
Dynamics of Lingulodinium polyedra Development in the Bulgarian Part of Black Sea (1992–2022)

 ,
,  ,
,  and
and 
Abstract
1. Introduction
1.1. Taxonomy of Lingulodinium polyedra
1.2. Morphology of L. polyedra
1.3. Ecology of L. polyedra
1.4. Harmful Algal Blooms (HABs) of L. polyedra Around the World
1.5. Distribution and Blooms of L. polyedra in the Black Sea
- A low-productive ecosystem before 1970 or the so-called “pristine” reference phase
- A highly productive eutrophic system during the 1980s or a phase of intensive anthropogenic eutrophication and destruction of the Black Sea ecosystem
- A transitional, relatively low productive system after the beginning of the 1990s or the so-called post-eutrophication phase in the development of the Black Sea ecosystem” [21] (p. 2).
1.6. Toxins Produced by L. polyedra
2. Materials and Methods
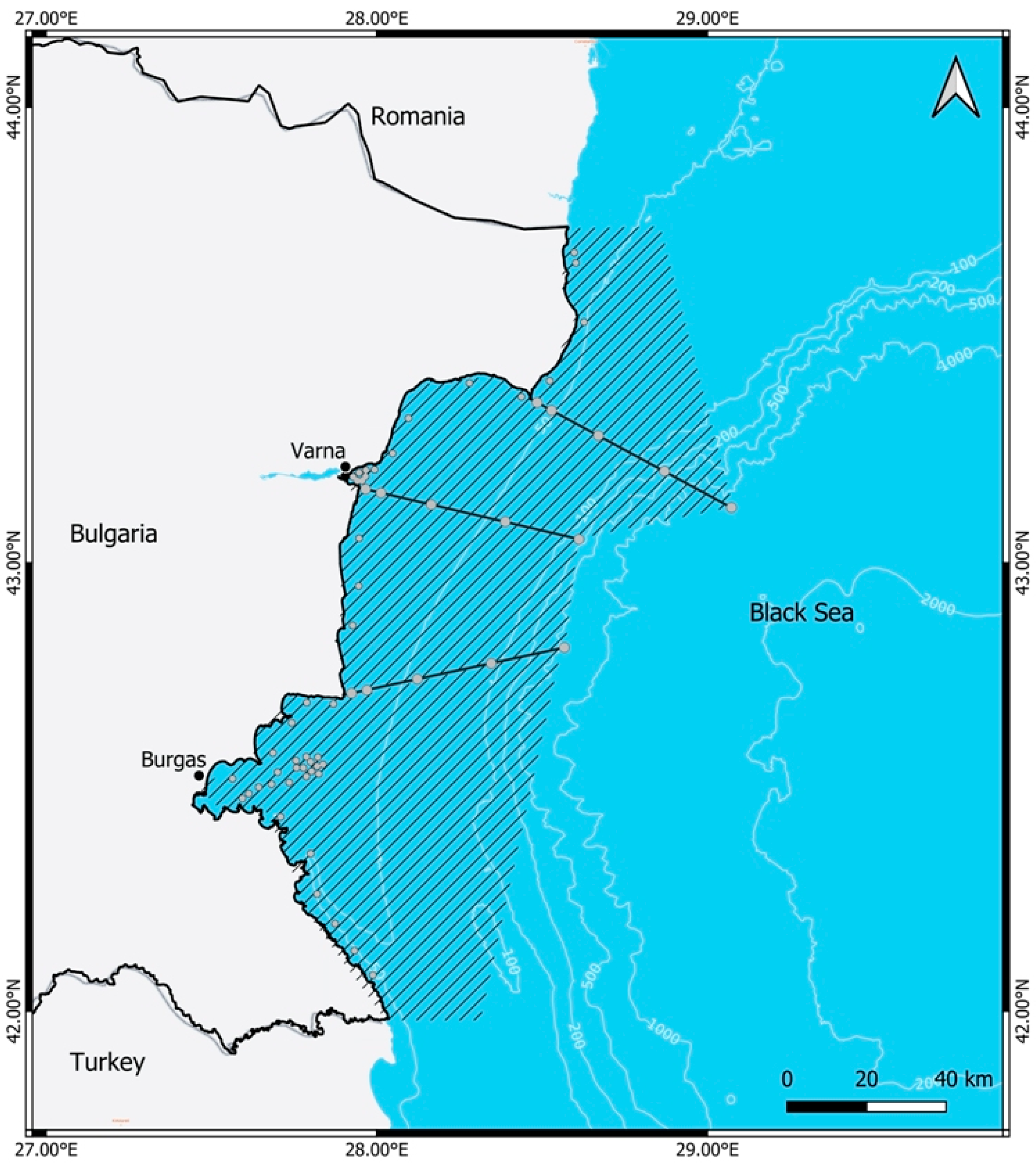
2.1. Study Area
2.2. Field Sampling
