
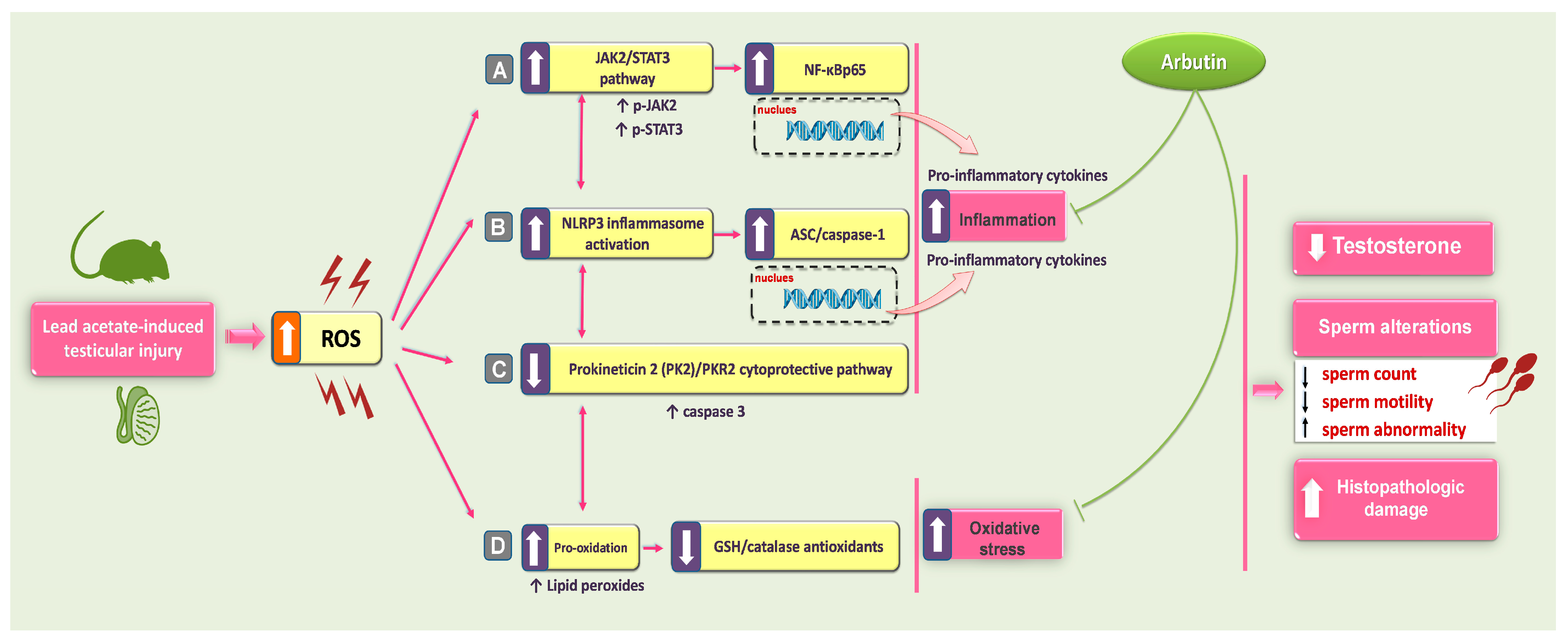
Targeting JAK2/STAT3, NLRP3/Caspase-1, and PK2/PKR2 Pathways with Arbutin Ameliorates Lead Acetate-Induced Testicular Injury in Rats
 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
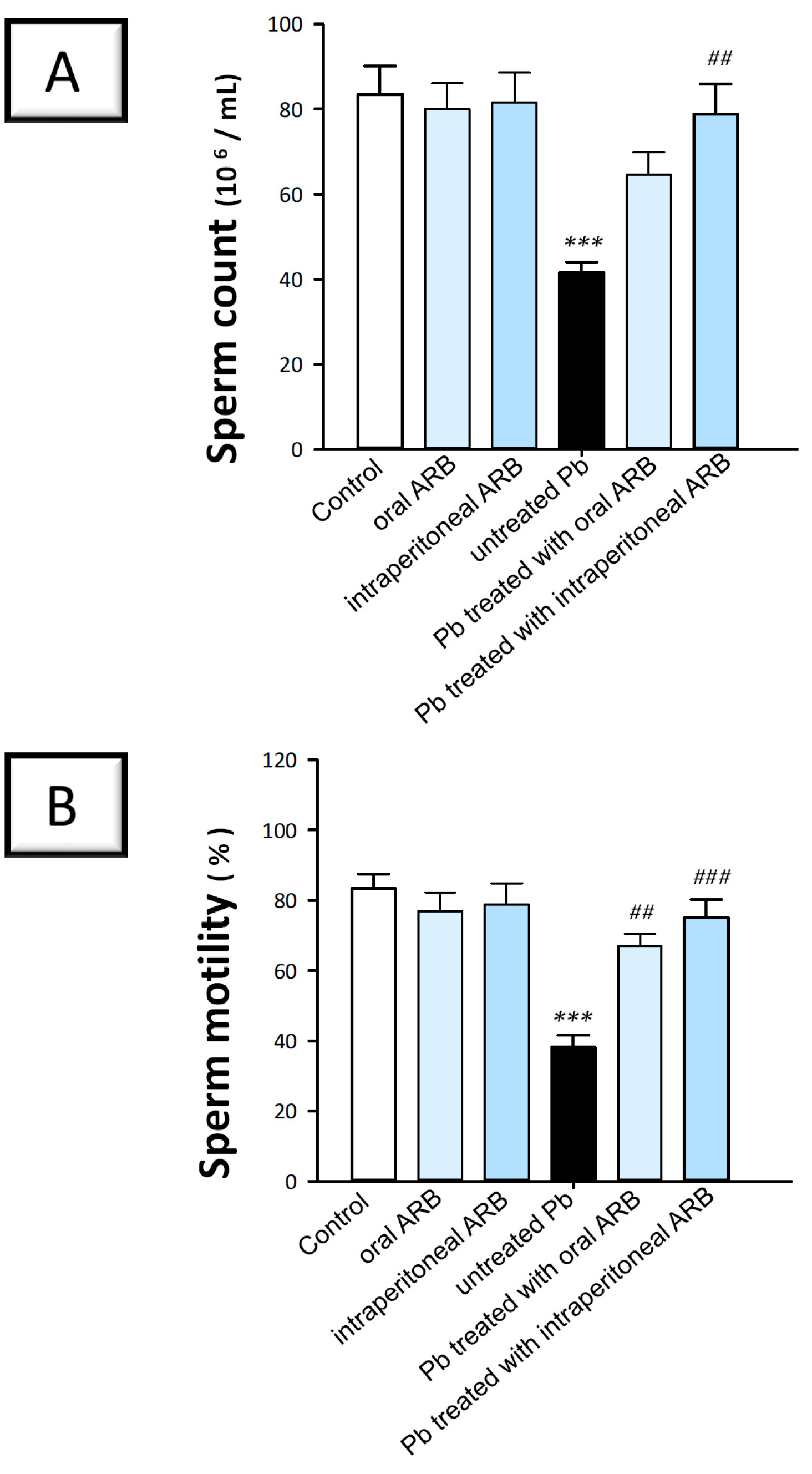
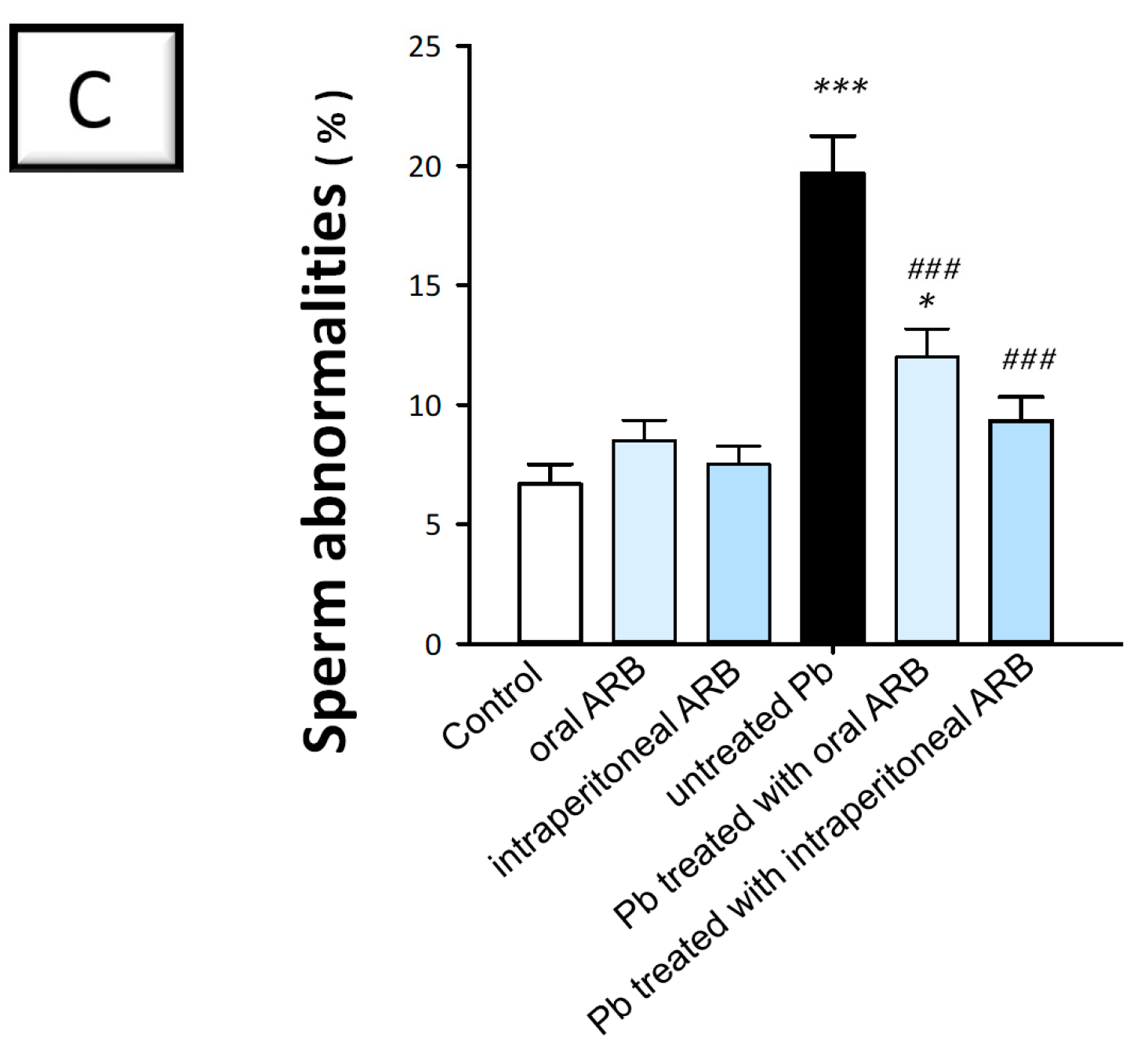
2.1. Arbutin Improved Lead Acetate-Evoked Alterations in Semen Analyses
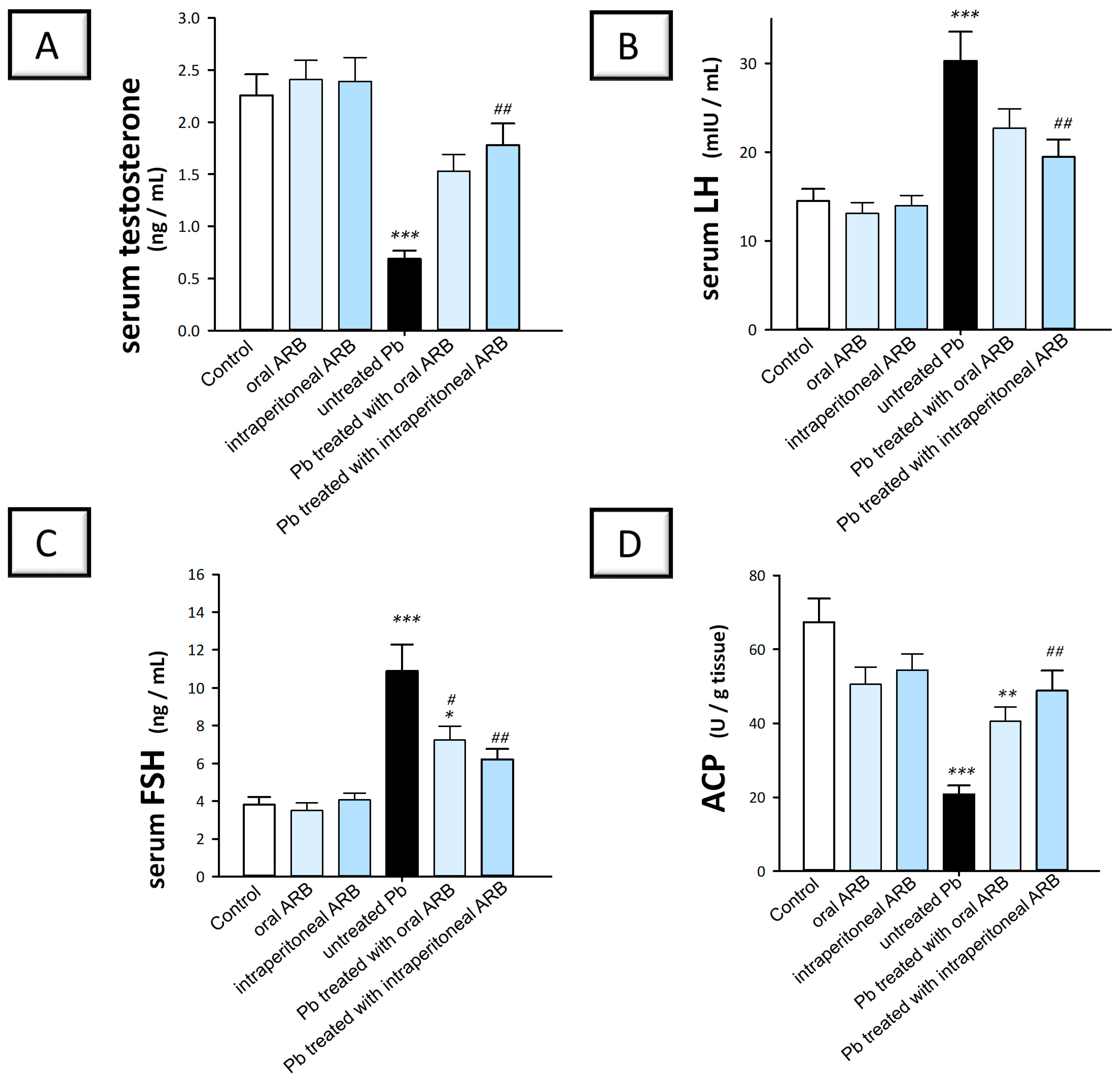
2.2. Arbutin Administration Rescued Serum Sex Hormones in Lead Acetate-Treated Rats
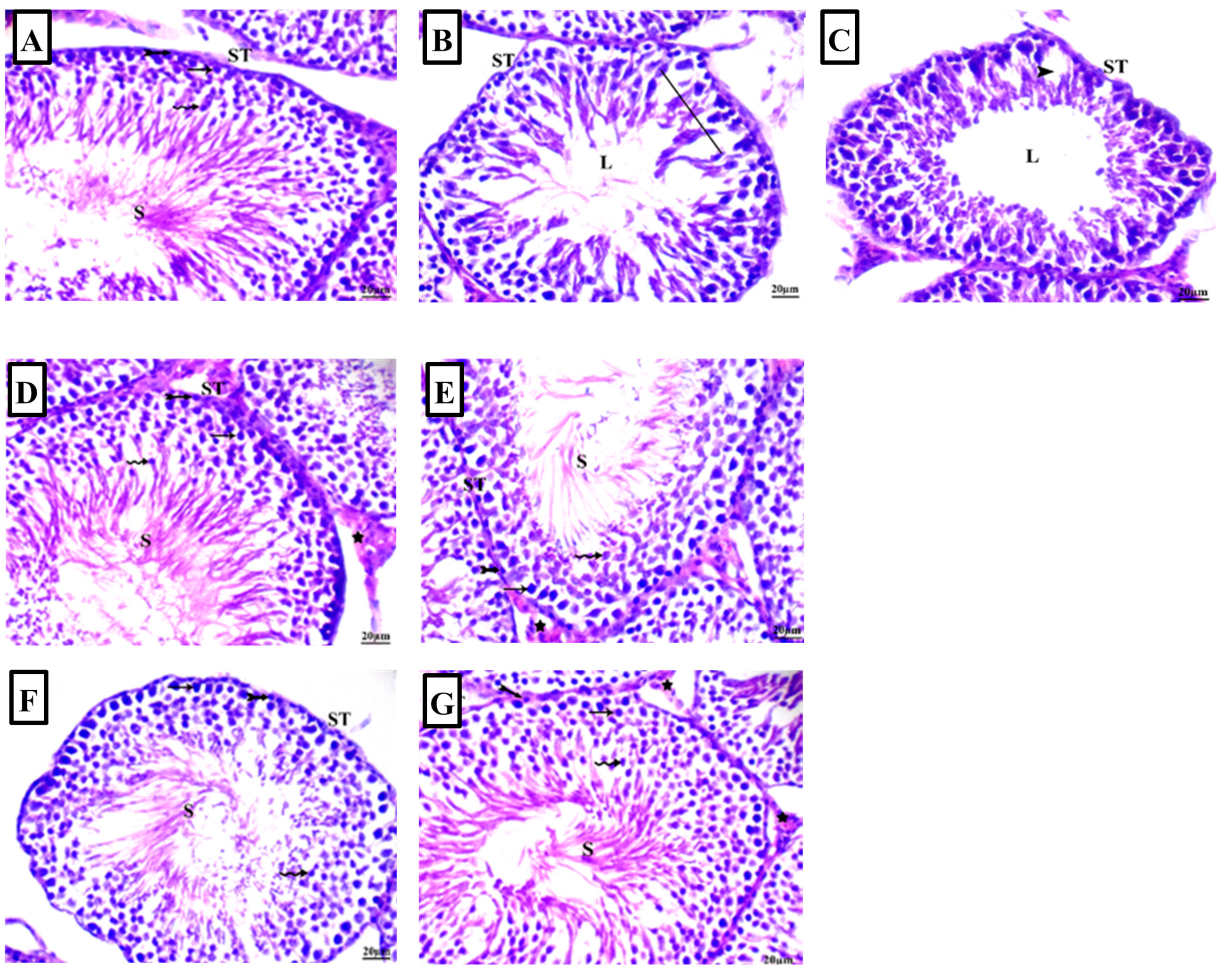
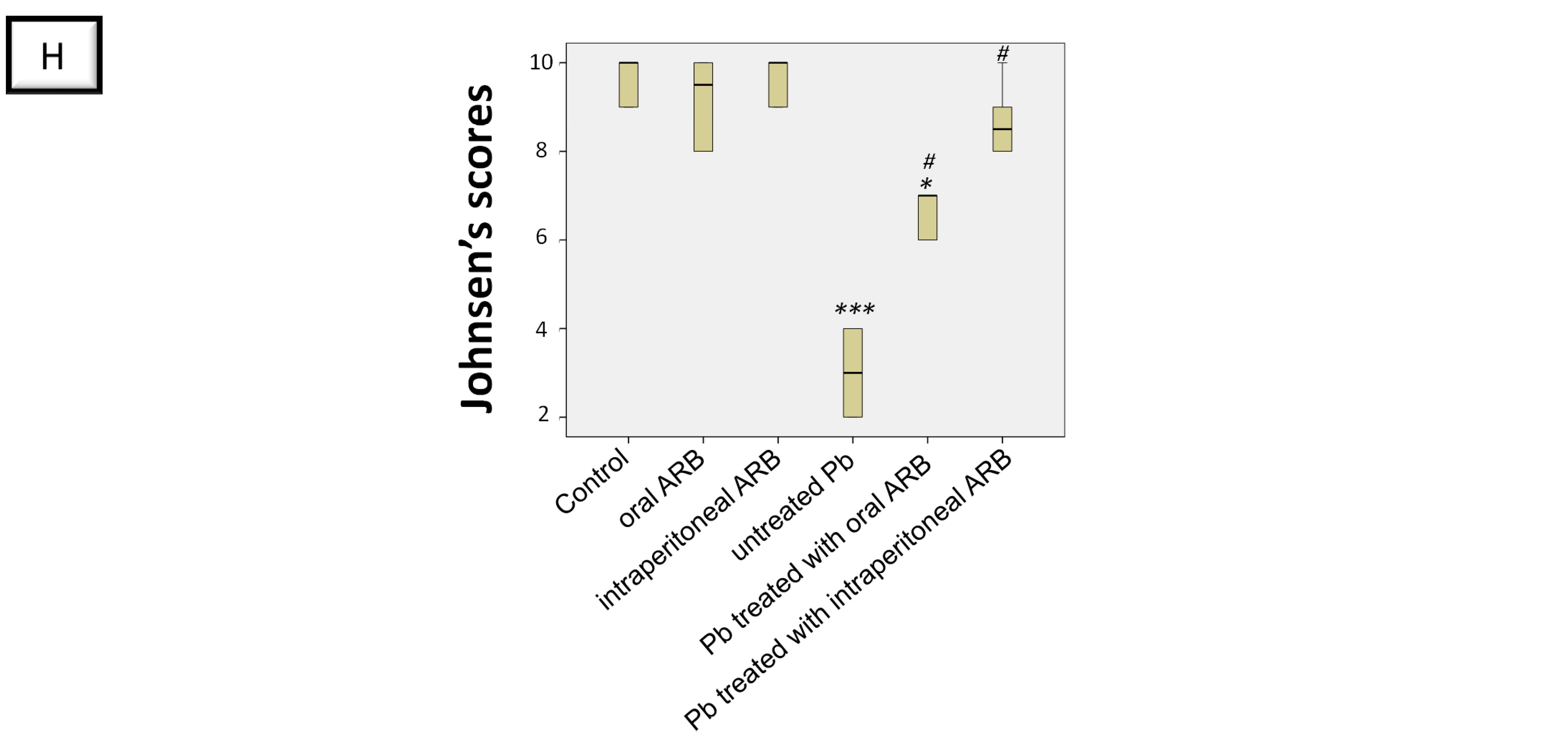
2.3. Arbutin Ameliorated Lead Acetate-Induced Histological Lesions in Testicular Tissue
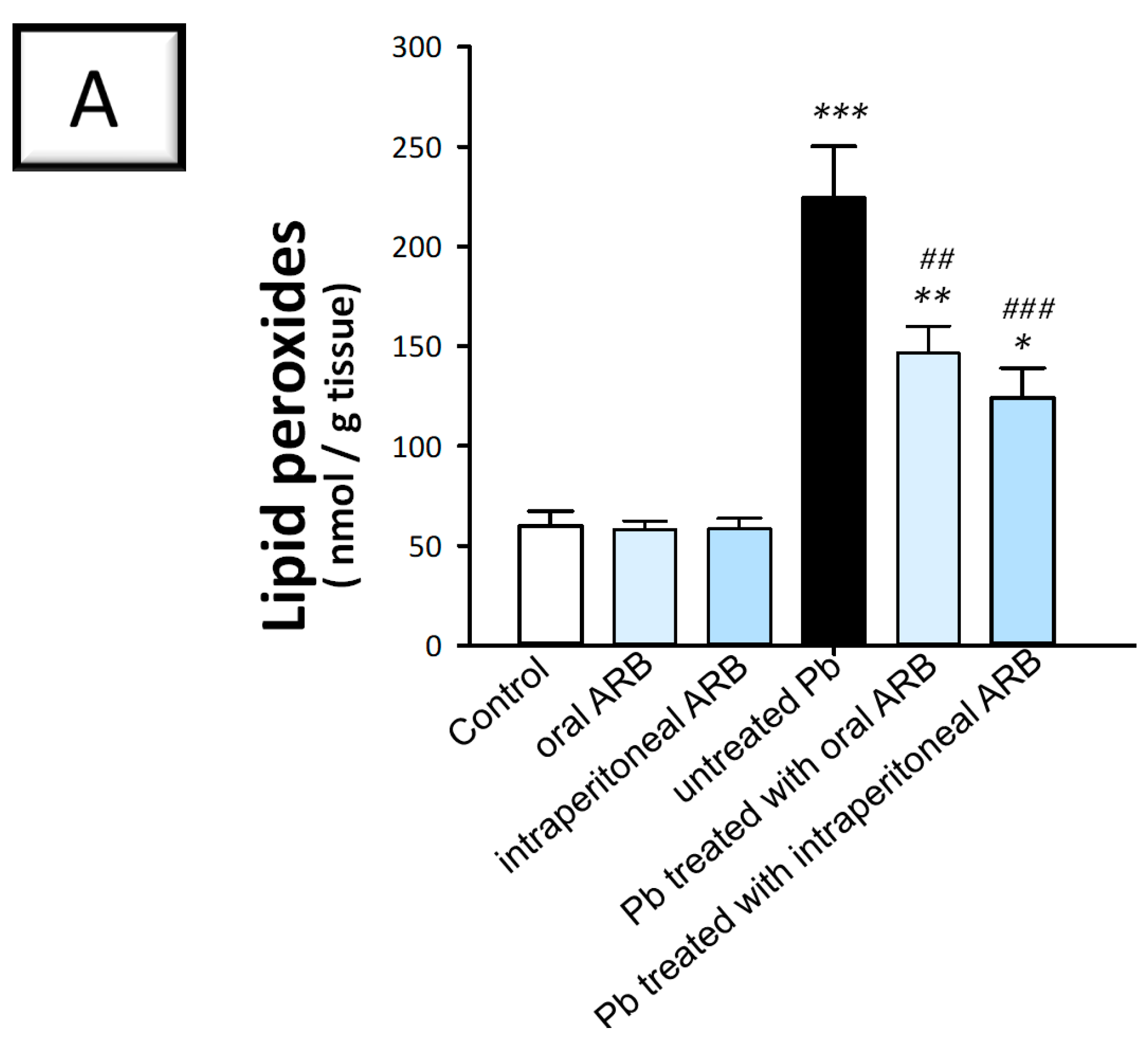
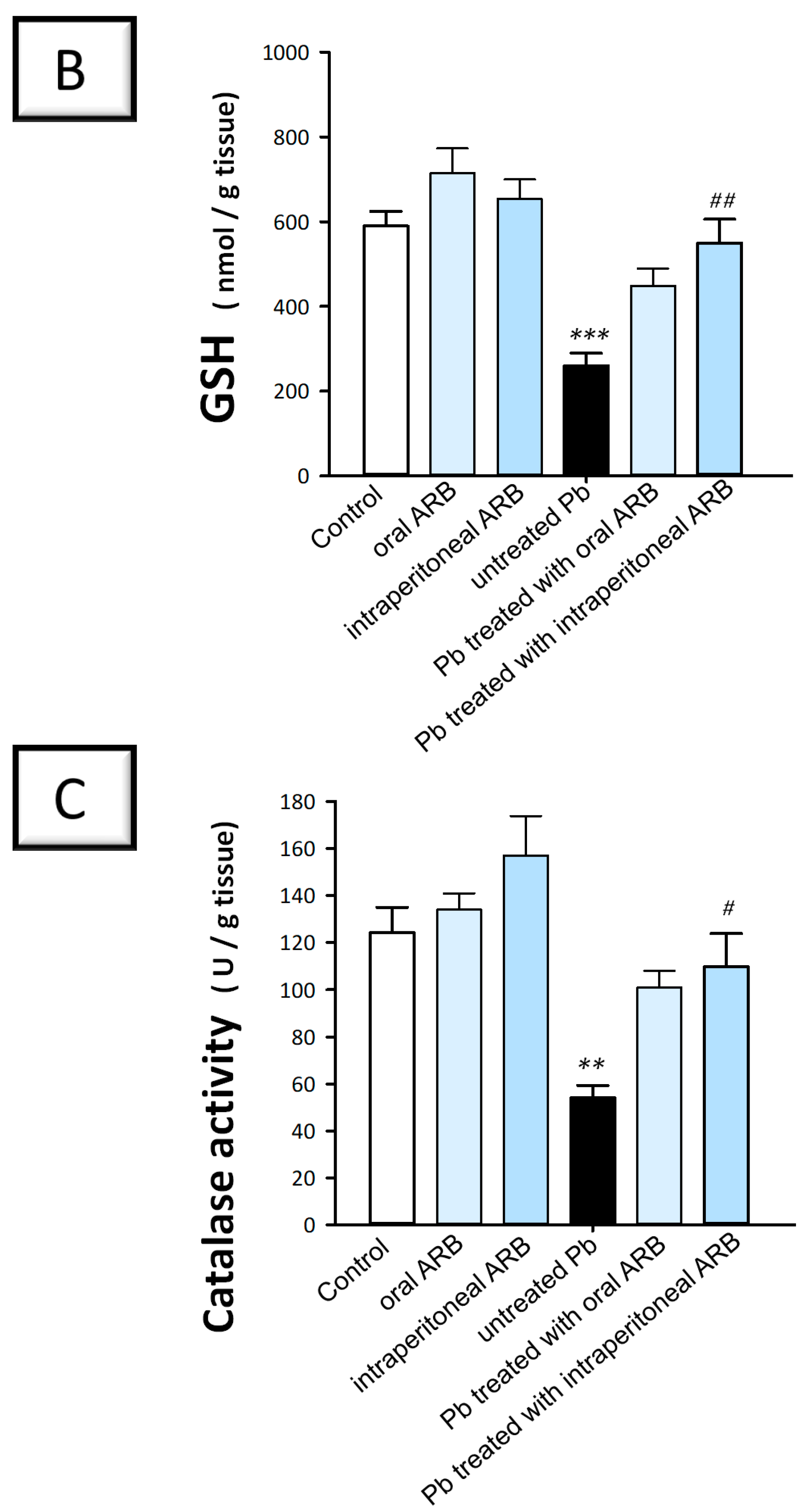
2.4. Arbutin Administration Mitigated Lead Acetate-Induced Oxidative Stress in the Testes of Rats
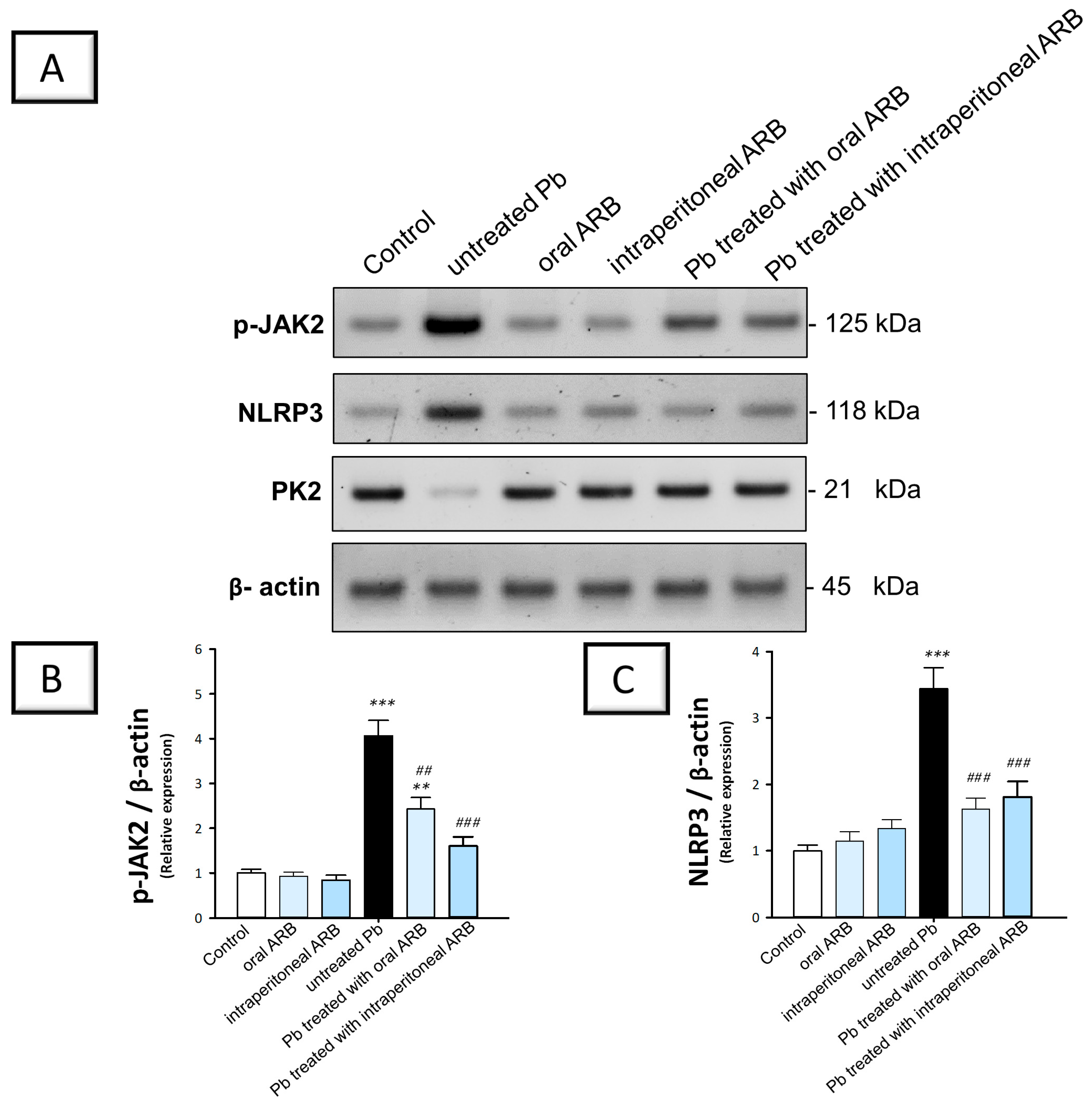
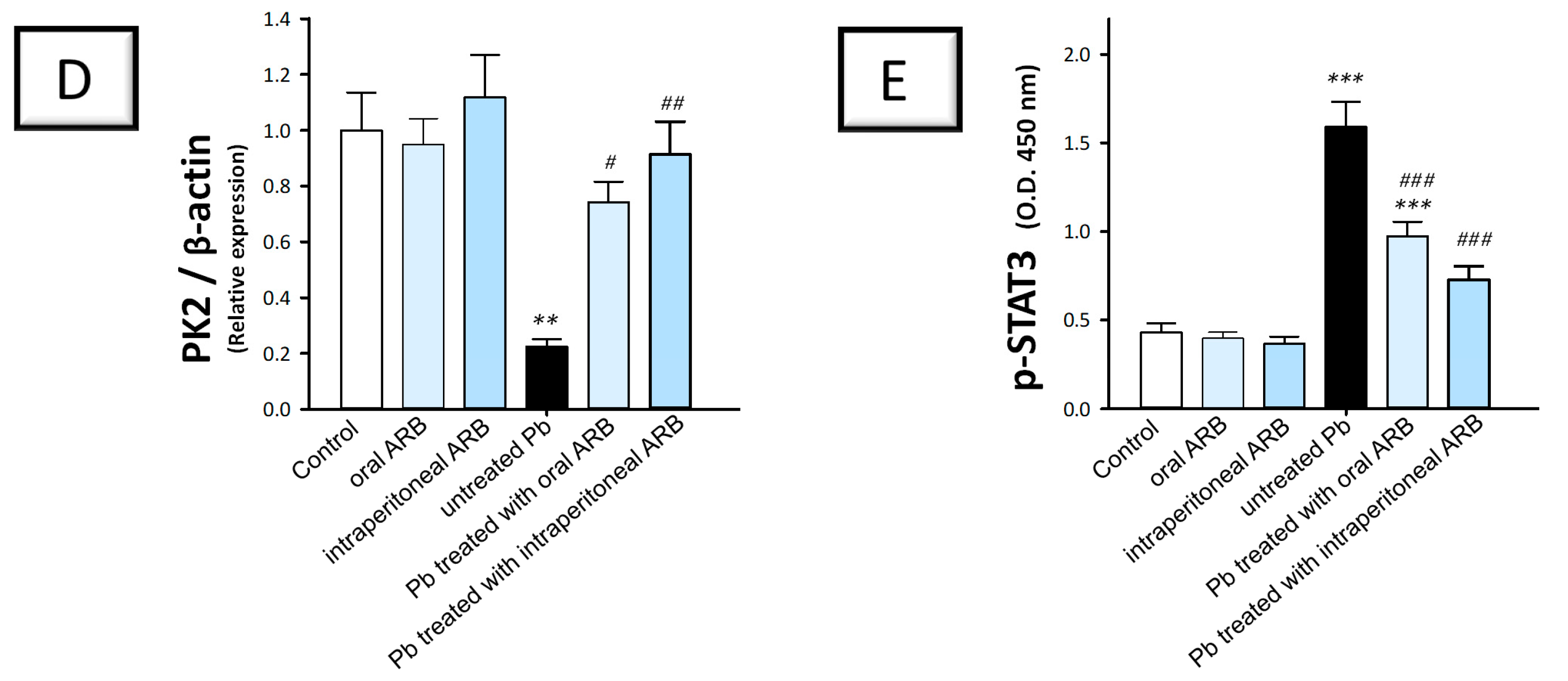
2.5. Arbutin Suppressed the JAK2/STAT3 Pathway in the Testicular Tissue of Lead Acetate-Intoxicated Rats
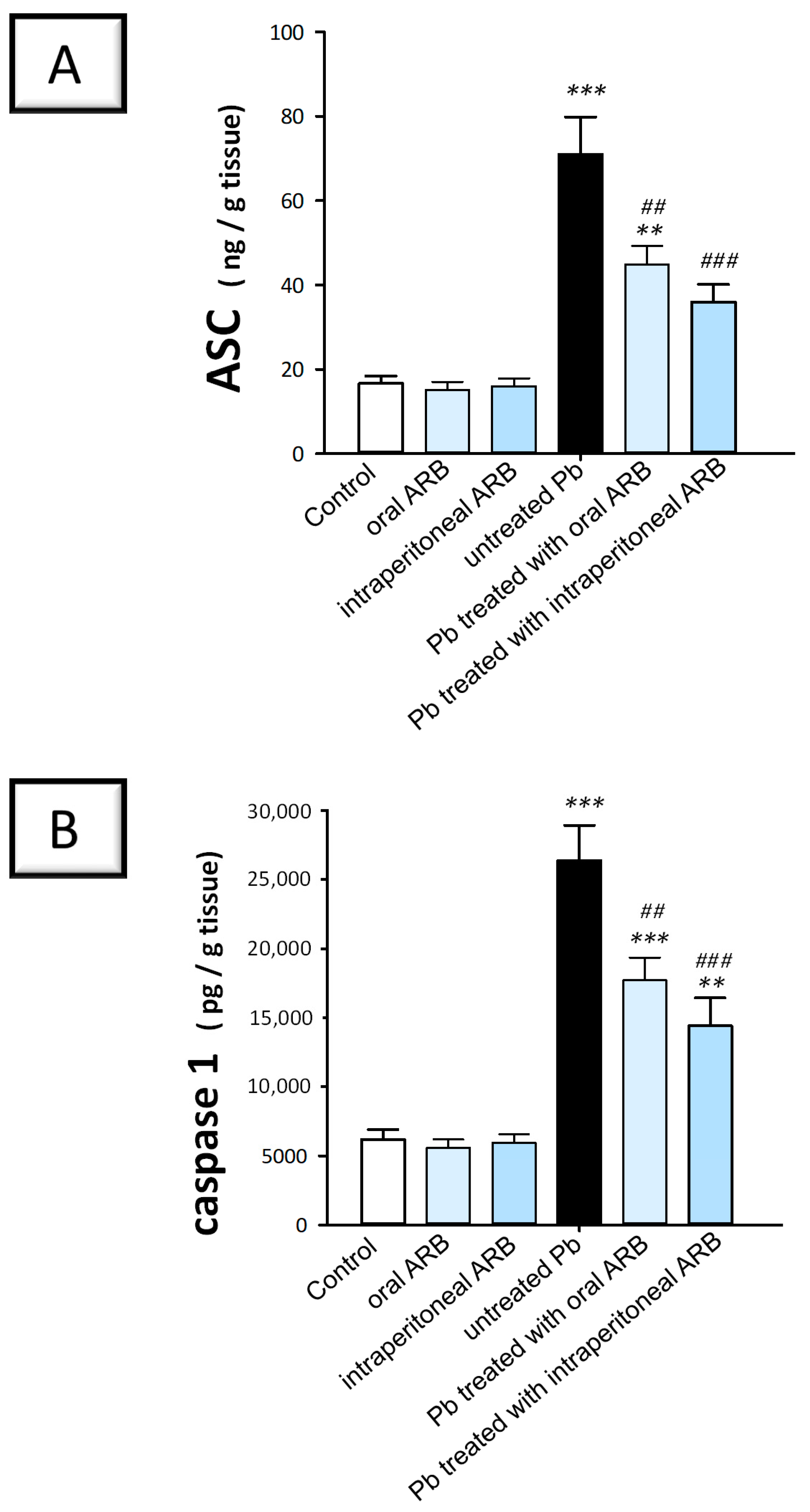
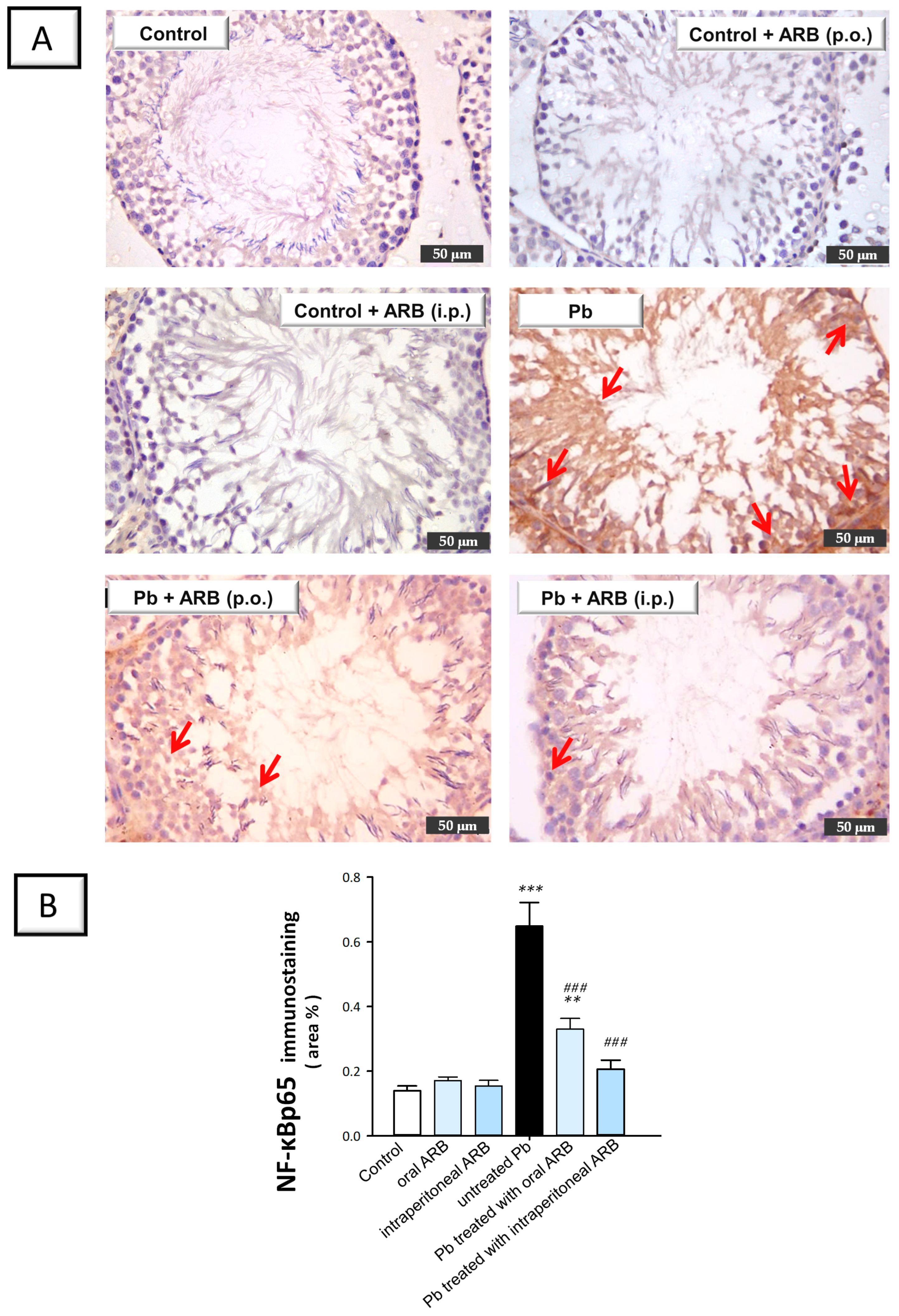
2.6. Arbutin Curtailed the Testicular NLRP3/Caspase-1/NF-κB Pathway in Lead Acetate-Intoxicated Rats
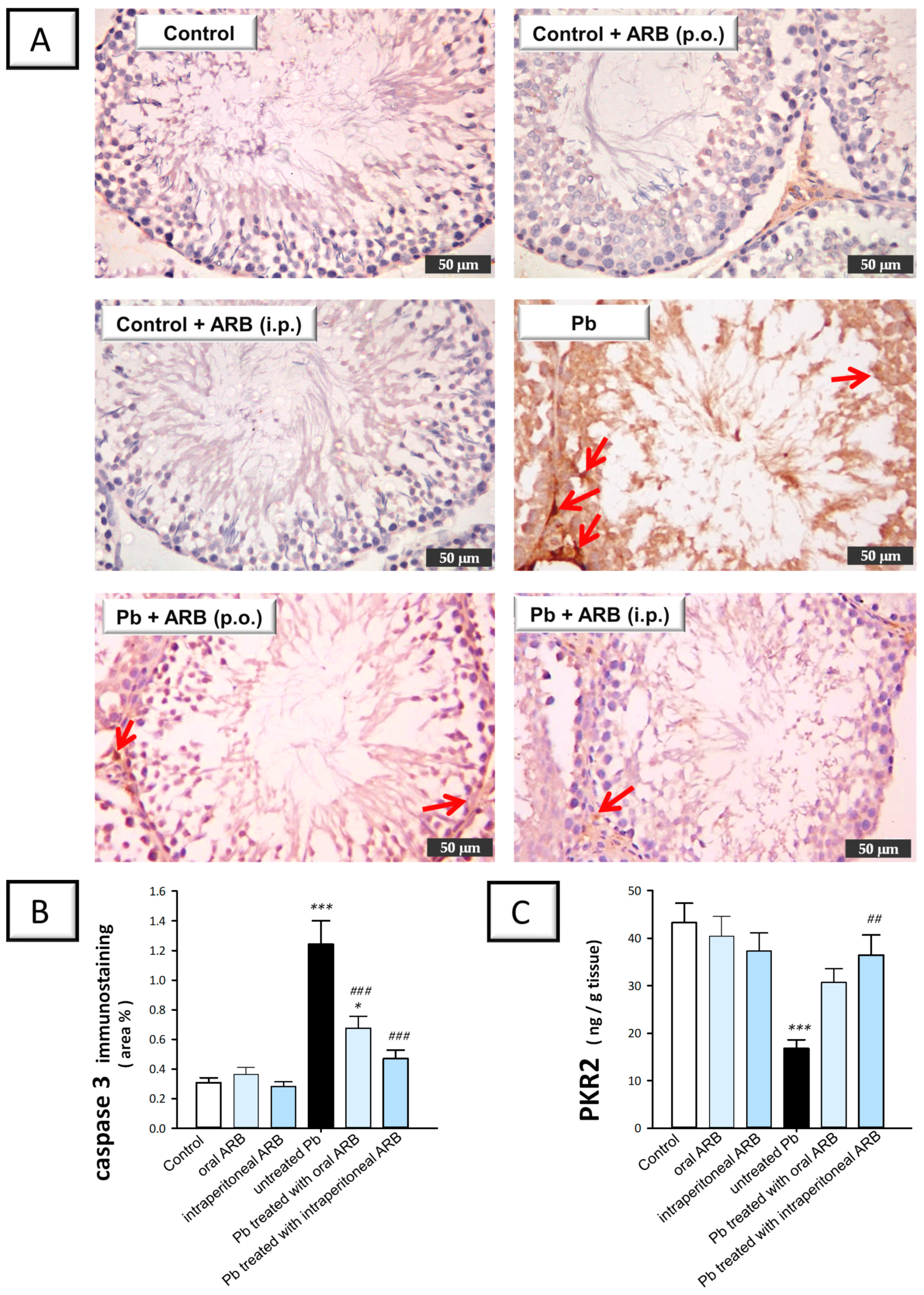
2.7. Arbutin Stimulated the PK2/PKR2 Pathway and Curtailed Caspase 3 Expression in the Testes of Lead Acetate-Intoxicated Rats
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Experimental Animals
4.3. Study Design
4.4. Determination of Sperm Parameters
4.5. Determination of Sex Hormones
4.6. Determination of Testicular Redox Milieu
4.7. Assessment of p-STAT3, ASC, Caspase 1, and PKR2
4.8. Western Blot Analysis
4.9. Histology
4.10. Immunohistochemistry (IHC)
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Massanyi, P.; Massanyi, M.; Madeddu, R.; Stawarz, R.; Lukac, N. Effects of Cadmium, Lead, and Mercury on the Structure and Function of Reproductive Organs. Toxics 2020, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prasad, M.N.V. Plant-lead interactions: Transport, toxicity, tolerance, and detoxification mechanisms. Ecotoxicol. Environ. Saf. 2018, 166, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.; Kahilo, K.; Kamal, T.; Hassan, M.; Saleh Elgawish, M. The