1. Introduction
Atopic dermatitis (AD) is a chronic inflammatory skin disease accompanied by severe itching and it is known worldwide as a disease in both children and adults that is caused by immune imbalance and skin barrier dysfunction. [
1,
2]. AD is a representative T-cell immune-mediated disease and is characterized by several factors, such as skin redness, dryness, itching, keratinocyte proliferation, immune cell infiltration, and lymph node T-cell expansion. In particular, Th1/Th2 immune imbalance caused by Th2-mediated immune response causes AD, along with damage to the skin barrier function [
3]. Immune response by immunologically abnormal Th2 cells is induced by high antibody production, and plays a role in activating B cell proliferation and class conversion, thereby causing cytokines interleukin (IL)-25, IL-33 and thymic stromal lymphopoietin (TSLP) to increase in keratinocytes of AD patients [
4,
5,
6]. Cytokines IL-25, IL-33, and TSLP play an important key role in the pro-inflammatory milieu of AD by stimulating Th2 cells directly or indirectly through the stimulation of multiple immune cells, including dendritic cells, mast cells, and eosinophils, and immunomodulatory disorders and contribute to barrier dysfunction [
7,
8]. These processes can also activate the cytokines IL-4, IL-5, IL-13 and IL-31, which are known to play a major role in skin infiltration in AD skin lesions [
9,
10,
11]. Currently, the most widely used drugs for the treatment of AD include corticosteroids, antihistamines, and immunosuppressive drugs, which are used by more than 60% of patients [
12,
13]. However, the use of these drugs to relieve symptoms of AD leads to various tolerance and side effects to the drug, including glaucoma, impaired wound healing, and growth retardation [
14]. Therefore, in order to reduce these problems, therapeutic agents that can replace the existing drugs are being developed to obtain safer and more effective therapeutics, and interest in the physiological activity of natural extracts derived from safer plants has started to increase recently [
15,
16].
Callicarpa dichotoma (Verbenaceae) is distributed evenly across Korea and central Japan, and it is known that there are about 40 species of plants that belong to
Callicarpa worldwide [
17]. In oriental traditional medicine, the plant of the genus
