

N-(coumarin-3-yl)cinnamamide Promotes Immunomodulatory, Neuroprotective, and Lung Function-Preserving Effects during Severe Malaria

 , , ,
, , ,  ,
,  and
and 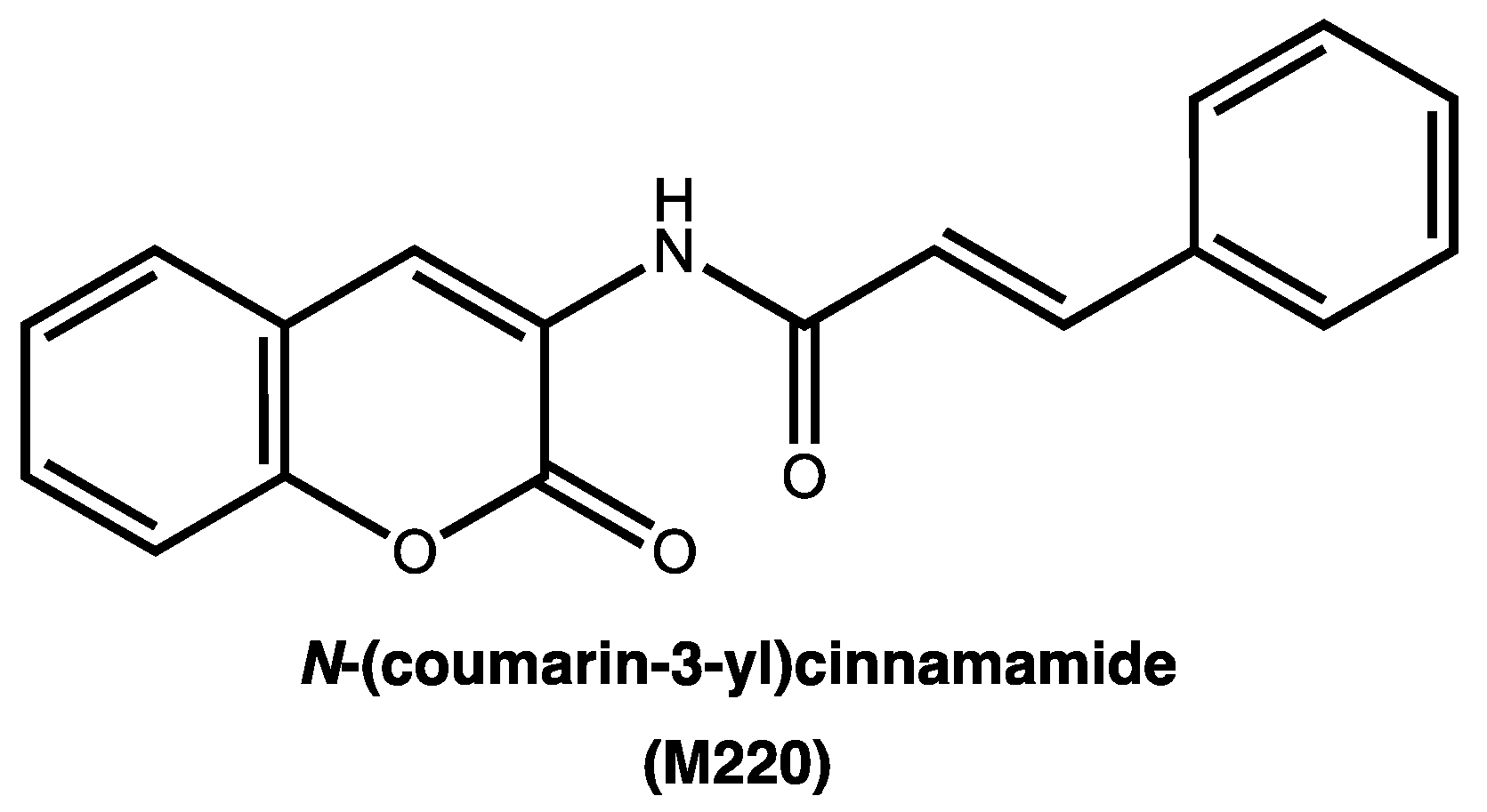{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. The synthesis of N-(coumarin-3-yl)cinnamamide (M220)
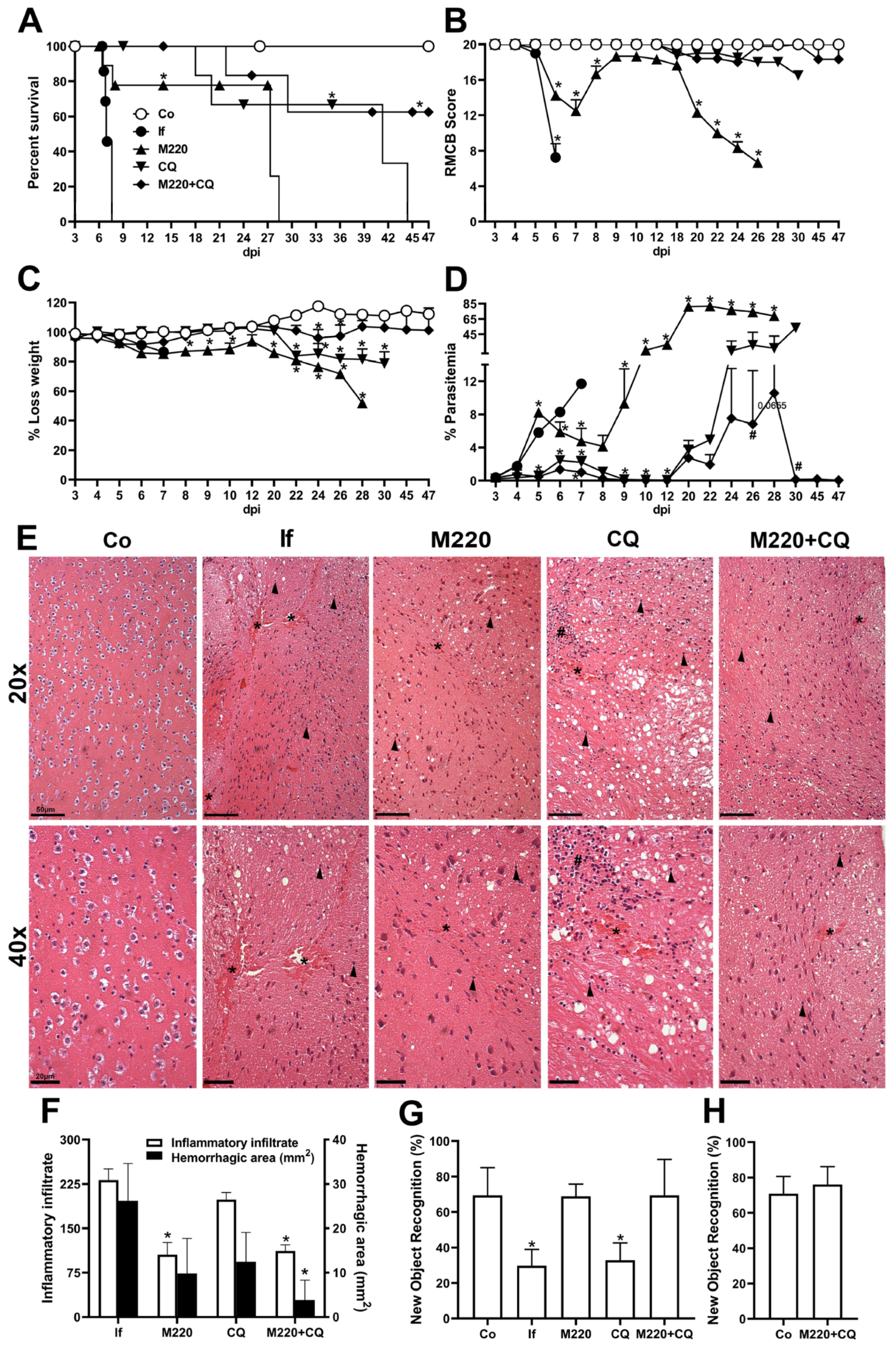
2.2. Administration of M220 Protects Mice from Mortality and Preserves Memory in PbA-Infected Mice
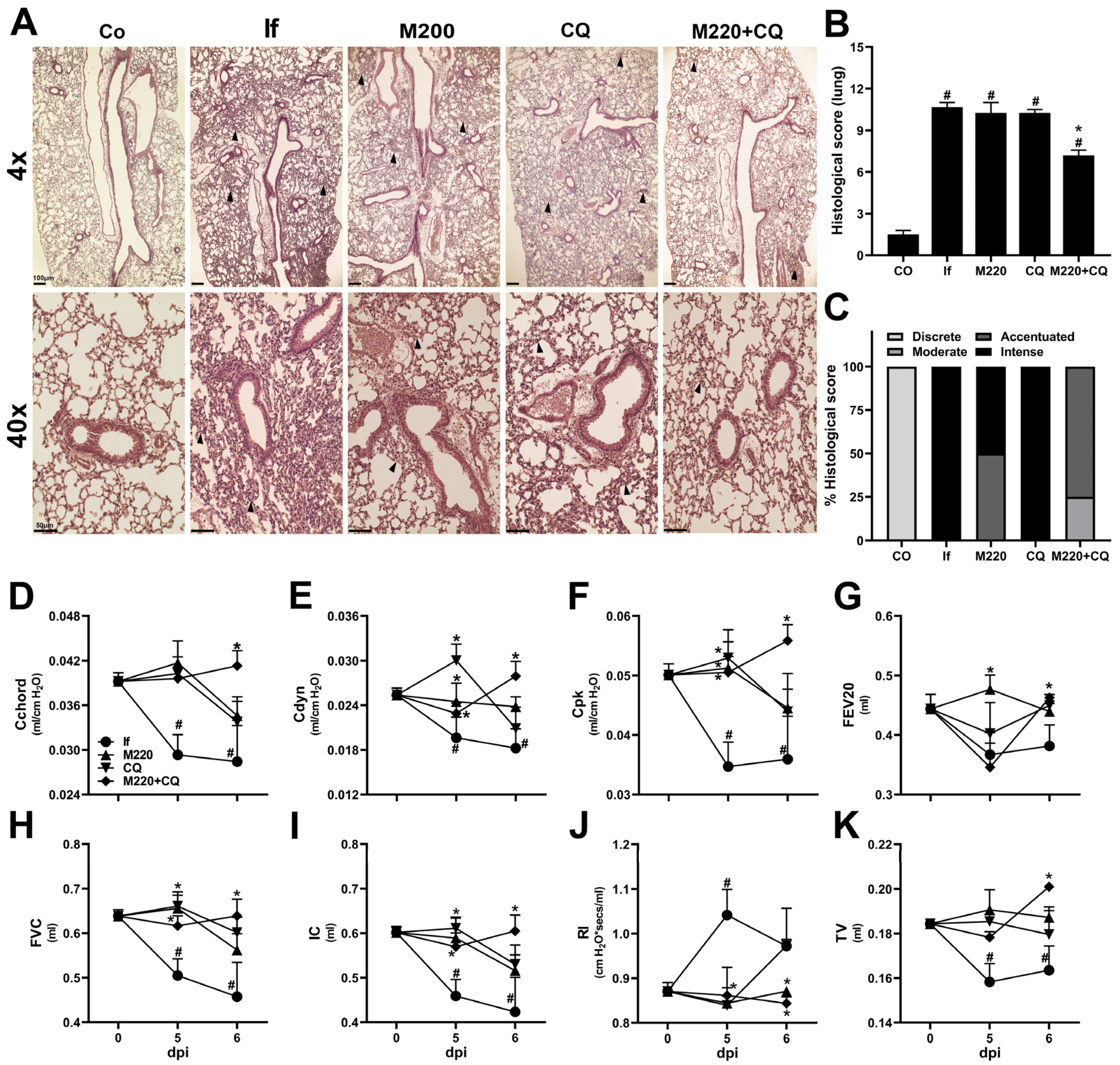