protective effect of epigallocatechin-3-gallate on testicular oxidative stress in lead-induced toxicity mediated by Cyp19 gene/estradiol level. Toxicology 2019, 422, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Elhemiely, A.A.; Yahia, R.; Gad, A.M. Naringenin alleviate reproductive toxicity evoked by lead acetate via attenuation of sperm profile and biochemical alterations in male Wistar rat: Involvement of TGFbeta/AKT/mTOR pathway. J. Biochem. Mol. Toxicol. 2023, 37, e23335. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Li, X.; Wang, Z.; Lin, X.; Tian, Y.; Zhao, Q.; Zheng, P. Anti-inflammatory effect of selenium on lead-induced testicular inflammation by inhibiting NLRP3 inflammasome activation in chickens. Theriogenology 2020, 155, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Al-Megrin, W.A.; Alomar, S.; Alkhuriji, A.F.; Metwally, D.M.; Mohamed, S.K.; Kassab, R.B.; Abdel Moneim, A.E.; El-Khadragy, M.F. Luteolin protects against testicular injury induced by lead acetate by activating the Nrf2/HO-1 pathway. IUBMB Life 2020, 72, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- Akarsu, S.A.; Gur, C.; Ileriturk, M.; Akaras, N.; Kucukler, S.; Kandemir, F.M. Effect of syringic acid on oxidative stress, autophagy, apoptosis, inflammation pathways against testicular damage induced by lead acetate. J. Trace Elem. Med. Biol. 2023, 80, 127315. [Google Scholar] [CrossRef]
- BaSalamah, M.A.; Abdelghany, A.H.; El-Boshy, M.; Ahmad, J.; Idris, S.; Refaat, B. Vitamin D alleviates lead induced renal and testicular injuries by immunomodulatory and antioxidant mechanisms in rats. Sci. Rep. 2018, 8, 4853. [Google Scholar] [CrossRef] [PubMed]
- Alnajem, A.; Al-Maghrebi, M. The Regulatory Effects of JAK2/STAT3 on Spermatogenesis and the Redox Keap1/Nrf2 Axis in an Animal Model of Testicular Ischemia Reperfusion Injury. Cells 2023, 12, 2292. [Google Scholar] [CrossRef]
- Khashab, F.; Al-Saleh, F.; Al-Kandari, N.; Fadel, F.; Al-Maghrebi, M. JAK Inhibition Prevents DNA Damage and Apoptosis in Testicular Ischemia-Reperfusion Injury via Modulation of the ATM/ATR/Chk Pathway. Int. J. Mol. Sci. 2021, 22, 13390. [Google Scholar] [CrossRef]