Callicarpa has been used for bruises, habitual pain, gastric and postpartum hemorrhage, and the leaves in particular have been used to treat inflammation and gastric ulcers [
18,
19]. The leaves and twigs of
C. dichotoma contain various types of phenylethanoid glycosides, essential oils, diterpenes, and flavonoids [
20]. In a previous study, the compound acteoside, isolated from
C. dichotoma, reduced neurotoxicity caused by glutamate and was reported to exhibit antioxidant anti-inflammatory and anti-hepatotoxic activity [
20]. Research on the ingredients and activities
of C. dichotoma is still incomplete. In addition, despite the anti-inflammatory effect of
C. dichotoma, its application to various inflammatory diseases has not been attempted. Therefore, in this study, the activity of
C. dichotoma was evaluated on an in vivo experimental animal model of mite-induced atopic dermatitis, and the effect of
C. dichotoma on the activity of T cells and keratinocytes, known as the main cause of atopic dermatitis, was investigated.
3. Discussion
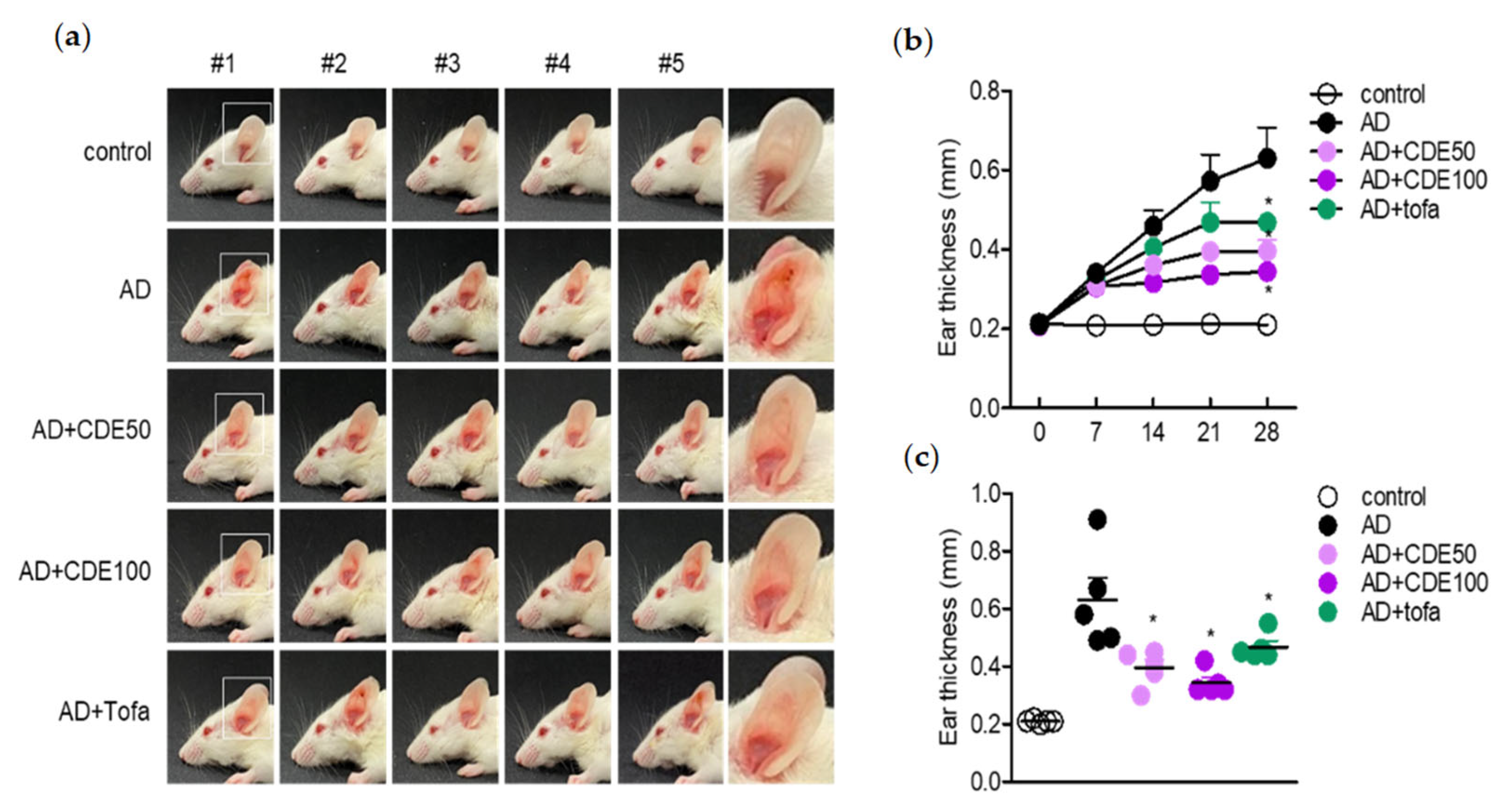
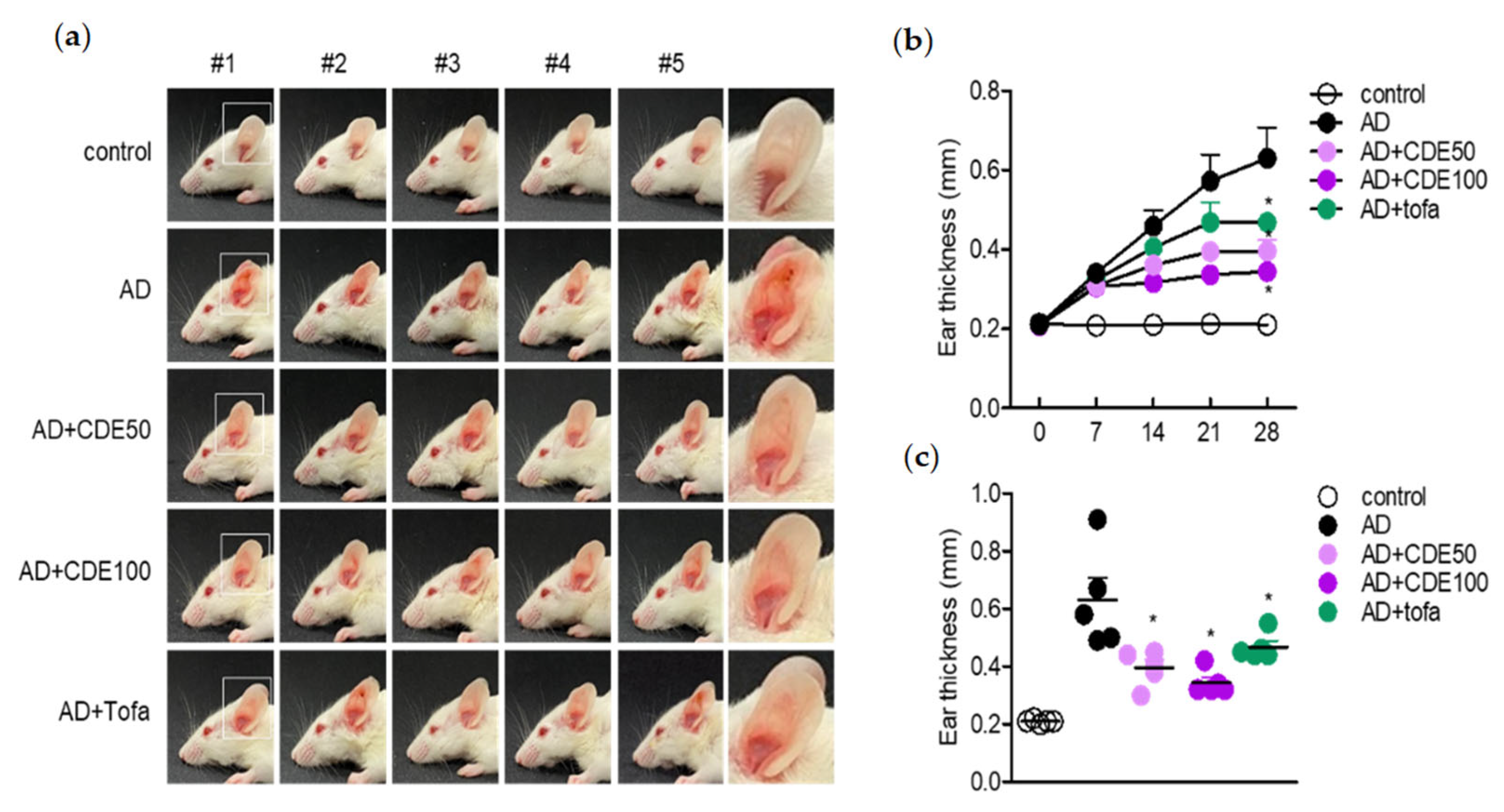
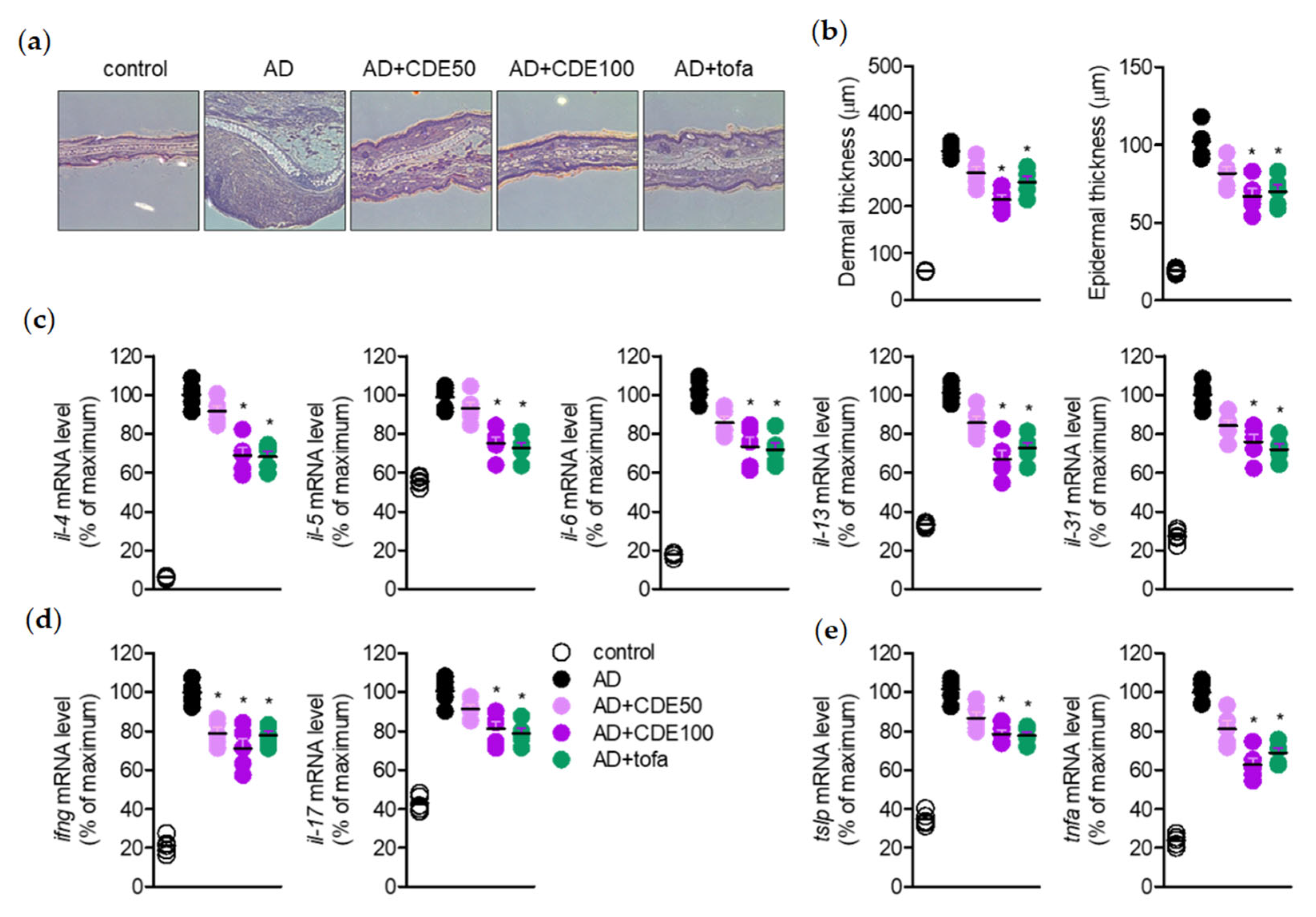
The present study explored the mitigating effect of CDE on the symptoms and pathogenesis of AD by modulating T cells and keratinocytes. As a barrier against external environmental stimuli, the keratin of the skin contributes to ILC2 and Th2 cell activation and hypersensitivity reaction and, eventually, along with the activation of T cells, it induces the characteristics of AD [
22]. Additionally, the immune response regulation through keratin protection from such damage to the skin barrier is reported as a new therapeutic strategy for AD [
23]. Therefore, in this study, the effect of CDE on AD in this novel treatment strategy was demonstrated in vitro and in vivo. In this study, CDE effectively restored the thickened ear thickness of AD-induced mice, and also stabilized the thickness of the epidermis and dermis of the ear tissue. These results suggest that CDE alleviates the typical symptoms of AD and exhibits therapeutic effects. In this process, modulation of the immune response through keratin protection is known to be an important strategy for AD treatment [
24]; therefore, the role of CDE in keratinocyte protection was also explored in this study.
