

Effects of Hispidulin on the Osteo/Odontogenic and Endothelial Differentiation of Dental Pulp Stem Cells

 ,
,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
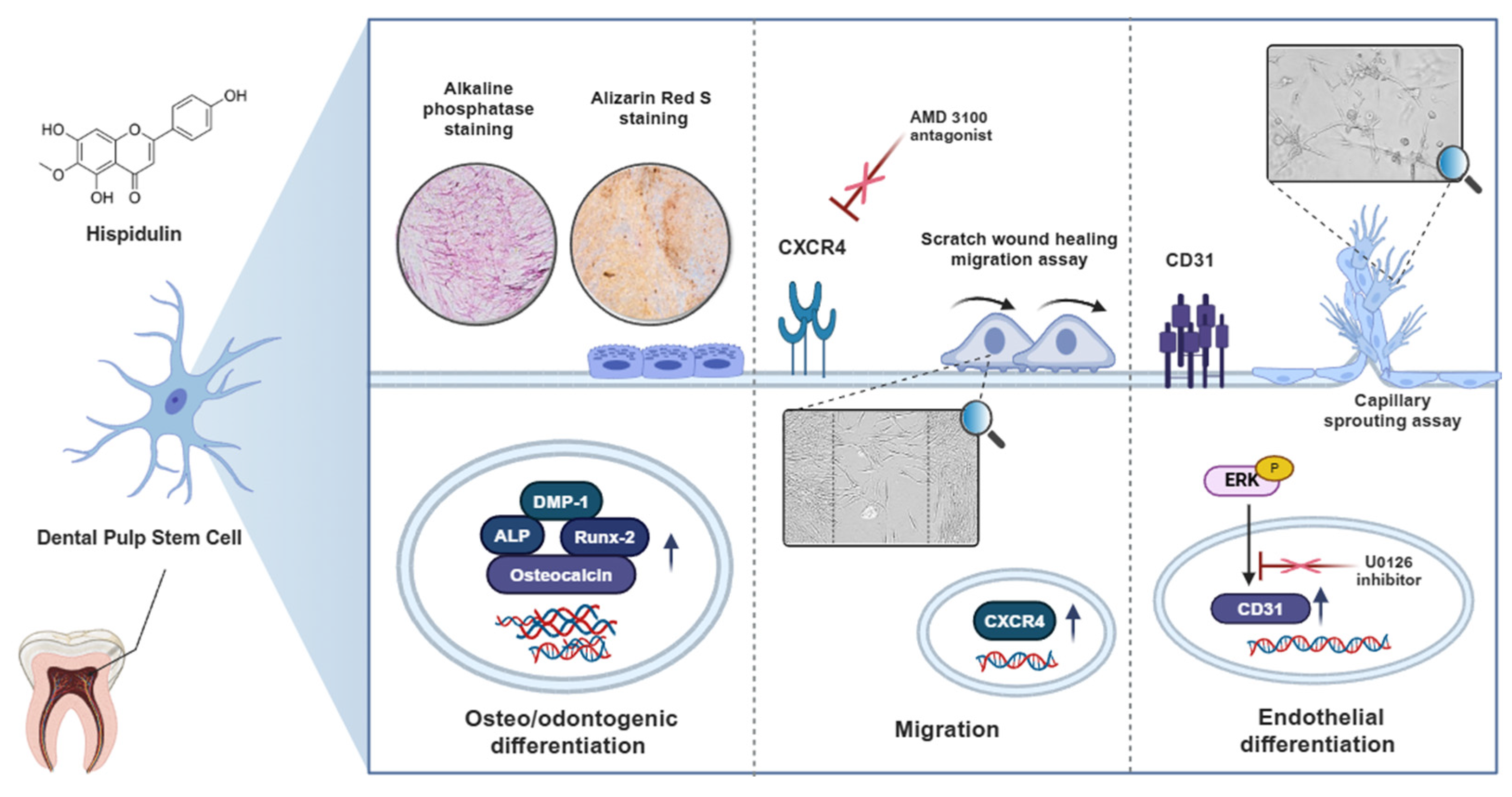
2. Results
2.1. Hispidulin Accelerates the Osteo/Odontogenic Differentiation of HDPSCs
2.2. Hispidulin Increases the Migration Ability of HDPSCs
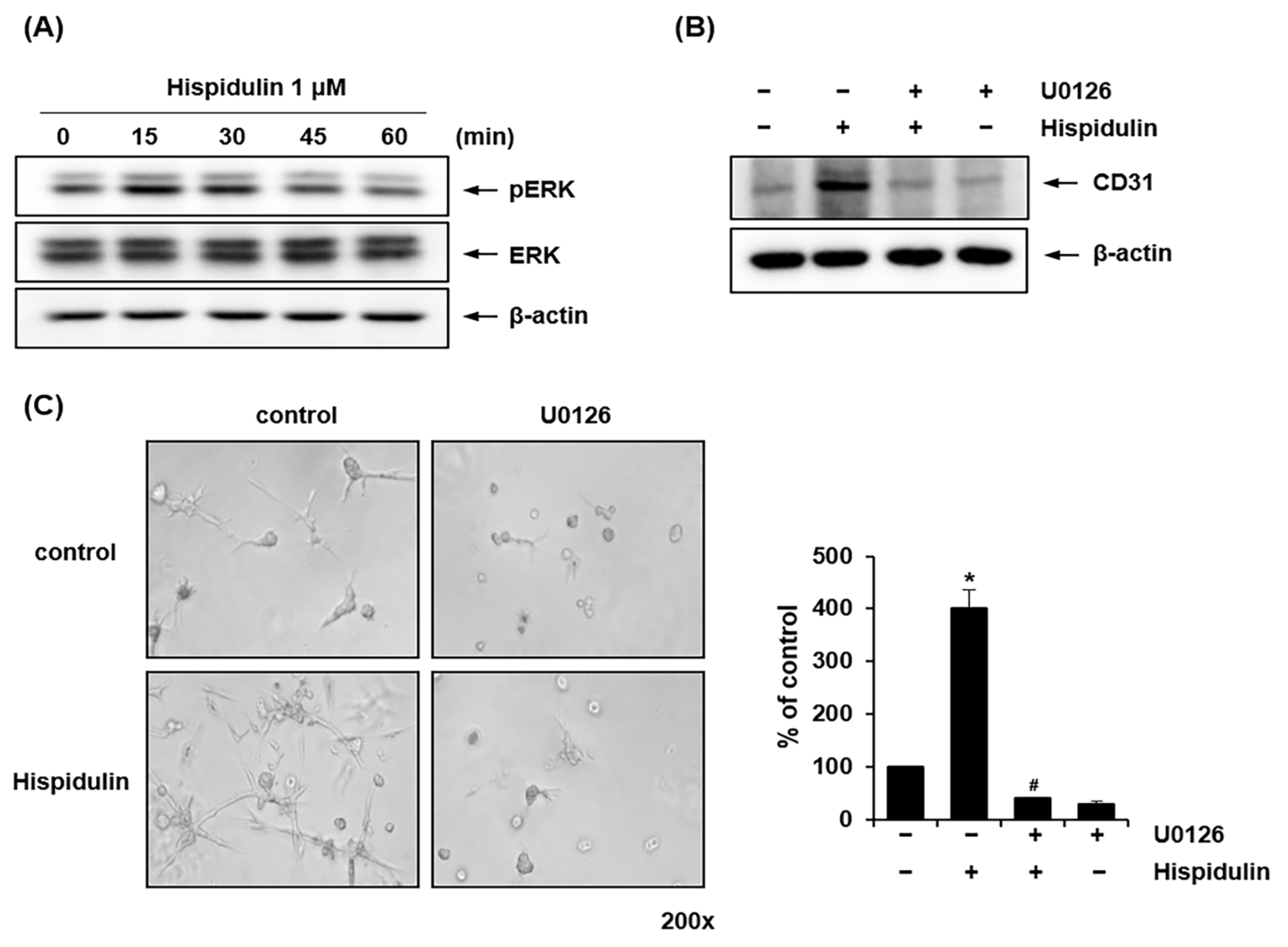
2.3. Hispidulin Enhances the Endothelial Differentiation of HDPSCs via CD31 Expression
2.4. ERK1/2 Mediates Hispidulin-Induced Endothelial Differentiation of HDPSCs
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Cell Proliferation
4.4. ALP Staining
4.5. ARS Staining
4.6. Scratch Wound Healing Migration Assay
4.7. Reverse Transcription-Quantitative PCR
4.8. Western Immunoblotting
4.9. Flow Cytometry Analysis
4.10. Capillary Sprouting Assay
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sui, B.; Wu, D.; Xiang, L.; Fu, Y.; Kou, X.; Shi, S. Dental Pulp Stem Cells: From Discovery to Clinical Application. J. Endod. 2020, 46, S46–S55. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Ohshima, H. Biological Characteristics of Dental Pulp Stem Cells and their Potential use in Regenerative Medicine. J. Oral Biosci. 2022, 64, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Katata, C.; Sasaki, J.I.; Li, A.; Abe, G.L.; Nör, J.E.; Hayashi, M.; Imazato, S. Fabrication of Vascularized DPSC Constructs for Efficient Pulp Regeneration. J. Dent. Res. 2021, 100, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.M.; Dong, Z.; Kaneko, T.; Zhang, Z.; Miyazawa, M.; Shi, S.; Smith, A.J.; Nör, J.E. Dental Pulp Tissue Engineering with Stem Cells from Exfoliated Deciduous Teeth. J. Endod. 2008, 34, 962–969. [Google Scholar] [CrossRef]
- Piva, E.; Silva, A.F.; Nör, J.E. Functionalized Scaffolds to Control Dental Pulp Stem Cell Fate. J. Endod. 2014, 40, 33. [Google Scholar] [CrossRef]
- Atif, M.; Ali, I.; Hussain, A.; Hyder, S.V.; Niaz, B.; Khan, F.A.; Maalik, A.; Farooq, U. Pharmacological Assessment of Hispidulin--a Natural Bioactive Flavone. Acta Pol. Pharm. 2015, 72, 829–842. [Google Scholar] [PubMed]