- Arab, H.H.; Abd El-Aal, S.A.; Ashour, A.M.; El-Sheikh, A.A.K.; Al Khabbaz, H.J.; Arafa, E.A.; Mahmoud, A.M.; Kabel, A.M. Targeting inflammation and redox perturbations by lisinopril mitigates Freund’s adjuvant-induced arthritis in rats: Role of JAK-2/STAT-3/RANKL axis, MMPs, and VEGF. Inflammopharmacology 2022, 30, 1909–1926. [Google Scholar] [CrossRef]
- Shafiey, S.I.; Ahmed, K.A.; Abo-Saif, A.A.; Abo-Youssef, A.M.; Mohamed, W.R. Galantamine mitigates testicular injury and disturbed spermatogenesis in adjuvant arthritic rats via modulating apoptosis, inflammatory signals, and IL-6/JAK/STAT3/SOCS3 signaling. Inflammopharmacology 2023, 32, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Z.; Kong, D.; Zhang, Y.; Yu, W.; Zha, W. Metformin Ameliorates Testicular Damage in Male Mice with Streptozotocin-Induced Type 1 Diabetes through the PK2/PKR Pathway. Oxid. Med. Cell Longev. 2019, 2019, 5681701. [Google Scholar] [CrossRef] [PubMed]
- Pitteloud, N.; Zhang, C.; Pignatelli, D.; Li, J.D.; Raivio, T.; Cole, L.W.; Plummer, L.; Jacobson-Dickman, E.E.; Mellon, P.L.; Zhou, Q.Y.; et al. Loss-of-function mutation in the prokineticin 2 gene causes Kallmann syndrome and normosmic idiopathic hypogonadotropic hypogonadism. Proc. Natl. Acad. Sci. USA 2007, 104, 17447–17452. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhao, X.; Yu, J.; Xie, J.; Li, C.; Liu, D.; Tang, C.; Wang, C. Lead-induced oxidative damage in rats/mice: A meta-analysis. J. Trace Elem. Med. Biol. 2020, 58, 126443. [Google Scholar] [CrossRef]
- Abu-Khudir, R.; Almutairi, H.H.; Abd El-Rahman, S.S.; El-Said, K.S. The Palliative and Antioxidant Effects of Hesperidin against Lead-Acetate-Induced Testicular Injury in Male Wistar Rats. Biomedicines 2023, 11, 2390. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, M.; Khezri, K.; Seyed Zakaryaei, A.; Mohammadamini, H. A comprehensive review of the therapeutic potential of alpha-arbutin. Phytother. Res. 2021, 35, 4136–4154. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, H.; Jie, H.; Ding, H.; Sun, H. Arbutin ameliorated ulcerative colitis of mice induced by dextran sodium sulfate (DSS). Bioengineered 2021, 12, 11707–11715. [Google Scholar] [CrossRef]
- Alruhaimi, R.S. Protective effect of arbutin against cyclophosphamide-induced oxidative stress, inflammation, and hepatotoxicity via Nrf2/HO-1 pathway in rats. Environ. Sci. Pollut. Res. Int. 2023, 30, 68101–68110. [Google Scholar] [CrossRef]