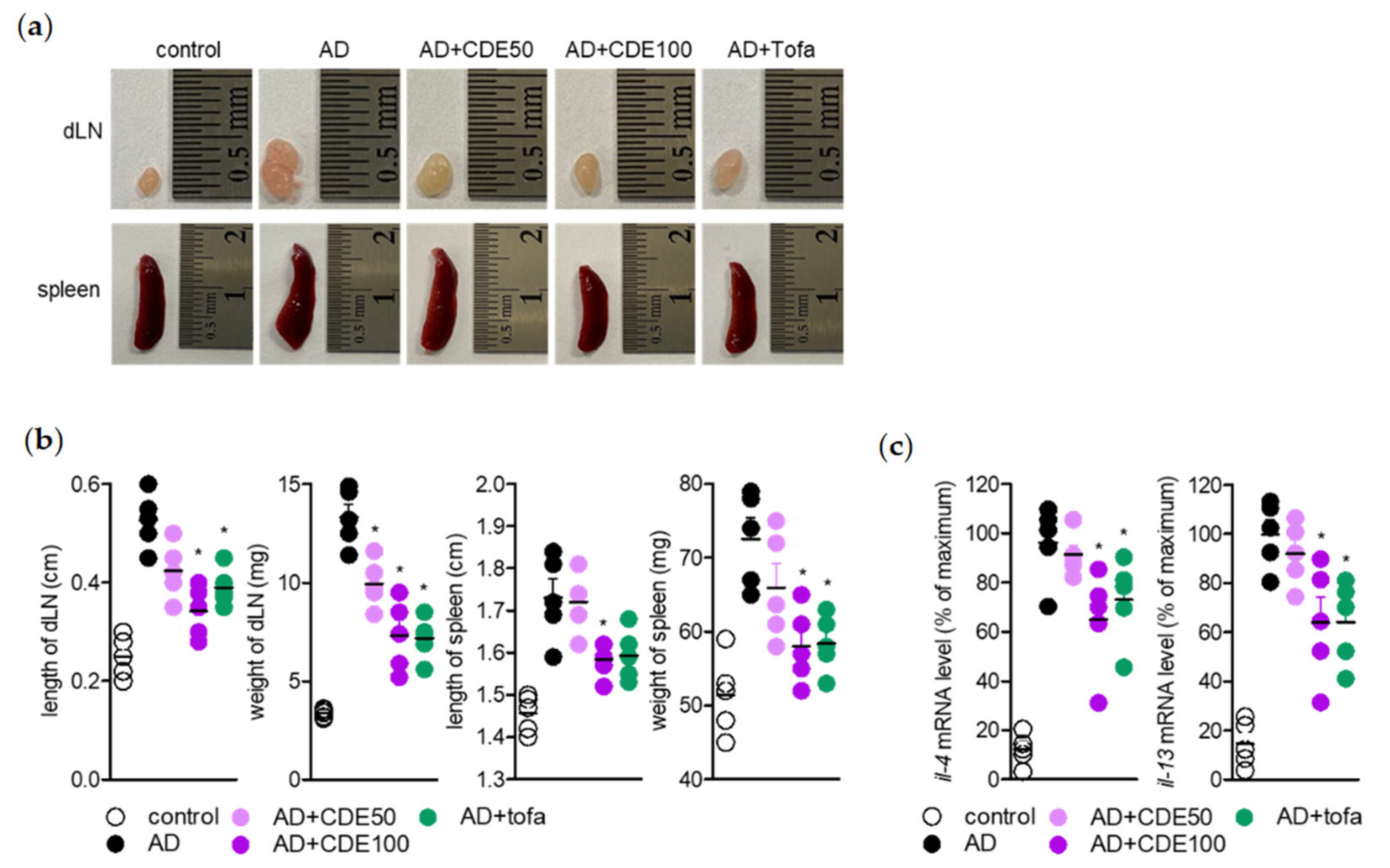
AD is a representative Th2 cytokine gene expression-induced immune regulation disorder and various pro-inflammatory cytokines, in particular IL-4 and IL-13, have been studied as major therapeutic targets for AD [
25,
26]. In order to obtain more convincing evidence of the AD symptom-alleviation effect of CDE, the analysis of these main therapeutic target cytokines for AD was conducted from ear tissues of AD-induced mice. In addition, enlarged ear thickness in the AD-induced mouse model is a known symptom of AD [
27], and the effect of CDE on these symptoms was evaluated together. CDE effectively down-regulated the gene expression levels of major pro-inflammatory cytokines, which are therapeutic targets for AD, and the ear thickness of the thickened mice was alleviated. Therefore, it could be concluded that CDE not only relieves typical symptoms of AD, but also exerts a therapeutic effect by directly down-regulating the therapeutic target cytokines.
