Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19?

 , , , ,
, , , , 

Abstract
1. Introduction
2. Zn(II) as an RNA-Dependent RNA Polymerase Inhibitor
3. Anticoronaviral Properties of Chloroquine and Hydroxychloroquine
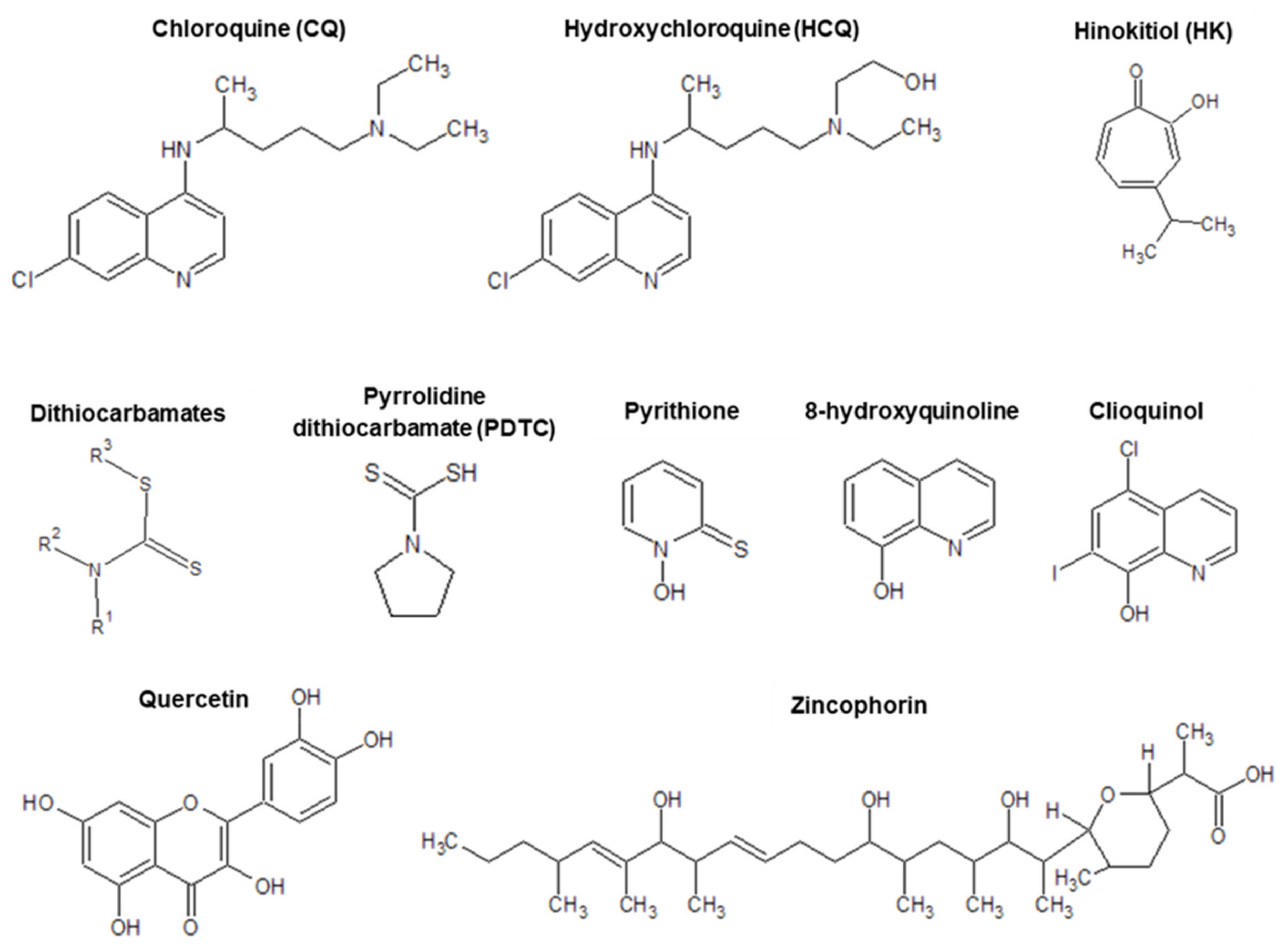
3.1. Zinc(II) Ionophores and Other Possible Modes of Action
3.2. Ongoing Clinical Trials
4. Antiviral Properties of Chloroquine and Hydroxychloroquine
5. Other Antiviral Zn(II) Ionophores
5.1. Pyrithione
5.2. Quercetin
5.3. Dithiocarbamates
6. Anticancer Properties of Zinc(II) Ionophores
6.1. Pyrithione
6.2. Clioquinol
6.3. Dithiocarbamates
7. Antibacterial Zn(II) Ionophores
7.1. Pyrithione
7.2. 8-Hydroxyquinoline
7.3. Zincophorin
7.4. Pyrrolidine Dithiocarbamate
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE2 | angiotensin-converting enzyme 2; |
| CCHFV | Crimean–Congo hemorrhagic fever virus; |
| CHIKV | Chikungunya virus; |
| DENV | Dengue virus; |
| DTC | dithiocarbamate; |
| EAV | equine arteritis virus; |
| EBOV | Ebola virus; |
| EC50 | effective statistically calculated concentration that induces a specific effect in 50% of experimental population under direct/specific conditions; |
| FCoV | feline coronavirus; |
| FIP | feline infectious peritonitis; |
| FIPV | feline infectious peritonitis virus; |
| FMDV | foot-and-mouth disease virus; |
| HA | hemagglutinin; |
| HAV | hepatitis A; |
| HBsAg | Hepatitis B surface antigen (HBsAg), |
| HBeAg | Hepatitis B antigen; |
| HBV | hepatitis B; |
| HCQ | hydroxychloroquine; |
| HCV | hepatitis C; |
| HIF-1 | hypoxia-inducible factor-1; |
| HIV | human immunodeficiency virus; |
| HK | hinokitol; |
| HSV | herpes simplex virus; |
| IC50 | inhibitory concentration: median value of inhibitor, where 50% of biological and biochemical function of organism inhibition is observed; |
| JEV | Japanese encephalitis virus; |
| NiRAN | nidovirus RdRp-associated nucleotidyltransferase; |
| NTP | nucleoside triphosphate; |
| PDTC | pyrrolidine dithiocarbamate; |
| PRRSV | porcine reproductive and respiratory syndrome virus; |
| PT | pyrithione; |
| RdRp | RNA-dependent RNA polymerase; |
| SARS-CoV | respiratory syndrome coronavirus; |
| SOD | superoxide dismutase-1; |
| UPS | ubiquitin-proteasome system; |
| ZIKV | Zika virus |
References
- Te Velthuis, A.J.W.; van den Worm, S.H.E.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn2+ Inhibits Coronavirus and Arterivirus RNA Polymerase Activity In Vitro and Zinc Ionophores Block the Replication of These Viruses in Cell Culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]
- Xue, J.; Moyer, A.; Peng, B.; Wu, J.; Hannafon, B.N.; Ding, W.-Q. Chloroquine Is a Zinc Ionophore. PLoS ONE 2014, 9, e109180. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Tian, Z.; Yang, X. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci. Trends 2020, 14, 72–73. [Google Scholar] [CrossRef]
- Gautret, P.; Lagier, J.-C.; Parola, P.; van Hoang, T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Li, C.; Chen, P.; Zhou, N.; Wang, L.; Li, J.; Jiang, H.; Wang, D.-W. Erratum to: Low dose of hydroxychloroquine reduces fatality of critically ill patients with COVID-19. Sci. China Life Sci. 2020. [Google Scholar] [CrossRef]
- Mahévas, M.; Tran, V.-T.; Roumier, M.; Chabrol, A.; Paule, R.; Guillaud, C.; Fois, E.; Lepeule, R.; Szwebel, T.-A.; Lescure, F.-X.; et al. Clinical efficacy of hydroxychloroquine in patients with covid-19 pneumonia who require oxygen: Observational comparative study using routine care data. BMJ 2020, 369, m1844. [Google Scholar] [CrossRef] [PubMed]
- Mehra, M.R.; Desai, S.S.; Ruschitzka, F.; Patel, A.N. RETRACTED: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020. [Google Scholar] [CrossRef]
- World Health Organization. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—25 May 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---25-may-2020 (accessed on 7 July 2020).
