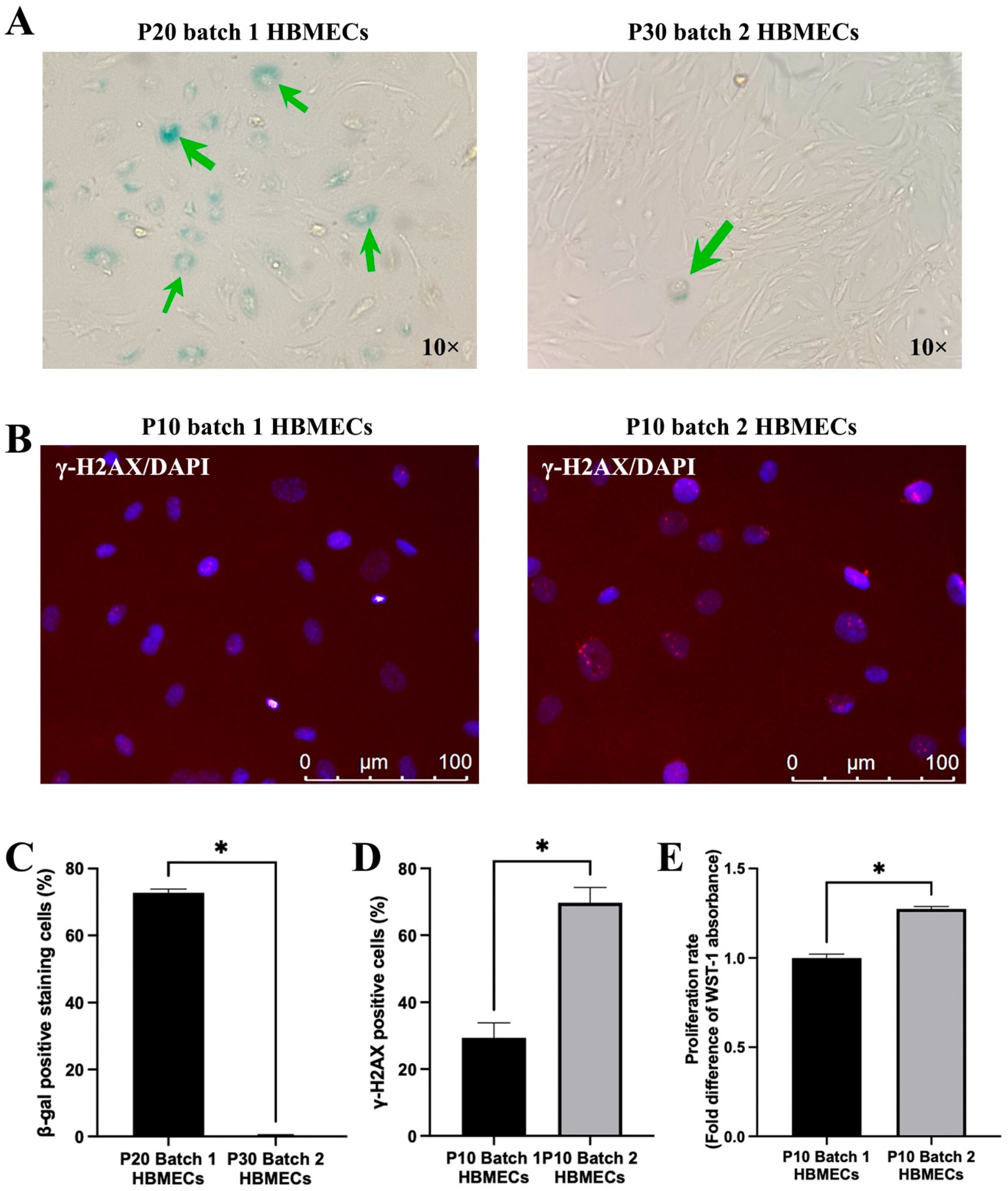
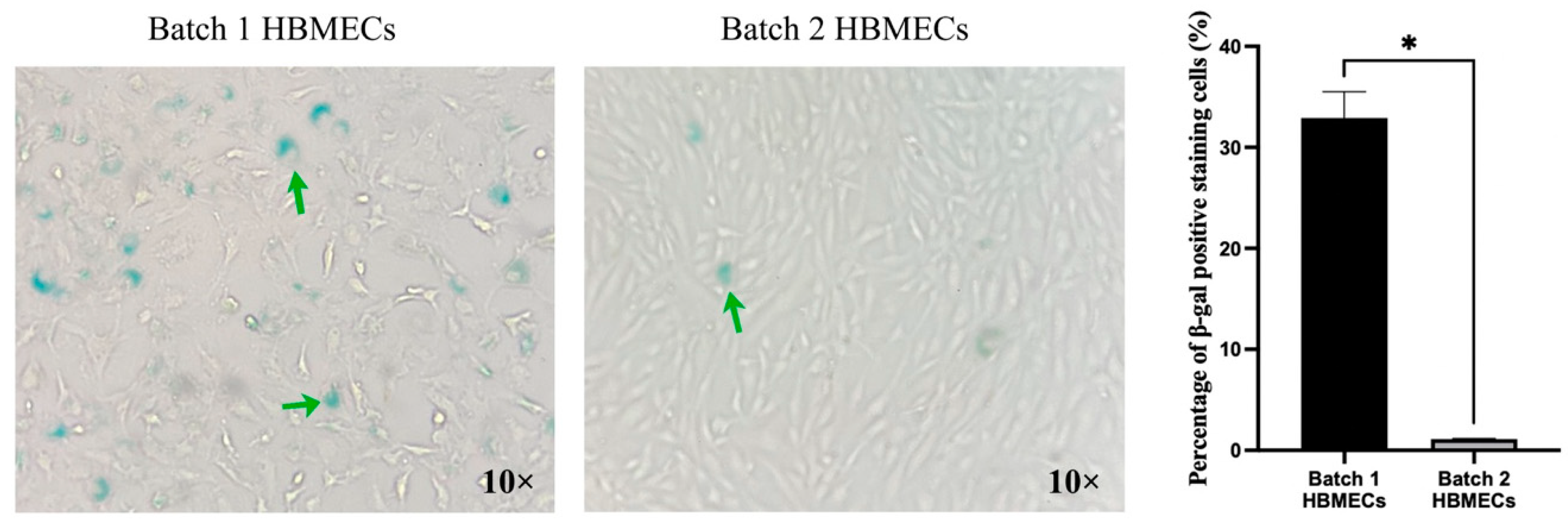
3.5. Endothelial Cells Respond to Senescence-Inducing Stimuli Differently
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sandoo, A.; van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The endothelium and its role in regulating vascular tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Dessalles, C.A.; Leclech, C.; Castagnino, A.; Barakat, A.I. Integration of substrate- and flow-derived stresses in endothelial cell mechanobiology. Commun. Biol. 2021, 4, 764. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.; Shao, B.; Bayraktutan, U. PKC-beta exacerbates in vitro brain barrier damage in hyperglycemic settings via regulation of RhoA/Rho-kinase/MLC2 pathway. J. Cereb. Blood Flow Metab. 2013, 33, 1928–1936. [Google Scholar] [CrossRef] [PubMed]
- Kadry, H.; Noorani, B.; Cucullo, L. A blood-brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 2020, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Piwocka, O.; Musielak, M.; Piotrowski, I.; Suchorska, W.M.; Trzeciak, T. From Donor to the Lab: A Fascinating Journey of Primary Cell Lines. Front. Cell Dev. Biol. 2021, 9, 711381. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef]
- Hashmat, A.; Ya, J.; Kadir, R.; Alwjwaj, M.; Bayraktutan, U. Hyperglycaemia perturbs blood-brain barrier integrity through its effects on endothelial cell characteristics and function. Tissue Barriers 2024, 2350821. [Google Scholar] [CrossRef] [PubMed]
- Ya, J.; Whitby, A.; Bayraktutan, U. Metabolites and Metabolic Functional Changes-Potential Markers for Endothelial Cell Senescence. Biomolecules 2024, 14, 1476. [Google Scholar] [CrossRef]
- Supp, D.M.; Hahn, J.M.; Combs, K.A.; McFarland, K.L.; Powell, H.M. Isolation and feeder-free primary culture of four cell types from a single human skin sample. STAR Protoc. 2022, 3, 101172. [Google Scholar] [CrossRef]
- Kadir, R.R.A.; Alwjwaj, M.; Bayraktutan, U. Establishment of an In Vitro Model of Human Blood-Brain Barrier to Study the Impact of Ischemic Injury. Methods Mol. Biol. 2022, 2492, 143–155. [Google Scholar] [CrossRef]