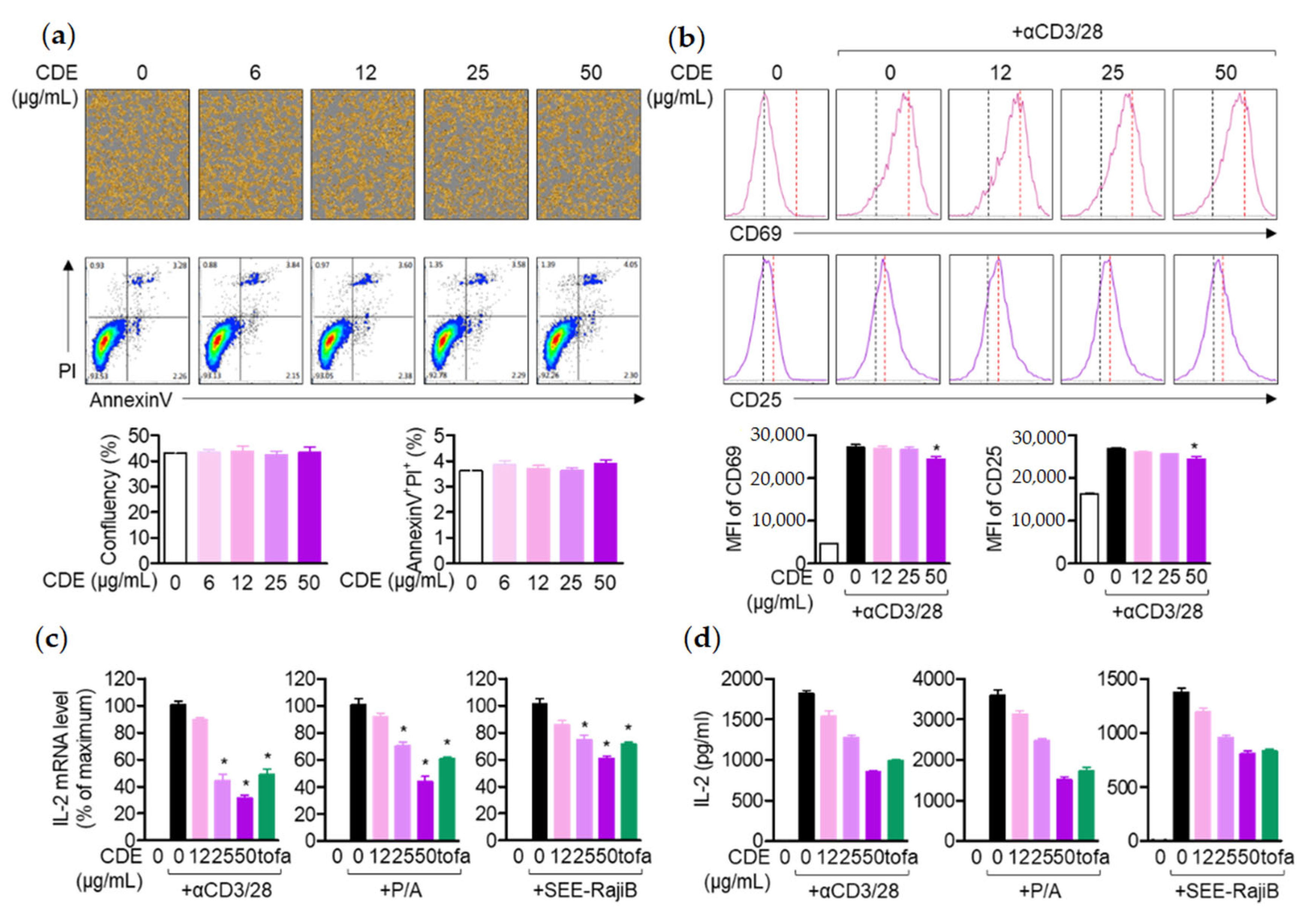
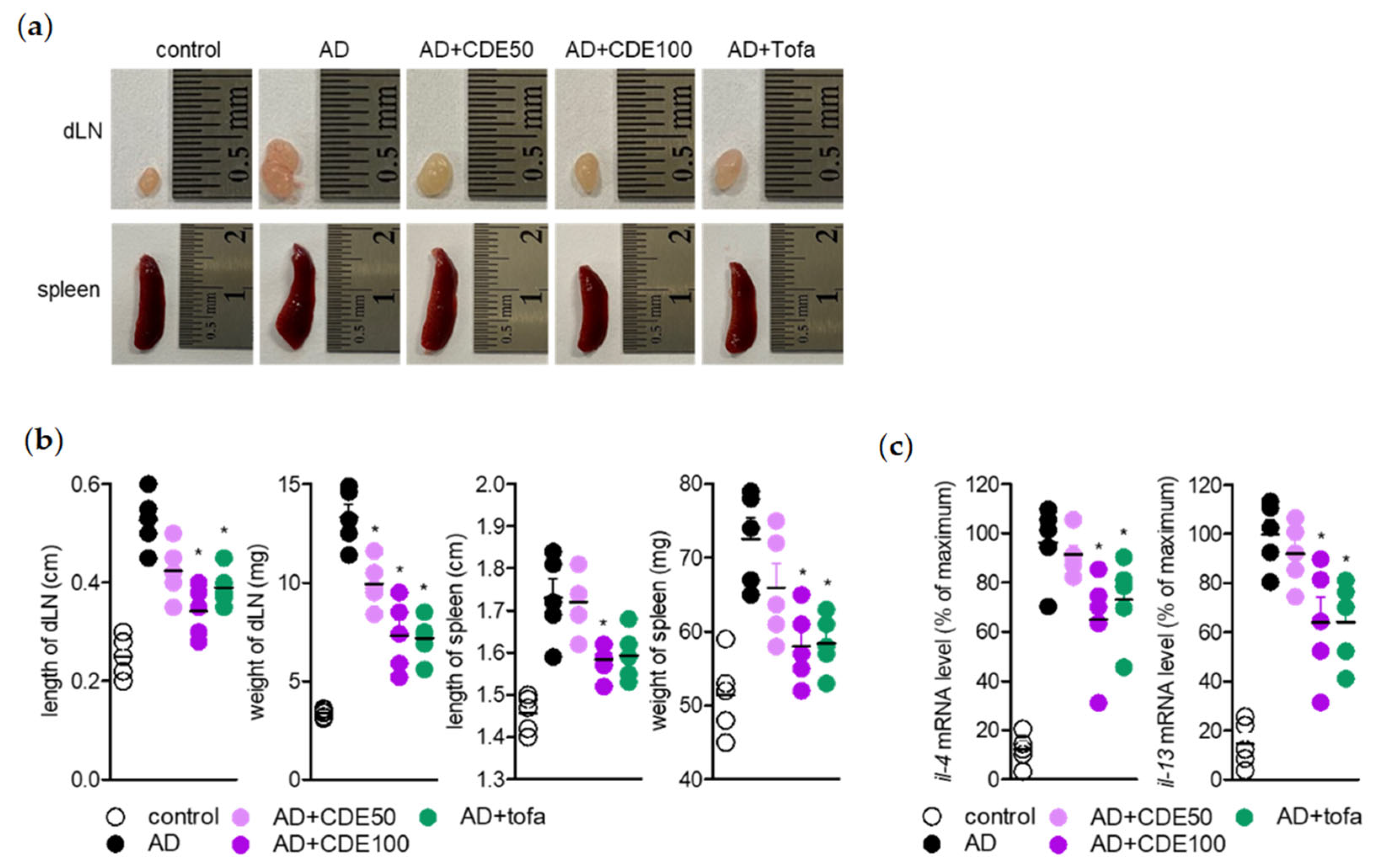
Regarding the AD alleviation effect of CDE, as mentioned above, the effect of CDE on lymph nodes and spleen, which are representative immune organs, was evaluated to determine whether it is due to the effect of regulating the immune response through keratin protection. The lymph node, a representative primary immune organ, is known to play an important key role in the maturation of B cells and T cells, and the spleen, a secondary immune organ, is also known as another important institution [
28,
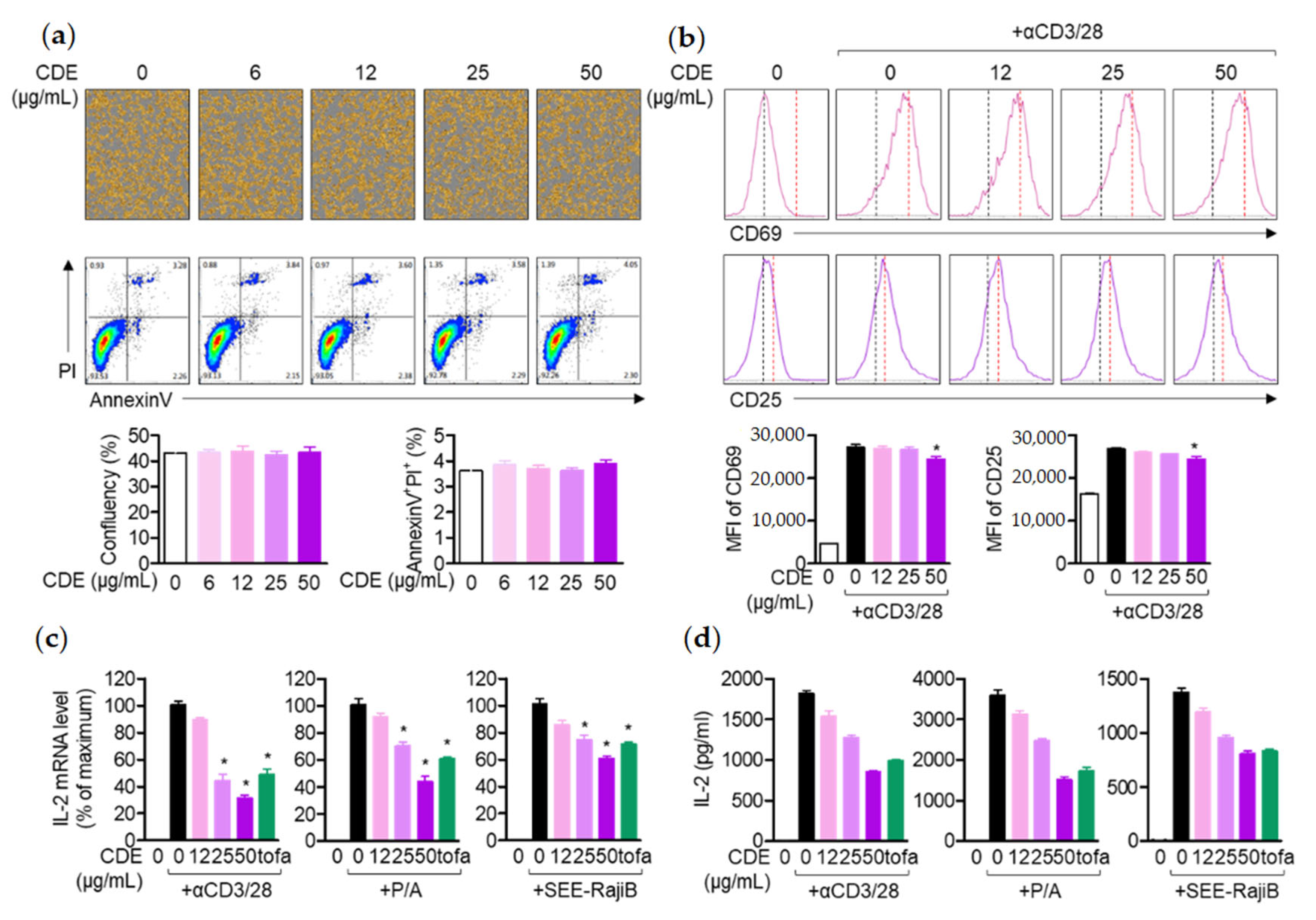
29]. According to the study results, CDE effectively restored the size of lymph nodes and spleen in mice with enlarged organs due to AD induction, which suggests that CDE can exert an AD-alleviating effect by modulating the systemic immune response. Therefore, as mentioned above, T cells activated in AD are known to induce the production of Th2 