- The Lancet Editors. Expression of concern: Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020, 395, e102. [Google Scholar] [CrossRef]
- Mehra, M.R.; Ruschitzka, F.; Patel, A.N. Retraction—Hydroxychloroquine or chloroquine with or without a macrolide for treatment of COVID-19: A multinational registry analysis. Lancet 2020, 395, 1820. [Google Scholar] [CrossRef]
- Mehra, M.R.; Desai, S.S.; Kuy, S.; Henry, T.D.; Patel, A.N. Cardiovascular Disease, Drug Therapy, and Mortality in Covid-19. N. Engl. J. Med. 2020, 382, e102. [Google Scholar] [CrossRef]
- Scholz, M.; Derwand, R. Does Zinc Supplementation Enhance the Clinical Efficacy of Chloroquine/Hydroxychloroquine to Win Todays Battle Against COVID-19? Preprints 2020, 109815. [Google Scholar] [CrossRef]
- Scholz, M.; Derwand, R.; Zelenko, V. COVID-19 Outpatients–Early Risk–Stratified Treatment with Zinc Plus Low Dose Hydroxychloroquine and Azithromycin: A Retrospective Case Series Study. Preprints 2020, 2020070025. [Google Scholar] [CrossRef]
- Carlucci, P.; Ahuja, T.; Petrilli, C.M.; Rajagopalan, H.; Jones, S.; Rahimian, J. Hydroxychloroquine and azithromycin plus zinc vs. hydroxychloroquine and azithromycin alone: Outcomes in hospitalized COVID-19 patients. medRixv 2020. [Google Scholar] [CrossRef]
- Ahlquist, P.; Noueiry, A.O.; Lee, W.-M.; Kushner, D.B.; Dye, B.T. Host Factors in Positive-Strand RNA Virus Genome Replication. JVI 2003, 77, 8181–8186. [Google Scholar] [CrossRef]
- Miller, W.A.; Koev, G. Synthesis of Subgenomic RNAs by Positive-Strand RNA Viruses. Virology 2000, 273, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Leveque, V.; Ma, H.; Johnson, K.A.; Klumpp, K. Assembly, purification, and pre-steady-state kinetic analysis of active RNA-dependent RNA polymerase elongation complex. J. Biol. Chem. 2012, 287, 10674–10683. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef]
- Stadler, K.; Masignani, V.; Eickmann, M.; Becker, S.; Abrignani, S.; Klenk, H.-D.; Rappuoli, R. SARS--beginning to understand a new virus. Nat. Rev. Microbiol. 2003, 1, 209–218. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest RNA virus genome. Virus Res. 2006, 117, 17–37. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat. Commun. 2019, 10, 2342. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, Y.; Weiss, S.; Arnold, E.; Sarafianos, S.G.; Ding, J. Molecular model of SARS coronavirus polymerase: Implications for biochemical functions and drug design. Nucleic Acids Res. 2003, 31, 7117–7130. [Google Scholar] [CrossRef] [PubMed]
- Clercq, E. Antivirals and antiviral strategies. Nat. Rev. Microbiol. 2004, 2, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Uchide, N.; Ohyama, K.; Bessho, T.; Yuan, B.; Yamakawa, T. Effect of antioxidants on apoptosis induced by influenza virus infection: Inhibition of viral gene replication and transcription with pyrrolidine dithiocarbamate. Antivir. Res. 2002, 56, 207–217. [Google Scholar] [CrossRef]
- Suara, R.O.; Crowe, J.E. Effect of zinc salts on respiratory syncytial virus replication. Antimicrob. Agents Chemother. 2004, 48, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Gaudernak, E.; Seipelt, J.; Triendl, A.; Grassauer, A.; Kuechler, E. Antiviral Effects of Pyrrolidine Dithiocarbamate on Human Rhinoviruses. JVI 2002, 76, 6004–6015. [Google Scholar] [CrossRef]
- Si, X.; McManus, B.M.; Zhang, J.; Yuan, J.; Cheung, C.; Esfandiarei, M.; Suarez, A.; Morgan, A.; Luo, H. Pyrrolidine dithiocarbamate reduces coxsackievirus B3 replication through inhibition of the ubiquitin-proteasome pathway. JVI 2005, 79, 8014–8023. [Google Scholar] [CrossRef]
- Korant, B.D.; Kauer, J.C.; Butterworth, B.E. Zinc ions inhibit replication of rhinoviruses. Nature 1974, 248, 588–590. [Google Scholar] [CrossRef]
- Polatnick, J.; Bachrach, H.L. Effect of Zinc and Other Chemical Agents on Foot-and-Mouth Disease Virus Replication. Antimicrob. Agents Chemother. 1978, 13, 731–734. [Google Scholar] [CrossRef]
- Lanke, K.; Krenn, B.M.; Melchers, W.J.G.; Seipelt, J.; van Kuppeveld, F.J.M. PDTC inhibits picornavirus polyprotein processing and RNA replication by transporting zinc ions into cells. J. Gen. Virol. 2007, 88, 1206–1217. [Google Scholar] [CrossRef]
- Krenn, B.M.; Gaudernak, E.; Holzer, B.; Lanke, K.; van Kuppeveld, F.J.M.; Seipelt, J. Antiviral Activity of the Zinc Ionophores Pyrithione and Hinokitiol against Picornavirus Infections. JVI 2009, 83, 58–64. [Google Scholar] [CrossRef]
- Haraguchi, Y.; Sakurai, H.; Hussain, S.; Anner, B.M.; Hoshino, H. Inhibition of HIV-1 infection by zinc group metal compounds. Antivir. Res. 1999, 43, 123–133. [Google Scholar] [CrossRef]
- Katz, E.; Margalith, E. Inhibition of vaccinia virus maturation by zinc chloride. Antimicrob. Agents Chemother. 1981, 19, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Kümel, G.; Schrader, S.; Zentgraf, H.; Daus, H.; Brendel, M. The mechanism of the antiherpetic activity of zinc sulphate. J. Gen. Virol. 1990, 71 Pt 12, 2989–2997. [Google Scholar] [CrossRef]
- Korant, B.D.; Butterworth, B.E. Inhibition by zinc of rhinovirus protein cleavage: Interaction of zinc with capsid polypeptides. JVI 1976, 18, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Novick, S.G.; Godfrey, J.C.; Godfrey, N.J.; Wilder, H.R. How does zinc modify the common cold? Clinical observations and implications regarding mechanisms of action. Med. Hypotheses 1996, 46, 295–302. [Google Scholar] [CrossRef]
- Oxford, J.S.; Perrin, D.D. Inhibition of the particle-associated RNA-dependent RNA polymerase activity of influenza viruses by chelating agents. J. Gen. Virol. 1974, 23, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Wright-Minogue, J.; Fang, J.W.S.; Baroudy, B.M.; Lau, J.Y.N.; Hong, Z. Characterization of Soluble Hepatitis C Virus RNA-Dependent RNA Polymerase Expressed in Escherichia coli. JVI 1999, 73, 1649–1654. [Google Scholar] [CrossRef]
- Hung, M.; Gibbs, C.S.; Tsiang, M. Biochemical characterization of rhinovirus RNA-dependent RNA polymerase. Antivir. Res. 2002, 56, 99–114. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System; Version 1.8; Schrödinger, LLC: New York, NY, USA, 2002.
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Cao, L.; Wang, T.; Wang, Q.; Lou, Z.; Rao, Z. SARS-CoV-2 RNA-Dependent RNA Polymerase in Complex with Cofactors in Reduced Condition. Available online: https://www.rcsb.org/structure/7btf (accessed on 15 April 2020).
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 269. [Google Scholar] [CrossRef]
- McChesney, E.W. Animal toxicity and pharmacokinetics of hydroxychloroquine sulfate. Am. J. Med. 1983, 75, 11–18. [Google Scholar] [CrossRef]
- COVID-19: Reminder of Risk of Serious Side Effects with Chloroquine and Hydroxychloroquine. Available online: http://urpl.gov.pl/sites/default/files/CQ-HCQ%20-%2004-2020%20-%20public%20health%20communication_0.pdf (accessed on 23 April 2020).
