
Anthocyanins and Anthocyanidins in the Management of Osteoarthritis: A Scoping Review of Current Evidence



Abstract
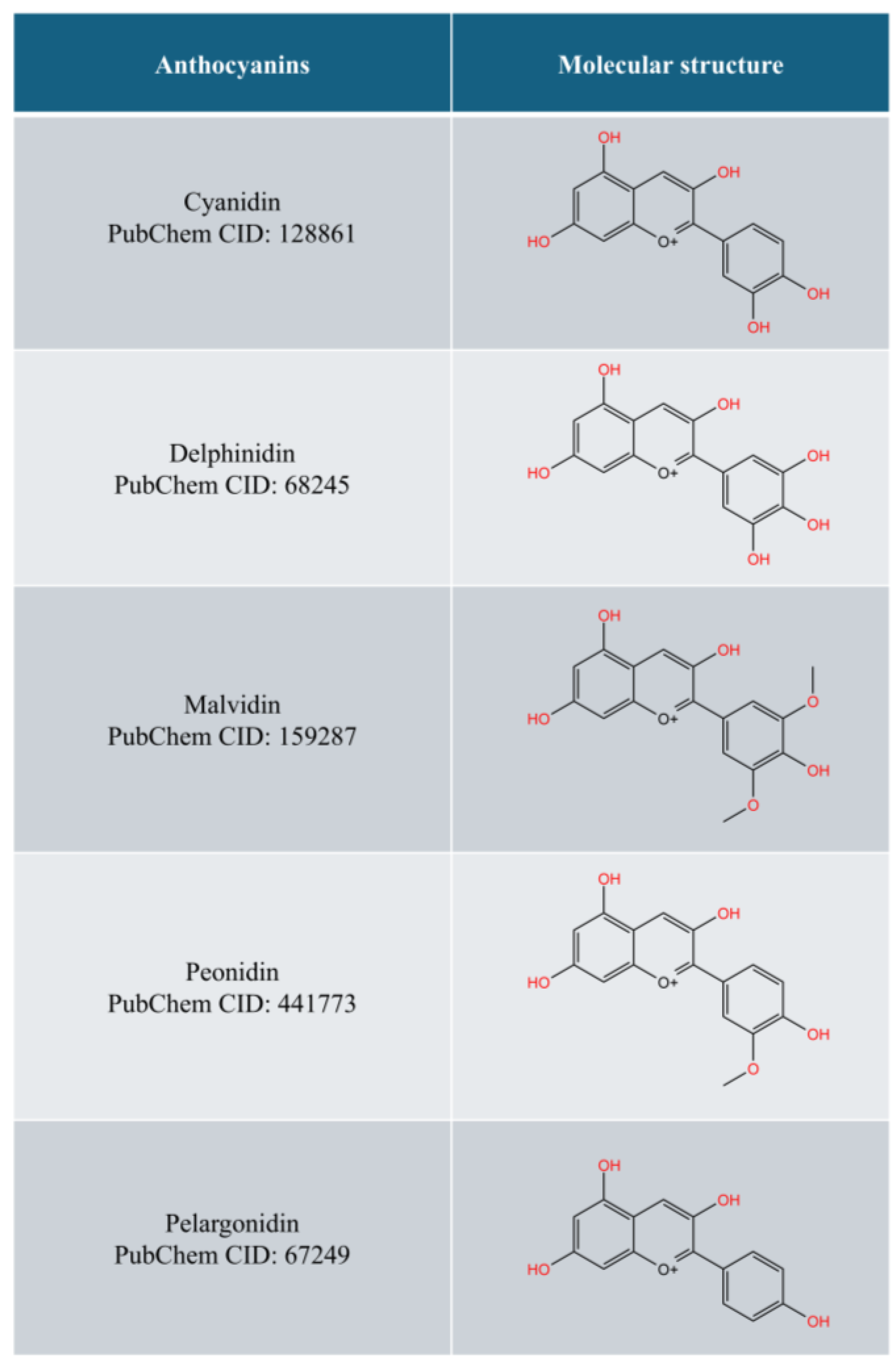
1. Introduction
2. Methods
2.1. Identifying the Research Question
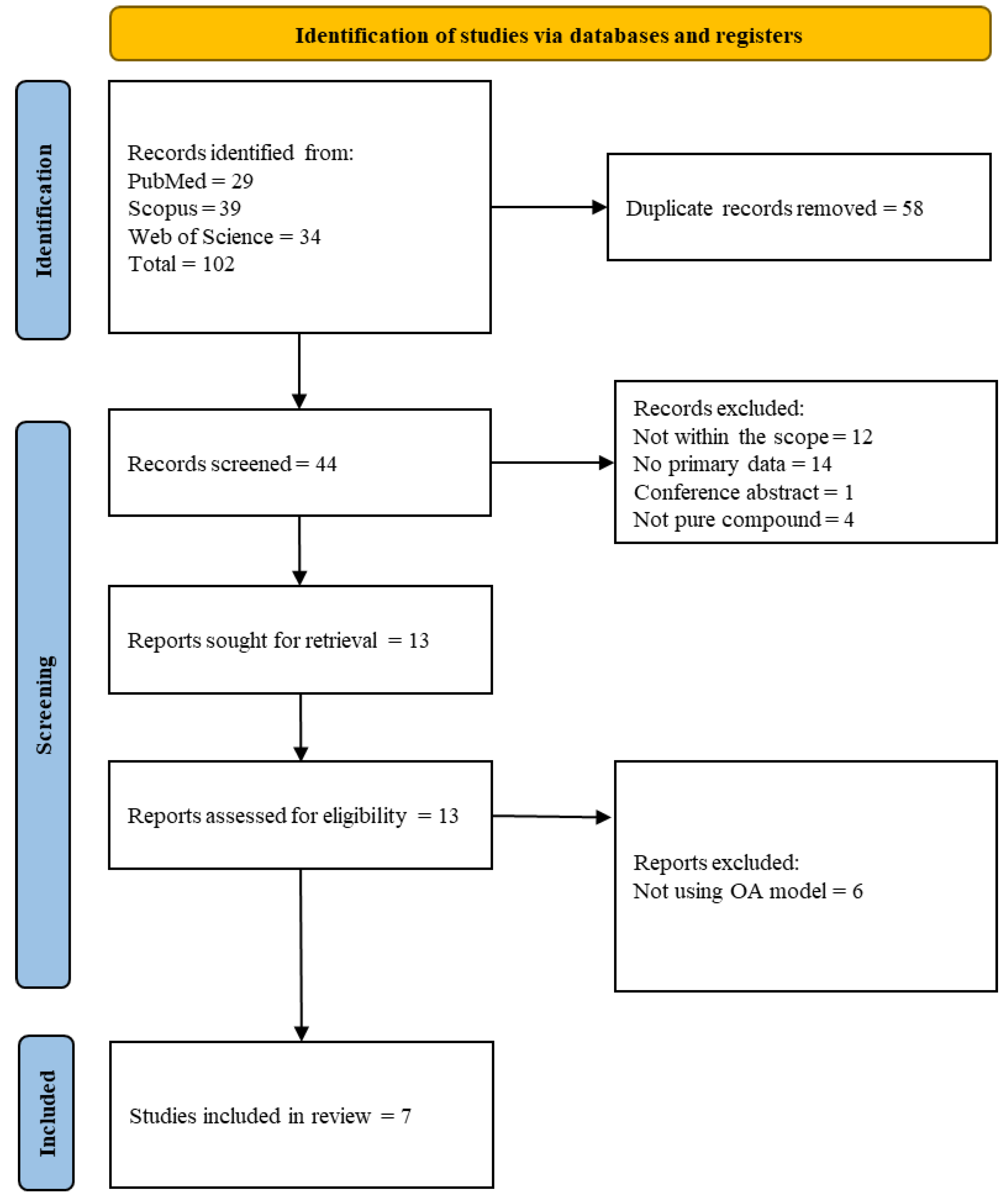
2.2. Identifying Relevant Studies
2.3. Study Selection
2.4. Charting the Data
2.5. Collating, Summarising, and Reporting the Results
3. Results
3.1. Study Characteristics
3.2. Effects on Chondrocyte Viability
3.3. Effects on Anabolic and Catabolic Processes of Chondrocytes
3.4. Effects of Cartilage Integrity
3.5. Effects on Pain Threshold
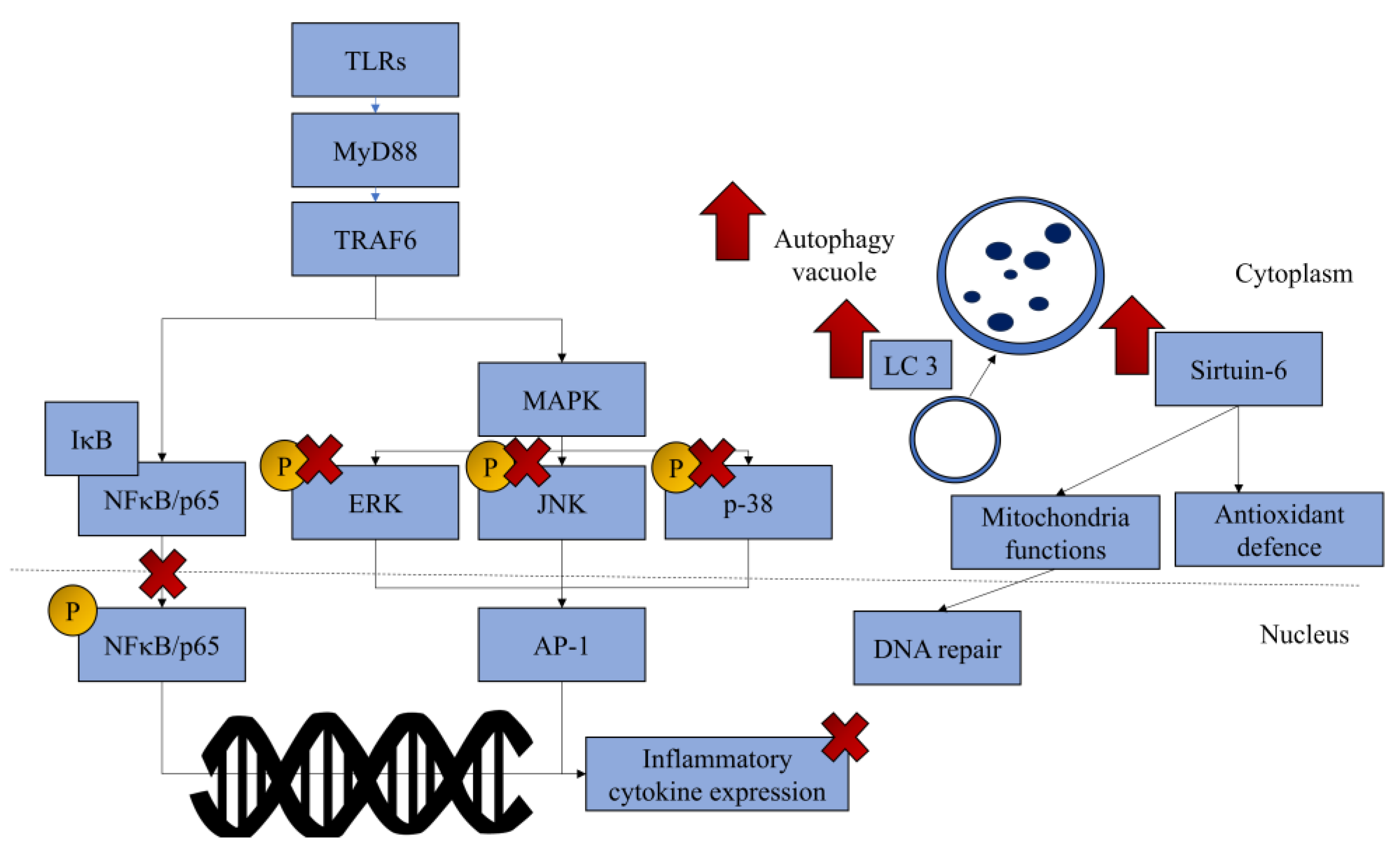
3.6. Mechanism of Actions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wenjian Zhao, Y.L.; Wang, Y.; Chin, K.-Y. Osteoarthritis: An integrative overview from pathogenesis to management. Malays. J. Pathol. 2024, 46, 369–378. [Google Scholar]
- Li, H.Z.; Liang, X.Z.; Sun, Y.Q.; Jia, H.F.; Li, J.C.; Li, G. Global, regional, and national burdens of osteoarthritis from 1990 to 2021: Findings from the 2021 global burden of disease study. Front. Med. 2024, 11, 1476853. [Google Scholar] [CrossRef]
- Nicholas, E.; Cheng, J.; Moley, P.J. Non-operative Treatment Options for Osteoarthritis in the Hip. HSS J. 2023, 19, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Cherniavskyi, V.V.; Baylo, A.E.; Onyshuk, L.O.; Tishchenko, V.V. Critical evaluation of the current role of SYSADOA in the management of osteoarthritis (literature review). Pain Jt. Spine 2024, 14, 96–105. [Google Scholar] [CrossRef]
- Atthakorn, J.; Chaturong, P. Complications after Total Knee Arthroplasty: Stiffness, Periprosthetic Joint Infection, and Periprosthetic Fracture. In Arthroplasty; Alessandro, Z., Hechmi, T., Eric, L., Eds.; IntechOpen: Rijeka, Croation, 2022; p. Ch. 8. [Google Scholar]
- Fang, S.; Zhang, B.; Xiang, W.; Zheng, L.; Wang, X.; Li, S.; Zhang, T.; Feng, D.; Gong, Y.; Wu, J.; et al. Natural products in osteoarthritis treatment: Bridging basic research to clinical applications. Chin. Med. 2024, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Chow, Y.Y.; Chin, K.Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediators Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharmacother. 2020, 129, 110452. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.Q.; Nie, L.Y.; Qian, L.N.; Zhao, K. Efficacy and Safety of Polyphenols for Osteoarthritis Treatment: A Meta-Analysis. Clin. Ther. 2021, 43, e241–e253.e242. [Google Scholar] [CrossRef]
- Arias, C.; Salazar, L.A. Autophagy and Polyphenols in Osteoarthritis: A Focus on Epigenetic Regulation. Int. J. Mol. Sci. 2021, 23, 421. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Wang, X.; Lin, X.; Mostafa, S.; Zou, H.; Wang, L.; Jin, B. Plant anthocyanins: Classification, biosynthesis, regulation, bioactivity, and health benefits. Plant Physiol. Biochem. 2024, 217, 109268. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef] [PubMed]