2.3. Sample Handling in the Laboratory
2.4. Climate Data
2.5. Studying the Dependence of L. polyedra on Sea Surface Temperature (SST) and North Atlantic Oscillation Indices (NAOI)
3. Results
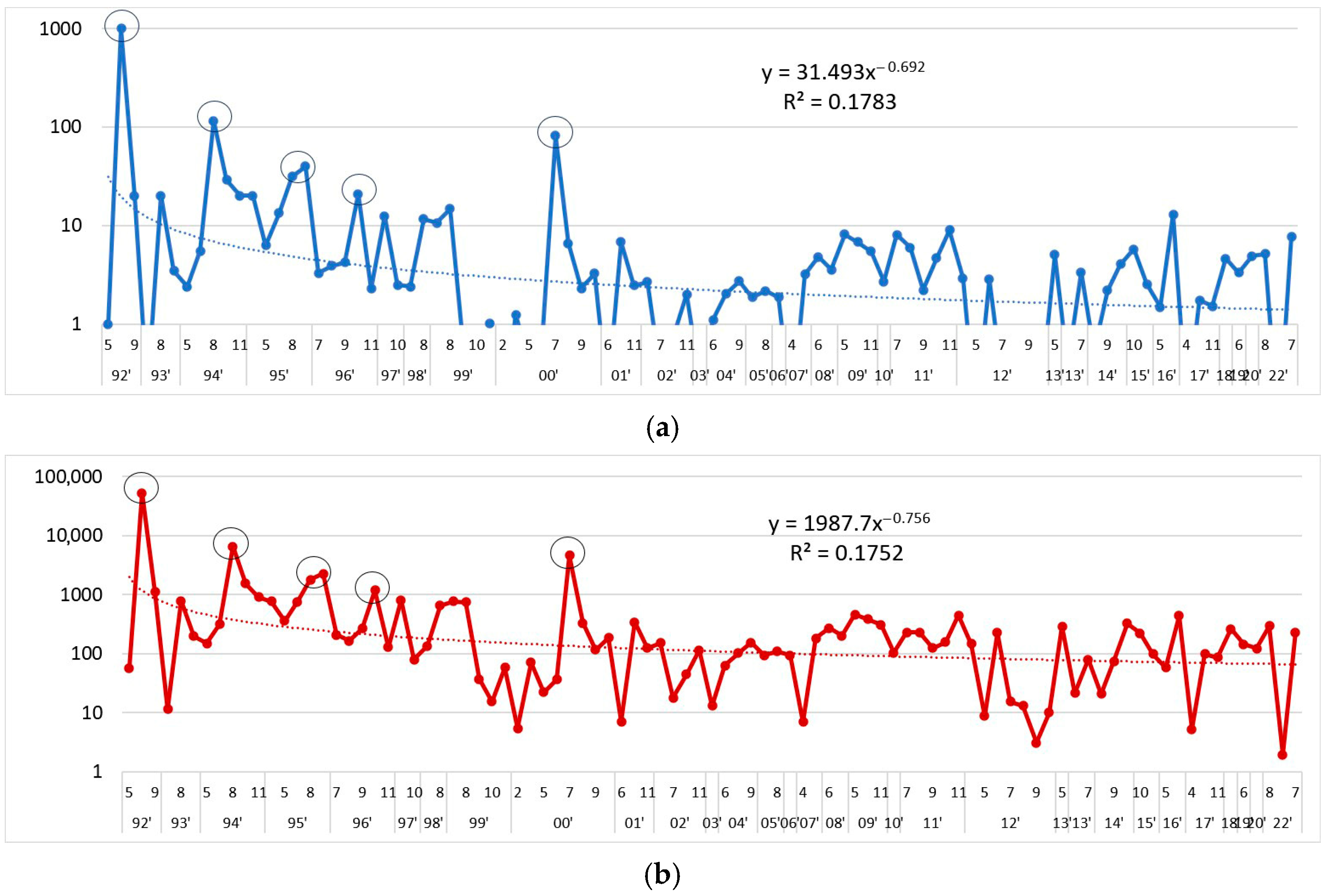
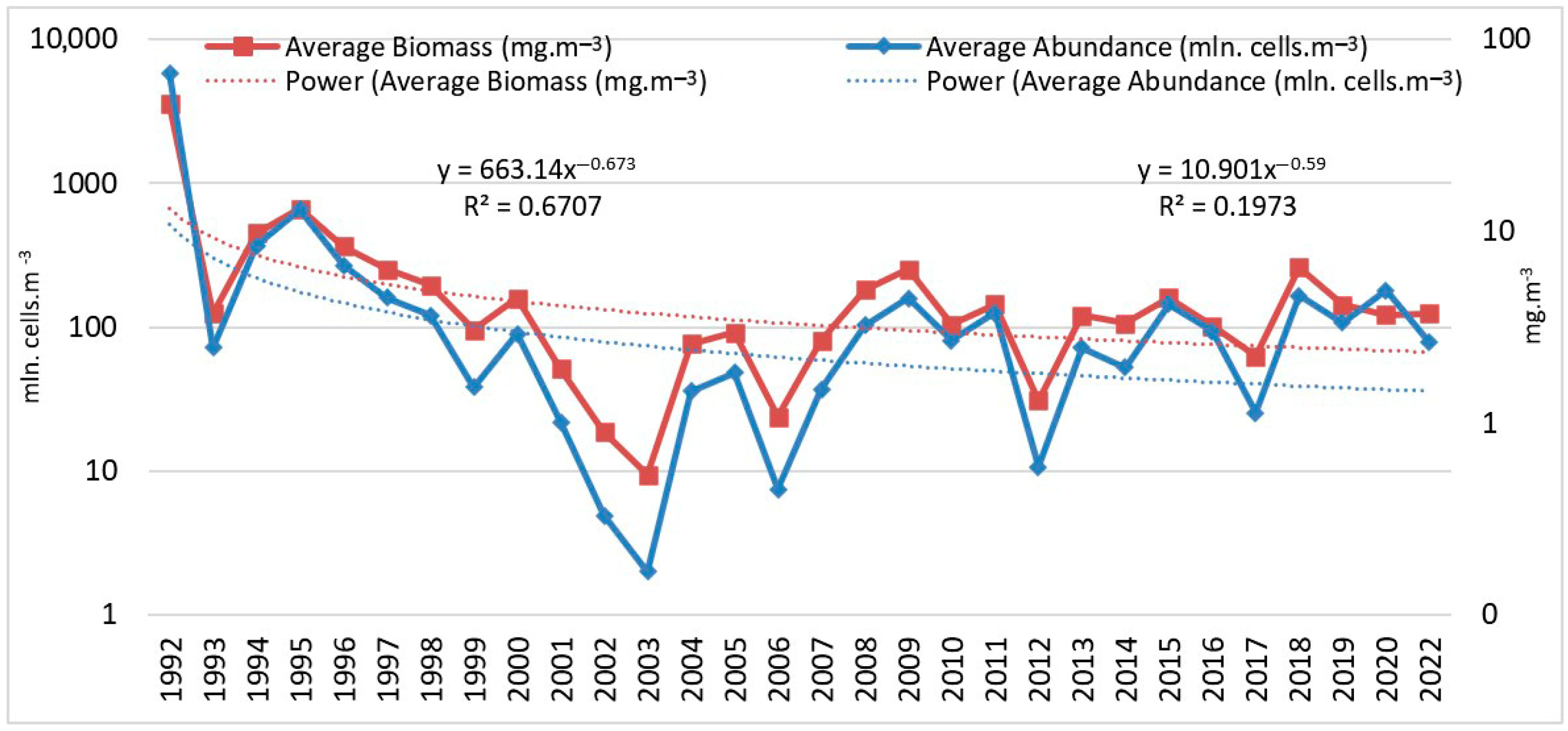
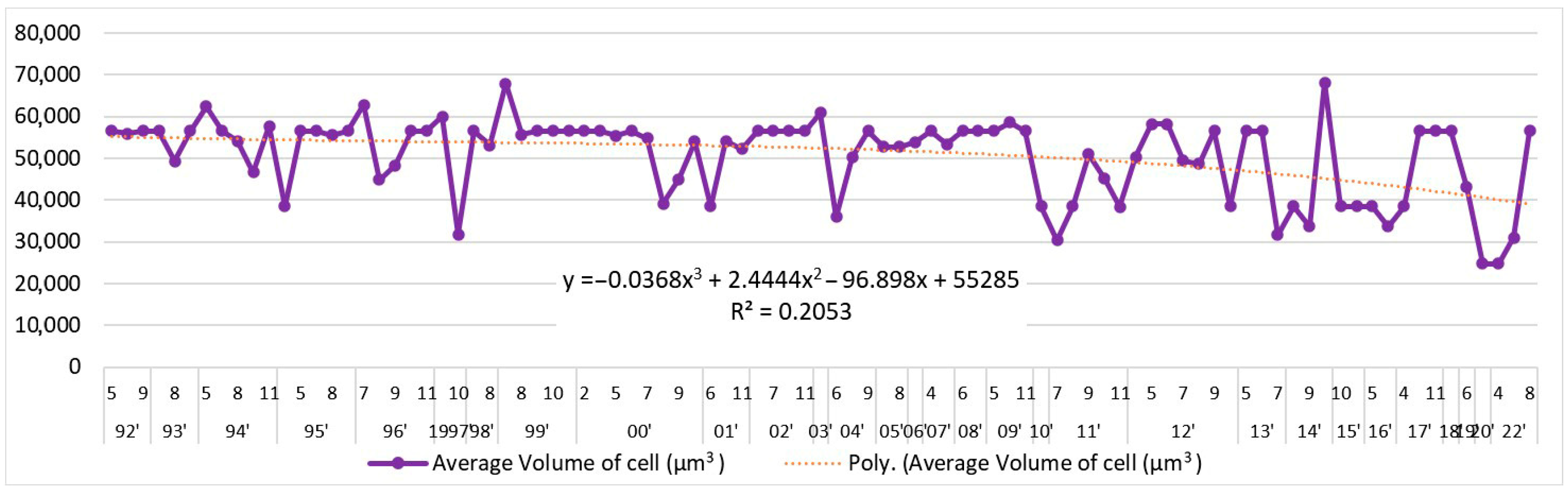
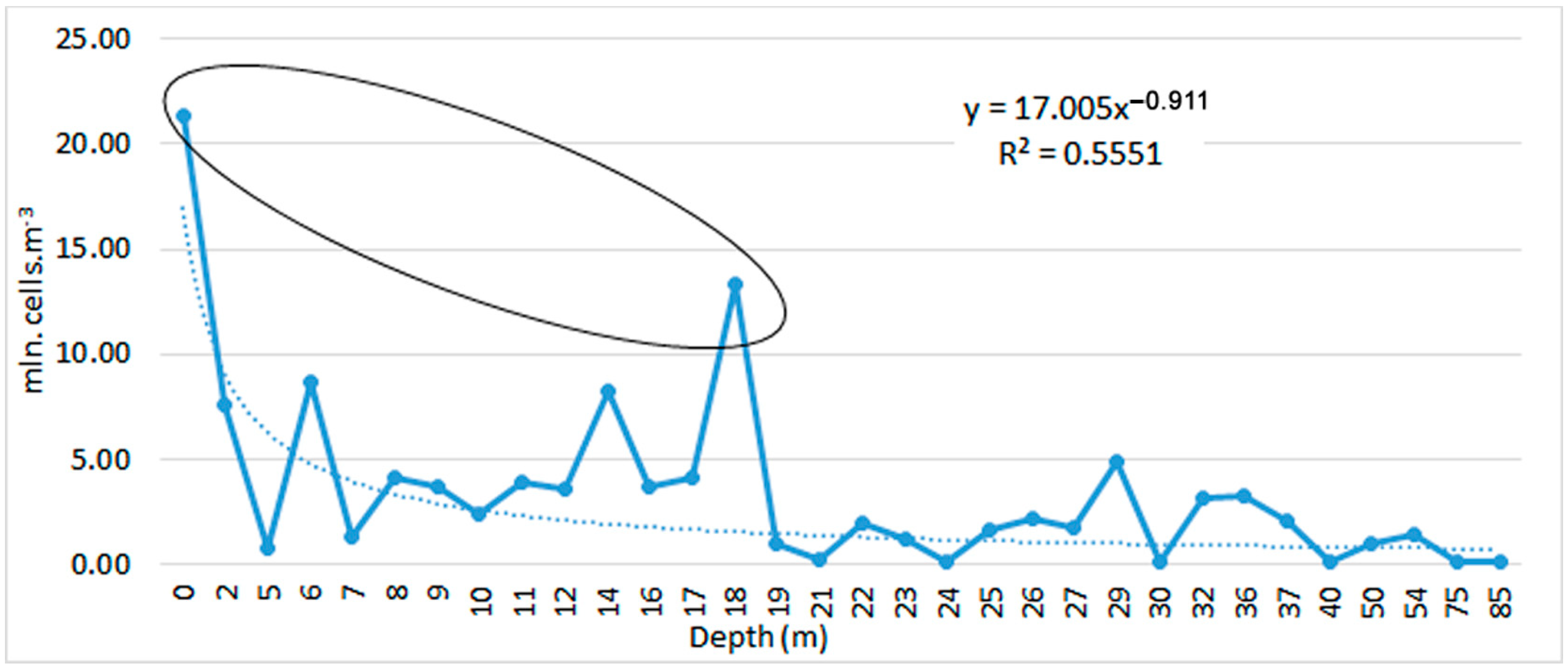
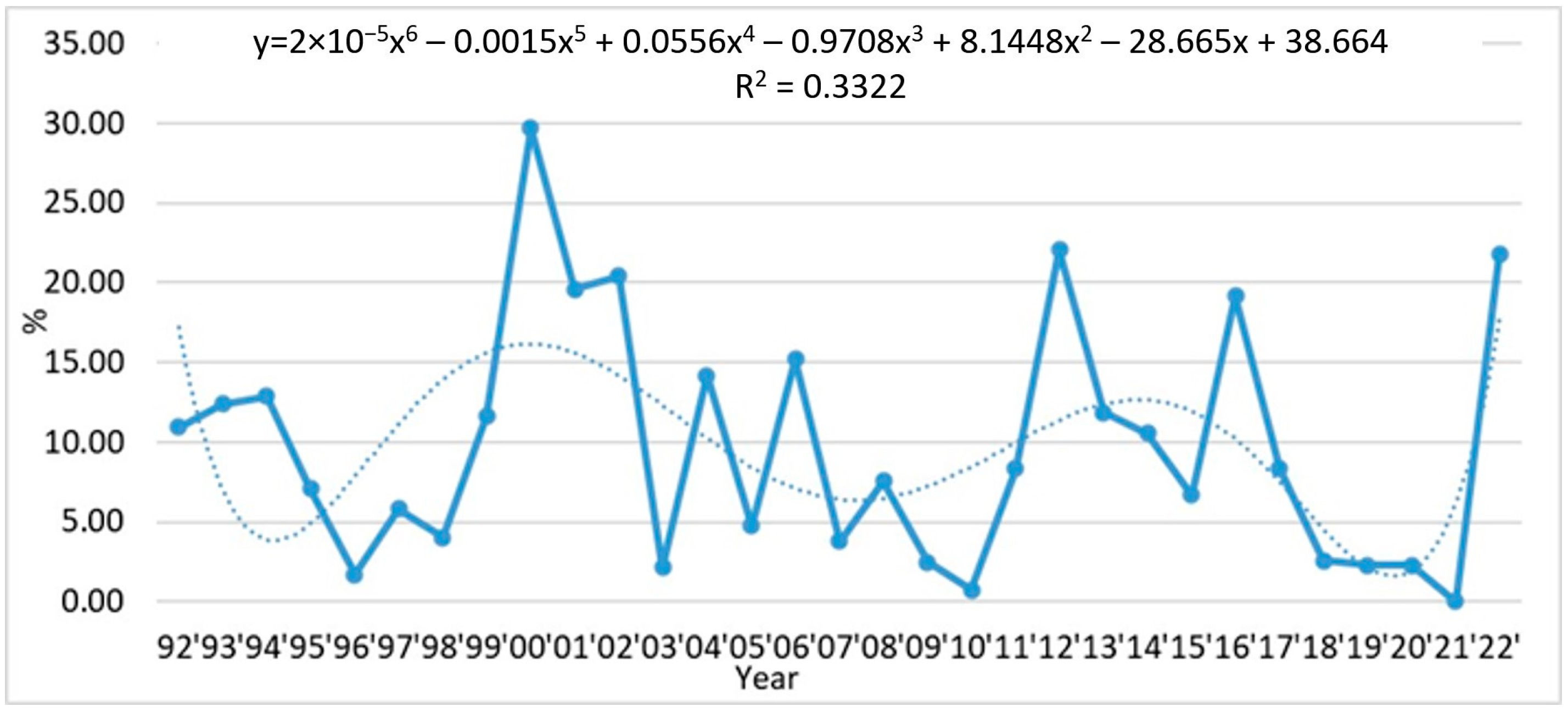
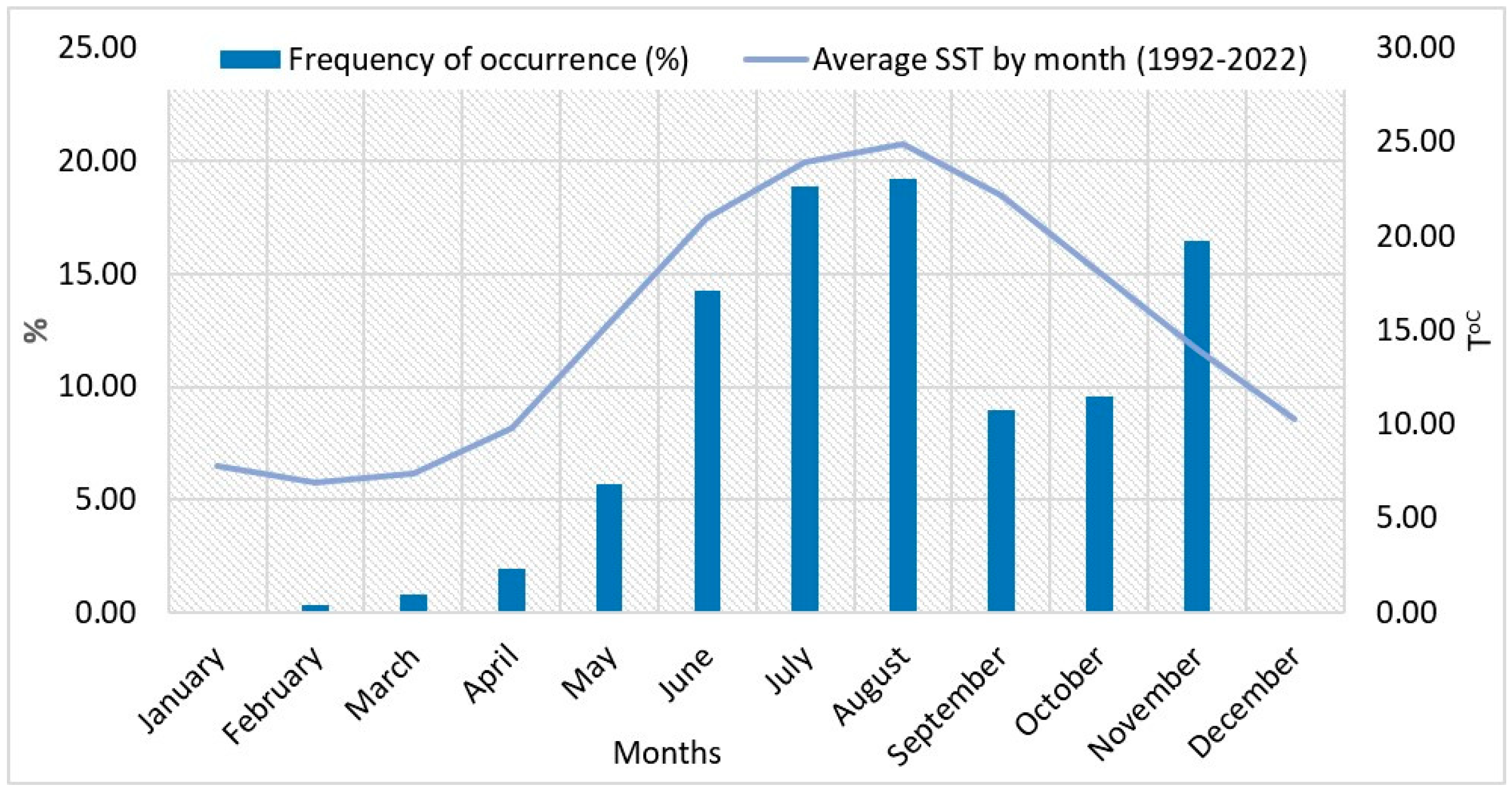