cytokines. This study assessed the direct effect of AD treatment of CDE on T cells and keratinocyte activities. As mentioned above, activated T cells in AD are known to induce the production of Th2 cytokines, and CDE effectively downregulated the expression of these cytokines. Therefore, in order to evaluate the direct effect of the AD treatment effect of CDE on the activity of T cells and keratinocytes, the expression levels of CD69 and CD25 were evaluated in Jurkat T cells stimulated with CD3/28. The
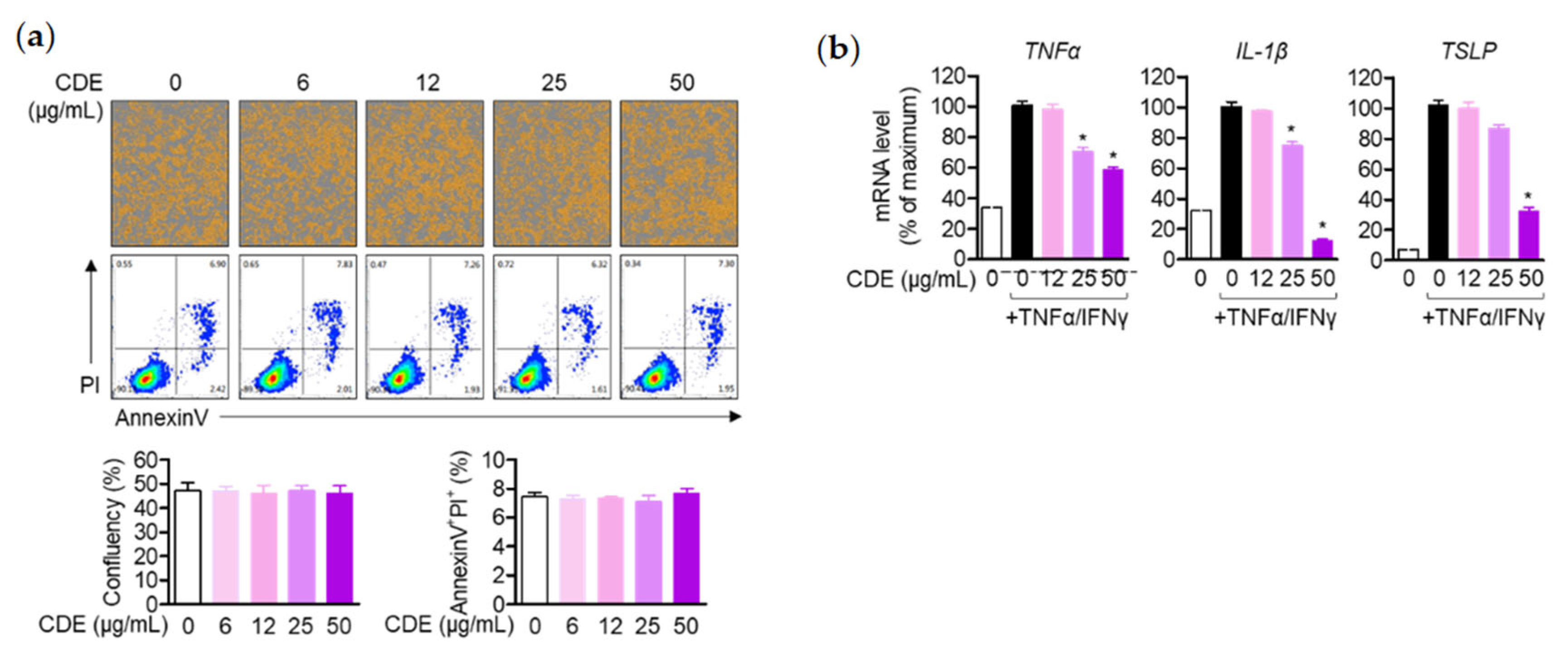
il2 gene expression level, a major cytokine of T cell activity, was evaluated. The data revealed that CDE did not show any effect on the toxicity of Jurkat T cells or keratinocytes in the indicated concentrations. Additionally, not only the expression levels of CD69 and CD25 of Jurkat T cells, but also the gene expression level of
il2 were down-regulated. Similarly, the gene expression levels of pro-inflammatory cytokines TNFα, IL-1β, and TSLP were suppressed in keratinocytes.