- Bonesi, M.; Leporini, M.; Tenuta, M.C.; Tundis, R. The Role of Anthocyanins in Drug Discovery: Recent Developments. Curr. Drug Discov. Technol. 2020, 17, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Frasca, G.; Panico, A.M.; Bonina, F.; Messina, R.; Rizza, L.; Musumeci, G.; Rapisarda, P.; Cardile, V. Involvement of inducible nitric oxide synthase and cyclooxygenase-2 in the anti-inflammatory effects of a red orange extract in human chondrocytes. Nat. Prod. Res. 2010, 24, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Chuntakaruk, H.; Kongtawelert, P.; Pothacharoen, P. Chondroprotective effects of purple corn anthocyanins on advanced glycation end products induction through suppression of NF-κB and MAPK signaling. Sci. Rep. 2021, 11, 1895. [Google Scholar] [CrossRef]
- Wongwichai, T.; Teeyakasem, P.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Anthocyanins and metabolites from purple rice inhibit IL-1β-induced matrix metalloproteinases expression in human articular chondrocytes through the NF-κB and ERK/MAPK pathway. Biomed. Pharmacother. 2019, 112, 108610. [Google Scholar] [CrossRef]
- Herrera-Bravo, J.; Beltrán, J.F.; Huard, N.; Saavedra, K.; Saavedra, N.; Alvear, M.; Lanas, F.; Salazar, L.A. Anthocyanins Found in Pinot Noir Waste Induce Target Genes Related to the Nrf2 Signalling in Endothelial Cells. Antioxidants 2022, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.M.; Ahmad, I.; Haqqi, T.M. Nrf2/ARE pathway attenuates oxidative and apoptotic response in human osteoarthritis chondrocytes by activating ERK1/2/ELK1-P70S6K-P90RSK signaling axis. Free Radic. Biol. Med. 2018, 116, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Semmarath, W.; Mapoung, S.; Umsumarng, S.; Arjsri, P.; Srisawad, K.; Thippraphan, P.; Yodkeeree, S.; Dejkriengkraikul, P. Cyanidin-3-O-glucoside and Peonidin-3-O-glucoside-Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS-CoV-2 Spike Glycoprotein S1-Induction In Vitro in A549 Lung Cells and THP-1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway. Nutrients 2022, 14, 2738. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef]
- Saulite, L.; Jekabsons, K.; Klavins, M.; Muceniece, R.; Riekstina, U. Effects of malvidin, cyanidin and delphinidin on human adipose mesenchymal stem cell differentiation into adipocytes, chondrocytes and osteocytes. Phytomedicine 2019, 53, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Arksey, H.; O’Malley, L. Scoping Studies: Towards a Methodological Framework. Int. J. Social. Res. Methodol. Theory Pract. 2005, 8, 19–32. [Google Scholar] [CrossRef]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, A.; Chen, D.; Haqqi, T.M. Delphinidin inhibits IL-1β-induced activation of NF-κB by modulating the phosphorylation of IRAK-1(Ser376) in human articular chondrocytes. Rheumatology 2013, 52, 998–1008. [Google Scholar] [CrossRef]