- Patel, K.; Patel, D.K. Medicinal Importance, Pharmacological Activities, and Analytical Aspects of Hispidulin: A Concise Report. J. Tradit. Complement. Med. 2016, 7, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Ashaq, A.; Maqbool, M.F.; Maryam, A.; Khan, M.; Shakir, H.A.; Irfan, M.; Qazi, J.I.; Li, Y.; Ma, T. Hispidulin: A Novel Natural Compound with Therapeutic Potential Against Human Cancers. Phytother. Res. 2021, 35, 771–789. [Google Scholar] [CrossRef]
- Kut, K.; Bartosz, G.; Soszyński, M.; Sadowska-Bartosz, I. Antioxidant Properties of Hispidulin. Nat. Prod. Res. 2022, 36, 6401–6404. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lee, H.; Park, H.; Kim, M.; Kim, Y.; Kim, H.J.; Bae, S.; Kim, Y.; Bae, M. Hispidulin Inhibits the Vascular Inflammation Triggered by Porphyromonas Gingivalis Lipopolysaccharide. Molecules 2023, 28, 6717. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, S.; Magaji, R.A.; Magaji, M.G.; Gaya, I.B.; Umar, B.; Yusha’u, Y.; Daku, A.B.; Chiroma, S.M.; Jaafar, A.; Mehat, M.Z.; et al. Neuroprotective Roles of Flavonoid “Hispidulin” in the Central Nervous System: A Review. Iran. J. Basic Med. Sci. 2024, 27, 1077–1084. [Google Scholar]
- Jeong, S.; Kim, Y.; Lee, D.; Kim, S.; Lee, S. Hispidulin Alleviates Mast Cell-Mediated Allergic Airway Inflammation through FcεR1 and Nrf2/HO-1 Signaling Pathway. Antioxidants 2024, 13, 528. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, Y.; Wu, C.; Zhang, X.; Zhang, X.; Xu, X. Targeting Bone Homeostasis Regulation: Potential of Traditional Chinese Medicine Flavonoids in the Treatment of Osteoporosis. Front. Pharmacol. 2024, 15, 1361864. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, D.; Liu, Q.; Xie, F.; Xiang, D.; Tang, G.; Luo, S. Osteogenesis Activity of Isocoumarin a through the Activation of the PI3K-Akt/Erk Cascade-Activated BMP/RUNX2 Signaling Pathway. Eur. J. Pharmacol. 2019, 858, 172480. [Google Scholar] [CrossRef] [PubMed]
- Nepal, M.; Choi, H.J.; Choi, B.; Yang, M.; Chae, J.; Li, L.; Soh, Y. Hispidulin Attenuates Bone Resorption and Osteoclastogenesis Via the RANKL-Induced NF-κB and NFATc1 Pathways. Eur. J. Pharmacol. 2013, 715, 96–104. [Google Scholar] [CrossRef]