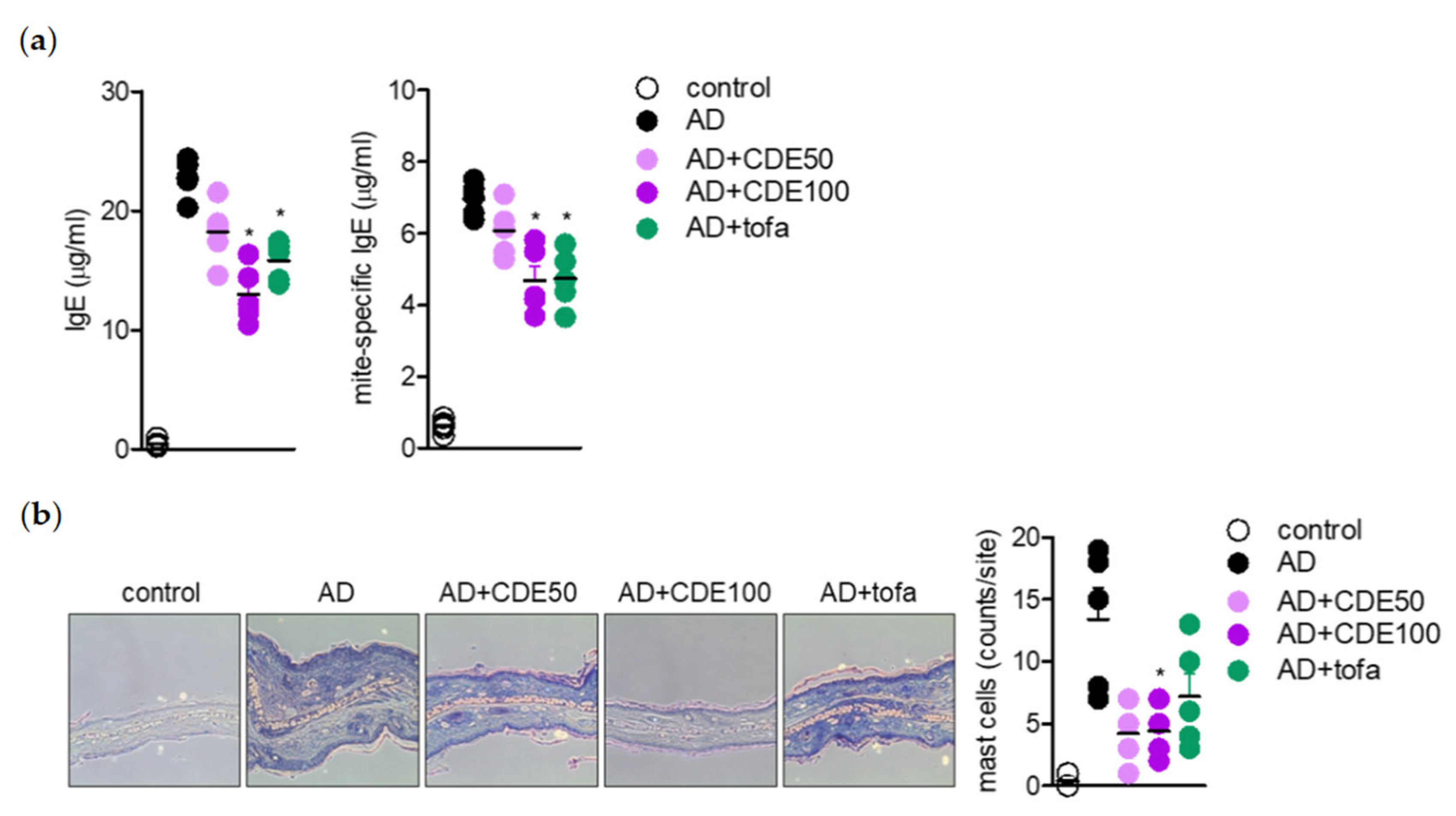
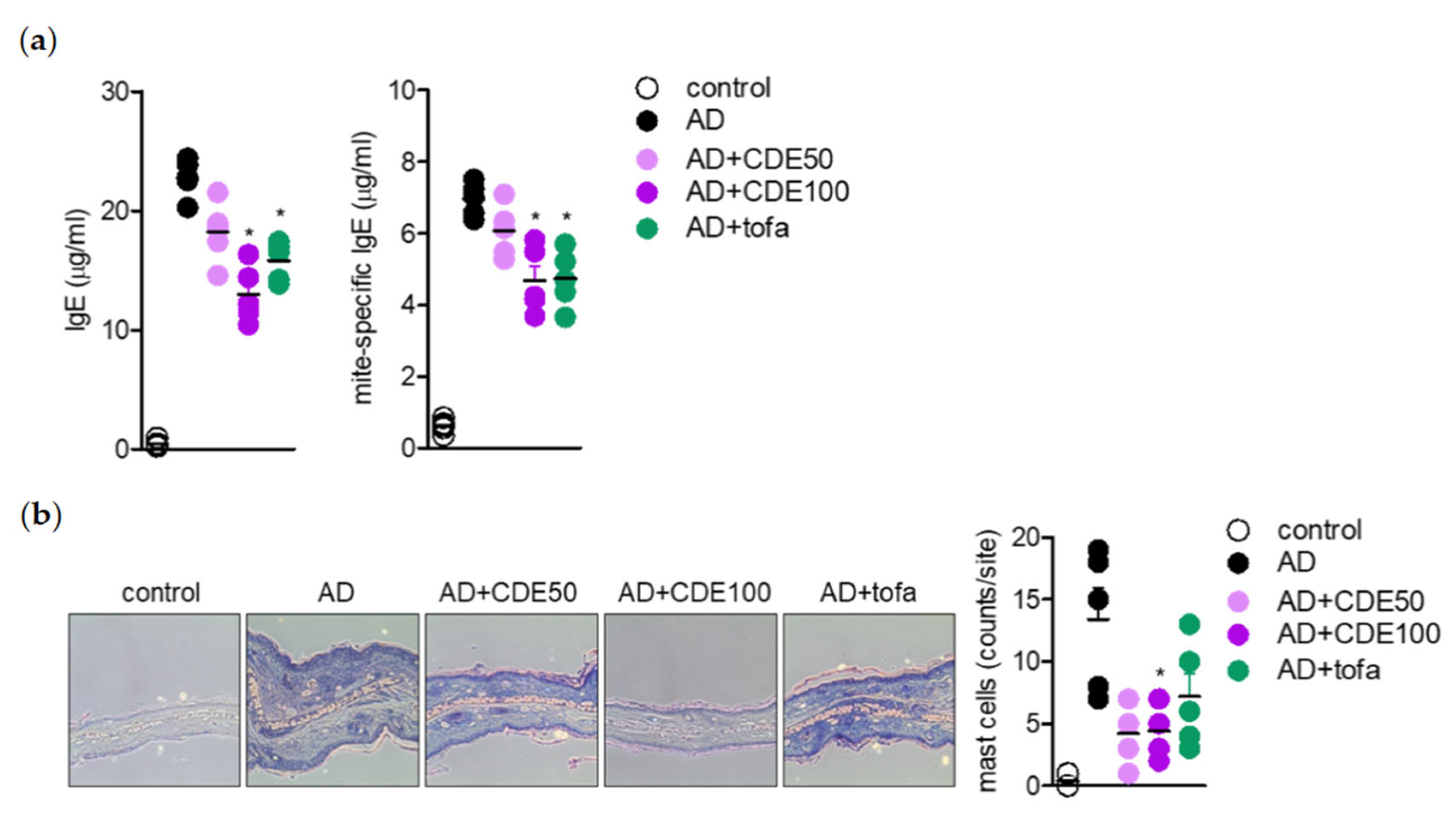
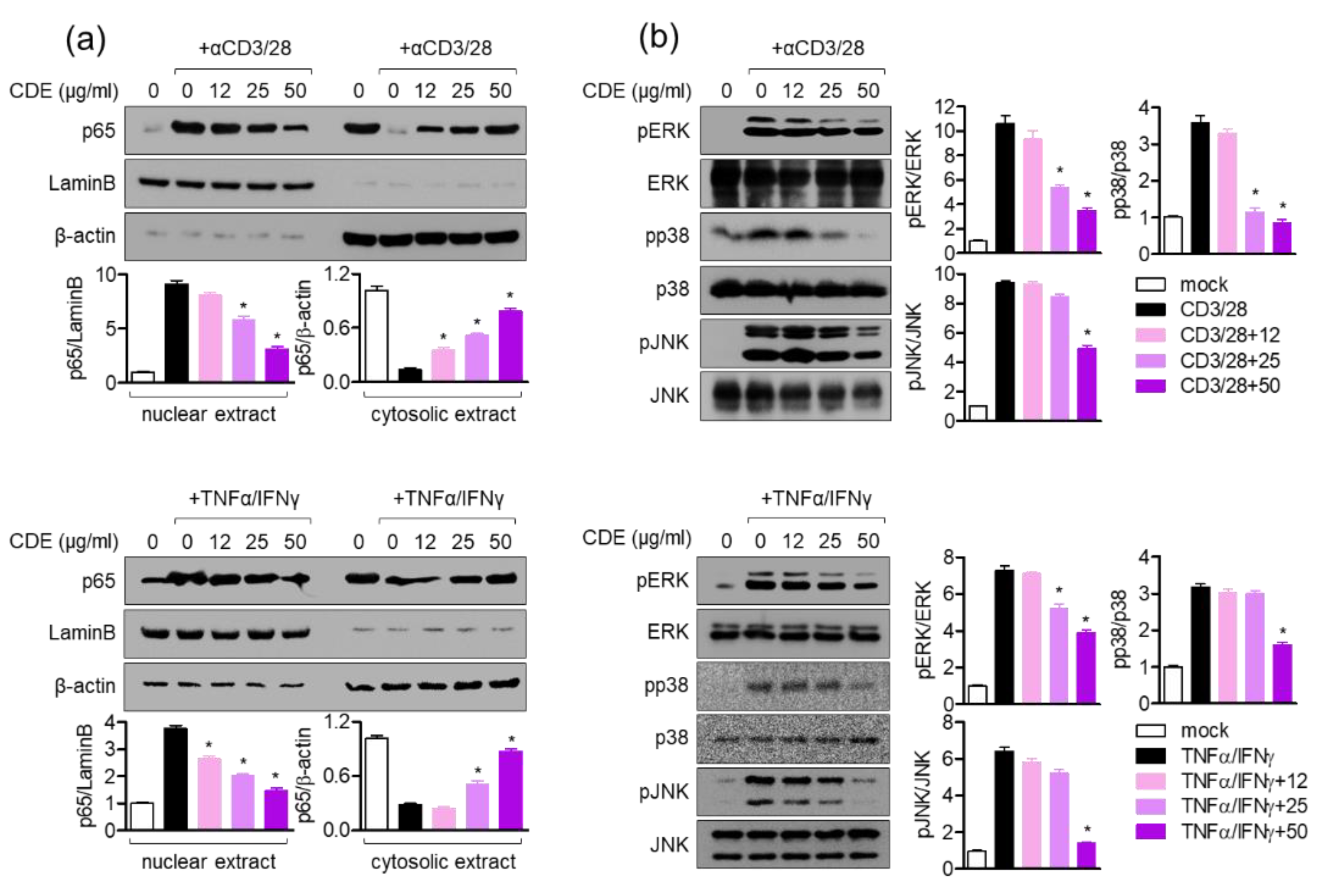
IL-4 is known to provide positive feedback to Th 2 cells and induce B isoform cells to differentiate into IgE antibody-producing cells, which is a major mediator and activator of allergic reactions, by inducing mitogen-activated protein kinase (MAPK) activation through the nuclear factor kappa-light-chain-enhancer of the activated B cells (NFκB) pathway to increase intracellular calcium and simultaneously induce histamine secretion [
30,
31]. In this study, CDE downregulated the
il4 gene expression level in AD-induced in vivo mice model; hence, the effect of CDE on the NFκB and MAPK pathways was evaluated. CDE inhibited the nuclear translocation of NFκB p65 in Jurkat T cells and keratinocytes stimulated with CD3/28 and TNFα/IFNγ, respectively, and downregulated the expression level of phosphorylated MAPK protein.