- Jiang, C.; Sun, Z.M.; Hu, J.N.; Jin, Y.; Guo, Q.; Xu, J.J.; Chen, Z.X.; Jiang, R.H.; Wu, Y.S. Cyanidin ameliorates the progression of osteoarthritis via the Sirt6/NF-κB axis in vitro and in vivo. Food Funct. 2019, 10, 5873–5885. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Li, H.; Luo, C.; Hu, W.; Weng, T.J.; Shuang, F. Pelargonidin ameliorates inflammatory response and cartilage degeneration in osteoarthritis via suppressing the NF-κB pathway. Arch. Biochem. Biophys. 2023, 743, 109668. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Shi, K.; Chen, G.; Shen, Y.; Pan, T. Malvidin attenuates pain and inflammation in rats with osteoarthritis by suppressing NF-κB signaling pathway. Inflamm. Res. 2017, 66, 1075–1084. [Google Scholar] [CrossRef]
- Lee, D.Y.; Park, Y.J.; Song, M.G.; Kim, D.R.; Zada, S.; Kim, D.H. Cytoprotective Effects of Delphinidin for Human Chondrocytes against Oxidative Stress through Activation of Autophagy. Antioxidants 2020, 9, 83. [Google Scholar] [CrossRef]
- Wang, B.W.; Jiang, Y.; Yao, Z.L.; Chen, P.S.; Yu, B.; Wang, S.N. Aucubin Protects Chondrocytes Against IL-1β-Induced Apoptosis In Vitro and Inhibits Osteoarthritis in Mice Model. Drug Des. Devel Ther. 2019, 13, 3529–3538. [Google Scholar] [CrossRef]
- Hwang, H.S.; Kim, H.A. Chondrocyte Apoptosis in the Pathogenesis of Osteoarthritis. Int. J. Mol. Sci. 2015, 16, 26035–26054. [Google Scholar] [CrossRef] [PubMed]
- Sumsuzzman, D.M.; Choi, J.; Khan, Z.A.; Kamenos, G.; Hong, Y. Melatonin Maintains Anabolic-Catabolic Equilibrium and Regulates Circadian Rhythm During Osteoarthritis Development in Animal Models: A Systematic Review and Meta-analysis. Front. Pharmacol. 2021, 12, 714974. [Google Scholar] [CrossRef]
- Arra, M.; Abu-Amer, Y. Cross-talk of inflammation and chondrocyte intracellular metabolism in osteoarthritis. Osteoarthritis Cartilage 2023, 31, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Ekeuku, S.O.; Ahmad, F.; Chin, K.-Y. Changes of Grip Strength, Articular Cartilage and Subchondral Bone in Monoiodoacetate-Induced Osteoarthritis in Rats. Sains Malays. 2022, 51, 3741–3754. [Google Scholar] [CrossRef]
- Ekeuku, S.O.; Nor Muhamad, M.L.; Aminuddin, A.A.; Ahmad, F.; Wong, S.K.; Mark-Lee, W.F.; Chin, K.-Y. Effects of emulsified and non-emulsified palm tocotrienol on bone and joint health in ovariectomised rats with monosodium iodoacetate-induced osteoarthritis. Biomed. Pharmacother. 2024, 170, 115998. [Google Scholar] [CrossRef]
- Yu, H.; Huang, T.; Lu, W.W.; Tong, L.; Chen, D. Osteoarthritis Pain. Int. J. Mol. Sci. 2022, 23, 4642. [Google Scholar] [CrossRef] [PubMed]
- Han, F.Y.; Brockman, D.A.; Nicholson, J.R.; Corradini, L.; Smith, M.T. Pharmacological characterization of the chronic phase of the monoiodoacetate-induced rat model of osteoarthritis pain in the knee joint. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1515–1522. [Google Scholar] [CrossRef]
- Bagaev, A.V.; Garaeva, A.Y.; Lebedeva, E.S.; Pichugin, A.V.; Ataullakhanov, R.I.; Ataullakhanov, F.I. Elevated pre-activation basal level of nuclear NF-kappaB in native macrophages accelerates LPS-induced translocation of cytosolic NF-kappaB into the cell nucleus. Sci. Rep. 2019, 9, 4563. [Google Scholar] [CrossRef] [PubMed]