- Rainsford, K.D.; Parke, A.L.; Clifford-Rashotte, M.; Kean, W.F. Therapy and pharmacological properties of hydroxychloroquine and chloroquine in treatment of systemic lupus erythematosus, rheumatoid arthritis and related diseases. Inflammopharmacology 2015, 23, 231–269. [Google Scholar] [CrossRef] [PubMed]
- Krafts, K.; Hempelmann, E.; Skórska-Stania, A. From methylene blue to chloroquine: A brief review of the development of an antimalarial therapy. Parasitol. Res. 2012, 111, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mercer, E.; Rekedal, L.; Garg, R.; Lu, B.; Massarotti, E.M.; Solomon, D.H. Hydroxychloroquine improves insulin sensitivity in obese non-diabetic individuals. Arthritis Res. Ther. 2012, 14, R135. [Google Scholar] [CrossRef] [PubMed]
- Solomon, D.H.; Massarotti, E.; Garg, R.; Liu, J.; Canning, C.; Schneeweiss, S. Association between disease-modifying antirheumatic drugs and diabetes risk in patients with rheumatoid arthritis and psoriasis. JAMA 2011, 305, 2525–2531. [Google Scholar] [CrossRef]
- Thomé, R.; Lopes, S.C.P.; Costa, F.T.M.; Verinaud, L. Chloroquine: Modes of action of an undervalued drug. Immunol. Lett. 2013, 153, 50–57. [Google Scholar] [CrossRef]
- Savarino, A.; Boelaert, J.R.; Cassone, A.; Majori, G.; Cauda, R. Effects of chloroquine on viral infections: An old drug against today’s diseases. Lancet Infect. Dis. 2003, 3, 722–727. [Google Scholar] [CrossRef]
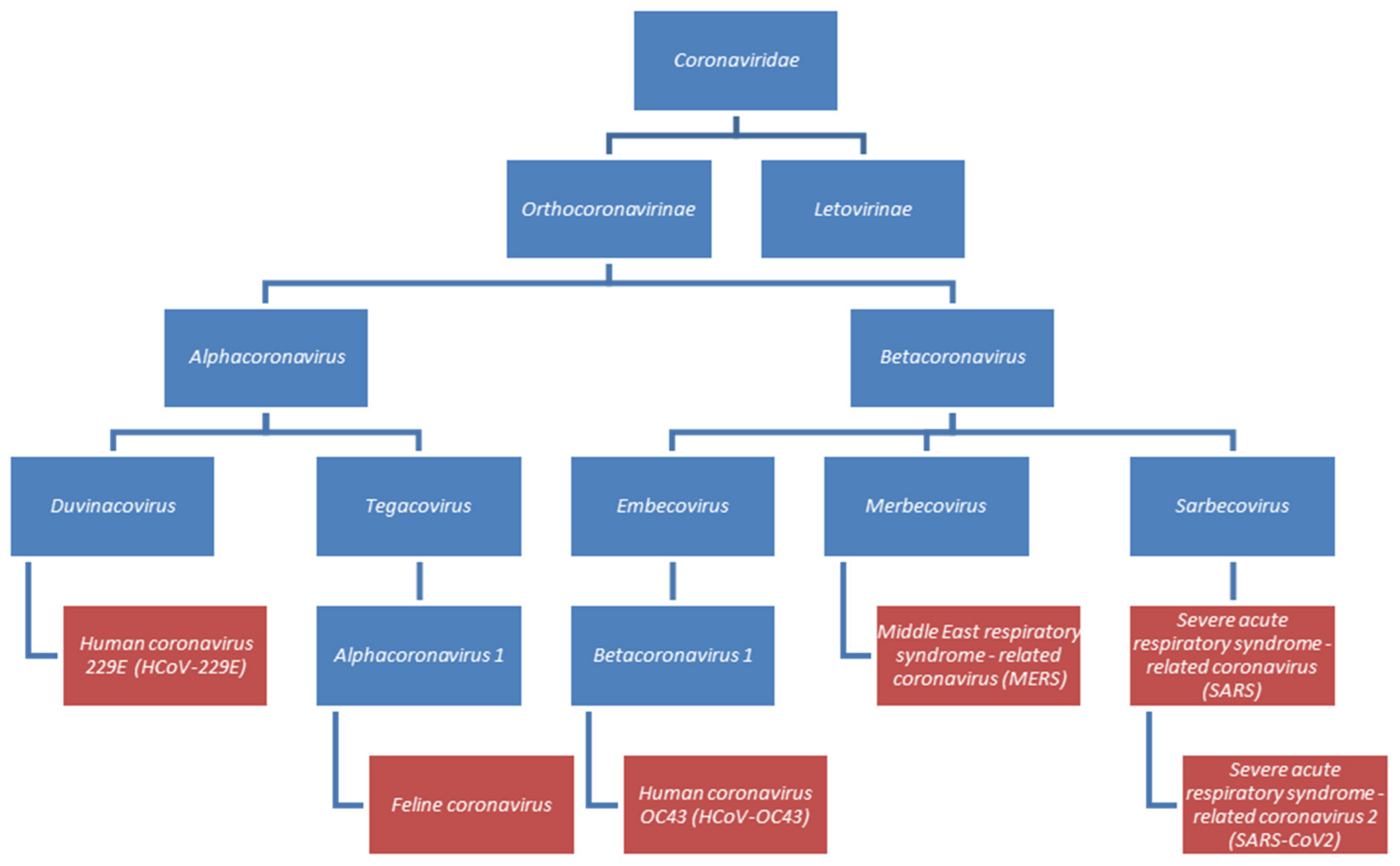
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Maes, P.; Neyts, J.; van Ranst, M. In vitro inhibition of severe acute respiratory syndrome coronavirus by chloroquine. Biochem. Biophys. Res. Commun. 2004, 323, 264–268. [Google Scholar] [CrossRef]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.-Y. Coronaviruses-drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef]
- Vincent, M.J.; Bergeron, E.; Benjannet, S.; Erickson, B.R.; Rollin, P.E.; Ksiazek, T.G.; Seidah, N.G.; Nichol, S.T. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol. J. 2005, 2, 69. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Shen, H.-M. Targeting the Endocytic Pathway and Autophagy Process as a Novel Therapeutic Strategy in COVID-19. Int. J. Biol. Sci. 2020, 16, 1724–1731. [Google Scholar] [CrossRef]
- Millet, J.K.; Whittaker, G.R. Physiological and molecular triggers for SARS-CoV membrane fusion and entry into host cells. Virology 2018, 517, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 15 April 2020).
- de Wilde, A.H.; Jochmans, D.; Posthuma, C.C.; Zevenhoven-Dobbe, J.C.; van Nieuwkoop, S.; Bestebroer, T.M.; van den Hoogen, B.G.; Neyts, J.; Snijder, E.J. Screening of an FDA-Approved Compound Library Identifies Four Small-Molecule Inhibitors of Middle East Respiratory Syndrome Coronavirus Replication in Cell Culture. Antimicrob. Agents Chemother. 2014, 58, 4875–4884. [Google Scholar] [CrossRef]
- Goldman, L.; La Cecil, R.F. Goldman’s Cecil Medicine, 24th ed.; Saunders: Philadelphia, PA, USA, 2011; ISBN 9781437736083. [Google Scholar]
- Pelczar, M.J. Microbiology (An Application Based Approach), 1st ed.; Tata McGraw Hill Education Private Limited: New Delhi, India, 2010; ISBN 978-0-07-015147-5. [Google Scholar]
- Keyaerts, E.; Li, S.; Vijgen, L.; Rysman, E.; Verbeeck, J.; van Ranst, M.; Maes, P. Antiviral Activity of Chloroquine against Human Coronavirus OC43 Infection in Newborn Mice. Antimicrob. Agents Chemother. 2009, 53, 3416–3421. [Google Scholar] [CrossRef]
- Kono, M.; Tatsumi, K.; Imai, A.M.; Saito, K.; Kuriyama, T.; Shirasawa, H. Inhibition of human coronavirus 229E infection in human epithelial lung cells (L132) by chloroquine: Involvement of p38 MAPK and ERK. Antivir. Res. 2008, 77, 150–152. [Google Scholar] [CrossRef]
- Kumar, R.; Khandelwal, N.; Thachamvally, R.; Tripathi, B.N.; Barua, S.; Kashyap, S.K.; Maherchandani, S.; Kumar, N. Role of MAPK/MNK1 signaling in virus replication. Virus Res. 2018, 253, 48–61. [Google Scholar] [CrossRef]
- Hartmann, K. Feline infectious peritonitis. Vet. Clin. N. Am. Small Anim. Pract. 2005, 35, 39–79. [Google Scholar] [CrossRef]
- Takano, T.; Katoh, Y.; Doki, T.; Hohdatsu, T. Effect of chloroquine on feline infectious peritonitis virus infection in vitro and in vivo. Antivir. Res. 2013, 99, 100–107. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. A Study of Hydroxychloroquine, Vitamin C, Vitamin D, and Zinc for the Prevention of COVID-19 Infection (HELPCOVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04335084 (accessed on 15 April 2020).
- ClinicalTrials.gov. Proflaxis Using Hydroxychloroquine Plus Vitamins-Zinc During COVID-19 Pandemia. Available online: https://clinicaltrials.gov/ct2/show/NCT04326725 (accessed on 15 April 2020).
- ClinicalTrials.gov. A Study of Quintuple Therapy to Treat COVID-19 Infection (HAZDpaC). Available online: https://clinicaltrials.gov/ct2/show/NCT04334512 (accessed on 15 April 2020).
- ClinicalTrials.gov. Anti-inflammatory/Antioxidant Oral Nutrition Supplementation in COVID-19 (ONSCOVID19). Available online: https://clinicaltrials.gov/ct2/show/NCT04323228 (accessed on 15 April 2020).
- ClinicalTrials.gov. Coronavirus 2019 (COVID-19)-Using Ascorbic Acid and Zinc Supplementation (COVIDAtoZ). Available online: https://clinicaltrials.gov/ct2/show/NCT04342728 (accessed on 15 April 2020).
- ClinicalTrials.gov. Hydroxychloroquine and Zinc with Either Azithromycin or Doxycycline for Treatment of COVID-19 in Outpatient Setting. Available online: https://clinicaltrials.gov/ct2/show/NCT04370782 (accessed on 7 July 2020).
- ClinicalTrials.gov. A Study of Hydroxychloroquine and Zinc in the Prevention of COVID-19 Infection in Military Healthcare Workers (COVID-Milit). Available online: https://clinicaltrials.gov/ct2/show/NCT04377646 (accessed on 7 July 2020).
- ClinicalTrials.gov. Impact of Zinc and Vitamin D3 Supplementation on the Survival of Aged Patients Infected With COVID-19 (ZnD3-CoVici). Available online: https://clinicaltrials.gov/ct2/show/NCT04351490 (accessed on 7 July 2020).
- ClinicalTrials.gov. Early Intervention in COVID-19: Favipiravir Verses Standard Care (PIONEER). Available online: https://clinicaltrials.gov/ct2/show/NCT04373733 (accessed on 28 August 2020).
- ClinicalTrials.gov. Zinc with Chloroquine/Hydroxychloroquine in Treatment of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04447534 (accessed on 7 July 2020).
- ClinicalTrials.gov. International ALLIANCE Study of Therapies to Prevent Progression of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04395768 (accessed on 7 July 2020).
- ClinicalTrials.gov. COVID-19 Prophylaxis with Hydroxychloroquine Associated with Zinc for High-Risk Healthcare Workers. Available online: https://clinicaltrials.gov/ct2/show/NCT04384458 (accessed on 7 July 2020).