- Khadir, F.; Pouramir, M.; Joorsaraee, S.G.; Feizi, F.; Sorkhi, H.; Yousefi, F. The effect of arbutin on lipid peroxidation and antioxidant capacity in the serum of cyclosporine-treated rats. Caspian J. Intern. Med. 2015, 6, 196–200. [Google Scholar]
- Demir, E.A.; Mentese, A.; Yilmaz, Z.S.; Alemdar, N.T.; Demir, S.; Aliyazicioglu, Y. Evaluation of the therapeutic effects of arbutin on cisplatin-induced ovarian toxicity in rats through endoplasmic reticulum stress and Nrf2 pathway. Reprod. Biol. 2023, 23, 100824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zeng, M.; Li, B.; Kan, Y.; Wang, S.; Cao, B.; Huang, Y.; Zheng, X.; Feng, W. Arbutin attenuates LPS-induced acute kidney injury by inhibiting inflammation and apoptosis via the PI3K/Akt/Nrf2 pathway. Phytomedicine 2021, 82, 153466. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, M.; Mittal, R.; Kumar, M.; Kushwah, A.S. Pharmacological Aspects of a Bioactive Compound Arbutin: A Comprehensive Review. J. Biointerface Res. Appl. Chem. 2022, 13, 119. [Google Scholar]
- Hassan, E.; El-Neweshy, M.; Hassan, M.; Noreldin, A. Thymoquinone attenuates testicular and spermotoxicity following subchronic lead exposure in male rats: Possible mechanisms are involved. Life Sci. 2019, 230, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.; Kahilo, K.; Kamal, T.; El-Neweshy, M.; Hassan, M. Protective effect of diallyl sulfide against lead-mediated oxidative damage, apoptosis and down-regulation of CYP19 gene expression in rat testes. Life Sci. 2019, 226, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Schindler, G.; Patzak, U.; Brinkhaus, B.; von Niecieck, A.; Wittig, J.; Krahmer, N.; Glockl, I.; Veit, M. Urinary excretion and metabolism of arbutin after oral administration of Arctostaphylos uvae ursi extract as film-coated tablets and aqueous solution in healthy humans. J. Clin. Pharmacol. 2002, 42, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Kumar, A.; Sindhu, R.K.; Kushwah, A.S. Arbutin attenuates monosodium L-glutamate induced neurotoxicity and cognitive dysfunction in rats. Neurochem. Int. 2021, 151, 105217. [Google Scholar] [CrossRef]