2.3. Administration of M220 Attenuates Lung Mechanical Dysfunction during PbA Infection in Mice
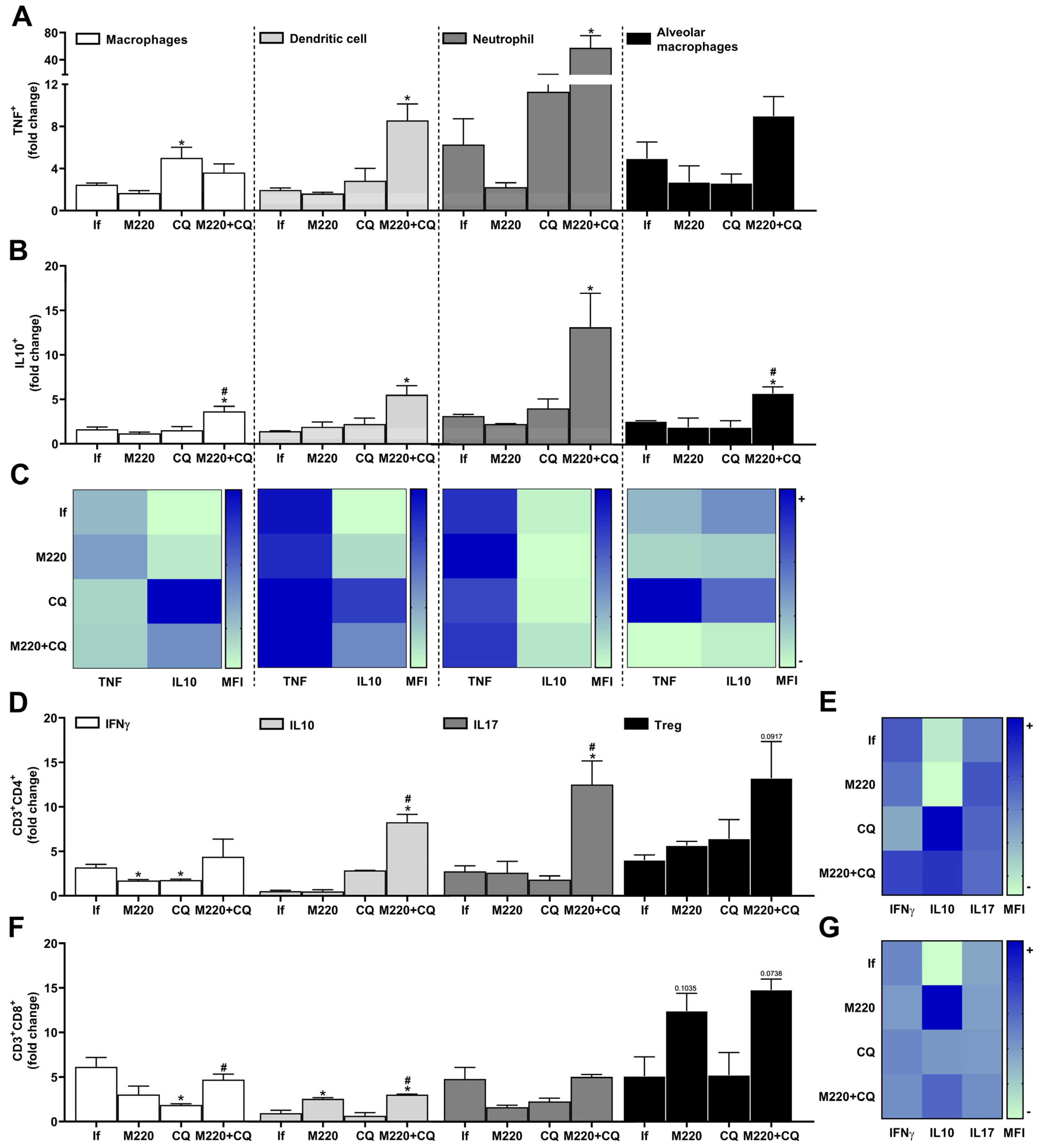
2.4. Effects of M220 on Brain Inflammation during PbA Infection in Mice
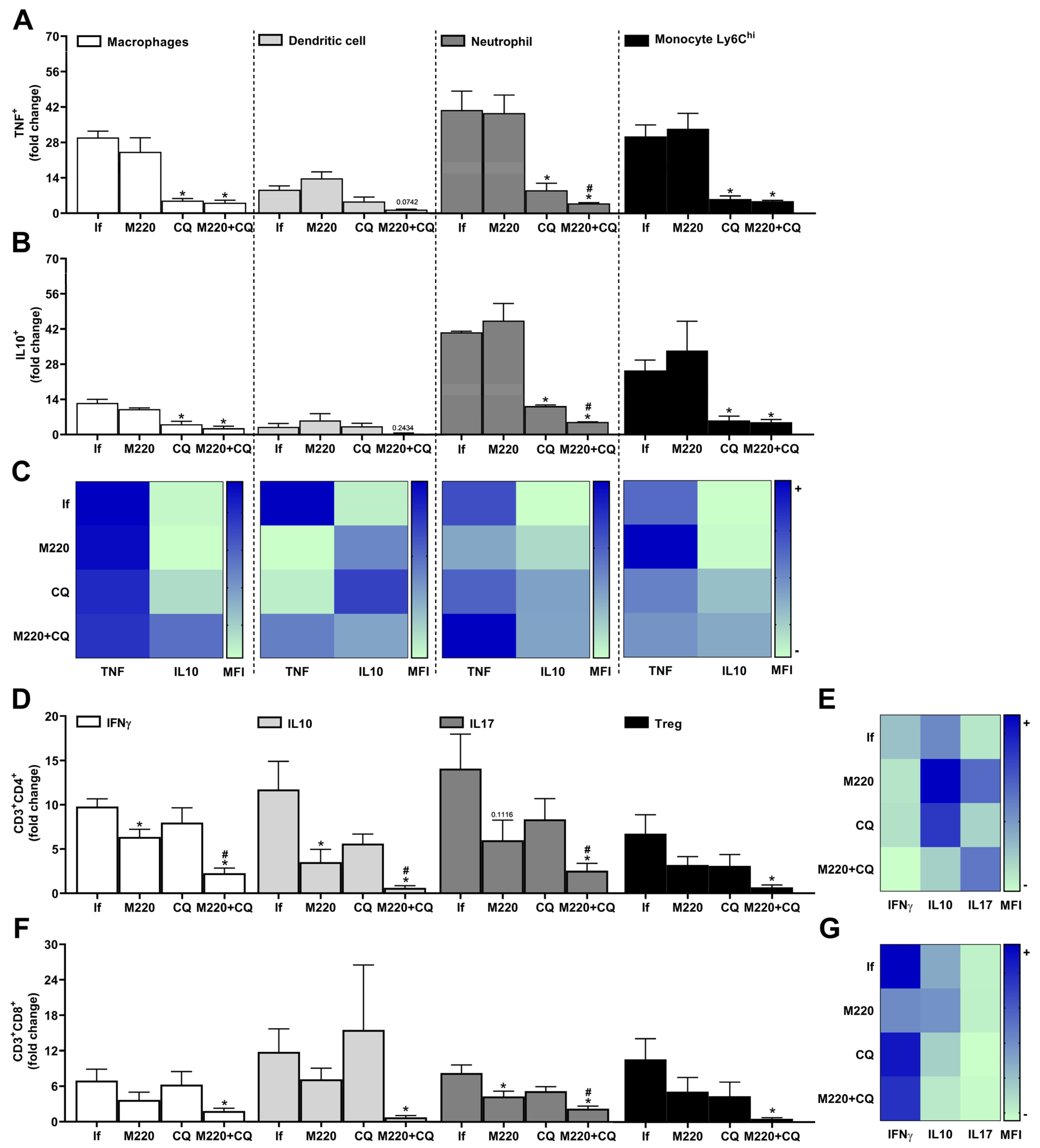
2.5. Effects of M220 on PbA-Induced Inflammatory Response in the Lungs
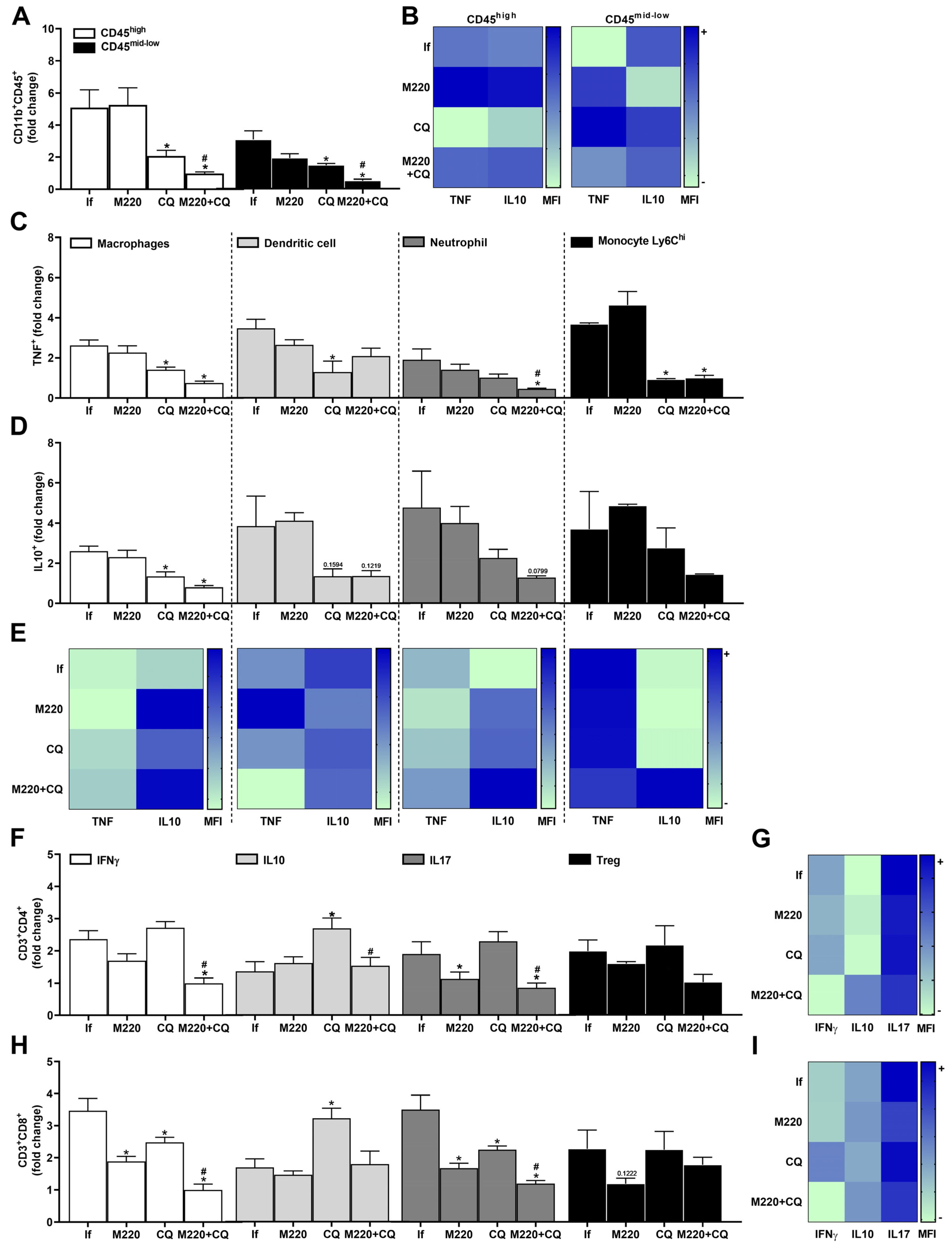
2.6. Effects of M220 on Innate and Adaptive Immune Cell Populations in the Spleen of PbA-Infected Mice
3. Discussion
4. Materials and Methods
4.1. Synthesis of N-(coumarin-3-yl)cinnamamide (M220)
4.2. Ethics Statement
4.3. Infection
4.4. Treatments
4.5. Novel Object Recognition Test
4.6. Spirometry
4.7. Immunophenotyping by Flow Cytometry
4.8. Histopathological and Morphometric Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, M.A.; Burrows, J.N.; Manyando, C.; van Huijsduijnen, R.H.; Van Voorhis, W.C.; Wells, T.N.C. Malaria. Nat. Rev. Dis. Primers 2017, 3, 17050. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, S.C.; Grau, G.E.R. Severe Malaria: What’s New on the Pathogenesis Front? Int. J. Parasitol. 2017, 47, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Vandermosten, L.; Pham, T.-T.; Possemiers, H.; Knoops, S.; Van Herck, E.; Deckers, J.; Franke-Fayard, B.; Lamb, T.J.; Janse, C.J.; Opdenakker, G.; et al. Experimental Malaria-Associated Acute Respiratory Distress Syndrome Is Dependent on the Parasite-Host Combination and Coincides with Normocyte Invasion. Malar. J. 2018, 17, 102. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. In World Malaria Report; World Health Organization: Geneva, Switzerland, 2022.