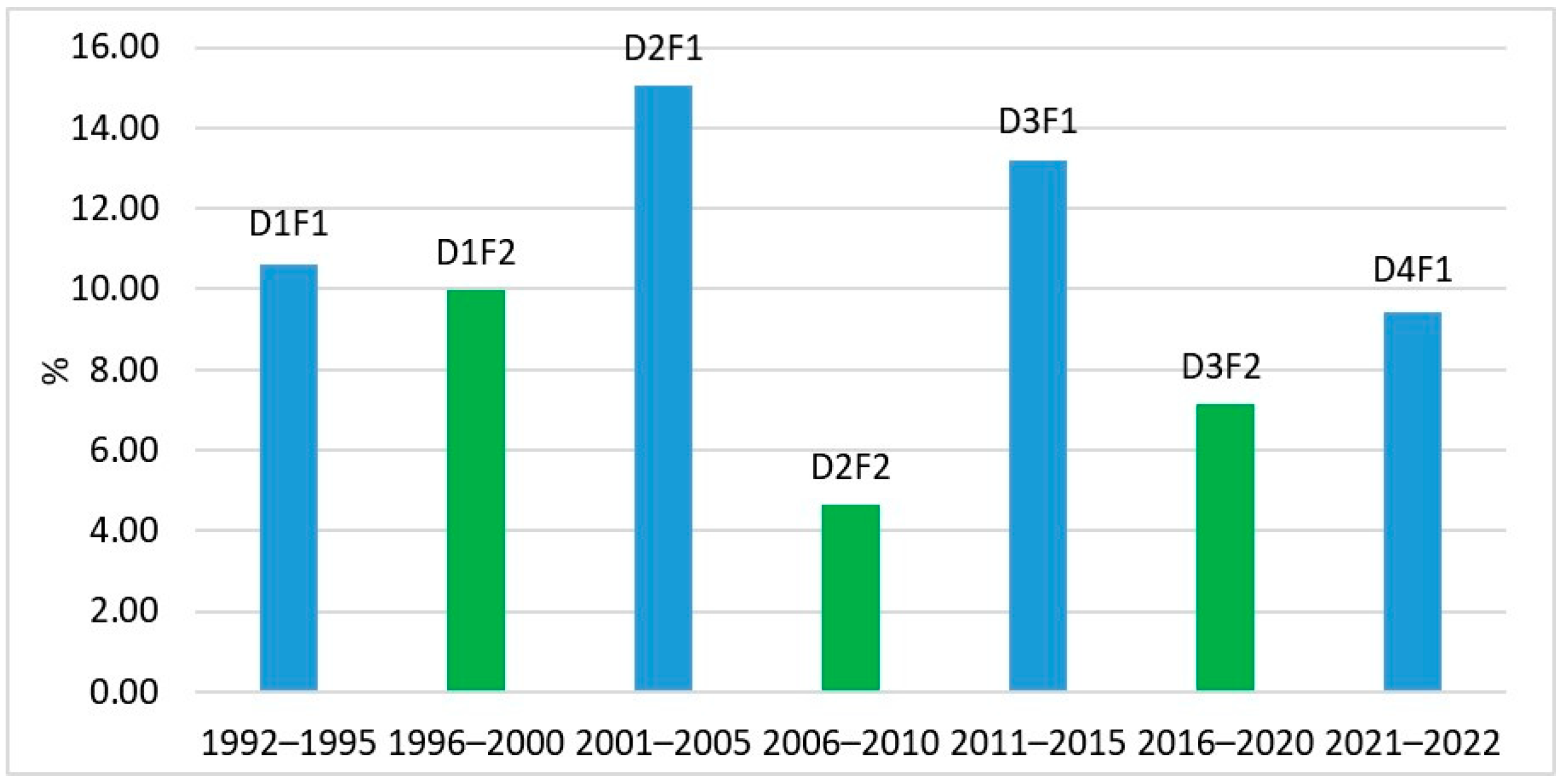
3.1. Tendencies in the Changes of L. polyedra Abundance, Biomass, Volume of Cells and Frequency of Occurrence in the Bulgarian Black Sea Waters
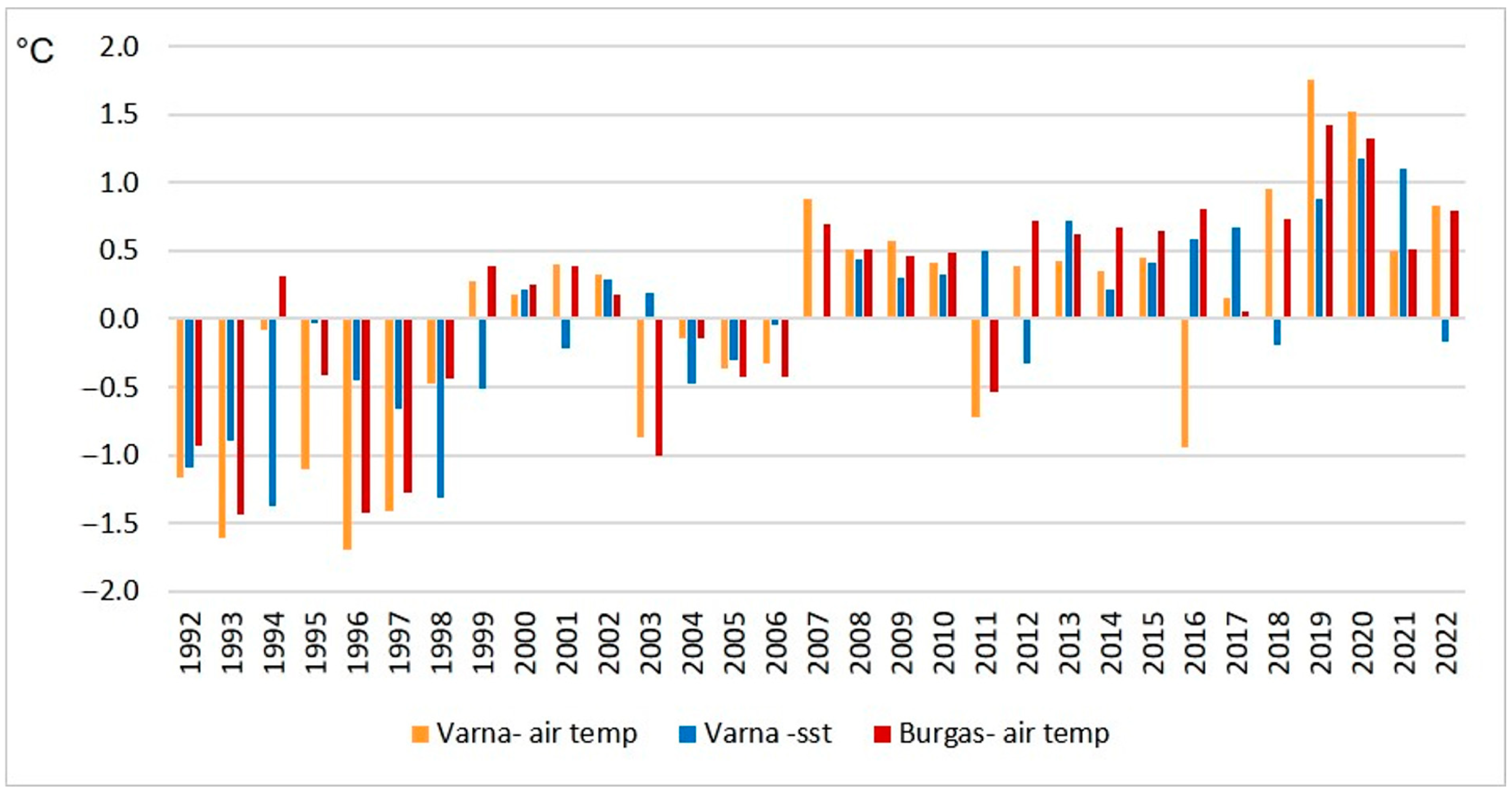
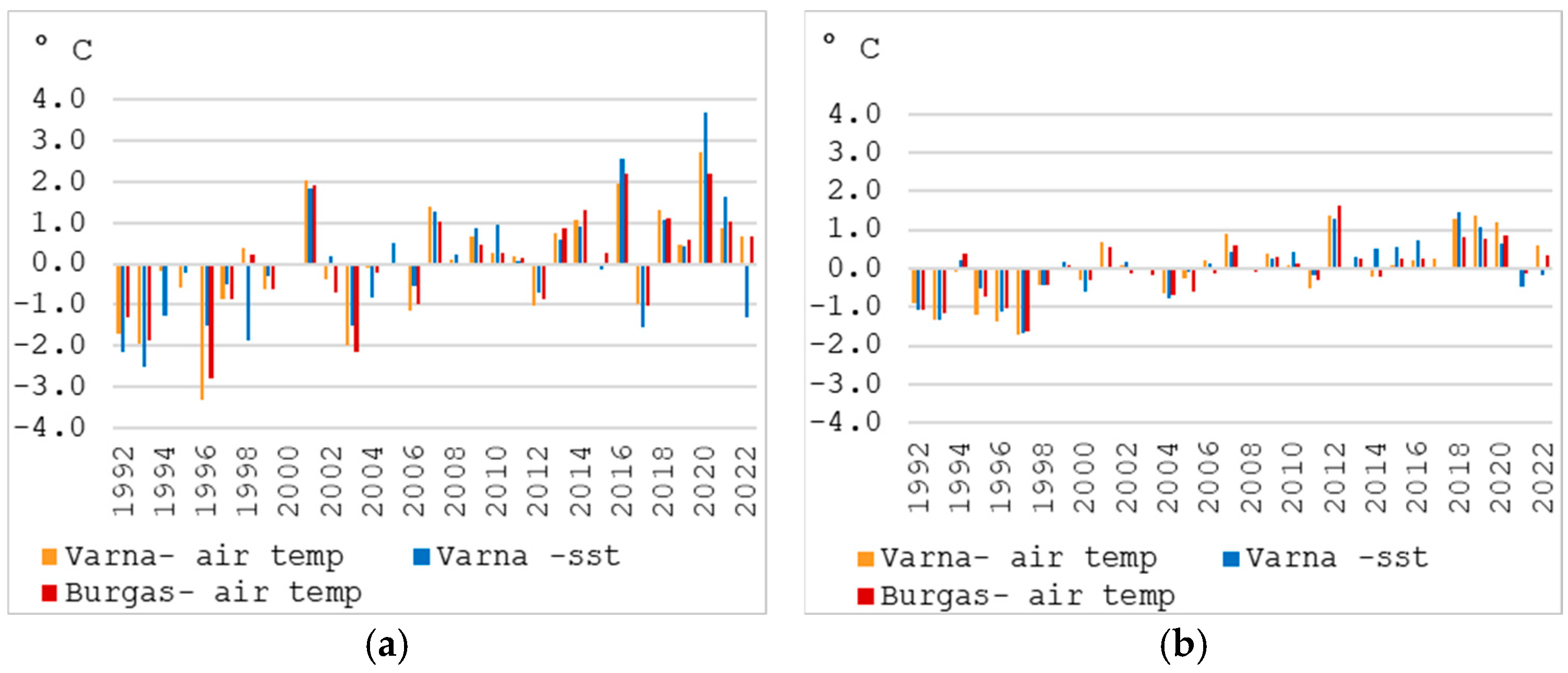
3.2. Climate Characteristics and Observed Changes in Air and Sea Surface Temperature (1992–2022)
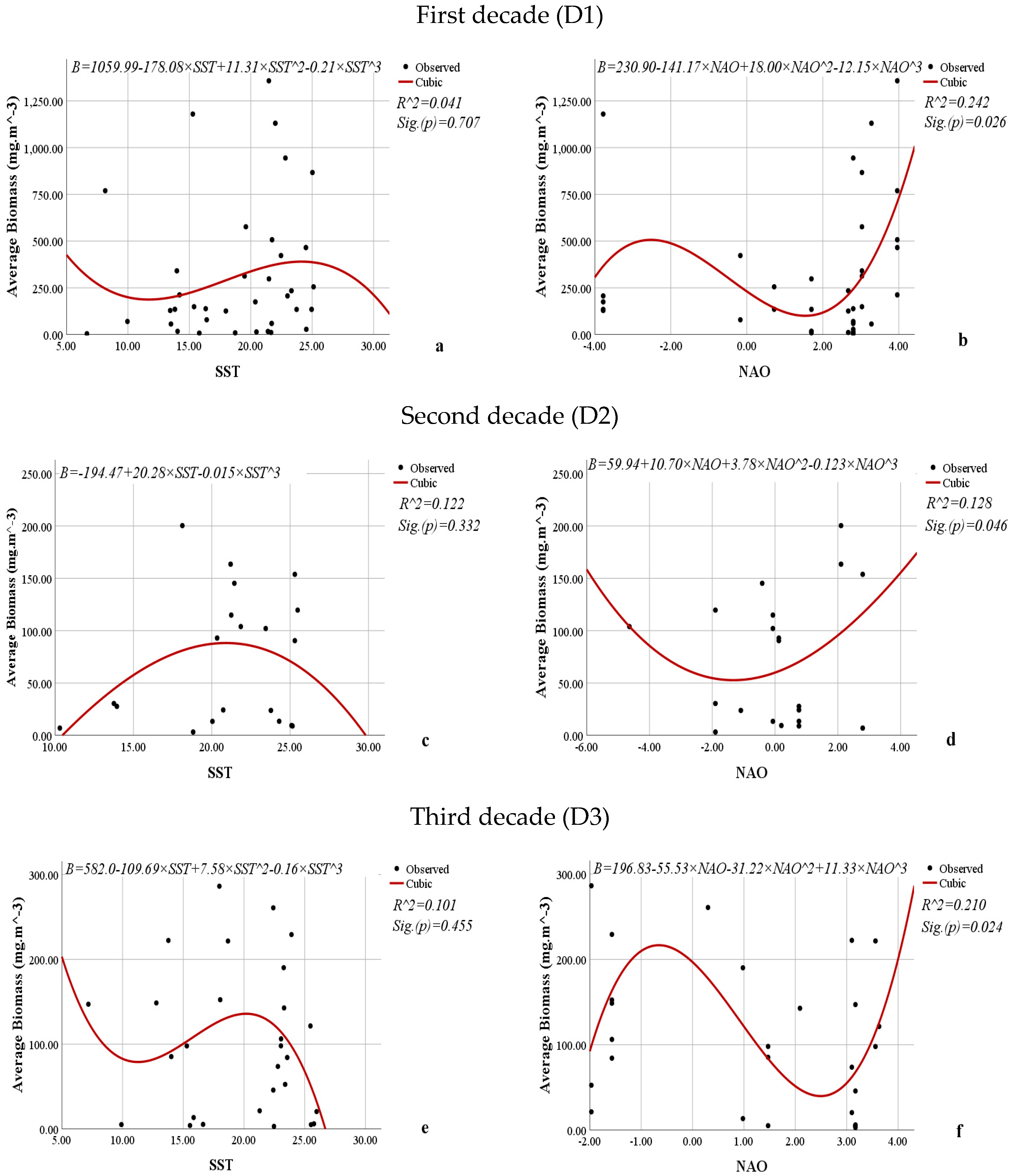
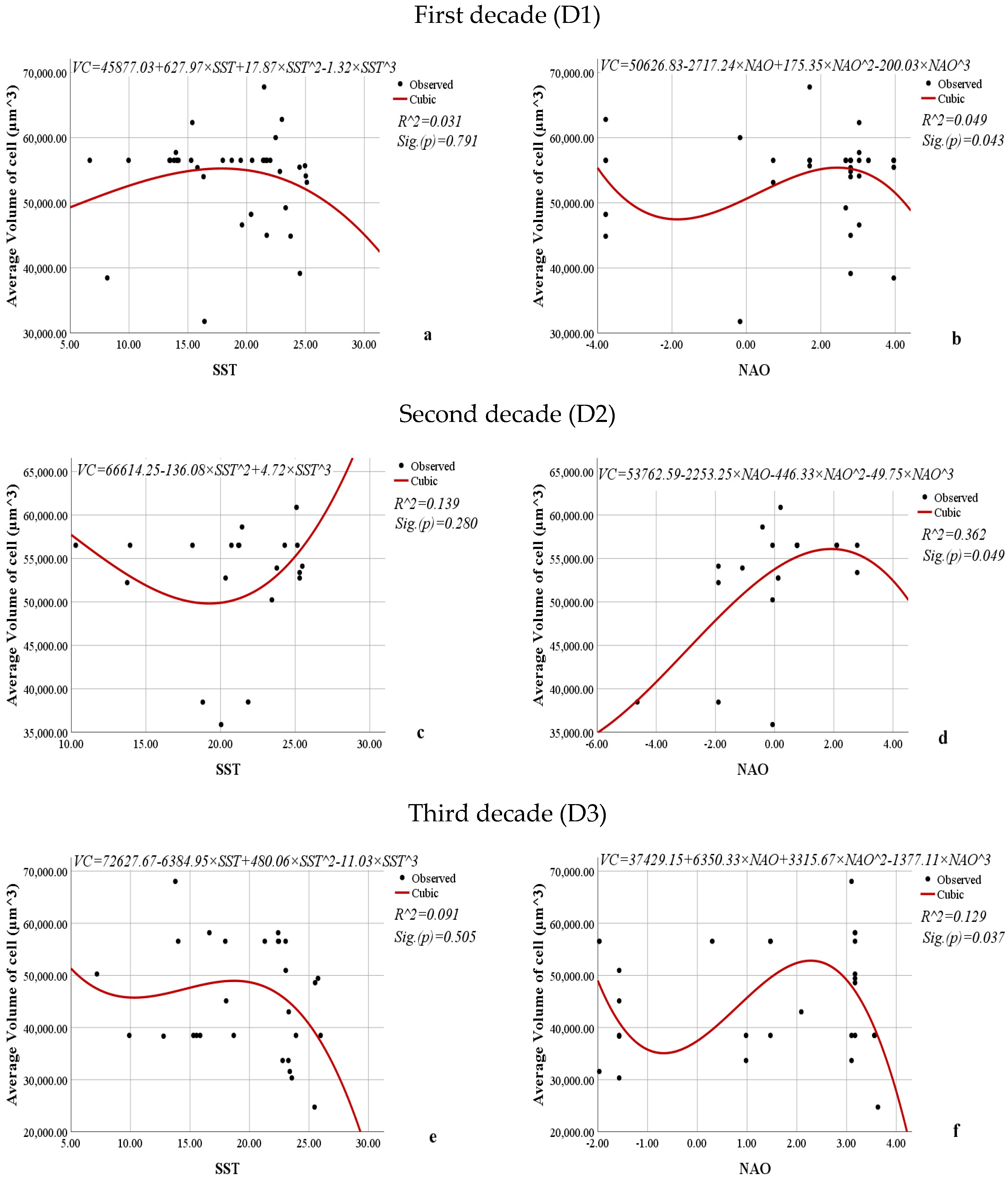
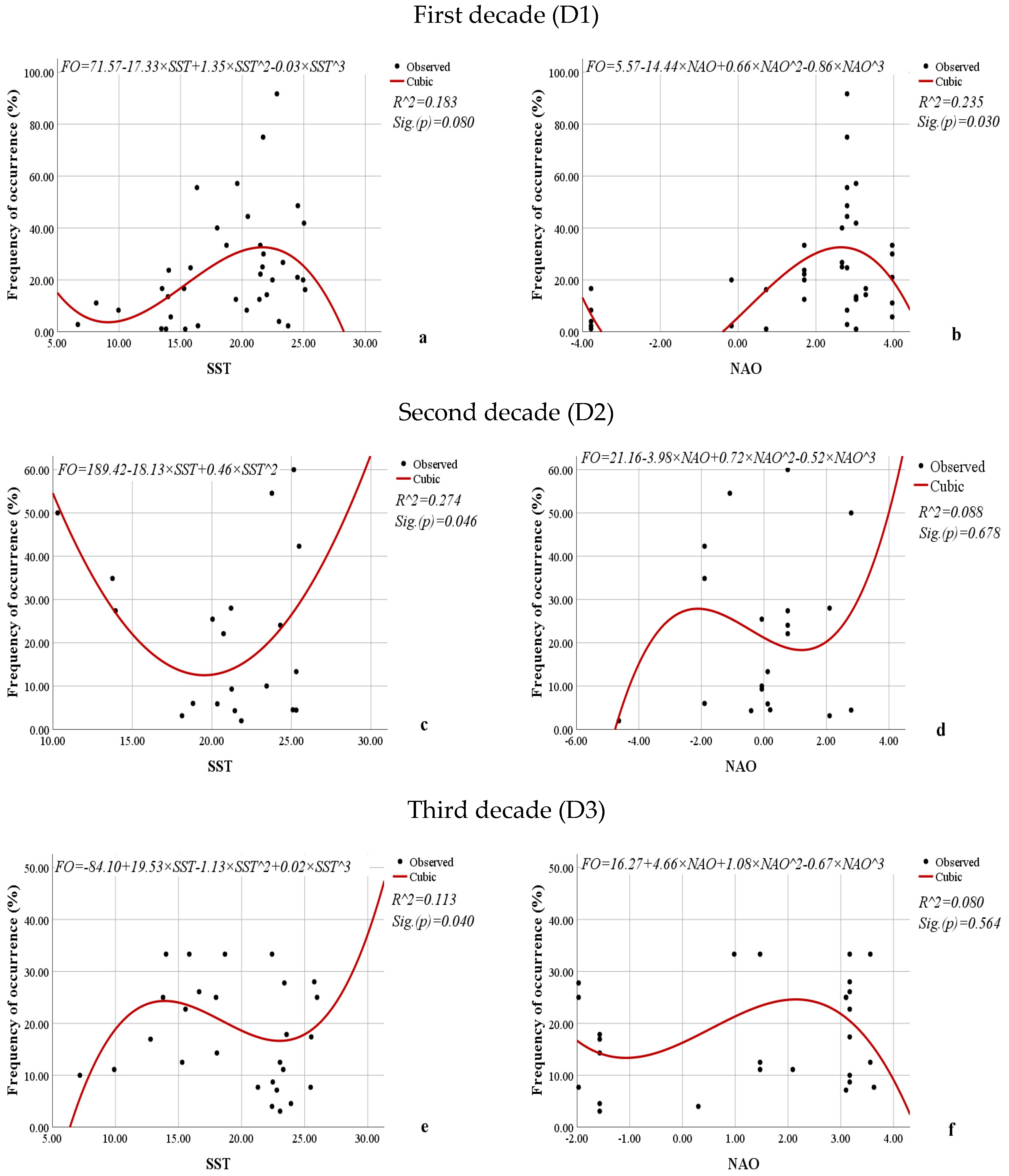
3.3. Relationship Between the Average Abundance (mln. cells·m−3), Biomass (mg·m−3), Volume of Cells (µm3), Frequency of Occurrence (%) of L. polyedra, and the Sea Surface Temperature and North Atlantic Oscillation Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guiry, M.D.; Guiry, G.M. World-Wide Electronic Publication, National University of Ireland, Galway. AlgaeBase. Available online: http://www.algaebase.org (accessed on 12 December 2024).