- Bearden, H.J.; Fuquay, J.W. Applied Animal Reproduction; Reston Publishing Company, Inc.: Reston, VA, USA, 1980. [Google Scholar]
- Arab, H.H.; Fikry, E.M.; Alsufyani, S.E.; Ashour, A.M.; El-Sheikh, A.A.K.; Darwish, H.W.; Al-Hossaini, A.M.; Saad, M.A.; Al-Shorbagy, M.Y.; Eid, A.H. Stimulation of Autophagy by Dapagliflozin Mitigates Cadmium-Induced Testicular Dysfunction in Rats: The Role of AMPK/mTOR and SIRT1/Nrf2/HO-1 Pathways. Pharmaceuticals 2023, 16, 1006. [Google Scholar] [CrossRef]
- Matsuyama, S.; DeFalco, T. Steroid hormone signaling: Multifaceted support of testicular function. Front. Cell Dev. Biol. 2023, 11, 1339385. [Google Scholar] [CrossRef]
- Noriega, N.C.; Howdeshell, K.L.; Furr, J.; Lambright, C.R.; Wilson, V.S.; Gray, L.E., Jr. Pubertal administration of DEHP delays puberty, suppresses testosterone production, and inhibits reproductive tract development in male Sprague-Dawley and Long-Evans rats. Toxicol. Sci. 2009, 111, 163–178. [Google Scholar] [CrossRef]
- Johnsen, S.G. Testicular biopsy score count--A method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Poli, G.; Fabi, C.; Sugoni, C.; Bellet, M.M.; Costantini, C.; Luca, G.; Brancorsini, S. The Role of NLRP3 Inflammasome Activation and Oxidative Stress in Varicocele-Mediated Male Hypofertility. Int. J. Mol. Sci. 2022, 23, 5233. [Google Scholar] [CrossRef] [PubMed]
- Moretti, E.; Collodel, G.; Fiaschi, A.I.; Micheli, L.; Iacoponi, F.; Cerretani, D. Nitric oxide, malondialdheyde and non-enzymatic antioxidants assessed in viable spermatozoa from selected infertile men. Reprod. Biol. 2017, 17, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Kim, K.-W.J.I.R. Anti-inflammatory effects of arbutin in lipopolysaccharide-stimulated BV2 microglial cells. Inflamm. Res. 2012, 61, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wang, J.; Zhao, T.; Wu, Y.; Wei, Y.; Chen, J.; Kang, L.; Shen, L.; Long, C.; Yang, Z.; et al. Stereological analysis and transcriptome profiling of testicular injury induced by di-(2-ethylhexyl) phthalate in prepubertal rats. Ecotoxicol. Environ. Saf. 2021, 220, 112326. [Google Scholar] [CrossRef] [PubMed]