- Craig, A.G.; Grau, G.E.; Janse, C.; Kazura, J.W.; Milner, D.; Barnwell, J.W.; Turner, G.; Langhorne, J. The Role of Animal Models for Research on Severe Malaria. PLoS Pathog. 2012, 8, e1002401. [Google Scholar] [CrossRef]
- Brant, F.; Miranda, A.S.; Esper, L.; Gualdrón-López, M.; Cisalpino, D.; de Souza, D.d.G.; Rachid, M.A.; Tanowitz, H.B.; Teixeira, M.M.; Teixeira, A.L.; et al. Suppressor of Cytokine Signaling 2 Modulates the Immune Response Profile and Development of Experimental Cerebral Malaria. Brain Behav. Immun. 2016, 54, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Deroost, K.; Pham, T.-T.; Opdenakker, G.; Van den Steen, P.E. The Immunological Balance between Host and Parasite in Malaria. FEMS Microbiol. Rev. 2016, 40, 208–257. [Google Scholar] [CrossRef] [PubMed]
- Gowda, D.C.; Wu, X. Parasite Recognition and Signaling Mechanisms in Innate Immune Responses to Malaria. Front. Immunol. 2018, 9, 3006. [Google Scholar] [CrossRef]
- Van den Steen, P.E.; Deroost, K.; Aelst, I.V.; Geurts, N.; Martens, E.; Struyf, S.; Nie, C.Q.; Hansen, D.S.; Matthys, P.; Damme, J.V.; et al. CXCR3 Determines Strain Susceptibility to Murine Cerebral Malaria by Mediating T Lymphocyte Migration toward IFN-γ-Induced Chemokines. Eur. J. Immunol. 2008, 38, 1082–1095. [Google Scholar] [CrossRef]
- Dunst, J.; Kamena, F.; Matuschewski, K. Cytokines and Chemokines in Cerebral Malaria Pathogenesis. Front. Cell. Infect. Microbiol. 2017, 7, 324. [Google Scholar] [CrossRef]
- Lagassé, H.A.D.; Anidi, I.U.; Craig, J.M.; Limjunyawong, N.; Poupore, A.K.; Mitzner, W.; Scott, A.L. Recruited Monocytes Modulate Malaria-Induced Lung Injury through CD36-Mediated Clearance of Sequestered Infected Erythrocytes. J. Leukoc. Biol. 2016, 99, 659–671. [Google Scholar] [CrossRef]
- Claser, C.; Nguee, S.Y.T.; Balachander, A.; Wu Howland, S.; Becht, E.; Gunasegaran, B.; Hartimath, S.V.; Lee, A.W.Q.; Theng Theng Ho, J.; Bing Ong, C.; et al. Lung Endothelial Cell Antigen Cross-Presentation to CD8+T Cells Drives Malaria-Associated Lung Injury. Nat. Commun. 2019, 10, 4241. [Google Scholar] [CrossRef] [PubMed]
- Götz, A.; Tang, M.S.; Ty, M.C.; Arama, C.; Ongoiba, A.; Doumtabe, D.; Traore, B.; Crompton, P.D.; Loke, P.; Rodriguez, A. Atypical Activation of Dendritic Cells by Plasmodium Falciparum. Proc. Natl. Acad. Sci. USA 2017, 114, E10568–E10577. [Google Scholar] [CrossRef] [PubMed]
- Hirako, I.C.; Ataide, M.A.; Faustino, L.; Assis, P.A.; Sorensen, E.W.; Ueta, H.; Araújo, N.M.; Menezes, G.B.; Luster, A.D.; Gazzinelli, R.T. Splenic Differentiation and Emergence of CCR5+CXCL9+CXCL10+ Monocyte-Derived Dendritic Cells in the Brain during Cerebral Malaria. Nat. Commun. 2016, 7, 13277. [Google Scholar] [CrossRef] [PubMed]
- Freitas do Rosario, A.P.; Langhorne, J. T Cell-Derived IL-10 and Its Impact on the Regulation of Host Responses during Malaria. Int. J. Parasitol. 2012, 42, 549–555. [Google Scholar] [CrossRef]
- Nakamae, S.; Kimura, D.; Miyakoda, M.; Sukhbaatar, O.; Inoue, S.-I.; Yui, K. Role of IL-10 in Inhibiting Protective Immune Responses against Infection with Heterologous Plasmodium Parasites. Parasitol. Int. 2019, 70, 5–15. [Google Scholar] [CrossRef]
- Braga, C.B.E.; Martins, A.C.; Cayotopa, A.D.E.; Klein, W.W.; Schlosser, A.R.; da Silva, A.F.; de Souza, M.N.; Andrade, B.W.B.; Filgueira-Júnior, J.A.; de Pinto, W.J.; et al. Side Effects of Chloroquine and Primaquine and Symptom Reduction in Malaria Endemic Area (Mâncio Lima, Acre, Brazil). Interdiscip. Perspect. Infect. Dis. 2015, 2015, 346853. [Google Scholar] [CrossRef]
- Mekonnen, D.A.; Abadura, G.S.; Behaksra, S.W.; Taffese, H.S.; Bayissa, G.A.; Bulto, M.G.; Tessema, T.S.; Tadesse, F.G.; Gadisa, E. Treatment of Uncomplicated Plasmodium Vivax with Chloroquine plus Radical Cure with Primaquine without G6PDd Testing Is Safe in Arba Minch, Ethiopia: Assessment of Clinical and Parasitological Response. Malar. J. 2023, 22, 135. [Google Scholar] [CrossRef]
- Macedo, T.S.; Villarreal, W.; Couto, C.C.; Moreira, D.R.M.; Navarro, M.; Machado, M.; Prudêncio, M.; Batista, A.A.; Soares, M.B.P. Platinum(II)–Chloroquine Complexes Are Antimalarial Agents against Blood and Liver Stages by Impairing Mitochondrial Function. Metallomics 2017, 9, 1548–1561. [Google Scholar] [CrossRef]
- Al-Bari, M.A.A. Chloroquine Analogues in Drug Discovery: New Directions of Uses, Mechanisms of Actions and Toxic Manifestations from Malaria to Multifarious Diseases. J. Antimicrob. Chemother. 2015, 70, 1608–1621. [Google Scholar] [CrossRef]
- Soentjens, P.; Delanote, M.; Van Gompel, A. Mefloquine-Induced Pneumonitis. J. Travel. Med. 2006, 13, 172–174. [Google Scholar] [CrossRef][Green Version]