- ClinicalTrials.gov. A Preventive Treatment for Migrant Workers at High-risk of Covid-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04446104 (accessed on 7 July 2020).
- ClinicalTrials.gov. New Antiviral Drugs for Treatment of COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04392427 (accessed on 7 July 2020).
- ClinicalTrials.gov. Covid-19 and Diabetes in West of Algeria (COVIDIAB-13). Available online: https://clinicaltrials.gov/ct2/show/NCT04412746 (accessed on 7 July 2020).
- ClinicalTrials.gov. The Study of Quadruple Therapy Zinc, Quercetin, Bromelain and Vitamin C on the Clinical Outcomes of Patients Infected with COVID-19. Available online: https://clinicaltrials.gov/ct2/show/NCT04468139 (accessed on 30 July 2020).
- ClinicalTrials.gov. Efficacy of Subcutaneous Ivermectin with or without Zinc and Nigella Sativa in COVID-19 Patients (SINZ-COVID-PK). Available online: https://clinicaltrials.gov/ct2/show/NCT04472585 (accessed on 30 July 2020).
- ClinicalTrials.gov. Clearing the Fog: Is Hydroxychloroquine Effective in Reducing COVID-19 Progression (COVID-19). Available online: https://clinicaltrials.gov/ct2/show/NCT04491994 (accessed on 30 July 2020).
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Milewska, A.; Chi, Y.; Szczepanski, A.; Barreto-Duran, E.; Liu, K.; Liu, D.; Guo, X.; Ge, Y.; Li, J.; Cui, L.; et al. HTCC as a highly effective polymeric inhibitor of SARS-CoV-2 and MERS-CoV. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rut, W.; Groborz, K.; Zhang, L.; Sun, X.; Zmudzinski, M.; Hilgenfeld, R.; Drag, M. Substrate specificity profiling of SARS-CoV-2 M pro protease provides basis for anti-COVID-19 drug design. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef]
- Han, Y.-S.; Chang, G.-G.; Juo, C.-G.; Lee, H.-J.; Yeh, S.-H.; Hsu, J.T.-A.; Chen, X. Papain-like protease 2 (PLP2) from severe acute respiratory syndrome coronavirus (SARS-CoV): Expression, purification, characterization, and inhibition. Biochemistry 2005, 44, 10349–10359. [Google Scholar] [CrossRef]
- Imbert, I.; Guillemot, J.-C.; Bourhis, J.-M.; Bussetta, C.; Coutard, B.; Egloff, M.-P.; Ferron, F.; Gorbalenya, A.E.; Canard, B. A second, non-canonical RNA-dependent RNA polymerase in SARS coronavirus. EMBO J. 2006, 25, 4933–4942. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Lin, W.-H.; Hsu, J.T.-A.; Hsieh, H.-P. Antiviral drug discovery against SARS-CoV. Curr. Med. Chem. 2006, 13, 2003–2020. [Google Scholar] [CrossRef]
- Adedeji, A.O.; Marchand, B.; te Velthuis, A.J.W.; Snijder, E.J.; Weiss, S.; Eoff, R.L.; Singh, K.; Sarafianos, S.G. Mechanism of nucleic acid unwinding by SARS-CoV helicase. PLoS ONE 2012, 7, e36521. [Google Scholar] [CrossRef]
- Boelaert, J.R.; Piette, J.; Sperber, K. The potential place of chloroquine in the treatment of HIV-1-infected patients. J. Clin. Virol. 2001, 20, 137–140. [Google Scholar] [CrossRef]
- Savarino, A.; Gennero, L.; Sperber, K.; Boelaert, J.R. The anti-HIV-1 activity of chloroquine. J. Clin. Virol. 2001, 20, 131–135. [Google Scholar] [CrossRef]
- Naarding, M.A.; Baan, E.; Pollakis, G.; Paxton, W.A. Effect of chloroquine on reducing HIV-1 replication in vitro and the DC-SIGN mediated transfer of virus to CD4 + T-lymphocytes. Retrovirology 2007, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Savarino, A.; Lucia, M.B.; Rastrelli, E.; Rutella, S.; Golotta, C.; Morra, E.; Tamburrini, E.; Perno, C.F.; Boelaert, J.R.; Sperber, K.; et al. Anti-HIV Effects of Chloroquine: Inhibition of viral particle glycosylation and synergism with protease inhibitors. J. Acquir. Immune Defic. Syndr. 2004, 35, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, F.; Smith, K.; Hoven, A. Chloroquine and Hydroxychloroquine as Inhibitors of Human Immunodeficiency Virus (HIV-1) Activity. Curr. Pharm. Des. 2004, 10, 2643–2648. [Google Scholar] [CrossRef]
- Savarino, A.; Gennero, L.; Chen, H.C.; Serrano, D.; Malavasi, F.; Boelaert, J.R.; Sperber, K. Anti-HIV effects of chloroquine: Mechanisms of inhibition and spectrum of activity. J. Acquir. Immune Defic. Syndr. 2001, 15, 2221–2229. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Yang, J.; Diagne, A. Chloroquine inhibits HIV-1 replication in human peripheral blood lymphocytes. Immunol. Lett. 1998, 64, 45–47. [Google Scholar] [CrossRef]
- Bishop, N.E. Examination of potential inhibitors of hepatitis A virus uncoating. Intervirology 1998, 41, 261–271. [Google Scholar] [CrossRef]
- Kouroumalis, E.A.; Koskinas, J. Treatment of chronic active hepatitis B (CAH B) with chloroquine: A preliminary report. Ann. Acad. Med. Singap. 1986, 15, 149–152. [Google Scholar]
- Mizui, T.; Yamashina, S.; Tanida, I.; Takei, Y.; Ueno, T.; Sakamoto, N.; Ikejima, K.; Kitamura, T.; Enomoto, N.; Sakai, T.; et al. Inhibition of hepatitis C virus replication by chloroquine targeting virus-associated autophagy. J. Gastroenterol. 2010, 45, 195–203. [Google Scholar] [CrossRef]
- Moraes Falcão, L.; Terrabuio, D.R.B.; Diniz, M.A.; da Silva Evangelista, A.; Souza, F.G.; R Cancado, E.L. Efficacy and safety of chloroquine plus prednisone for the treatment of autoimmune hepatitis in a randomized trial. JGH Open 2020, 4, 371–377. [Google Scholar] [CrossRef]
- Kronenberger, P.; Vrijsen, R.; Boeyé, A. Chloroquine induces empty capsid formation during poliovirus eclipse. JVI 1991, 65, 7008–7011. [Google Scholar] [CrossRef] [PubMed]
- Ooi, E.E.; Chew, J.S.W.; Loh, J.P.; Chua, R.C.S. In vitro inhibition of human influenza a virus replication by chloroquine. Virol. J. 2006, 3, 39. [Google Scholar] [CrossRef] [PubMed]
- Vigerust, D.J.; McCullers, J.A. Chloroquine is effective against influenza A virus in vitro but not in vivo. Influenza Other Respir. Viruses 2007, 1, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Aoki, H.; Tsurumi, T.; Sugiura, Y.; Nishiyama, Y.; Suzuki, S.; Maeno, K. Mechanism of Uncoating of Influenza B Virus in MDCK Cells: Action of Chloroquine. J. Gen. Virol. 1983, 64, 1149–1156. [Google Scholar] [CrossRef]
- Yan, Y.; Zou, Z.; Sun, Y.; Li, X.; Xu, K.-F.; Wei, Y.; Jin, N.; Jiang, C. Anti-malaria drug chloroquine is highly effective in treating avian influenza A H5N1 virus infection in an animal model. Cell Res. 2013, 23, 300–302. [Google Scholar] [CrossRef]
- Delvecchio, R.; Higa, L.M.; Pezzuto, P.; Valadão, A.L.; Garcez, P.P.; Monteiro, F.L.; Loiola, E.C.; Dias, A.A.; Silva, F.J.M.; Aliota, M.T.; et al. Chloroquine, an Endocytosis Blocking Agent, Inhibits Zika Virus Infection in Different Cell Models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Ji, X.; Quanquin, N.; Deng, Y.-Q.; Tian, M.; Aliyari, R.; Zuo, X.; Yuan, L.; Afridi, S.K.; et al. Chloroquine, a FDA-approved Drug, Prevents Zika Virus Infection and its Associated Congenital Microcephaly in Mice. EBioMedicine 2017, 24, 189–194. [Google Scholar] [CrossRef]