- Darwish, H.A.; Arab, H.H.; Abdelsalam, R.M. Chrysin alleviates testicular dysfunction in adjuvant arthritic rats via suppression of inflammation and apoptosis: Comparison with celecoxib. Toxicol. Appl. Pharmacol. 2014, 279, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Moran-Martinez, J.; Carranza-Rosales, P.; Morales-Vallarta, M.; Heredia-Rojas, J.A.; Bassol-Mayagoitia, S.; Denys Betancourt-Martinez, N.; Cerda-Flores, R.M. Chronic environmental exposure to lead affects semen quality in a Mexican men population. Iran. J. Reprod. Med. 2013, 11, 267–274. [Google Scholar] [PubMed]
- Arab, H.H.; Gad, A.M.; Fikry, E.M.; Eid, A.H. Ellagic acid attenuates testicular disruption in rheumatoid arthritis via targeting inflammatory signals, oxidative perturbations and apoptosis. Life Sci. 2019, 239, 117012. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Gad, A.M.; Reda, E.; Yahia, R.; Eid, A.H. Activation of autophagy by sitagliptin attenuates cadmium-induced testicular impairment in rats: Targeting AMPK/mTOR and Nrf2/HO-1 pathways. Life Sci. 2021, 269, 119031. [Google Scholar] [CrossRef]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell Mol. Life Sci. 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Boo, Y.C. Arbutin as a skin depigmenting agent with antimelanogenic and antioxidant properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.H.; Gad, A.M.; Fikry, E.M.; Arab, H.H. Venlafaxine and carvedilol ameliorate testicular impairment and disrupted spermatogenesis in rheumatoid arthritis by targeting AMPK/ERK and PI3K/AKT/mTOR pathways. Toxicol. Appl. Pharmacol. 2019, 364, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Metryka, E.; Chibowska, K.; Gutowska, I.; Falkowska, A.; Kupnicka, P.; Barczak, K.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (Pb) Exposure Enhances Expression of Factors Associated with Inflammation. Int. J. Mol. Sci. 2018, 19, 1813. [Google Scholar] [CrossRef]
- Arab, H.H.; Elhemiely, A.A.; El-Sheikh, A.A.K.; Khabbaz, H.J.A.; Arafa, E.A.; Ashour, A.M.; Kabel, A.M.; Eid, A.H. Repositioning Linagliptin for the Mitigation of Cadmium-Induced Testicular Dysfunction in Rats: Targeting HMGB1/TLR4/NLRP3 Axis and Autophagy. Pharmaceuticals 2022, 15, 852. [Google Scholar] [CrossRef]