- Maguire, G.P.; Handojo, T.; Pain, M.C.F.; Kenangalem, E.; Price, R.N.; Tjitra, E.; Anstey, N.M. Lung Injury in Uncomplicated and Severe Falciparum Malaria: A Longitudinal Study in Papua, Indonesia. J. Infect. Dis. 2005, 192, 1966–1974. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.S.; Gupta, J.; Sharma, S.; Sahu, D. An Insight into the Therapeutic Applications of Coumarin Compounds and Their Mechanisms of Action. Eur. J. Pharm. Sci. 2020, 152, 105424. [Google Scholar] [CrossRef] [PubMed]
- Stefanachi, A.; Leonetti, F.; Pisani, L.; Catto, M.; Carotti, A. Coumarin: A Natural, Privileged and Versatile Scaffold for Bioactive Compounds. Molecules 2018, 23, 250. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, F.; Pinna, C.; Dallavalle, S.; Tamborini, L.; Pinto, A. An Overview of Coumarin as a Versatile and Readily Accessible Scaffold with Broad-Ranging Biological Activities. Int. J. Mol. Sci. 2020, 21, 4618. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.J.; Rodríguez-Enríquez, F.; Borges, F.; Santana, L.; Uriarte, E.; Estrada, M.; Rodríguez-Franco, M.I.; Laguna, R.; Viña, D. 3-Amidocoumarins as Potential Multifunctional Agents against Neurodegenerative Diseases. ChemMedChem 2015, 10, 2071–2079. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Hu, J.; Song, X.; Wu, D.; Kong, L.; Sun, Y.; Wang, D.; Wang, Y.; Chen, N.; Liu, G. Coumarin Derivatives Protect against Ischemic Brain Injury in Rats. Eur. J. Med. Chem. 2013, 67, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.J.; Vilar, S.; Gonzalez-Franco, R.M.; Uriarte, E.; Santana, L.; Friedman, C.; Tatonetti, N.P.; Viña, D.; Fontenla, J.A. Novel (Coumarin-3-Yl)Carbamates as Selective MAO-B Inhibitors: Synthesis, in Vitro and in Vivo Assays, Theoretical Evaluation of ADME Properties and Docking Study. Eur. J. Med. Chem. 2013, 63, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Enríquez, F.; Viña, D.; Uriarte, E.; Laguna, R.; Matos, M.J. 7-Amidocoumarins as Multitarget Agents against Neurodegenerative Diseases: Substitution Pattern Modulation. ChemMedChem 2021, 16, 179–186. [Google Scholar] [CrossRef]
- Matos, M.J.; Novo, P.; Mayán, L.; Torres, I.; Uriarte, E.; Yáñez, M.; Fontenla, J.Á.; Ortuso, F.; Alcaro, S.; Procopio, F.; et al. 8-Amide and 8-Carbamate Substitution Patterns as Modulators of 7-Hydroxy-4-Methylcoumarin’s Antidepressant Profile: Synthesis, Biological Evaluation and Docking Studies. Eur. J. Med. Chem. 2023, 248, 115091. [Google Scholar] [CrossRef]
- Dwivedi, A.P.; Kumar, S.; Varshney, V.; Singh, A.B.; Srivastava, A.K.; Sahu, D.P. Synthesis and Antihyperglycemic Activity of Novel N-Acyl-2-Arylethylamines and N-Acyl-3-Coumarylamines. Bioorganic Med. Chem. Lett. 2008, 18, 2301–2305. [Google Scholar] [CrossRef]
- Taylor, W.R.J.; Hanson, J.; Turner, G.D.H.; White, N.J.; Dondorp, A.M. Respiratory Manifestations of Malaria. Chest 2012, 142, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Sharma, S.K.; Bollineni, S. Acute Lung Injury and Acute Respiratory Distress Syndrome in Malaria. J. Vector Borne Dis. 2008, 45, 179–193. [Google Scholar] [PubMed]
- Ghosh, D.; Stumhofer, J.S. The Spleen: “Epicenter” in Malaria Infection and Immunity. J. Leukoc. Biol. 2021, 110, 753–769. [Google Scholar] [CrossRef] [PubMed]
- Zekar, L.; Sharman, T. Plasmodium Falciparum Malaria; StatPearls Publishing: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Demissie, Y.; Ketema, T. Complicated Malaria Symptoms Associated with Plasmodium Vivax among Patients Visiting Health Facilities in Mendi Town, Northwest Ethiopia. BMC Infect. Dis. 2016, 16, 436. [Google Scholar] [CrossRef]
- Brant, F.; Miranda, A.S.; Esper, L.; Rodrigues, D.H.; Kangussu, L.M.; Bonaventura, D.; Soriani, F.M.; Pinho, V.; Souza, D.G.; Rachid, M.A.; et al. Role of the Aryl Hydrocarbon Receptor in the Immune Response Profile and Development of Pathology during Plasmodium Berghei Anka Infection. Infect. Immun. 2014, 82, 3127–3140. [Google Scholar] [CrossRef] [PubMed]
- Bartoloni, A.; Zammarchi, L. Clinical Aspects of Uncomplicated and Severe Malaria. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012026. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Reznik, S.E.; Spray, D.C.; Weiss, L.M.; Tanowitz, H.B.; Gulinello, M.; Desruisseaux, M.S. Persistent Cognitive and Motor Deficits after Successful Antimalarial Treatment in Murine Cerebral Malaria. Microbes Infect. 2010, 12, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Treatment of Malaria, 3rd ed.; World Health Organization: Geneva, Switzerland, 2015; Available online: https://apps.who.int/iris/handle/10665/162441 (accessed on 8 December 2020)ISBN 978-92-4-154912-7.