- Zhang, S.; Yi, C.; Li, C.; Zhang, F.; Peng, J.; Wang, Q.; Liu, X.; Ye, X.; Li, P.; Wu, M.; et al. Chloroquine inhibits endosomal viral RNA release and autophagy-dependent viral replication and effectively prevents maternal to fetal transmission of Zika virus. Antivir. Res. 2019, 169, 104547. [Google Scholar] [CrossRef]
- Dowall, S.D.; Bosworth, A.; Watson, R.; Bewley, K.; Taylor, I.; Rayner, E.; Hunter, L.; Pearson, G.; Easterbrook, L.; Pitman, J.; et al. Chloroquine inhibited Ebola virus replication in vitro but failed to protect against infection and disease in the in vivo guinea pig model. J. Gen. Virol. 2015, 96, 3484–3492. [Google Scholar] [CrossRef]
- Ferraris, O.; Moroso, M.; Pernet, O.; Emonet, S.; Ferrier Rembert, A.; Paranhos-Baccalà, G.; Peyrefitte, C.N. Evaluation of Crimean-Congo hemorrhagic fever virus in vitro inhibition by chloroquine and chlorpromazine, two FDA approved molecules. Antivir. Res. 2015, 118, 75–81. [Google Scholar] [CrossRef]
- Tsiang, H.; Superti, F. Ammonium chloride and chloroquine inhibit rabies virus infection in neuroblastoma cells. Brief report. Arch. Virol. 1984, 81, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Santhosh, S.R.; Tiwari, M.; Lakshmana Rao, P.V.; Parida, M. Assessment of in vitro prophylactic and therapeutic efficacy of chloroquine against Chikungunya virus in vero cells. J. Med. Virol. 2010, 82, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Delogu, I.; de Lamballerie, X. Chikungunya disease and chloroquine treatment. J. Med. Virol. 2011, 83, 1058–1059. [Google Scholar] [CrossRef] [PubMed]
- Farias, K.J.S.; Machado, P.R.L.; da Fonseca, B.A.L. Chloroquine Inhibits Dengue Virus Type 2 Replication in Vero Cells but Not in C6/36 Cells. Sci. World J. 2013, 2013, 1–5. [Google Scholar] [CrossRef]
- Farias, K.J.S.; Machado, P.R.L.; de Almeida Junior, R.F.; de Aquino, A.A.; da Fonseca, B.A.L. Chloroquine interferes with dengue-2 virus replication in U937 cells. Microbiol. Immunol. 2014, 58, 318–326. [Google Scholar] [CrossRef]
- Farias, K.J.S.; Machado, P.R.L.; Muniz, J.A.P.C.; Imbeloni, A.A.; da Fonseca, B.A.L. Antiviral Activity of Chloroquine Against Dengue Virus Type 2 Replication in Aotus Monkeys. Viral Immunol. 2015, 28, 161–169. [Google Scholar] [CrossRef]
- Freiberg, A.N.; Worthy, M.N.; Lee, B.; Holbrook, M.R. Combined chloroquine and ribavirin treatment does not prevent death in a hamster model of Nipah and Hendra virus infection. Arch. Virol. 2010, 91, 765–772. [Google Scholar] [CrossRef]
- Koyama, A.H.; Uchida, T. Inhibition of multiplication of herpes simplex virus type 1 by ammonium chloride and chloroquine. Virology 1984, 138, 332–335. [Google Scholar] [CrossRef]
- Sperber, K.; Louie, M.; Kraus, T.; Proner, J.; Sapira, E.; Lin, S.; Stecher, V.; Mayer, L. Hydroxychloroquine treatment of patients with human immunodeficiency virus type 1. Clin. Ther. 1995, 17, 622–636. [Google Scholar] [CrossRef]
- Sperber, K.; Chiang, G.; Chen, H.; Ross, W.; Chusid, E.; Gonchar, M.; Chow, R.; Liriano, O. Comparison of hydroxychloroquine with zidovudine in asymptomatic patients infected with human immunodeficiency virus type 1. Clin. Ther. 1997, 19, 913–923. [Google Scholar] [CrossRef]
- Sperber, K.; Kalb, T.H.; Stecher, V.J.; Banerjee, R.; Mayer, L. Inhibition of human immunodeficiency virus type 1 replication by hydroxychloroquine in T cells and monocytes. AIDS Res. Hum. Retrovir. 1993, 9, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Fesen, M.R.; Kohn, K.W.; Leteurtre, F.; Pommier, Y. Inhibitors of human immunodeficiency virus integrase. Proc. Natl. Acad. Sci. USA 1993, 90, 2399–2403. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C.; Lin, J.K.; Chen, S.S. Inhibition of HIV-1 Tat-mediated transactivation by quinacrine and chloroquine. Biochem. Biophys. Res. Commun. 1996, 226, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Morra, E.; Savarino, A.; Gennero, L.; Pescarmona, G.P. Effects of chloroquine on iron metabolism in a lymphocytic cell line. J. Clin. Virol. 2000, 16, 91–92. [Google Scholar]
- Tsai, W.P.; Nara, P.L.; Kung, H.F.; Oroszlan, S. Inhibition of human immunodeficiency virus infectivity by chloroquine. AIDS Res. Hum. Retrovir. 1990, 6, 481–489. [Google Scholar] [CrossRef]
- Tricou, V.; Minh, N.N.; Van, T.P.; Lee, S.J.; Farrar, J.; Wills, B.; Tran, H.T.; Simmons, C.P. A randomized controlled trial of chloroquine for the treatment of dengue in Vietnamese adults. PLoS Negl. Trop. Dis. 2010, 4, e785. [Google Scholar] [CrossRef]
- Borges, M.C.; Castro, L.A.; Fonseca, B.A.L. Chloroquine use improves dengue-related symptoms. Mem. Inst. Oswaldo Cruz 2013, 108, 596–599. [Google Scholar] [CrossRef]
- Madrid, P.B.; Chopra, S.; Manger, I.D.; Gilfillan, L.; Keepers, T.R.; Shurtleff, A.C.; Green, C.E.; Iyer, L.V.; Dilks, H.H.; Davey, R.A.; et al. A systematic screen of FDA-approved drugs for inhibitors of biological threat agents. PLoS ONE 2013, 8, e60579. [Google Scholar] [CrossRef]
- Guo, C.; Zhu, Z.; Wang, X.; Chen, Y.; Liu, X. Pyrithione inhibits porcine reproductive and respiratory syndrome virus replication through interfering with NF-κB and heparanase. Vet. Microbiol. 2017, 201, 231–239. [Google Scholar] [CrossRef]
- Qiu, M.; Chen, Y.; Chu, Y.; Song, S.; Yang, N.; Gao, J.; Wu, Z. Zinc ionophores pyrithione inhibits herpes simplex virus replication through interfering with proteasome function and NF-κB activation. Antivir. Res. 2013, 100, 44–53. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef] [PubMed]
- Uchide, N.; Toyoda, H. Antioxidant therapy as a potential approach to severe influenza-associated complications. Molecules 2011, 16, 2032–2052. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Sun, G.; Guo, W.; Huang, Y.; Sun, W.; Zhao, F.; Hu, K. Inhibition of hepatitis B virus replication by quercetin in human hepatoma cell lines. Virol. Sin. 2015, 30, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Rojas, Á.; Del Campo, J.A.; Clement, S.; Lemasson, M.; García-Valdecasas, M.; Gil-Gómez, A.; Ranchal, I.; Bartosch, B.; Bautista, J.D.; Rosenberg, A.R.; et al. Effect of Quercetin on Hepatitis C Virus Life Cycle: From Viral to Host Targets. Sci. Rep. 2016, 6, 55. [Google Scholar] [CrossRef]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Golan-Goldhirsh, A.; Benhar, I.; Tur-Kaspa, R.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral. Hepat. 2012, 19, e81–e88. [Google Scholar] [CrossRef]
- Qiu, X.; Kroeker, A.; He, S.; Kozak, R.; Audet, J.; Mbikay, M.; Chrétien, M. Prophylactic Efficacy of Quercetin 3-β- O -d-Glucoside against Ebola Virus Infection. Antimicrob. Agents Chemother. 2016, 60, 5182–5188. [Google Scholar] [CrossRef]
- Chiow, K.H.; Phoon, M.C.; Putti, T.; Tan, B.K.H.; Chow, V.T. Evaluation of antiviral activities of Houttuynia cordata Thunb. extract, quercetin, quercetrin and cinanserin on murine coronavirus and dengue virus infection. Asian Pac. J. Trop. Med. 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Johari, J.; Kianmehr, A.; Mustafa, M.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785–16795. [Google Scholar] [CrossRef]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-O-D-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef]