- Moonen, J.R.; Lee, E.S.; Schmidt, M.; Maleszewska, M.; Koerts, J.A.; Brouwer, L.A.; van Kooten, T.G.; van Luyn, M.J.; Zeebregts, C.J.; Krenning, G.; et al. Endothelial-to-mesenchymal transition contributes to fibro-proliferative vascular disease and is modulated by fluid shear stress. Cardiovasc. Res. 2015, 108, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Qadan, M.A.; Piuzzi, N.S.; Boehm, C.; Bova, W.; Moos, M., Jr.; Midura, R.J.; Hascall, V.C.; Malcuit, C.; Muschler, G.F. Variation in primary and culture-expanded cells derived from connective tissue progenitors in human bone marrow space, bone trabecular surface and adipose tissue. Cytotherapy 2018, 20, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Chan, Y.K.; Goodman, D.B.; Guo, X.; Chavez, A.; Lim, E.T.; Church, G.M. Enabling multiplexed testing of pooled donor cells through whole-genome sequencing. Genome Med. 2018, 10, 31. [Google Scholar] [CrossRef]
- Kent, D.G. Battle of the sexes: Understanding donor:recipient sex differences in transplantation biology. Hemasphere 2024, 8, e70000. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Giri, A.K.; Aittokallio, T. Fully-automated and ultra-fast cell-type identification using specific marker combinations from single-cell transcriptomic data. Nat. Commun. 2022, 13, 1246. [Google Scholar] [CrossRef]
- Alizadeh, E.; Castle, J.; Quirk, A.; Taylor, C.D.L.; Xu, W.; Prasad, A. Cellular morphological features are predictive markers of cancer cell state. Comput. Biol. Med. 2020, 126, 104044. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.M.; Izumo, S. Mechanism of endothelial cell shape change and cytoskeletal remodeling in response to fluid shear stress. J. Cell Sci. 1996, 109 Pt 4, 713–726. [Google Scholar] [CrossRef]
- Vion, A.C.; Perovic, T.; Petit, C.; Hollfinger, I.; Bartels-Klein, E.; Frampton, E.; Gordon, E.; Claesson-Welsh, L.; Gerhardt, H. Endothelial Cell Orientation and Polarity Are Controlled by Shear Stress and VEGF Through Distinct Signaling Pathways. Front. Physiol. 2020, 11, 623769. [Google Scholar] [CrossRef]