- Saklatvala, J. Inflammatory signaling in cartilage: MAPK and NF-kappaB pathways in chondrocytes and the use of inhibitors for research into pathogenesis and therapy of osteoarthritis. Curr. Drug Targets 2007, 8, 305–313. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.-Y.; Ahmad, F.; Ima-Nirwana, S. Regulation of inflammatory response and oxidative stress by tocotrienol in a rat model of non-alcoholic fatty liver disease. J. Funct. Foods 2020, 74, 104209. [Google Scholar] [CrossRef]
- Bao, J.; Chen, Z.; Xu, L.; Wu, L.; Xiong, Y. Rapamycin protects chondrocytes against IL-18-induced apoptosis and ameliorates rat osteoarthritis. Aging 2020, 12, 5152–5167. [Google Scholar] [CrossRef] [PubMed]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef]
- Mandraju, R.; Troutman, T.D.; Pasare, C. Toll-Like Receptor Function and Signaling. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Malemud, C.J. Chapter Seven—Matrix Metalloproteinases and Synovial Joint Pathology. In Progress in Molecular Biology and Translational Science; Matrix Metalloproteinases and Tissue Remodeling in Health and Disease: Target Tissues and Therapy; Khalil, R.A., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 148, pp. 305–325. [Google Scholar]
- Rim, Y.A.; Nam, Y.; Ju, J.H. The Role of Chondrocyte Hypertrophy and Senescence in Osteoarthritis Initiation and Progression. Int. J. Mol. Sci. 2020, 21, 2358. [Google Scholar] [CrossRef]
- Sun, K.; Wu, Y.; Zeng, Y.; Xu, J.; Wu, L.; Li, M.; Shen, B. The role of the sirtuin family in cartilage and osteoarthritis: Molecular mechanisms and therapeutic targets. Arthritis Res. Ther. 2022, 24, 286. [Google Scholar] [CrossRef]
- Cao, Z.; Huang, S.; Dou, C.; Xiang, Q.; Dong, S. Cyanidin suppresses autophagic activity regulating chondrocyte hypertrophic differentiation. J. Cell Physiol. 2018, 233, 2332–2342. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, J.; Li, J.J.; Zhou, H.; Zhu, X.W.; Zhang, P.H.; Huang, B.; Zhao, W.T.; Zhao, X.F.; Chen, E.S. Chondrocyte autophagy mechanism and therapeutic prospects in osteoarthritis. Front. Cell Dev. Biol. 2024, 12, 1472613. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Bahar, M.E.; Kim, C.-W.; Seo, M.-S.; Song, M.-G.; Song, S.-Y.; Kim, S.-Y.; Kim, D.-R.; Kim, D.-H. Autophagy in Osteoarthritis: A Double-Edged Sword in Cartilage Aging and Mechanical Stress Response: A Systematic Review. J. Clin. Med. 2024, 13, 3005. [Google Scholar] [CrossRef] [PubMed]
- Mironidou-Tzouveleki, M. Inflammation and pain. Ann. Gen. Psychiatry 2010, 9, S2. [Google Scholar] [CrossRef]
- Godyla-Jabłoński, M.; Raczkowska, E.; Jodkowska, A.; Kucharska, A.Z.; Sozański, T.; Bronkowska, M. Effects of Anthocyanins on Components of Metabolic Syndrome—A Review. Nutrients 2024, 16, 1103. [Google Scholar] [CrossRef] [PubMed]