- Huang, Q.; Sivaramakrishna, R.P.; Ludwig, K.; Korte, T.; Böttcher, C.; Herrmann, A. Early steps of the conformational change of influenza virus hemagglutinin to a fusion active state. Biochim. Biophys. Acta Biomembr. 2003, 1614, 3–13. [Google Scholar] [CrossRef]
- Gaspari, P.; Banerjee, T.; Malachowski, W.P.; Muller, A.J.; Prendergast, G.C.; DuHadaway, J.; Bennett, S.; Donovan, A.M. Structure-activity study of brassinin derivatives as indoleamine 2,3-dioxygenase inhibitors. J. Med. Chem. 2006, 49, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-L.; Feng, Y.-P.; Jiang, Y.-Y.; Liu, S.-Y.; Ding, G.-Y.; Li, R.-T. Synthesis and in vitro antitumor activity of 4(3H)-quinazolinone derivatives with dithiocarbamate side chains. Bioorg. Med. Chem. Lett. 2005, 15, 1915–1917. [Google Scholar] [CrossRef]
- Li, Y.; Qi, H.; Li, X.; Hou, X.; Lu, X.; Xiao, X. A novel dithiocarbamate derivative induces cell apoptosis through p53-dependent intrinsic pathway and suppresses the expression of the E6 oncogene of human papillomavirus 18 in HeLa cells. Apoptosis 2015, 20, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, E.C.; Kern, P.; Dietrich, M.; Ernst, M.; Flad, H.D.; Bock, P.; Group, G.D.S. Inhibition of HIV progression by dithiocarb. Lancet 1990, 335, 679–682. [Google Scholar] [CrossRef]
- Brooks, H.A.; Gardner, D.; Poyser, J.P.; King, T.J. The structure and absolute stereochemistry of zincophorin (antibiotic M144255): A monobasic carboxylic acid ionophore having a remarkable specificity for divalent cations. J. Antibiot. 1984, 37, 1501–1504. [Google Scholar] [CrossRef]
- Gräfe, U.; Schade, W.; Roth, M.; Radics, L.; Incze, M.; Ujszászy, K. Griseochelin, a novel carboxylic acid antibiotic from Streptomyces griseus. J. Antibiot. 1984, 37, 836–846. [Google Scholar] [CrossRef][Green Version]
- Radics, L. The constitution of griseochelin. J. Chem. Soc. Chem. Commun. 1984, 1010, 599. [Google Scholar] [CrossRef]
- Gräfe, U.; Tonew, E.; Schade, W.; Reinhardt, G.; Hartl, A. Ger. (East). Patent DD 231 793 1986. [Google Scholar]
- Tonew, E.; Tonew, M.; Graefe, U.; Zöpel, P. Griseochelinmethylester, ein neues Polyether-Derivat mit antiviraler Wirksamkeit. Pharmazie 1988, 43, 717–719. [Google Scholar]
- Huang, R.; Wallqvist, A.; Covell, D.G. Anticancer metal compounds in NCI’s tumor-screening database: Putative mode of action. Biochem. Pharmacol. 2005, 69, 1009–1039. [Google Scholar] [CrossRef]
- Potocki, S.; Rowinska-Zyrek, M.; Witkowska, D.; Pyrkosz, M.; Szebesczyk, A.; Krzywoszynska, K.; Kozlowski, H. Metal transport and homeostasis within the human body: Toxicity associated with transport abnormalities. Curr. Med. Chem. 2012, 19, 2738–2759. [Google Scholar] [CrossRef] [PubMed]
- Magda, D.; Lecane, P.; Wang, Z.; Hu, W.; Thiemann, P.; Ma, X.; Dranchak, P.K.; Wang, X.; Lynch, V.; Wei, W.; et al. Synthesis and Anticancer Properties of Water-Soluble Zinc Ionophores. Cancer Res. 2008, 68, 5318–5325. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-Q.; Lind, S.E. Metal ionophores: An emerging class of anticancer drugs. IUBMB Life 2009, 61, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, D.B.; Richardson, D.R. Iron chelators as anti-neoplastic agents: Current developments and promise of the PIH class of chelators. Curr. Med. Chem. 2003, 10, 1035–1049. [Google Scholar] [CrossRef][Green Version]
- Kontoghiorghes, G.J.; Efstathiou, A.; Ioannou-Loucaides, S.; Kolnagou, A. Chelators Controlling Metal Metabolism and Toxicity Pathways: Applications in Cancer Prevention, Diagnosis and Treatment. Hemoglobin 2008, 32, 217–227. [Google Scholar] [CrossRef]
- Gaur, K.; Vázquez-Salgado, A.; Duran-Camacho, G.; Dominguez-Martinez, I.; Benjamín-Rivera, J.; Fernández-Vega, L.; Carmona Sarabia, L.; Cruz García, A.; Pérez-Deliz, F.; Méndez Román, J.; et al. Iron and Copper Intracellular Chelation as an Anticancer Drug Strategy. Inorganics 2018, 6, 126. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.H.; Moon, S.J.; Chung, K.C.; Hsu, C.Y.; Seo, J.T.; Ahn, Y.S. Pyrithione, a zinc ionophore, inhibits NF-kappaB activation. Biochem. Biophys. Res. Commun. 1999, 259, 505–509. [Google Scholar] [CrossRef]
- Knight, D.W.; Hartung, J. 1-Hydroxypyridine-2(1H)-thione. In Encyclopedia of Reagents for Organic Synthesis; John Wiley & Sons, Ltd.: Chichester, UK, 2001; p. 1875. ISBN 0471936235. [Google Scholar]
- Feng, P.; Li, T.L.; Guan, Z.X.; Franklin, R.B.; Costello, L.C. Effect of zinc on prostatic tumorigenicity in nude mice. Ann. N. Y. Acad. Sci. 2003, 1010, 316–320. [Google Scholar] [CrossRef]
- Ding, W.-Q.; Liu, B.; Vaught, J.L.; Yamauchi, H.; Lind, S.E. Anticancer activity of the antibiotic clioquinol. Cancer Res. 2005, 65, 3389–3395. [Google Scholar] [CrossRef]
- Ritchie, C.W.; Bush, A.I.; Mackinnon, A.; Macfarlane, S.; Mastwyk, M.; MacGregor, L.; Kiers, L.; Cherny, R.; Li, Q.-X.; Tammer, A.; et al. Metal-protein attenuation with iodochlorhydroxyquin (clioquinol) targeting Abeta amyloid deposition and toxicity in Alzheimer disease: A pilot phase 2 clinical trial. Arch. Neurol. 2003, 60, 1685–1691. [Google Scholar] [CrossRef]
- Misra, H.P. Reaction of copper-zinc superoxide dismutase with diethyldithiocarbamate. J. Biol. Chem. 1979, 254, 11623–11628. [Google Scholar] [PubMed]
- Schreck, R.; Meier, B.; Männel, D.N.; Dröge, W.; Baeuerle, P.A. Dithiocarbamates as potent inhibitors of nuclear factor kappa B activation in intact cells. J. Exp. Med. 1992, 175, 1181–1194. [Google Scholar] [CrossRef] [PubMed]
- Verhaegh, G.W.; Richard, M.J.; Hainaut, P. Regulation of p53 by metal ions and by antioxidants: Dithiocarbamate down-regulates p53 DNA-binding activity by increasing the intracellular level of copper. Mol. Cell. Biol. 1997, 17, 5699–5706. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.H.; Momand, J. Pyrrolidine dithiocarbamate prevents p53 activation and promotes p53 cysteine residue oxidation. J. Biol. Chem. 1998, 273, 18898–18905. [Google Scholar] [CrossRef] [PubMed]
- Nobel, C.I.; Kimland, M.; Lind, B.; Orrenius, S.; Slater, A.F. Dithiocarbamates induce apoptosis in thymocytes by raising the intracellular level of redox-active copper. J. Biol. Chem. 1995, 270, 26202–26208. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, J.H.; Xu, J.; Hsu, C.Y.; Ahn, Y.S. Pyrrolidine Dithiocarbamate Induces Bovine Cerebral Endothelial Cell Death by Increasing the Intracellular Zinc Level. J. Neurochem. 1999, 72, 1586–1592. [Google Scholar] [CrossRef]
- Puschner, B.; Dechant, J.E. Common Toxins in Equine Practice. In Equine Medicine, 7th ed.; Robinson, N.E., Sprayberry, K.A., Eds.; Elsevier: St. Louis, MO, USA, 2015; pp. 922–927. ISBN 9781455745555. [Google Scholar]
- Patel, M.B.; Garrad, E.; Meisel, J.W.; Negin, S.; Gokel, M.R.; Gokel, G.W. Synthetic ionophores as non-resistant antibiotic adjuvants. RSC Adv. 2019, 9, 2217–2230. [Google Scholar] [CrossRef]