4. Materials and Methods
4.1. Plant Material
C. dichotoma was obtained at the Daegu Yangnyeong Herbal Medicine Market in 2021, and a C. dichotoma voucher specimen was deposited at Chungnam National University, College of Pharmacy.
4.2. Extract of C. dichotoma Leaf
The dried leaves of C. dichotoma (387.6 g) were manually separated and ground, and the dried leaves were extracted with EtOH (1 L) after 1 day at room temperature, and extracted twice for 2 h at 80 °C. Thereafter, the alcohol extract was concentrated in a rotary vacuum to obtain a residue (94.6 g).
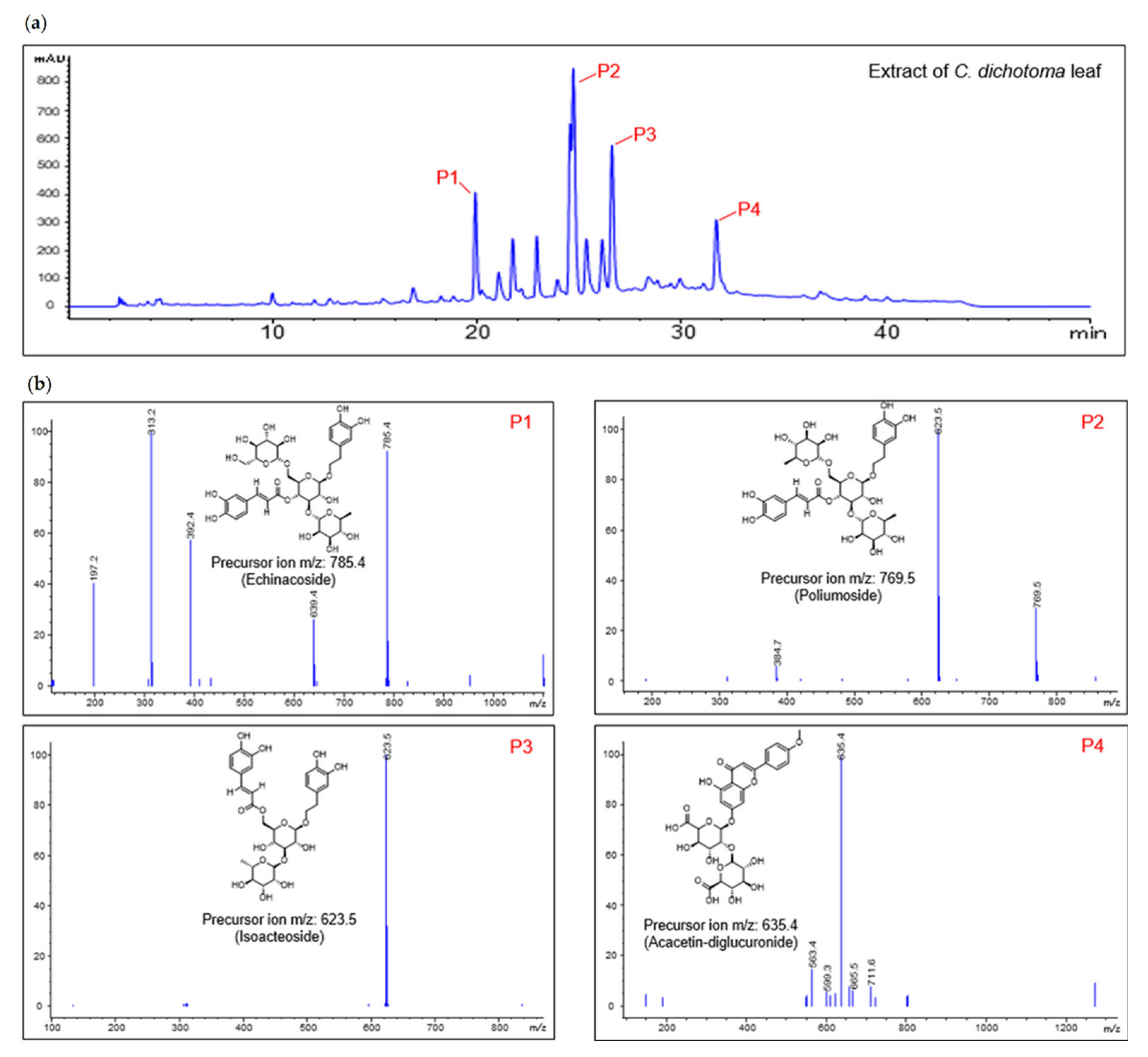
4.3. Condition of Liquid Chromatography–Mass Spectrometry Analysis
To analyze the extract of C. dichotoma leaf, we used the single quadrupole negative ion mode on an eclipse plus C18 (5 μm × 4.6 mm × 250 mm; Agilent, Santa Clara, CA, USA) column and a high-performance liquid chromatography (Agilent model 1260 series, Santa Clara, CA, USA)–mass spectrometry (Agilent 6120 series, Santa Clara, CA, USA) system.
4.4. Reagents and Antibodies
Dinitrochlorobenzene (DNCB), and RIPA buffer were purchased from Sigma Chemical Co. (St. Louis, MO, USA). House dust mite (Dermatophagoides farina) extract was obtained from Greer (Lenoir, NC, USA). Mouse IgE ELISA kit was purchased from R&D Systems (Minneapolis, MN, USA). Antibodies for p65, p38, ERK, JNK, lamin B and β-actin were obtained from Cell Signaling Technology (Danvers, MA, USA). Antibodies for phosphorylated ERK, p38 and JNK were also purchased from Cell Signaling Technology (Danvers, MA, USA).
4.5. Animals
Six to eight-week-old female BALB/c mice were obtained from Samtako and housed in specific pathogen-free (SPF) conditions. All experiments were approved by the Animal Care and Use Committee of the College of Pharmacy, Keimyung University (approval number: KM2020-007, approved on July 2nd 2020).
4.6. Induction of Atopic Dermatitis
AD was induced by repeatedly applying mite extract and DNCB to the ears of mice. The control mice group were treated with DNCB/mite extract alone (AD), the experimental mice group were treated with DNCB/mite extract concurrently (mite extract and CDE (AD + CDE)), and for the positive control mice, DNCB/mite extract and tofacitinib (AD + tofa) were administered simultaneously. To induce AD disease, both ears were wrapped with surgical tape 5 times (Seo-il Chemistry, Hwasung, Korea). After stripping of the tape, 20 μL of DNCB (1%) was topically administered to each ear. After 4 days, the ears were painted with 20 μL of tick extract (10 mg/mL). Tick extract/DNCB treatment was repeated weekly for 4 weeks. Treatment started one day after the second DNCB application. It was repeated daily for 4 days thereafter. After a 2-day break, this 5-day on and 2-day off CDE dosing protocol was repeated for 4 weeks. Ear thickness was assessed 24 h after application of DNCB or mite extract using a dial thickness meter (Kori Seiki MFG Co., Tokyo, Japan). Animals were sacrificed 28 days after induction.
4.7. H&E Staining
H&E staining was performed to evaluate the histopathological effect of CDE on AD-induced mice. Ears removed from the experimental animals were fixed in 10% paraformaldehyde and embedded in paraffin. Paraffin-embedded ears were cut to a thickness of 5 μm, deparaffinized, and stained with hematoxylin and eosin (H&E). Dermal and epidermal thickness were measured on H&E-stained slides.