- Nedunchezhiyan, U.; Varughese, I.; Sun, A.R.; Wu, X.; Crawford, R.; Prasadam, I. Obesity, Inflammation, and Immune System in Osteoarthritis. Front. Immunol. 2022, 13, 907750. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.C.; Chang, C.C.; Wu, W.T.; Lee, R.P.; Yao, T.K.; Peng, C.H.; Yeh, K.T. Effect of Osteoporosis Treatments on Osteoarthritis Progression in Postmenopausal Women: A Review of the Literature. Curr. Rheumatol. Rep. 2024, 26, 188–195. [Google Scholar] [CrossRef]
- Zhu, X.; Chan, Y.T.; Yung, P.S.H.; Tuan, R.S.; Jiang, Y. Subchondral Bone Remodeling: A Therapeutic Target for Osteoarthritis. Front. Cell Dev. Biol. 2020, 8, 607764. [Google Scholar] [CrossRef]
- Quek, Y.Y.; Cheng, L.J.; Ng, Y.X.; Hey, H.W.D.; Wu, X.V. Effectiveness of anthocyanin-rich foods on bone remodeling biomarkers of middle-aged and older adults at risk of osteoporosis: A systematic review, meta-analysis, and meta-regression. Nutr. Rev. 2023, 82, 1187–1207. [Google Scholar] [CrossRef]
- Mao, W.; Huang, G.; Chen, H.; Xu, L.; Qin, S.; Li, A. Research Progress of the Role of Anthocyanins on Bone Regeneration. Front. Pharmacol. 2021, 12, 773660. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Vitaglione, P.; Donnarumma, G.; Napolitano, A.; Galvano, F.; Gallo, A.; Scalfi, L.; Fogliano, V. Protocatechuic Acid Is the Major Human Metabolite of Cyanidin-Glucosides123. J. Nutr. 2007, 137, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.F.; Hsu, Y.H.; Lin, Y.C.; Nguyen, T.T.; Chen, H.W.; Nabilla, S.C.; Hou, S.Y.; Chang, F.C.; Chung, R.J. 3d printing of collagen/oligomeric proanthocyanidin/oxidized hyaluronic acid composite scaffolds for articular cartilage repair. Polymers 2021, 13, 3123. [Google Scholar] [CrossRef]
- Jeyaraman, M.; Ram, P.R.; Jeyaraman, N.; Yadav, S. The Gut-Joint Axis in Osteoarthritis. Cureus 2023, 15, e48951. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Hao, L.; Huang, G. Exploring the Interconnection between Metabolic Dysfunction and Gut Microbiome Dysbiosis in Osteoarthritis: A Narrative Review. Biomedicines 2024, 12, 2182. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Wang, F.; Li, Y.; Zheng, X. Black Current Anthocyanins Improve Lipid Metabolism and Modulate Gut Microbiota in High-Fat Diet-Induced Obese Mice. Mol. Nutr. Food Res. 2021, 65, e2001090. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Zhou, Y.; Zheng, X. Black rice anthocyanins alleviate hyperlipidemia, liver steatosis and insulin resistance by regulating lipid metabolism and gut microbiota in obese mice. Food Funct. 2021, 12, 10160–10170. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Deng, R.; Chu, Q.; Zheng, X. Pomegranate peel anthocyanins prevent diet-induced obesity and insulin resistance in association with modulation of the gut microbiota in mice. Eur. J. Nutr. 2022, 61, 1837–1847. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Yang, M.; Liu, M. Rutin prevents inflammation induced by lipopolysaccharide in RAW 264.7 cells via conquering the TLR4-MyD88-TRAF6-NF-κB signalling pathway. J. Pharm. Pharmacol. 2021, 73, 110–117. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Concept | Description |
|---|---|