- Frimpong, A.; Amponsah, J.; Adjokatseh, A.S.; Agyemang, D.; Bentum-Ennin, L.; Ofori, E.A.; Kyei-Baafour, E.; Akyea-Mensah, K.; Adu, B.; Mensah, G.I.; et al. Asymptomatic Malaria Infection Is Maintained by a Balanced Pro- and Anti-Inflammatory Response. Front. Microbiol. 2020, 11, 559255. [Google Scholar] [CrossRef]
- Davis, S.N.; Wu, P.; Camci, E.D.; Simon, J.A.; Rubel, E.W.; Raible, D.W. Chloroquine Kills Hair Cells in Zebrafish Lateral Line and Murine Cochlear Cultures: Implications for Ototoxicity. Hear. Res. 2020, 395, 108019. [Google Scholar] [CrossRef]
- Eyo, U.B.; Murugan, M.; Wu, L.-J. Microglia-Neuron Communication in Epilepsy: Microglia in Epilepsy. Glia 2017, 65, 5–18. [Google Scholar] [CrossRef]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cheeran, M.; Lokensgard, J.R.; Peterson, P.K. Role of Microglia in Central Nervous System Infections. Clin. Microbiol. Rev. 2004, 17, 942–964. [Google Scholar] [CrossRef] [PubMed]
- Figarella, K.; Wolburg, H.; Garaschuk, O.; Duszenko, M. Microglia in Neuropathology Caused by Protozoan Parasites. Biol. Rev. 2020, 95, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Spanos, J.P.; Hsu, N.-J.; Jacobs, M. Microglia Are Crucial Regulators of Neuro-Immunity during Central Nervous System Tuberculosis. Front. Cell. Neurosci. 2015, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sundar, S. Combined Neutralization of Interferon Gamma and Tumor Necrosis Factor Alpha Induces IL-4 Production but Has No Direct Additive Impact on Parasite Burden in Splenic Cultures of Human Visceral Leishmaniasis. PLoS ONE 2018, 13, e0199817. [Google Scholar] [CrossRef] [PubMed]
- Hirako, I.C.; Assis, P.A.; Galvão-Filho, B.; Luster, A.D.; Antonelli, L.R.; Gazzinelli, R.T. Monocyte-Derived Dendritic Cells in Malaria. Curr. Opin. Microbiol. 2019, 52, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Swanson, P.A.; Hart, G.T.; Russo, M.V.; Nayak, D.; Yazew, T.; Peña, M.; Khan, S.M.; Janse, C.J.; Pierce, S.K.; McGavern, D.B. CD8+ T Cells Induce Fatal Brainstem Pathology during Cerebral Malaria via Luminal Antigen-Specific Engagement of Brain Vasculature. PLoS Pathog. 2016, 12, e1006022. [Google Scholar] [CrossRef] [PubMed]
- Niewold, P.; Cohen, A.; van Vreden, C.; Getts, D.R.; Grau, G.E.; King, N.J.C. Experimental Severe Malaria Is Resolved by Targeting Newly-Identified Monocyte Subsets Using Immune-Modifying Particles Combined with Artesunate. Commun. Biol. 2018, 1, 227. [Google Scholar] [CrossRef] [PubMed]
- Marquet, S.; Conte, I.; Poudiougou, B.; Argiro, L.; Cabantous, S.; Dessein, H.; Burté, F.; Oumar, A.A.; Brown, B.J.; Traore, A.; et al. The IL17F and IL17RA Genetic Variants Increase Risk of Cerebral Malaria in Two African Populations. Infect. Immun. 2016, 84, 590–597. [Google Scholar] [CrossRef]
- Raballah, E.; Kempaiah, P.; Karim, Z.; Orinda, G.O.; Otieno, M.F.; Perkins, D.J.; Ong’echa, J.M. CD4 T-Cell Expression of IFN-γ and IL-17 in Pediatric Malarial Anemia. PLoS ONE 2017, 12, e0175864. [Google Scholar] [CrossRef]
- Villegas-Mendez, A.; Greig, R.; Shaw, T.N.; de Souza, J.B.; Gwyer Findlay, E.; Stumhofer, J.S.; Hafalla, J.C.R.; Blount, D.G.; Hunter, C.A.; Riley, E.M.; et al. IFN-γ–Producing CD4+ T Cells Promote Experimental Cerebral Malaria by Modulating CD8+ T Cell Accumulation within the Brain. J. Immunol. 2012, 189, 968–979. [Google Scholar] [CrossRef]
- Monteiro, S.; Ferreira, F.M.; Pinto, V.; Roque, S.; Morais, M.; de Sá-Calçada, D.; Mota, C.; Correia-Neves, M.; Cerqueira, J.J. Absence of IFNγ Promotes Hippocampal Plasticity and Enhances Cognitive Performance. Transl. Psychiatry 2016, 6, e707. [Google Scholar] [CrossRef] [PubMed]
- Aitken, E.H.; Negri, E.M.; Barboza, R.; Lima, M.R.; Álvarez, J.M.; Marinho, C.R.; Caldini, E.G.; Epiphanio, S. Ultrastructure of the Lung in a Murine Model of Malaria-Associated Acute Lung Injury/Acute Respiratory Distress Syndrome. Malar. J. 2014, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care 2020, 9, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gu, Y.; Tu, Q.; Wang, K.; Gu, X.; Ren, T. Blockade of Interleukin-17 Restrains the Development of Acute Lung Injury. Scand. J. Immunol. 2016, 83, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Xiaoyu, Z.; Zhixin, S.; Di, Q.; Xinyu, D.; Jing, X.; Jing, H.; Wang, D.; Xi, Z.; Chunrong, Z.; et al. Rapamycin Attenuates Acute Lung Injury Induced by LPS through Inhibition of Th17 Cell Proliferation in Mice. Sci. Rep. 2016, 6, 20156. [Google Scholar] [CrossRef] [PubMed]