4.8. Cell Culture
Jurkat T cells (KCLB number: 40152) were purchased from the Korean Cell Line Bank (Seoul, Korea) and Raji B (ATCC cat#. CCL-86) cells were purchased from ATCC (Manassas, VA, USA). Cells were cultured in RPMI medium (Welgene, Gyeongsan, Korea), supplemented with penicillin G (100 units/mL), streptomycin (100 μg/mL), 10% fetal bovine serum (FBS), and L-glutamine (2 mM). In addition, the HaCaT cell line was kindly donated by Prof. Eun-Kyung Kim (Department of Food Science and Nutrition, Dong-A University, Busan, Korea) and cultured in DMEM medium (Welgene, Gyeongsan-si, Korea), supplemented with 10% fetal bovine serum (FBS), penicillin G (100 units/mL), streptomycin (100 μg/mL), and L-glutamine (2 mM). The passage of HaCaT cells was maintained at three to eight cells for experiments. Cells were grown at 37 °C in a humidified incubator that contained 5% CO2 and 95% air.
4.9. Analysis of mRNA Level Using Real-Time Quantitative PCR
To evaluate the mRNA level of each gene, cells were lysed in TRIZOL reagent. Then, reverse transcription of RNA was performed using the RT PreMix kit (Enzynomics, Korea). The primers used in this study were as follows: human IL-2, 5′-CAC GTC TTG CAC TTG TCA C-3′ and 5′-CCT TCT TGG GCA TGT AAA ACT-3′; human tnfa, 5′-ACC TCA TCT ACT CCC AGG TC-3′ and 5′-AAG ACC CCT CCC AGA TAG AT-3′; human IL-1b, 5′-GGA TAT GGA GCA ACA AGT GG-3′ and 5′-ATG TAC CAG TTG GGG AAC TG-3′; human tslp, 5′-TAG CAA TCG GCC ACA TTG CCT-3′ and 5′-GAA GCG ACG CCA CAA TCC TTG-3′; human GAPDH, 5′-CGG AGT CAA CGG ATT TGG TCG TAT-3′ and 5′-AGC CTT CTC CAT GGT GGT GAA GAC-3′; mouse IL-4, 5′-ACA GGA GAA GGG ACG CCA T-3′ and 5′-GAA GCC GTA CAG ACG AGC TCA-3′; mouse Il-5, 5′-GAA GTG TGG CGA GGA GAG AC-3′ and 5′-GCA CAG TTT TGT GGG GTT TT-3′; mouse IL-6, 5′-CCG GAG AGG AGA CTT CAC AG–3′ and 5′-GGA AAT TGG GGT AGG AAG GA–3′; mouse IL-13, 5′-GCA ACA TCA ACA GGA CCA GA–3′ and 5′-GTC AGG GAA TCC AGG GCT AC–3′; mouse IL-31, 5′- TCG GTC ATC ATA GCA CAT CTG GAG–3′ and 5′- GCACAG TCC CTT TGG AGT TAA GTC –3′; mouse Ifng, 5′-TCA AGT GGC ATA GAT GTG GAA GAA-3′ and 5′-TGG CTC TGC AGG ATT TTC ATG-3′; mouse Il17, 5′-TCC CCT CTG TCA TCT GGG AAG-3′ and 5′-CTC GAC CCT GAA AGT GAA GG-3′; mouse TSLP, 5′-GCA ACA TCA ACA GGA CCA GA–3′ and 5′-GTC AGG GAA TCC AGG GCT AC–3′; mouse tnfa, 5′-AAG CCT GTA GCC CAC GTC GTA-3′ and 5′-GGC ACC ACT AGT TGG TTG TCT TTG-3′; mouse Gapdh, 5′–GCA CAG TCA AGG CCG AGA AT–3′ and 5′–GCC TTC TCC ATG GTG GTG AA–3′. PCR amplification was performed in a DNA Engine Opticon 1 continuous fluorescence detection system (MJ Research, Waltham, MA, USA), using SYBR Premix Ex Taq. It contained 1 μL of cDNA/control and gene-specific primers. Each PCR reaction was performed using the following conditions: 95 °C 30 s, 60 °C 30 s, 72 °C 30 s, and plate reading (detection of fluorescent product) was carried out for 40 cycles, followed by 7 min of extension at 72 °C. Melting curve analysis was performed to characterize the dsDNA product by slowly raising the temperature (0.1 °C/s) from 60 °C to 95 °C, with fluorescence data collected at 0.2 °C intervals. mRNA levels of genes were normalized to Gapdh. The gene expression was calculated using the following equation: gene expression = 2−ΔΔCT, where ΔΔCT = (CT target−CT gapdh).
4.10. Analysis of Cell Confluency
Jurkat cells (1 × 104 cells/well, 96 well) and keratinocytes (1 × 104 cells/well, 96 well) were treated with the indicated concentration (6–50 μg/mL) of CDE for 24 h, then the cells were automatically marked in yellow by the IncuCyte live cell imaging system (Sartorius, Göttingen, Germany).
4.11. AnnexinV/PI Apoptosis Assay
To evaluate the effects of CDE on apoptosis, the AnnexinV/PI apoptosis kit was used. For determination of apoptosis after treatment with CDE, the AnnexinV/PI apoptosis kit was used. The Jurkat cells (5 × 105/well, 12 well) and keratinocyte cells (5 × 105/well, 12 well) were treated with CDE at the indicated concentrations (6–50 μg/mL) for 24 h and then stained with AnnexinV and PI, according to the manufacturer’s instructions. After one day, flow cytometry was performed and gated using BD software.
4.12. T Cell Stimulation
In this study, three methods were used to stimulate T cells. In the first method, Jurkat T cells were replaced on plates coated with anti-CD3 antibody (20 μg/mL) and stimulated by treatment with anti-CD28 soluble antibody (7 μg/mL). In addition, for PMA/A23187 stimulation, Jurkat T cells were stimulated by treatment with 100 nM PMA and 1 μM A23187, and as the final method, Jurkat T cells were stimulated with the same number of previously pulsed SEE superantigens (1 μg/mL) for 1 h and were stimulated by co-culture with Raji B cells.
4.13. Western Blot Analysis
Western blot analysis was performed to detect the level of each target protein in the cells. Lysates were obtained by lysing the harvested cells in RIPA buffer for 30 min on ice and centrifugation at 14,000 rpm for 20 min at 4 °C. For nuclear isolation of NFκB p65, NE-PER nuclear and cytoplasmic extraction reagents (Thermo Fisher Science, Waltham, MA, USA) were used according to the manufacturer’s instructions. Lysates (30–40 μg) were loaded for separation on an 8–12% SDS-PAGE gel. Total proteins were transferred to PVDF membranes (Bio-Rad, Hercules, CA, USA) and the membranes were placed in 5% skim milk in TBS that contained 0.1% Tween 20 (TBS-T) for 1 h. After rinsing with TBST, the membranes were incubated with the indicated primary antibodies in 3% skim milk in TBST overnight at 4 °C. Excess primary antibody was discarded by washing the membrane 3 times with TBST. The membranes were then incubated with 0.1 μg/mL of peroxidase-labeled secondary antibody (for rabbits or mice) for 2 h. After three washes with TBST, the bands were treated with ECL Western blotting detection reagent (Thermo Fisher Scientific, Waltham, MA, USA), using an ImageQuant LAS 4000 (GE Healthcare, Chicago, IL, USA). All detected bands were quantified by ImageJ and normalized to the intensity of the loading control protein (β-actin). In the ‘mock’ group, the ratio between the experimental protein and the loading control protein was calculated to be 1×. All normalized proportions were expressed as fold changes compared to the ‘mock’ group.
4.14. Statistics
Mean values ± SEM were calculated from the data collected from the three independent experiments performed on separate days. For each mice experiment group, the mean ± SEM were analyzed from the data obtained from the experiments of 5 mice per group and displayed as bar or dot graphs. One-way ANOVA was used to determine the significance (p value) and Tukey’s post-hoc test was used after one-way ANOVA. * indicates that differences between the two indicated groups were considered significant at p < 0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}