| Population | Chondrocytes induced with OA changes Animals induced with OA Patients with OA |
| Concept | Anthocyanins or anthocyanidins |
| Context | In vitro, in vivo, or clinical trials |
| Authors (Year) | Study Type | OA Model/Subject Characteristics | Treatments | Parameters Increased vs. OA Control | Parameters Decreased vs. OA Control | Parameters Unchanged vs. OA Control | Limitations | Remarks |
|---|---|---|---|---|---|---|---|---|
| Haseeb et al. (2013) [24] | In vitro | Chondrocytes from OA patients’ femoral head undergoing hip replacement surgery, induced with IL-1β (1–5 ng/mL for inflammatory assays, 10 ng/mL for NFκB signalling). | Treatment: delphinidin (10 µg/mL for inflammatory assays, 50 µg/mL for NFκB signalling) first for 2 h, followed by IL-1β exposure (24 h for inflammation assays, 30 min for NFκB signalling). OA control: IL-1β exposure, no treatment. Positive control: BAY 11-7082 (2.5 mM for inflammatory assays, 10 mM for NFκB signalling). | NA | Inflammation markers: COX-2 mRNA and protein expressions, PGE2 production NFκB signalling: p-IRAK-1Ser376, p-IKKα/β, IKKβ mRNA and protein expression, IκBα degradation, NFκB activation and nuclear translocation, p-NF-κB-inducing kinase | p-TGF-β-activated kinase 1 | Anabolic and catabolic markers of chondrocytes not studied. | Delphinidin inhibits COX-2 expression and PGE2 by suppressing the activation of NFκB signalling by preventing the phosphorylation of IRAK-1Ser376 |
| Dai et al. (2017) [27] | In vitro, in vivo | In vivo: Wistar rats (8–12 weeks, sex not specified), injected with MIA (2 mg/kg) intra-articularly. In vitro: Apoptosis and inflammation assays: Chondrocytes from sham and OA group of the in vivo study. NFκB signalling: Chondrocytes from the femoral head of C57BL/6 mice exposed to IL-1β. Luciferase assay: 293T cells transfected with IjBa-specific siRNA. | In vivo: Treatment: malvidin (p.o., 10, 20 mg/kg/day) for 14 days. OA control: MIA injection without treatment (2 mg/kg). Positive control: NA. | In vivo: Day 14: Paw pressure and joint compression threshold (reduction in pain). | In vitro: Day 14: Apoptosis: Senescence-associated β-galactosidase-stained chondrocytes Days 1 and 7: Inflammation: Expression of IL-1β, IL-6, TNF-α, MMP3, MMP9, MMP13 NFκB: p65 nuclear translocation | In vitro: NFκB signalling: p-IKKα/β, IKKβ protein expression Luciferase assay: silencing of IκBα did not affect the effects of malvidin | Cartilage degradation not illustrated | Malvidin prevents pain and inflammation in rats with OA by suppressing the activation of NFκB signalling, but its mechanism is independent of IκBα |