- Liang, J.; Gu, S.; Mao, X.; Tan, Y.; Wang, H.; Li, S.; Zhou, Y. Endothelial Cell Morphology Regulates Inflammatory Cells Through MicroRNA Transferred by Extracellular Vesicles. Front. Bioeng. Biotechnol. 2020, 8, 369. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.; Gossen, M.; Lendlein, A.; Jung, F. Venous and Arterial Endothelial Cells from Human Umbilical Cords: Potential Cell Sources for Cardiovascular Research. Int. J. Mol. Sci. 2021, 22, 978. [Google Scholar] [CrossRef] [PubMed]
- Wijelath, E.S.; Carlsen, B.; Cole, T.; Chen, J.; Kothari, S.; Hammond, W.P. Oncostatin M induces basic fibroblast growth factor expression in endothelial cells and promotes endothelial cell proliferation, migration and spindle morphology. J. Cell Sci. 1997, 110 Pt 7, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Clere, N.; Renault, S.; Corre, I. Endothelial-to-Mesenchymal Transition in Cancer. Front. Cell Dev. Biol. 2020, 8, 747. [Google Scholar] [CrossRef] [PubMed]
- Gibieza, P.; Petrikaite, V. The regulation of actin dynamics during cell division and malignancy. Am. J. Cancer Res. 2021, 11, 4050–4069. [Google Scholar] [PubMed]
- Yadunandanan Nair, N.; Samuel, V.; Ramesh, L.; Marib, A.; David, D.T.; Sundararaman, A. Actin cytoskeleton in angiogenesis. Biol. Open 2022, 11, bio058899. [Google Scholar] [CrossRef]
- Weber, C.R. Dynamic properties of the tight junction barrier. Ann. N. Y. Acad. Sci. 2012, 1257, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical expression of endothelial markers CD31, CD34, von Willebrand factor, and Fli-1 in normal human tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Coracina, A.; Baesso, I.; Agostini, C.; Tiengo, A.; Avogaro, A.; de Kreutzenberg, S.V. Peripheral blood CD34+KDR+ endothelial progenitor cells are determinants of subclinical atherosclerosis in a middle-aged general population. Stroke 2006, 37, 2277–2282. [Google Scholar] [CrossRef]
- Zhang, Z.; Gan, Q.; Han, J.; Tao, Q.; Qiu, W.Q.; Madri, J.A. CD31 as a probable responding and gate-keeping protein of the blood-brain barrier and the risk of Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2023, 43, 1027–1041. [Google Scholar] [CrossRef] [PubMed]
- Venkataramani, V.; Kuffer, S.; Cheung, K.C.P.; Jiang, X.; Trumper, L.; Wulf, G.G.; Strobel, P. CD31 Expression Determines Redox Status and Chemoresistance in Human Angiosarcomas. Clin. Cancer Res. 2018, 24, 460–473. [Google Scholar] [CrossRef]