- Rutkowski, J.; Brzezinski, B. Structures and Properties of Naturally Occurring Polyether Antibiotics. Biomed Res. Int. 2013, 2013, 1–31. [Google Scholar] [CrossRef]
- Shen, J.; Brodbelt, J.S. Characterization of ionophore–metal complexes by infrared multiphoton photodissociation and collision activated dissociation in a quadrupole ion trap mass spectrometer. Analyst 2000, 125, 641–650. [Google Scholar] [CrossRef]
- Sugarman, B. Zinc and infection. Rev. Infect. Dis. 1983, 5, 137–147. [Google Scholar] [CrossRef]
- Wątły, J.; Potocki, S.; Rowińska-Żyrek, M. Zinc Homeostasis at the Bacteria/Host Interface-From Coordination Chemistry to Nutritional Immunity. Chemistry 2016, 22, 15992–16010. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.-A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surf. A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Schwartz, J.R. Zinc Pyrithione: A Topical Antimicrobial with Complex Pharmaceutics. J. Drugs Dermatol. 2016, 15, 140–144. [Google Scholar] [PubMed]
- Blanchard, C.; Brooks, L.; Ebsworth-Mojica, K.; Didione, L.; Wucher, B.; Dewhurst, S.; Krysan, D.; Dunman, P.M.; Wozniak, R.A.F. Zinc Pyrithione Improves the Antibacterial Activity of Silver Sulfadiazine Ointment. mSphere 2016, 1, 322. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.E.; Harper, A.D.; Davison, E.K.; Jeong, J.Y.; Mros, S.; Harbison-Price, N.; van Zuylen, E.M.; Knottenbelt, M.K.; Heikal, A.; Ferguson, S.A.; et al. Substituted sulfonamide bioisosteres of 8-hydroxyquinoline as zinc-dependent antibacterial compounds. Bioorg. Med. Chem. Lett. 2020, 30, 127110. [Google Scholar] [CrossRef] [PubMed]
- Di Vaira, M.; Bazzicalupi, C.; Orioli, P.; Messori, L.; Bruni, B.; Zatta, P. Clioquinol, a drug for Alzheimer’s disease specifically interfering with brain metal metabolism: Structural characterization of its zinc(II) and copper(II) complexes. Inorg. Chem. 2004, 43, 3795–3797. [Google Scholar] [CrossRef]
- Alfonso, I.; Quesada, R. Biological activity of synthetic ionophores: Ion transporters as prospective drugs? Chem. Sci. 2013, 4, 3009. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, Y.; Lind, S.E.; Ding, W.-Q. Clioquinol targets zinc to lysosomes in human cancer cells. Biochem. J. 2009, 417, 133–139. [Google Scholar] [CrossRef]
- Mao, X.; Li, X.; Sprangers, R.; Wang, X.; Venugopal, A.; Wood, T.; Zhang, Y.; Kuntz, D.A.; Coe, E.; Trudel, S.; et al. Clioquinol inhibits the proteasome and displays preclinical activity in leukemia and myeloma. Leukemia 2009, 23, 585–590. [Google Scholar] [CrossRef]
- Ding, W.-Q.; Yu, H.-J.; Lind, S.E. Zinc-binding compounds induce cancer cell death via distinct modes of action. Cancer Lett. 2008, 271, 251–259. [Google Scholar] [CrossRef]
- Kavlock, R.J. Pesticides as Endocrine-Disrupting Chemicals. In Methods of Tissue Engineering; Atala, A., Lanza, R.P., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 727–746. ISBN 9780124262607. [Google Scholar]
- Bessho, R.; Matsubara, K.; Kubota, M.; Kuwakado, K.; Hirota, H.; Wakazono, Y.; Lin, Y.W.; Okuda, A.; Kawai, M.; Nishikomori, R.; et al. Pyrrolidine dithiocarbamate, a potent inhibitor of nuclear factor κB (NF-κB) activation, prevents apoptosis in human promyelocytic leukemia HL-60 cells and thymocytes. Biochem. Pharmacol. 1994, 48, 1883–1889. [Google Scholar] [CrossRef]
- Nguyen, M.; Vendier, L.; Stigliani, J.-L.; Meunier, B.; Robert, A. Structures of the Copper and Zinc Complexes of PBT2, a Chelating Agent Evaluated as Potential Drug for Neurodegenerative Diseases. Eur. J. Inorg. Chem. 2017, 2017, 600–608. [Google Scholar] [CrossRef]
- Adlard, P.A.; Cherny, R.A.; Finkelstein, D.I.; Gautier, E.; Robb, E.; Cortes, M.; Volitakis, I.; Liu, X.; Smith, J.P.; Perez, K.; et al. Rapid Restoration of Cognition in Alzheimer’s Transgenic Mice with 8-Hydroxy Quinoline Analogs Is Associated with Decreased Interstitial Aβ. Neuron 2008, 59, 43–55. [Google Scholar] [CrossRef]
- Lannfelt, L.; Blennow, K.; Zetterberg, H.; Batsman, S.; Ames, D.; Harrison, J.; Masters, C.L.; Targum, S.; Bush, A.I.; Murdoch, R.; et al. Safety, efficacy, and biomarker findings of PBT2 in targeting Aβ as a modifying therapy for Alzheimer’s disease: A phase IIa, double-blind, randomised, placebo-controlled trial. Lancet Neurol. 2008, 7, 779–786. [Google Scholar] [CrossRef]
- Harbison-Price, N.; Ferguson, S.A.; Heikal, A.; Taiaroa, G.; Hards, K.; Nakatani, Y.; Rennison, D.; Brimble, M.A.; El-Deeb, I.M.; Bohlmann, L.; et al. Multiple Bactericidal Mechanisms of the Zinc Ionophore PBT2. mSphere 2020, 5, 5649. [Google Scholar] [CrossRef] [PubMed]
- Walther, E.; Boldt, S.; Kage, H.; Lauterbach, T.; Martin, K.; Roth, M.; Hertweck, C.; Sauerbrei, A.; Schmidtke, M.; Nett, M. Zincophorin–biosynthesis in Streptomyces griseus and antibiotic properties. GMS Infect. Dis. 2016, 4, 1–9. [Google Scholar] [CrossRef]
- Farver, C.F.; Zander, D.S. Molecular Basis of Pulmonary Disease. Mol. Pathol. 2009, 417, 305–364. [Google Scholar] [CrossRef]
- Schoen, C.; Unzicker, C.; Stuhler, G.; Elias, J.; Einsele, H.; Grigoleit, G.U.; Abele-Horn, M.; Mielke, S. Life-Threatening Infection Caused by Daptomycin-Resistant Corynebacterium jeikeium in a Neutropenic Patient. J. Clin. Microbiol. 2009, 47, 2328–2331. [Google Scholar] [CrossRef]
- Mochizuki, T.; Satsu, H.; Shimizu, M. Signaling pathways involved in tumor necrosis factor alpha-induced upregulation of the taurine transporter in Caco-2 cells. FEBS Lett. 2005, 579, 3069–3074. [Google Scholar] [CrossRef]
- Sharma, P.K.; Kumar, S.; Kumar, P.; Kaushik, P.; Kaushik, D.; Dhingra, Y.; Aneja, K.R. Synthesis and biological evaluation of some pyrazolylpyrazolines as anti-inflammatory–antimicrobial agents. Eur. J. Med. Chem. 2010, 45, 2650–2655. [Google Scholar] [CrossRef]
- Kang, M.-S.; Choi, E.-K.; Choi, D.-H.; Ryu, S.-Y.; Lee, H.-H.; Kang, H.-C.; Koh, J.-T.; Kim, O.-S.; Hwang, Y.-C.; Yoon, S.-J.; et al. Antibacterial activity of pyrrolidine dithiocarbamate. FEMS Microbiol. Lett. 2008, 280, 250–254. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
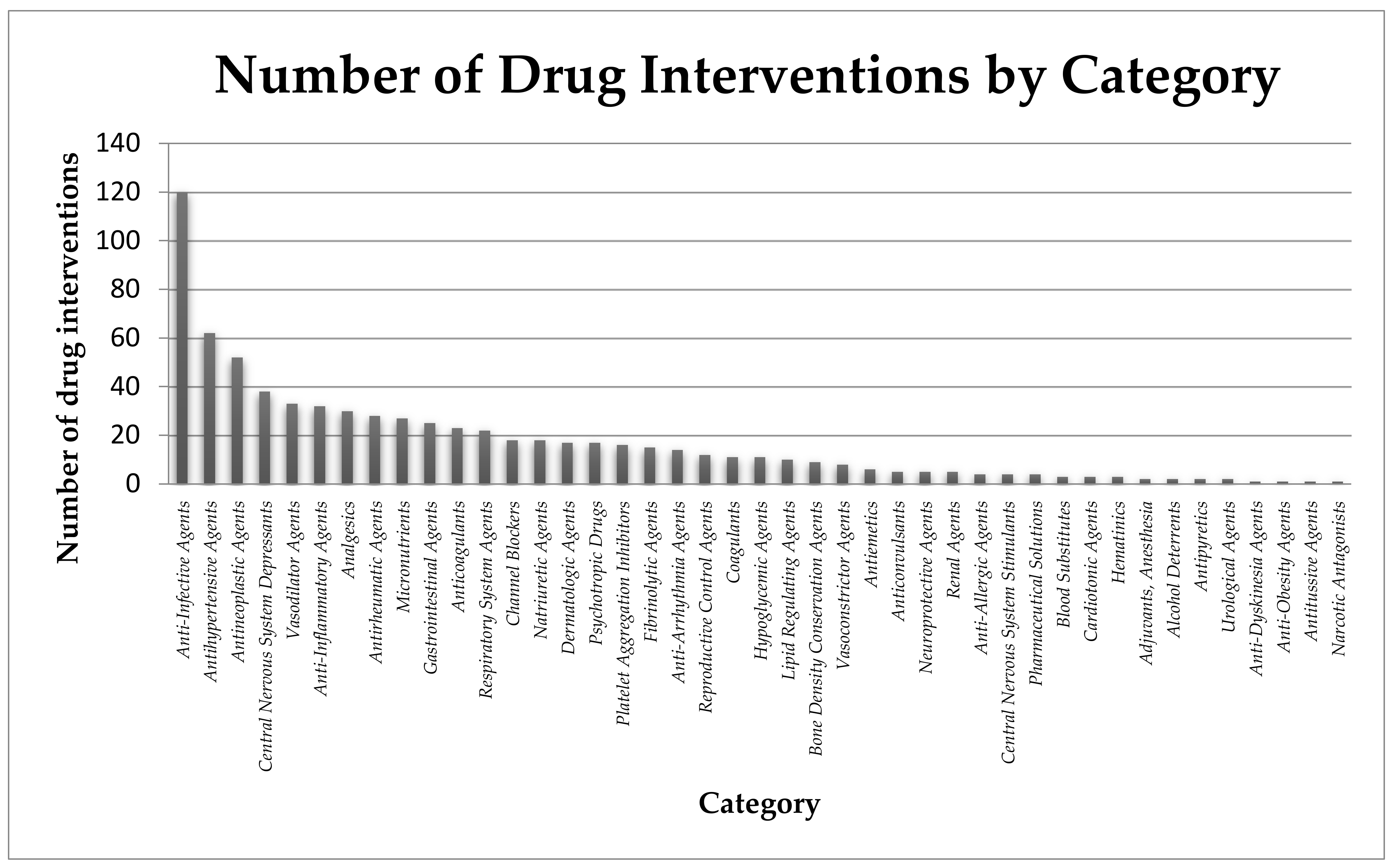
| Main Drug | Other Interventions | Clinical Trial Identifier | Duration | Location | Purpose | Estimated Enrollment | Inclusion Criteria: | Ref. |
|---|---|---|---|---|---|---|---|---|
| Hydroxychloroquine | Vitamin C, vitamin D, zinc(II) | NCT04335084 | 24 weeks | Ventura, CA, USA | prevention | 600 participants, medical workers who are exposed to COVID-19 | Patients >18 years old without COVID-19 symptoms; male or female that are considered to be high-risk individuals | [69] |
| Hydroxy-chloroquine (Plaquenil) (200 mg) | Vitamin C, vitamin D, zinc(II) | NCT04326725 | 16 weeks | Istanbul, Turkey | prevention | 80 participants, healthcare professionals and their first degree relatives | Patients aged between 20 and 90 years without COVID-19 symptoms; male or female | [70] |