- Li, M.; Sun, X.; Ma, L.; Jin, L.; Zhang, W.; Xiao, M.; Yu, Q. SDF-1/CXCR4 Axis Induces Human Dental Pulp Stem Cell Migration through FAK/PI3K/Akt and GSK3β/Β-Catenin Pathways. Sci. Rep. 2017, 7, 40161. [Google Scholar] [CrossRef]
- Kim, Y.; Park, H.; Kim, M.; Kim, Y.; Kim, H.J.; Bae, S.; Nör, J.E.; Bae, M. Naringenin Stimulates Osteogenic/Odontogenic Differentiation and Migration of Human Dental Pulp Stem Cells. J. Dent. Sci. 2023, 18, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Sakai, V.T.; Zhang, Z.; Dong, Z.; Neiva, K.G.; Machado, M.A.A.M.; Shi, S.; Santos, C.F.; Nör, J.E. SHED Differentiate into Functional Odontoblasts and Endothelium. J. Dent. Res. 2010, 89, 791–796. [Google Scholar] [CrossRef]
- Li, A.; Sasaki, J.I.; Inubushi, T.; Abe, G.L.; Nör, J.E.; Yamashiro, T.; Imazato, S. Role of Heparan Sulfate in Vasculogenesis of Dental Pulp Stem Cells. J. Dent. Res. 2023, 102, 207–216. [Google Scholar] [CrossRef]
- Bento, L.W.; Zhang, Z.; Imai, A.; Nör, F.; Dong, Z.; Shi, S.; Araujo, F.B.; Nör, J.E. Endothelial Differentiation of SHED Requires MEK1/ERK Signaling. J. Dent. Res. 2013, 92, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Z.; Luo, Y.; Li, X.; Huang, G.; Chen, H.; Li, A.; Qin, S. The Role of Flavonoids in the Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Pharmacol. 2022, 13, 849513. [Google Scholar] [CrossRef]
- Sudarshan, K.; Boda, A.K.; Dogra, S.; Bose, I.; Yadav, P.N.; Aidhen, I.S. Discovery of an Isocoumarin Analogue that Modulates Neuronal Functions Via Neurotrophin Receptor TrkB. Bioorg. Med. Chem. Lett. 2019, 29, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, H.; Zhang, M.; Wu, Y.; Xu, X.; Yang, M.; Mei, L. Assessing the Effect and Related Mechanism of Naringenin on the Proliferation, Osteogenic Differentiation and Endothelial Differentiation of Human Periodontal Ligament Stem Cells. Biochem. Biophys. Res. Commun. 2021, 534, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Hu, G.; Li, J.; Zheng, J.; Li, Y. Icariin Accelerates Bone Regeneration by Inducing Osteogenesis-Angiogenesis Coupling in Rats with Type 1 Diabetes Mellitus. World J. Diabetes 2024, 15, 769–782. [Google Scholar] [CrossRef]