- Kadir, R.R.A.; Alwjwaj, M.; Bayraktutan, U. Protein kinase C-beta distinctly regulates blood-brain barrier-forming capacity of Brain Microvascular endothelial cells and outgrowth endothelial cells. Metab. Brain Dis. 2022, 37, 1815–1827. [Google Scholar] [CrossRef]
- Lu, X.; Gong, J.; Dennery, P.A.; Yao, H. Endothelial-to-mesenchymal transition: Pathogenesis and therapeutic targets for chronic pulmonary and vascular diseases. Biochem. Pharmacol. 2019, 168, 100–107. [Google Scholar] [CrossRef]
- Reskiawan, A.K.R.; Alwjwaj, M.; Ahmad Othman, O.; Rakkar, K.; Sprigg, N.; Bath, P.M.; Bayraktutan, U. Inhibition of oxidative stress delays senescence and augments functional capacity of endothelial progenitor cells. Brain Res. 2022, 1787, 147925. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Akahori, T.; Komaki, M.; Li, Q.; Kanayasu-Toyoda, T.; Ishii-Watabe, A.; Kobayashi, A.; Yamaguchi, T.; Abe, M.; Amagasa, T.; et al. A comparison of the tube forming potentials of early and late endothelial progenitor cells. Exp. Cell Res. 2008, 314, 430–440. [Google Scholar] [CrossRef]
- Katsuta, E.; Sawant Dessai, A.; Ebos, J.M.; Yan, L.; Ouchi, T.; Takabe, K. H2AX mRNA expression reflects DNA repair, cell proliferation, metastasis, and worse survival in breast cancer. Am. J. Cancer Res. 2022, 12, 793–804. [Google Scholar] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ya, J.; Bayraktutan, U. Donor Variability Alters the Characteristics of Human Brain Microvascular Endothelial Cells. Curr. Issues Mol. Biol. 2025, 47, 73. https://doi.org/10.3390/cimb47020073
Ya J, Bayraktutan U. Donor Variability Alters the Characteristics of Human Brain Microvascular Endothelial Cells. Current Issues in Molecular Biology. 2025; 47(2):73. https://doi.org/10.3390/cimb47020073
Chicago/Turabian StyleYa, Jingyuan, and Ulvi Bayraktutan. 2025. "Donor Variability Alters the Characteristics of Human Brain Microvascular Endothelial Cells" Current Issues in Molecular Biology 47, no. 2: 73. https://doi.org/10.3390/cimb47020073
APA StyleYa, J., & Bayraktutan, U. (2025). Donor Variability Alters the Characteristics of Human Brain Microvascular Endothelial Cells. Current Issues in Molecular Biology, 47(2), 73. https://doi.org/10.3390/cimb47020073