- Petrova, V.J. Composition and quantitative distribution of phytoplankton in Varna Bay. Proc. Bot. Inst. BAS 1960, 7, 247–277. (In Bulgarian) [Google Scholar]
- Petrova, V.J. The phytoplankton of Lake Varna. Proc. Cent. Sci. Res. Inst. Fish Resour.-Varna 1961, 1, 183–220. (In Bulgarian) [Google Scholar]
- Faust, M.; Gulledge, R. Identifying Harmful Marine Dinoflagellates. Contrib. U. S. Natl. Herb. 2002, 42, 1–144. Available online: http://www.jstor.org/stable/23493225 (accessed on 8 July 2024).
- Kofoid, C.A. Dinoflagellata of the San Diego region, IV. The genus Gonyaulax, with notes on its skeletal morphology and a discussion of its generic and specific characters. Univ. Cal. Publ. Zool. 1911, 8, 187–269. [Google Scholar]
- Kiselev, I.A. Dinoflagellata of the Seas and Fresh Waters of the USSR. Determinants of Fauna of the USSR; Zoological Institute of the Academy of Sciences of the USSR: Moscow, Russia, 1950; pp. 1–280. (In Russian) [Google Scholar]
- Nielsen, L.T.; Kiørboe, T. Feeding currents facilitate a mixotrophic way of life. ISME J. 2015, 9, 2117–2127. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Taylor, F.J.R. The Biology of Dinoflagellates; Blackwell: Oxford, UK, 1987; pp. 1–785. [Google Scholar]
- Prego, R.; Bao, R.; Varela, M.; Carballeira, R. Naturally and Anthropogenically Induced Lingulodinium polyedra Dinoflagellate Red Tides in the Galician Rias (NW Iberian Pterenkoeninsula). Toxins 2024, 16, 280. [Google Scholar] [CrossRef] [PubMed]
- Lundholm, N.; Churro, C.; Escalera, L.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Larsen, J.; Mertens, K.; Moestrup, Ø.; Murray, S.; et al. (Eds.) (2009 Onwards). IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. Lingulodinium polyedra (F. Stein) J.D. Dodge, 1989. Available online: https://www.marinespecies.org/hab/aphia.php?p=taxdetails&id=233592 (accessed on 29 October 2024).
- Amorim, A.; Palma, A.S.; Sampayo, M.A.; Moita, M.T. On a Lingulodinium polyedrum Bloom in Setúbal Bay, Portugal. In Harmful Algal Blooms 2000; Hallegraef, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Hobart, Australia, 2001; pp. 133–136. [Google Scholar]
- Meave del Castillo, M.E.; Zamudio-Resendiz, M.E. Planktonic algal blooms from 2000 to 2015 in Acapulco Bay, Guerrero, Mexico. Acta Bot. Mex. 2018, 125, 61–93. [Google Scholar] [CrossRef]
- Mertens, K.N.; Retho, M.; Manach, S.; Zoffoli, M.L.; Doner, A.; Schapira, M.; Bilien, G.; Séchet, V.; Lacour, T.; Robert, E.; et al. An unprecedented bloom of Lingulodinium polyedra on the French Atlantic coast during summer 2021. Harmful Algae 2023, 125, 102426. [Google Scholar] [CrossRef] [PubMed]
- Zernov, S.A. On the problem of studying life in Black Sea. Proc. Emp. Acad. Sci. 1913, 32, 1–310. (In Russian) [Google Scholar]
- Terenko, G.; Krakhmalnyi, A. Red tide of the Lingulodinium polyedrum (Dinophyceae) in Odessa Bay (Black Sea). Turk. J. Fish. Aquat. Sci. 2022, 22, TRJFAS20312. [Google Scholar] [CrossRef]
- Moncheva, S.P. On the thermohaline characteristics of development of some massive phytoplankton species from the northern part of the Bulgarian coast. Oceanology 1991, 21, 14–20. (In Bulgarian) [Google Scholar]
- Dzhembekova, N.; Moncheva, S. Recent trends of potentially toxic phytoplankton species along the Bulgarian Black Sea area. In Proceedings of the 12 International Conference on Marine Science and Technology “Black Sea 2014”, Varna, Bulgaria, 25–27 September 2014; pp. 321–329, ISSN 1314-0957. [Google Scholar]
- Ryabushko, L.I. Potentially Dangerous Microalgae of the Azov-Black Sea Basin; ECOSI–Hydrophysics: Sevastopol, Ukraine, 2003; pp. 1–288. (In Russian) [Google Scholar]
- Velikova, V.; Moncheva, S.; Petrova, D. Phytoplankton dynamics and Red Tides (1987–1997) in the Bulgarian Black Sea. Water Sci. Technol. 1999, 39, 27–36. [Google Scholar] [CrossRef]
- Oguz, T.; Velikova, V. Abrupt transition of the northwestern Black Sea shelf ecosystem from a eutrophic to an alternative pristine state. Mar. Ecol. Prog. Ser. 2010, 405, 231–242. Available online: http://www.jstor.org/stable/24873891 (accessed on 2 July 2022). [CrossRef]
- Klisarova, D.; Gerdzhikov, D.; Nikolova, N.; Gera, M.; Veleva, P. Influence of Some Environmental Factors on Summer Phytoplankton Community Structure in the Varna Bay, Black Sea (1992–2019). Water 2023, 15, 1677. [Google Scholar] [CrossRef]
- Valkanov, A. Catalogue of Our Black Sea Fauna; State Publishing House “Science and Art”: Sofia, Bulgaria, 1957; pp. 1–62. (In Bulgarian) [Google Scholar]
- Petrova, V.Y. Phytoplankton in the Black Sea in front of the Bulgarian coast during the period 1958–1960. Proc. Inst. Fish Farming Fish.-Varna 1964, 5, 5–32. (In Bulgarian) [Google Scholar]
- Petrova, V.Y. Peculiarities in the development of phytoplankton in the Black Sea in front of the Bulgarian coast in 1961–1963. Proc. Res. Inst. Fish. Oceanogr.-Varna 1965, 6, 63–74. (In Bulgarian) [Google Scholar]
- Petrova, V. Phytoplankton in the Black Sea in front of the Bulgarian coast for the period 1954–1957. Proc. Cent. Res. Inst. Fish Farming Fish.-Varna 1963, 3, 31–60. (In Bulgarian) [Google Scholar]
- Moncheva, S. Phytoplankton in the food of cultured mussels Mytilus galloprovicialis Lam. in the area of Cape Kaliakra. Proc. Inst. Fish Resour.-Varna 1983, 20, 145–152. (In Bulgarian) [Google Scholar]
- Marasović, I.; Pucher-Petković, T.; Petrova-Karadjova, V. Prorocentrum minimum (Dinophyceae) in the Adriatic and Black Seas. J. Mar. Biol. Assoc. U. K. 1990, 70, 473–476. [Google Scholar] [CrossRef]
- Petrova-Karadjova, V. Phytoplankton blooms in the Black Sea. Sci. Rep. Sci. Dev. Contemp. Social. Pract. 1979, 2, 8–12. (In Bulgarian) [Google Scholar]
- Petrova-Karadjova, V.J. Changes in the planktonic flora in the Bulgarian Black Sea waters under the influence of eutrophication. Proc. Inst. Fish Resour. Varna 1984, 21, 105–112. (In Bulgarian) [Google Scholar]
- Petrova-Karadjova, V.J. The “red tide” of Prorocentrum micans Ehr. and Exuviaella cordata Ost. in the Bay of Varna in November 1984. Hydrobiology 1985, 26, 70–78. (In Bulgarian) [Google Scholar]
- Temniskova, D. Assoc. Prof. Vyara Petrova-Karadzhova: A life dedicated to the sea. In memory of the 75th anniversary of her birth. Phytol. Balc. 2005, 11, 103–118. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/11_2/11_2_01_Temniskova.pdf (accessed on 25 June 2024).