- Grellier, M.; Granja, P.L.; Fricain, J.; Bidarra, S.J.; Renard, M.; Bareille, R.; Bourget, C.; Amédée, J.; Barbosa, M.A. The Effect of the Co-Immobilization of Human Osteoprogenitors and Endothelial Cells within Alginate Microspheres on Mineralization in a Bone Defect. Biomaterials 2009, 30, 3271–3278. [Google Scholar] [CrossRef]
- Li, A.; Sasaki, J.; Abe, G.L.; Katata, C.; Sakai, H.; Imazato, S. Vascularization of a Bone Organoid Using Dental Pulp Stem Cells. Stem Cells Int. 2023, 2023, 5367887. [Google Scholar] [CrossRef] [PubMed]
- Mantesso, A.; Zhang, Z.; Warner, K.A.; Herzog, A.E.; Pulianmackal, A.J.; Nör, J.E. Pulpbow: A Method to Study the Vasculogenic Potential of Mesenchymal Stem Cells from the Dental Pulp. Cells 2021, 10, 2804. [Google Scholar] [CrossRef] [PubMed]
- Duncan, H.F.; Cooper, P.R.; Smith, A.J. Dissecting Dentine-Pulp Injury and Wound Healing Responses: Consequences for Regenerative Endodontics. Int. Endod. J. 2019, 52, 261–266. [Google Scholar] [CrossRef]
- Yi, B.; Dissanayaka, W.L.; Zhang, C. Growth Factors and Small-Molecule Compounds in Derivation of Endothelial Lineages from Dental Stem Cells. J. Endod. 2020, 46, S63–S70. [Google Scholar] [CrossRef]
- Li, J.; Zhu, Y.; Li, N.; Wu, T.; Zheng, X.; Heng, B.C.; Zou, D.; Xu, J. Upregulation of ETV2 Expression Promotes Endothelial Differentiation of Human Dental Pulp Stem Cells. Cell Transplant. 2021, 30, 963689720978739. [Google Scholar] [CrossRef]
- Sakai, V.T.; Cordeiro, M.M.; Dong, Z.; Zhang, Z.; Zeitlin, B.D.; Nör, J.E. Tooth Slice/Scaffold Model of Dental Pulp Tissue Engineering. Adv. Dent. Res. 2011, 23, 325–332. [Google Scholar] [CrossRef]
- Prescott, R.S.; Alsanea, R.; Fayad, M.I.; Johnson, B.R.; Wenckus, C.S.; Hao, J.; John, A.S.; George, A. In Vivo Generation of Dental Pulp-Like Tissue by using Dental Pulp Stem Cells, a Collagen Scaffold, and Dentin Matrix Protein 1 After Subcutaneous Transplantation in Mice. J. Endod. 2008, 34, 421–426. [Google Scholar] [CrossRef]