- Bazrafkan, M.; Nikmehr, B.; Shahverdi, A.; Hosseini, S.R.; Hassani, F.; Poorhassan, M.; Mokhtari, T.; Abolhassani, F.; Choobineh, H.; Beyer, C.; et al. Lipid Peroxidation and Its Role in the Expression of NLRP1a and NLRP3 Genes in Testicular Tissue of Male Rats: A Model of Spinal Cord Injury. Iran. Biomed. J. 2018, 22, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Q.; Ihsan, A.; Huang, L.; Dai, M.; Hao, H.; Cheng, G.; Liu, Z.; Wang, Y.; Yuan, Z. JAK/STAT pathway plays a critical role in the proinflammatory gene expression and apoptosis of RAW264.7 cells induced by trichothecenes as DON and T-2 toxin. Toxicol. Sci. 2012, 127, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, X.; Meng, F.; Deng, S.; Dai, H.; Bao, B.; Feng, J.; Li, H.; Wang, B. Biological Network Model of Effect of Chronic Intermittent Hypoxia on Spermatogenesis in Rats. Med. Sci. Monit. 2020, 26, e925579. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Yu, L.; Zhao, K.; Chen, B.; Li, C.; Yang, F.; Yuan, H.; Zhang, H. Effects of prokineticin 2 on testicular inflammation in rats. Am. J. Reprod. Immunol. 2018, 79, e12843. [Google Scholar] [CrossRef]
- Gao, Y.; Liao, W.; Zhang, R.; Lei, Y.; Chen, T.; Wu, L.; Li, M.; Liu, X.; Cai, F. PK2/PKRs pathway is involved in the protective effect of artemisinin against trimethyltin chloride-induced hippocampal injury. Toxicology 2023, 486, 153432. [Google Scholar] [CrossRef]
- Yang, Z.; Wu, Y.; Wang, L.; Qiu, P.; Zha, W.; Yu, W. Prokineticin 2 (PK2) rescues cardiomyocytes from high glucose/high palmitic acid-induced damage by regulating the AKT/GSK3β pathway in vitro. Oxid. Med. Cell Longev. 2020, 2020, 3163629. [Google Scholar] [CrossRef]
- Gordon, R.; Neal, M.L.; Luo, J.; Langley, M.R.; Harischandra, D.S.; Panicker, N.; Charli, A.; Jin, H.; Anantharam, V.; Woodruff, T.M.; et al. Prokineticin-2 upregulation during neuronal injury mediates a compensatory protective response against dopaminergic neuronal degeneration. Nat. Commun. 2016, 7, 12932. [Google Scholar] [CrossRef]
- Mirshahvalad, S.; Feizi, F.; Barkhordar, A.; Bahadoram, M.; Houshmand, G.; Pouramir, M. Hepatoprotective effects of arbutin against liver damage induced by carbon tetrachloride in rats. Jundishapur J. Nat. Pharm. Prod. 2016, 11, e33392. [Google Scholar] [CrossRef]
- Dadgar, M.; Pouramir, M.; Dastan, Z.; Ghasemi-Kasman, M.; Ashrafpour, M.; Moghadamnia, A.A.; Khafri, S.; Pourghasem, M. Arbutin attenuates behavioral impairment and oxidative stress in an animal model of Parkinson’s disease. Avicenna J. Phytomed 2018, 8, 533–542. [Google Scholar] [PubMed]