- Mee, L.D. The Black Sea in a crisis: A need for concerted international action. Ambio 1992, 21, 278–285. [Google Scholar]
- Moncheva, S. On some biological aspects of the “blooming” phenomena. In Proceedings of the Scientific-Practical Conference, Status of Research, Rational Utilization and Protection of Natural Resources of the Varna Region, Varna, Bulgaria, 30 October 1989; pp. 110–121. (In Bulgarian). [Google Scholar]
- Oguz, T.; Abaza, V.; Akatov, V.; Aktan, Y.; Arashkevich, E.; Birkun, A.; Boicenco, L.; Chikina, M.V.; Cociasu, A.; Daskalov, G.M.; et al. State of the Environment of the Black Sea (2001–2006/7); Black Sea Commission Publications: Istanbul, Turkey, 2008; pp. 1–421. ISBN 978-9944-245-33-3. [Google Scholar]
- Alexandrov, B.; Krutov, A.; Korshenko, A.; Lavrova, O.; Raykov, V.; Ivanova, P.; Dencheva, K.; Antonidze, E.; Golumbeanu, M.; Gvilava, M.; et al. State of the Environment of the Black Sea (2009–2014/5); Krutov, A., Ed.; Black Sea Commission Publications: Istanbul, Turkey, 2019; pp. 1–811. ISBN 978-605-84837-0-5. [Google Scholar]
- Shtereva, G.; Velikova, V.; Doncheva, V. Human impact on marine water nutrients enrichment. J. Environ. Prot. Ecol. 2015, 16, 40–48. [Google Scholar]
- Moncheva, S.; Doncheva, V.; Kamburska, L. On the long-term response of harmful algal blooms to the evolution of eutrophication off the Bulgarian Black Sea coast: Are the recent changes a sign of recovery of the ecosystem-the uncertainties. In Proceedings of the IX International Conference on Harmful Algal Blooms, Hobart, Tasmania, 7–11 February 2000; UNESCO-IOC: Paris, France, 2001; pp. 177–182. [Google Scholar]
- Moncheva, S.; Slabakova, V.; Doncheva, V. Dominant habitat types in the water column—Phytoplankton. In Initial Assessment of the State of the Marine Environment, According to Article 8 of the WFD and the NOEMS; Publication of Institute of Oceanology: Varna, Bulgaria, 2013; pp. 168–180. (In Bulgarian) [Google Scholar]
- Mee, L.D.; Friedrich, J.; Gomoiu, M.T. Restoring the Black Sea in Times of Uncertainty. Oceanography 2005, 18, 32–43. [Google Scholar] [CrossRef]
- Zaitzev, Y.P. Impact of eutrophication on the Black Sea fauna. Gen. Fish. Counc. Mediterr. Stud. Rev. 1993, 64, 63–86. [Google Scholar]
- Bruno, M.; Gucci, P.M.; Pierdominici, E.; Ioppolo, A.; Volterra, L. Presence of saxitoxin in toxic extracts from Gonyaulax polyedra. Toxicon 1990, 28, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Wang, D. Neurotoxins from Marine Dinoflagellates: A Brief Review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Lee, J. Environmental Bloom of Lingulodinium polyedrum in Southern California: Potential Health Risks. Master’s Thesis, University of California, San Diego, CA, USA, 2015. Available online: https://escholarship.org/uc/item/3zb9b1rp (accessed on 20 February 2022).
- Paz, B.; Riobó, P.; Fernández, M.L.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.D.A.; Smith, G.J.; Kudela, R.M. Phylogenetic relationships of yessotoxin-producing dinoflagellates based on the large subunit and internal transcribed spacer ribosomal DNA domains. Appl. Environ. Microbiol. 2009, 7, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Knechtges, P. Food Safety: Theory and Practice; Jones & Bartlett Learning: Bulrington, VT, USA, 2012; pp. 1–459. [Google Scholar]
- Stobo, L.A.; Lewis, J.; Quilliam, M.A.; Hardstaff, W.R.; Gallacher, S.; Webster, L.; Smith, E.; McKenzie, M. Detection of Yessotoxin in UK and Canadian Isolates of Phytoplankton and Optimization and Validation of LC-MS Methods; Bates, S., Ed.; Gulf Fisheries Centre: Moncton, NB, Canada, 2003; pp. 8–14.
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin Levels and Profiles in Microalgae from the North-Western Adriatic Sea—15 Years of Studies on Cultured Species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Mertens, K.N.; Derrien, A.; David, O.; Shin, H.H.; Li, Z.; Cao, X.; Cabrini, M.; Klisarova, D.; Gu, H. Gonyaulax montresoriae sp. nov. (Dinophyceae) from the Adriatic Sea produces predominantly yessotoxin. Harmful Algae 2025, 141, 102761. [Google Scholar] [CrossRef]
- Granéli, E.; Tuner, J.T. Ecology of Harmful Algae; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1–413. [Google Scholar]
- Yasakova, O.N. Seasonal dynamics of potentially toxic and harmful species of planktonic algae in Novorossiysk Bay (Black Sea). Biol. Sea 2013, 39, 98–105. (In Russian) [Google Scholar]
- Morton, S.L.; Vershinin, A.; Leighfield, T.; Smith, L.; Quilliam, M. Identification of yessotoxin in mussels from the Caucasian Black Sea Coast of the Russian Federation. Toxicon 2007, 50, 581–584. [Google Scholar] [CrossRef]
- European Marine Observation and Data Network (EMODnet). Available online: https://emodnet.ec.europa.eu/en/bathymetry (accessed on 4 November 2024).
- Basemap. Available online: https://becagis.vn/becagis-for-community/?lang=en (accessed on 4 November 2024).
- Morozova-Vodyanitskaya, N.V. Phytoplankton of the Black Sea, part II. Proc. Sevastopol Biol. Stn. 1954, 8, 11–99. (In Russian) [Google Scholar]
- Moncheva, S.; Parr, B. Manual for Phytoplankton Sampling and Analysis in the Black Sea; Black Sea Commission: Istanbul, Turkey, 2010; pp. 1–68. [Google Scholar]
- Edler, L. Recommendations for Marine Biological Studies in the Baltic Sea Phytoplankton and Chlorophyll; Baltic Marine Biologists: Uppsala, Sweden, 1979; pp. 5–38. [Google Scholar]
- Olenina, I.; Hajdu, S.; Edler, L.; Andersson, A.; Wasmund, N.; Busch, S.; Göbel, J.; Gromisz, S.; Huseby, S.; Huttunen, M.; et al. Biovolumes and size-classes of phytoplankton in the Baltic Sea. HELCOM Balt. Sea Environ. Proc. 2006, 106, 1–144. [Google Scholar]
- Lebour, M.V. The Dinoflagellates of Northern Seas; Marine Biological association of the United Kingdom: Plymouth, UK, 1925; pp. 1–251. [Google Scholar]
- Dajoz, R. Précis D’écologie, 7th ed.; Dunod: Paris, France, 2000; pp. 1–615. (In French) [Google Scholar]
- Hurrell, J.; Phillips, A.; National Center for Atmospheric Research Staff (Eds.) Last Modified 2023-07-10 “The Climate Data Guide: Hurrell North Atlantic Oscillation (NAO) Index (Station-Based)”. Available online: https://climatedataguide.ucar.edu/climate-data (accessed on 13 December 2024).