- Murdock, B.J.; Huffnagle, G.B.; Olszewski, M.A.; Osterholzer, J.J. Interleukin-17A Enhances Host Defense against Cryptococcal Lung Infection through Effects Mediated by Leukocyte Recruitment, Activation, and Gamma Interferon Production. Infect. Immun. 2014, 82, 937–948. [Google Scholar] [CrossRef]
- Murakami, T.; Hatano, S.; Yamada, H.; Iwakura, Y.; Yoshikai, Y. Two Types of Interleukin 17A–Producing Γδ T Cells in Protection Against Pulmonary Infection With Klebsiella Pneumoniae. J. Infect. Dis. 2016, 214, 1752–1761. [Google Scholar] [CrossRef]
- Wang, H.; Li, S.; Cui, Z.; Qin, T.; Shi, H.; Ma, J.; Li, L.; Yu, G.; Jiang, T.; Li, C. Analysis of Spleen Histopathology, Splenocyte Composition and Haematological Parameters in Four Strains of Mice Infected with Plasmodium Berghei K173. Malar. J. 2021, 20, 249. [Google Scholar] [CrossRef]
- Bronte, V.; Pittet, M.J. The Spleen in Local and Systemic Regulation of Immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef]
- Carroll, R.W.; Wainwright, M.S.; Kim, K.-Y.; Kidambi, T.; Gómez, N.D.; Taylor, T.; Haldar, K. A Rapid Murine Coma and Behavior Scale for Quantitative Assessment of Murine Cerebral Malaria. PLoS ONE 2010, 5, e13124. [Google Scholar] [CrossRef]
- Russo, R.C.; Savino, B.; Mirolo, M.; Buracchi, C.; Germano, G.; Anselmo, A.; Zammataro, L.; Pasqualini, F.; Mantovani, A.; Locati, M.; et al. The Atypical Chemokine Receptor ACKR2 Drives Pulmonary Fibrosis by Tuning Influx of CCR2 + and CCR5 + IFNγ-Producing γδT Cells in Mice. Am. J. Physiol. -Lung Cell. Mol. Physiol. 2018, 314, L1010–L1025. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Santos, F.; Leal-Silva, T.; de Lima Silva Padrão, L.; Ruas, A.C.L.; Nogueira, D.S.; Kraemer, L.; Oliveira, F.M.S.; Caliari, M.V.; Russo, R.C.; Fujiwara, R.T.; et al. Concomitant Experimental Coinfection by Plasmodium Berghei NK65-NY and Ascaris Suum Downregulates the Ascaris-Specific Immune Response and Potentiates Ascaris-Associated Lung Pathology. Malar. J. 2021, 20, 296. [Google Scholar] [CrossRef] [PubMed]
- Resende, N.M.; Gazzinelli-Guimarães, P.H.; Barbosa, F.S.; Oliveira, L.M.; Nogueira, D.S.; Gazzinelli-Guimarães, A.C.; Gonçalves, M.T.P.; Amorim, C.C.O.; Oliveira, F.M.S.; Caliari, M.V.; et al. New Insights into the Immunopathology of Early Toxocara Canis Infection in Mice. Parasites Vectors 2015, 8, 354. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaio, P.; Cramer, A.; de Melo Oliveira, N.F.; Porto, S.; Kramer, L.; Nonato Rabelo, R.A.; Pereira, R.d.D.; de Oliveira Santos, L.L.; Nascimento Barbosa, C.L.; Silva Oliveira, F.M.; et al. N-(coumarin-3-yl)cinnamamide Promotes Immunomodulatory, Neuroprotective, and Lung Function-Preserving Effects during Severe Malaria. Pharmaceuticals 2024, 17, 46. https://doi.org/10.3390/ph17010046
Gaio P, Cramer A, de Melo Oliveira NF, Porto S, Kramer L, Nonato Rabelo RA, Pereira RdD, de Oliveira Santos LL, Nascimento Barbosa CL, Silva Oliveira FM, et al. N-(coumarin-3-yl)cinnamamide Promotes Immunomodulatory, Neuroprotective, and Lung Function-Preserving Effects during Severe Malaria. Pharmaceuticals. 2024; 17(1):46. https://doi.org/10.3390/ph17010046
Chicago/Turabian StyleGaio, Paulo, Allysson Cramer, Natália Fernanda de Melo Oliveira, Samuel Porto, Lucas Kramer, Rayane Aparecida Nonato Rabelo, Rafaela das Dores Pereira, Laura Lis de Oliveira Santos, César Luís Nascimento Barbosa, Fabrício Marcus Silva Oliveira, and et al. 2024. "N-(coumarin-3-yl)cinnamamide Promotes Immunomodulatory, Neuroprotective, and Lung Function-Preserving Effects during Severe Malaria" Pharmaceuticals 17, no. 1: 46. https://doi.org/10.3390/ph17010046
APA StyleGaio, P., Cramer, A., de Melo Oliveira, N. F., Porto, S., Kramer, L., Nonato Rabelo, R. A., Pereira, R. d. D., de Oliveira Santos, L. L., Nascimento Barbosa, C. L., Silva Oliveira, F. M., Martins Teixeira, M., Castro Russo, R., Matos, M. J., & Simão Machado, F. (2024). N-(coumarin-3-yl)cinnamamide Promotes Immunomodulatory, Neuroprotective, and Lung Function-Preserving Effects during Severe Malaria. Pharmaceuticals, 17(1), 46. https://doi.org/10.3390/ph17010046