- Arab, H.H.; Eid, A.H.; Mahmoud, A.M.; Senousy, M.A. Linagliptin mitigates experimental inflammatory bowel disease in rats by targeting inflammatory and redox signaling. Life Sci. 2021, 273, 119295. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Arab, H.H.; Eid, A.H.; Gad, A.M.; Yahia, R.; Mahmoud, A.M.; Kabel, A.M. Inhibition of oxidative stress and apoptosis by camel milk mitigates cyclosporine-induced nephrotoxicity: Targeting Nrf2/HO-1 and AKT/eNOS/NO pathways. J. Food Sci. Nutr. 2021, 9, 3177–3190. [Google Scholar] [CrossRef]
- Arab, H.H.; Abd El Aal, H.A.; Alsufyani, S.E.; El-Sheikh, A.A.K.; Arafa, E.A.; Ashour, A.M.; Kabel, A.M.; Eid, A.H. Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy. Pharmaceuticals 2022, 15, 1402. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arab, H.H.; Alsufyani, S.E.; Ashour, A.M.; Gad, A.M.; Elhemiely, A.A.; Gadelmawla, M.H.A.; Mahmoud, M.A.; Khames, A. Targeting JAK2/STAT3, NLRP3/Caspase-1, and PK2/PKR2 Pathways with Arbutin Ameliorates Lead Acetate-Induced Testicular Injury in Rats. Pharmaceuticals 2024, 17, 909. https://doi.org/10.3390/ph17070909
Arab HH, Alsufyani SE, Ashour AM, Gad AM, Elhemiely AA, Gadelmawla MHA, Mahmoud MA, Khames A. Targeting JAK2/STAT3, NLRP3/Caspase-1, and PK2/PKR2 Pathways with Arbutin Ameliorates Lead Acetate-Induced Testicular Injury in Rats. Pharmaceuticals. 2024; 17(7):909. https://doi.org/10.3390/ph17070909
Chicago/Turabian StyleArab, Hany H., Shuruq E. Alsufyani, Ahmed M. Ashour, Amany M. Gad, Alzahraa A. Elhemiely, Mohamed H. A. Gadelmawla, Marwa Ahmed Mahmoud, and Ali Khames. 2024. "Targeting JAK2/STAT3, NLRP3/Caspase-1, and PK2/PKR2 Pathways with Arbutin Ameliorates Lead Acetate-Induced Testicular Injury in Rats" Pharmaceuticals 17, no. 7: 909. https://doi.org/10.3390/ph17070909
APA StyleArab, H. H., Alsufyani, S. E., Ashour, A. M., Gad, A. M., Elhemiely, A. A., Gadelmawla, M. H. A., Mahmoud, M. A., & Khames, A. (2024). Targeting JAK2/STAT3, NLRP3/Caspase-1, and PK2/PKR2 Pathways with Arbutin Ameliorates Lead Acetate-Induced Testicular Injury in Rats. Pharmaceuticals, 17(7), 909. https://doi.org/10.3390/ph17070909