- Schneider, D.P.; Deser, C.; Fasullo, J.; Trenberth, K.E. Climate Data Guide Spurs Discovery and Understanding. Eos Trans. AGU 2013, 94, 121–122. [Google Scholar] [CrossRef]
- Klisarova, D. Phytomar 2.0—Software; Institute of Fish Resources: Varna, Bulgaria, 2008. [Google Scholar]
- Velikova, V.; Atanasova, V.; Manasieva, S.; Daskalov, G. Some aspects regarding the recent state of the Black Sea ecosystem. Proc. Inst. Fish.-Varna 1996, 24, 105–116. (In Bulgarian) [Google Scholar]
- Oguz, T.; Dippner, J.W.; Kaymaz, Z. Climatic regulation of the Black Sea hydro-meteorological and ecological properties at interannual-to-decadal time scales. J. Mar. Syst. 2006, 60, 235–254. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Rachev, G.; Asenova, N. Contemporary Changes in Air Temperature and Precipitation in Bulgaria. Annu. Sofia Univ. St. Kliment Ohridski Fac. Geol. Geogr. 2018, 110, 7–24. (In Bulgarian) [Google Scholar]
- Velikova, V.; Cociasu, A.; Popa, L.; Boicenco, L.; Petrova, D. Phytoplankton community and hydrochemical characteristics of the Western Black Sea. Water Sci. Technol. 2005, 51, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Shtereva, G. Water quality in Varna Bay after the flood in June 2014. Proceeding US-Varna Ser. Mar. Sci. 2014, 1, 67–74. (In Bulgarian) [Google Scholar]
- Velikova, V.; Petrova, D. State of phytoplankton community in the lakes of Beloslav and of Varna during the period from 1991 till 1997. Proc. Inst. Fish Resour. 1999, 25, 103–124. (In Bulgarian) [Google Scholar]
- Stoykov, S.; Kolarov, P.; Stanev, T.; Murdjeva, D.; Atanassova, V.; Kolemanova, K. Ecological state of the Bourgas bay biota (1991–1992). Proc. Inst. Fish.-Varna 1994, 22, 5–57. (In Bulgarian) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Longitude (Eastern) | Latitude (Northern) | |
|---|---|---|
| Durankulak | 28.9539 | 43.6849 |
| Shabla | 28.9799 | 43.5333 |
| Varna | 28.3017 | 43.2026 |
| Tsarevo | 28.2239 | 42.1703 |
| Variable | ||||
|---|---|---|---|---|
| D1 | D2 | D3 | D4 | |
| (n = 38) | (n = 22) | (n = 28) | (n = 3) | |
| Average abundance (mln. cells·m−3) | 10.37 ± 28.78 a | 1.75 ± 1.64 a | 2.38 ± 1.99 a | 2.62 ± 2.54 |
| R2 = 0.047; Sig.(p) = 0.043 | ||||
| Average biomass (mg·m−3) | 553.40 ± 1525.57 a | 93.69 ± 91.42 a | 101.85 ± 87.20 a | 124.01 ± 149.69 |
| R2 = 0.050; Sig.(p) = 0.013 | ||||
| Average volume of cell (µm3) | 54,034.98 ± 6849.47 a | 53,041.24 ± 6701.50 b | 45,456.77 ± 10,846.40 ab | 37,427.54 ± 16,852.03 |
| R2 = 0.223; Sig.(p) = 0.001 | ||||
| Frequency of occurrence (%) | 24.93 ± 21.79 | 20.42 ± 18.13 | 18.91 ± 11.58 | 28.17 ± 13.38 |
| R2 = 0.026; Sig.(p) = 0.510 | ||||
| Month | Max Abundance (mln. cells·m−3) | Average Abundance (mln. cells·m−3) | Min Abundance (mln. cells·m−3) | Max Biomass (mg·m−3) | Average Biomass (mg·m−3) | Min Biomass (mg·m−3) |
|---|---|---|---|---|---|---|
| 2 | 0.10 | 0.08 | 0.06 | 5.42 | 4.55 | 3.67 |
| 3 | 20.00 | 11.46 | 2.93 | 769.30 | 458.21 | 147.12 |
| 4 | 1.24 | 0.39 | 0.08 | 70.24 | 21.09 | 1.94 |
| 5 | 8.14 | 2.29 | 0.04 | 460.18 | 130.29 | 2.08 |
| 6 | 13.33 | 2.60 | 0.03 | 762.13 | 146.11 | 1.52 |
| 7 | 81.44 | 3.11 | 0.05 | 4603.07 | 151.94 | 1.85 |
| 8 | 1000.00 | 13.15 | 0.05 | 52,752.00 | 686.72 | 1.40 |
| 9 | 40.08 | 5.45 | 0.05 | 2265.52 | 285.76 | 3.06 |
| 10 | 28.98 | 5.86 | 0.06 | 1528.81 | 298.17 | 3.55 |
| 11 | 20.00 | 2.40 | 0.05 | 906.83 | 128.27 | 2.08 |
| Variable | D1 | D2 | D3 | |||
|---|---|---|---|---|---|---|
| Period (n) | Period (n) | Period (n) | ||||
| Average abundance (mln. cells·m−3) | D1F1 (16) | 19.61 ± 42.96 a | D2F1 (13) | 0.97 ± 0.89 a | D3F1 (20) | 2.22 ± 1.96 |
| D1F2 (22) | 3.65 ± 5.29 a | D2F2 (9) | 2.89 ± 1.84 a | D3F2 (8) | 2.78 ± 2.14 | |
| Average biomass (mg·m−3) | D1F1 (16) | 1035.50 ± 2277.40 b | D2F1 (13) | 50.03 ± 45.63 b | D3F1 (20) | 96.72 ± 89.51 |
| D1F2 (22) | 202.78 ± 300.10 b | D2F2 (9) | 156.74 ± 106.03 b | D3F2 (8) | 114.68 ± 85.56 | |
| Average volume of cell (µm3) | D1F1 (16) | 54,497.53 ± 5503.18 | D2F1 (13) | 52,301.22 ± 7254.58 | D3F1 (20) | 46,237.62 ± 10,572.75 |
| D1F2 (22) | 53,698.58 ± 7794.01 | D2F2 (9) | 54,110.14 ± 6063.871 | D3F2 (8) | 43,504.68 ± 12,010.96 | |
| Frequency of occurrence (%) | D1F1 (16) | 25.78 ± 17.63 | D2F1 (13) | 21.93 ± 16.51 | D3F1 (20) | 18.32 ± 9.52 |
| D1F2 (22) | 24.32 ± 24.78 | D2F2 (9) | 18.24 ± 21.09 | D3F2 (8) | 20.38 ± 16.37 | |
| № | Regions | Distance from the Shore | Max of Abundance (mln. cells·m−3) | Max of Biomass (mg·m−3) |
|---|---|---|---|---|
| 1 | Biala | offshore | 3.35 | 70.96 |
| 2 | Albena | offshore | 0.16 | 8.79 |
| 3 | Balchik | offshore | 4.91 | 121.48 |
| 4 | Beloslav lake | offshore | 11.59 | 799.35 |
| 5 | Burgas bay | offshore | 40.08 | 2114.22 |
| 6 | cape Emine | offshore | 18.29 | 965.08 |
| 7 | cape Emine | open sea | 14.67 | 674.27 |
| 8 | cape Galata | offshore | 81.44 | 4603.07 |
| 9 | cape Galata | open sea | 9.43 | 532.90 |
| 10 | cape Kaliakra | offshore | 9.01 | 191.15 |
| 11 | cape Kaliakra | open sea | 1.19 | 67.49 |
| 12 | cape Maslen | offshore | 0.55 | 31.25 |
| 13 | Dvoinitca | offshore | 0.43 | 21.41 |
| 14 | Ilandzhik | offshore | 1.51 | 85.36 |
| 15 | Kamchia | offshore | 0.34 | 19.00 |
| 16 | Krapec | offshore | 0.26 | 9.91 |
| 17 | Rusalka | offshore | 2.43 | 93.32 |
| 18 | Shabla | offshore | 5.67 | 160.28 |
| 19 | Sozopol | offshore | 2.72 | 104.73 |
| 20 | Varna bay | offshore | 1600.00 | 84,403.20 |
| 21 | Varna lake | offshore | 8.14 | 460.18 |
| 22 | Varvara | offshore | 0.66 | 37.58 |
| 23 | Veleka | offshore | 1.94 | 87.86 |
| n = 88 | D1 | D2 | D3 | |||
|---|---|---|---|---|---|---|
| SST | NAO | SST | NAO | SST | NAO | |
| Average abundance (mln. cells·m−3) | 0.118 | 0.110 | 0.242 | 0.038 | 0.051 | −0.300 |
| Average biomass (mg·m−3) | 0.172 | 0.090 | 0.216 | 0.155 | −0.085 | −0.303 |
| Average volume of cell (µm3) | 0.003 | 0.037 | 0.056 | 0.554 * | −0.188 | 0.067 |
| Frequency of occurrence (%) | 0.374 * | 0.374 * | −0.152 | 0.103 | −0.054 | 0.162 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klisarova, D.; Gerdzhikov, D.; Dragomirova, P.; Nikolova, N.; Gera, M.; Veleva, P. Dynamics of Lingulodinium polyedra Development in the Bulgarian Part of Black Sea (1992–2022). Diversity 2025, 17, 105. https://doi.org/10.3390/d17020105
Klisarova D, Gerdzhikov D, Dragomirova P, Nikolova N, Gera M, Veleva P. Dynamics of Lingulodinium polyedra Development in the Bulgarian Part of Black Sea (1992–2022). Diversity. 2025; 17(2):105. https://doi.org/10.3390/d17020105
Chicago/Turabian StyleKlisarova, Daniela, Dimitar Gerdzhikov, Petya Dragomirova, Nina Nikolova, Martin Gera, and Petya Veleva. 2025. "Dynamics of Lingulodinium polyedra Development in the Bulgarian Part of Black Sea (1992–2022)" Diversity 17, no. 2: 105. https://doi.org/10.3390/d17020105
APA StyleKlisarova, D., Gerdzhikov, D., Dragomirova, P., Nikolova, N., Gera, M., & Veleva, P. (2025). Dynamics of Lingulodinium polyedra Development in the Bulgarian Part of Black Sea (1992–2022). Diversity, 17(2), 105. https://doi.org/10.3390/d17020105