| Jiang et al. (2019) [25] | In vitro, in vivo | In vitro: OA patients’ primary chondrocytes undergoing total knee replacement, exposed to IL-1β (10 ng/mL). In vivo: Animals: 10-week-old C57BL/6 male mice. OA induction: MMD. | In vitro: Co-incubation of chondrocytes with cyanidin (12.5, 25, and 50 µM). In vivo: Treatment: 5 mg/kg/day cyanidin for 8 weeks, oral. Negative control: normal saline. Positive control: NA. | In vitro: The protein expression of aggrecan, collagen II, SOX9 (12.5–50 µM, except collagen II at 25 µM). The protein expression and fluorescence intensity of sirtuin 6 (25–50 µM). * Silencing sirtuin 6 abolished all these changes. In vivo: Collagen II- and sirtuin 6-positive cells. | In vitro: Inflammation: mRNA and protein expression of iNOS, COX2, TNF-α, IL-6, the levels of NO and PGE2 (25 and 50 µM) Cartilage degradation markers: protein expression of MMP-13 and ADAMST4 (25 and 50 µM) NFκB signalling: phosphorylation of p65 and pIκB * Silencing sirtuin 6 abolished all these changes In vivo: OARSI scores, synovitis scores MMP-13 positive cells in the cartilage | NA | The anti-inflammatory effects were not replicated in the in vivo model | Cyanidin activates sirtuin 6 to suppress inflammation and achieve its chondroprotective effects |
| Lee et al. (2020) [28] | In vitro | C28/I2 human chondrocyte cells exposed to hydrogen peroxide (500 µM). | Delphinidin (40 µM). Negative control: not treated. Positive control: NAC (5 mM). | Cell viability Anti-apoptosis proteins: Bcl-Xl levels. Antioxidant response proteins: Nfr2, p-NFκB. Autophagy markers: LC3 expression, autophagic vacuoles (staining) [increased by H2O2, further increased in the presence of delphinidin]. * These changes were inversed in the presence of chloroquine, an autophagy inhibitor. | Pro-apoptosis markers: c-caspase-3 and c-PARP Apoptotic cells (TUNEL assay) | NA | Not human primary cell line Upstream mechanism of autophagy not investigated | Delphinidin protects chondrocytes subjected to oxidative stress by activating Nrf2, NFκB, and autophagy of the cells. |
| Chuntakaruk et al. (2021) [15] | In vitro studies | Porcine cartilage exposed to AGE (25 μg/mL). Non-OA patients’ joint material, exposed to AGE (10 μg/mL). | Purple corn anthocyanins, containing cyanidin-3-O-glucoside chloride (C3G, 53.39 ± 0.54 μg/g of crude extract), pelargonidin-3-O-glucoside (P3G, 34.21 ± 0.13 μg/g of crude extract), peonidin-3-O-glucoside chloride (P3GC, 33.18 ± 0.12 μg/g of crude extract), and malvidin-3-O-glucoside (M3OG, 16.50 ± 0.05 μg/g of crude extract): 6.25–25 μg/mL for explant assays for 35 days. C3G: 1.25–5 μM; P3G: 2.5–10 μM; P3GC: 2.5–10 μM; Protocatechuic acid (PCA): 2.5–10 μM. | Retention of uronic acid in the explant: PCA. Glycosaminoglycan retention in the explant (Safranin O staining): PCA. | Glycosaminoglycan and hyaluronic acid release from cartilage explant: PCA mRNA expression of MMP1, 3, 13: PCA and all constituents p-IKK/IKK, p-IκB/IκB, p-p65/p65: PCA and all constituents p-ERK/ERK, p-p-38/p38, p-JNK/JNK: PCA and all constituents | Chondrocyte morphology (H&E staining) P3G not effective on p-p38 | Unclear details on human cartilage donor AGE-RAGE signalling study not performed | Purple corn anthocyanin and its constituents protect against cartilage loss due to AGE through NFκB and MAPK signalling pathways |