| Hydroxy-chloroquine (Plaquenil) | Azithromycin (Zithromax), vitamins C and D Zinc(II) | NCT04482686 | 24 weeks | Ventura, CA, USA | treatment | 600 participants, patients with COVID-19 infection | Patients aged between 18 and 75 years; healthy male or female with positive test for COVID-19; diabetic and obese (BMI > 30) patients will be included in the trial but randomization will be stratified | [71] |
| - | Oral nutrition supplement (ONS): protein (14800 mg), fat (22200 mg), carbohydrate (25000 mg), EPA (1100 mg), DHA (450 mg), GLA (950 mg), vitamins A (2840 IU), C (205 mg), and E (75 IU), selenium (0.018 mg), zinc(II) (5.7 mg) | NCT04323228 | 12 weeks | King Saud University Saudi Arabia | supportive care | 30 participants patients with COVID-19 infection | Patients aged between 18 and 65 years; male or female in stable condition (i.e., not requiringICU admission) | [72] |
| - | Vitamin C (8000 mg) Zinc(II) Gluconate (50 mg) | NCT04342728 | 4 weeks | Cleveland, OH, USA | supportive care | 520 patients diagnosed with COVID-19 | Male or female patients >18 years old with COVID-19 diagnosed in an outpatient setting with any symptom (fever or chills, shortness of breath or difficulty breathing, cough, fatigue, muscle/body aches, headache, new loss of taste, new loss of smell, congestion or runny nose, nausea, vomiting, diarrhea | [73] |
| Hydroxy-chloroquine (400 mg, 200 mg)/azithromycin (500 mg, 250 mg) hydroxy-chloroquine (400 mg, 200 mg)/doxycycline (200 mg) | Zinc(II) (220 mg) | NCT04370782 | 8 weeks | Roslyn, NY, USA | treatment | 750 participants with COVID-19 infection | Patients 30 years and older; male or female with high initial clinical suspicion by physician based on signs and symptoms (fever, cough, myalgias, fatigue, shortness of breath) | [74] |
| Hydroxy-chloroquine (400 mg) | Zinc(II) (15 mg) | NCT04377646 | 8 weeks | Tunis, Tunisia | prevention | 660 participants military health professionals exposed to SARS-CoV-2 | Patients aged between 18 and 65 years; male or female without COVID-19 symptoms and with negative SARS-CoV-2 diagnostic test; no self-medication with chloroquine, hydroxychloroquine or antivirals | [75] |
| - | Vitamin D3 (2000 IU) Zinc(II) (30 mg) | NCT04351490 | 8 weeks | Lille, France | treatment | 3140 participants, 60 years and older patients infected with COVID-19 | Patients 60 years and older; male or female; institutionalized | [76] |
| Hydroxy-chloroquine, azithromycin, favipiravir (1800 mg, 800 mg) | Zinc(II) | NCT04373733 | 4 weeks | London, UK | treatment | 450 participants infected with COVID-19 | Male or female patients > 18 years old with suspected or confirmed COVID-19 infection (fever ≥37.8 °C and at least one of the following respiratory symptoms, which must be of acute onset: persistent cough, hoarseness, nasal discharge or congestion, shortness of breath, sore throat, wheezing or sneezing) | [77] |
| Chloroquine | Zinc(II) | NCT04447534 | 2 weeks | Tanta, Egypt | treatment | 200 participants infected with COVID-19 | Patients 18 years and older with positive COVID-19 (inclusion/exclusion criteria not very precise; expected treatment outcome difficult to predict) | [78] |
| Hydroxy-chloroquine (400 mg, 200 mg), azithromycin (500 mg, 250 mg) | Vitamin C (400 mg/kg/day, 200 mg/kg/day Zinc(II) (30 mg) Vitamin D3 (5000 IU) Vitamin B12 (0.5 mg) | NCT04395768 | 18 months | Melbourne Australia | treatment | 200 participants infected with COVID-19 | Patients who were > 18 years old; male or female with active COVID-19 infection | [79] |
| Hydroxy-chloroquine (800 mg, 400 mg) | Zinc(II) (66 mg) | NCT04384458 | 7 weeks | Fortaleza, Ceará, Brazil | prevention | 400 healthcare workers exposed to SARSCoV-2 | Patients aged between 18 and 70 years; male or female which works in a healthcare facility delivering direct care to patients with either proven or suspected COVID-19 | [80] |
| Hydroxy-chloroquine (400 mg; 200 mg) | Ivermectin (12 mg) Povidone-Iodine Zinc(II) (80 mg) Vitamin C (500 mg) | NCT04446104 | 6 weeks | Singapore | prevention | 5000 participants, migrant workers at High-risk of COVID-19 | Patients aged between 21 and 60 years; male with weight more than 40 kg | [81] |
| Nitazoxanide, ribavirin, ivermectin | Zinc(II) | NCT04392427 | 2 years | MansouraEgypt | treatment | 100 participants infected with COVID-19 | Patients 12 years and older; male or female with the COVID-19 PCR positive test and referred to the quarantine; patient without any comorbidities and no sensitivity or contraindication to the three drugs | [82] |
| Hydroxy-chloroquine (200 mg) | Azithromycin (500 mg, 250 mg) Zinc(II) (220 mg) | NCT04412746 | 3 months | Algeria | treatment | 100 participants with diabetes and infected with COVID-19 | Patients 16 years and older; male or female; hospitalized patients with diabetes known before the admission; new onset diabetes discovered at admission | [83] |
| Quercetin (500 mg) | Bromelain (500 mg) Zinc(II) (50 mg) Vitamin C (1000 mg) | NCT04468139 | 4 weeks | Saudi Arabia | treatment | 60 participants infected with COVID-19 | Male or female patients aged ≥ 18 years with moderate to severe disease (inclusion/exclusion criteria not very precise) | [84] |
| Ivermectin (0.2 mg/kg) | Nigella Sativa (80 mg/kg) Zinc(II) (20 mg) | NCT04472585 | 4 weeks | Punjab, Pakistan | treatment | 40 participants infected with COVID-19 | Male of female patients > 18 years old with a positive SARS-CoV-2 PCR test; patients with mild to moderate disease | [85] |
| Hydroxy-chloroquine | Vitamin C (2000 mg) Vitamin D (Alfacalcidiol 0.001 mg) Zinc(II) (50 mg) Paracetamol | NCT04491994 | 3 weeks | Punjab, Pakistan | treatment | 500 participants infected with COVID-19 | Patients aged between 18 and 80 years; male or female, with mild disease | [86] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hecel, A.; Ostrowska, M.; Stokowa-Sołtys, K.; Wątły, J.; Dudek, D.; Miller, A.; Potocki, S.; Matera-Witkiewicz, A.; Dominguez-Martin, A.; Kozłowski, H.; et al. Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19? Pharmaceuticals 2020, 13, 228. https://doi.org/10.3390/ph13090228
Hecel A, Ostrowska M, Stokowa-Sołtys K, Wątły J, Dudek D, Miller A, Potocki S, Matera-Witkiewicz A, Dominguez-Martin A, Kozłowski H, et al. Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19? Pharmaceuticals. 2020; 13(9):228. https://doi.org/10.3390/ph13090228
Chicago/Turabian StyleHecel, Aleksandra, Małgorzata Ostrowska, Kamila Stokowa-Sołtys, Joanna Wątły, Dorota Dudek, Adriana Miller, Sławomir Potocki, Agnieszka Matera-Witkiewicz, Alicia Dominguez-Martin, Henryk Kozłowski, and et al. 2020. "Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19?" Pharmaceuticals 13, no. 9: 228. https://doi.org/10.3390/ph13090228
APA StyleHecel, A., Ostrowska, M., Stokowa-Sołtys, K., Wątły, J., Dudek, D., Miller, A., Potocki, S., Matera-Witkiewicz, A., Dominguez-Martin, A., Kozłowski, H., & Rowińska-Żyrek, M. (2020). Zinc(II)—The Overlooked Éminence Grise of Chloroquine’s Fight against COVID-19? Pharmaceuticals, 13(9), 228. https://doi.org/10.3390/ph13090228