- Xu, J.; Liu, X.; Jiang, Y.; Chu, L.; Hao, H.; Liua, Z.; Verfaillie, C.; Zweier, J.; Gupta, K.; Liu, Z. MAPK/ERK Signaling Mediates VEGF-Induced Bone Marrow Stem Cell Differentiation into Endothelial Cell. J. Cell. Mol. Med. 2008, 12, 2395–2406. [Google Scholar] [CrossRef] [PubMed]
- Almalki, S.G.; Agrawal, D.K. ERK Signaling is Required for VEGF-A/VEGFR2-Induced Differentiation of Porcine Adipose-Derived Mesenchymal Stem Cells into Endothelial Cells. Stem Cell Res. Ther. 2017, 8, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.A.; Lee, J. Hispidulin Modulates Epithelial-Mesenchymal Transition in Breast Cancer Cells. Oncol. Lett. 2021, 21, 155. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.G.; Gong, T.; Wang, Y.Y.; Zou, T.; Heng, B.C.; Yang, Y.Q.; Zhang, C.F. Inhibition of TGF-β Signaling in SHED Enhances Endothelial Differentiation. J. Dent. Res. 2018, 97, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Nör, F.; Oh, M.; Cucco, C.; Shi, S.; Nör, J.E. Wnt/Β-Catenin Signaling Determines the Vasculogenic Fate of Postnatal Mesenchymal Stem Cells. Stem Cells 2016, 34, 1576–1587. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Park, H.-J.; Kim, M.-K.; Kim, H.J.; Kim, Y.-I.; Bae, S.-K.; Bae, M.-K. Effects of Hispidulin on the Osteo/Odontogenic and Endothelial Differentiation of Dental Pulp Stem Cells. Pharmaceuticals 2024, 17, 1740. https://doi.org/10.3390/ph17121740
Kim Y, Park H-J, Kim M-K, Kim HJ, Kim Y-I, Bae S-K, Bae M-K. Effects of Hispidulin on the Osteo/Odontogenic and Endothelial Differentiation of Dental Pulp Stem Cells. Pharmaceuticals. 2024; 17(12):1740. https://doi.org/10.3390/ph17121740
Chicago/Turabian StyleKim, Yeon, Hyun-Joo Park, Mi-Kyoung Kim, Hyung Joon Kim, Yong-Il Kim, Soo-Kyung Bae, and Moon-Kyoung Bae. 2024. "Effects of Hispidulin on the Osteo/Odontogenic and Endothelial Differentiation of Dental Pulp Stem Cells" Pharmaceuticals 17, no. 12: 1740. https://doi.org/10.3390/ph17121740
APA StyleKim, Y., Park, H.-J., Kim, M.-K., Kim, H. J., Kim, Y.-I., Bae, S.-K., & Bae, M.-K. (2024). Effects of Hispidulin on the Osteo/Odontogenic and Endothelial Differentiation of Dental Pulp Stem Cells. Pharmaceuticals, 17(12), 1740. https://doi.org/10.3390/ph17121740