| Wongwichai et al. (2019) [16] | In vitro studies | Explant study: Porcine articular cartilage exposed to IL-1β (25 ng/mL for 3 days, 10 ng/mL for 35 days in the presence of oncostatin M 10 ng/mL). Chondrocyte study: Non-OA human articular chondrocytes, exposed to IL-1β (2 ng/mL, duration varies on experiments). | PCA: Explant study: 6.25–50 µg/mL; chondrocyte study: 2.5–10 µM. Anthocyanidins: - Cyanidin-3-O-glucoside chloride (C3G): 2.5–10 µM. Peonidin-3-O-glucoside chloride (P3G): 2.5–10 µM. Anthocyanins: Cyanidin chloride (CC): 2.5–10 µM; Peonidin chloride (PC): 2.5–10 µM; PCA: 2.5–10 µM. | NA | Explant study: PCA: release of glycosaminoglycan (7–14 days), hyaluronic acid (7–28 days), hydroxyproline (21–35 days) Chondrocyte study: mRNA expression of MMP-1, MMP-3, MMP-13: C3G, P3G, PC, PCA p-IKK/IKK, p-IκB/IκB: C3G, P3G, CC, PC, PCA p-p65/p65: C3G, P3G, PCA p-ERK/ERK: C3G, P3G, PC, PA p-JNK/JNK: PC | Explant study: PCA: Uronic acid retention, cartilage and chondrocyte morphology (H&E), glycosaminoglycan retention (Safranin-O staining), collagen retention (staining) p-p65/p65: CC, PC p-p38/p38: C3G, P3G, CC, PC, PCA p-JNK/JNK: C3G, P3G, CC, PCA | Unclear details on human cartilage donor | Anthocyanidins inhibit NFκB activation more effectively than anthocyanins in preventing cartilage degradation |
| Zeng et al. (2023) [26] | In vitro, in vivo | In vitro: Primary chondrocytes from neonatal mice’s knee cartilage, exposed to IL-1β (10 mg/mL). In vivo: Medial meniscus destabilisation in C57BL/6 mice. | In vitro: pelargonidin (10–40 μM) for 24–72 h. In vivo: pelargonidin (10 mg/kg/d, 20 mg/kg/d) for 8 weeks. Negative control: normal saline. Positive control: NA. | In vitro: Toluidine blue staining, collagen II protein expression. In vivo: Aggrecan expression (IHC). | In vitro: mRNA expression of IL-6, TNF-α, COX-2 and iNOS; protein expression of COX-2, iNOS, ADAMTS5 and MMP13 p-p65/p65 expression and nuclear translocation of p65 In vivo: OARSI score (Safranin O) MMP13 expression (IHC) | NA | Single cell line, not from human Inflammatory effects not demonstrated in vivo | Pelargonidin prevents cartilage degradation by suppressing NFκB activation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Chin, K.-Y.; Ekeuku, S.O. Anthocyanins and Anthocyanidins in the Management of Osteoarthritis: A Scoping Review of Current Evidence. Pharmaceuticals 2025, 18, 301. https://doi.org/10.3390/ph18030301
Ma X, Chin K-Y, Ekeuku SO. Anthocyanins and Anthocyanidins in the Management of Osteoarthritis: A Scoping Review of Current Evidence. Pharmaceuticals. 2025; 18(3):301. https://doi.org/10.3390/ph18030301
Chicago/Turabian StyleMa, Xiaodong, Kok-Yong Chin, and Sophia Ogechi Ekeuku. 2025. "Anthocyanins and Anthocyanidins in the Management of Osteoarthritis: A Scoping Review of Current Evidence" Pharmaceuticals 18, no. 3: 301. https://doi.org/10.3390/ph18030301
APA StyleMa, X., Chin, K.-Y., & Ekeuku, S. O. (2025). Anthocyanins and Anthocyanidins in the Management of Osteoarthritis: A Scoping Review of Current Evidence. Pharmaceuticals, 18(3), 301. https://doi.org/10.3390/ph18030301