Abstract
Plant community stability and diversity play crucial roles in maintaining the stable performance of grassland ecosystem functions. In this study, we selected 55 representative grassland community sample plots in the Habahe Forest region of the Altai Mountains, Xinjiang. We examined the number of species, the number of individuals, and the cover of each species in the sample plots. Additionally, we determined the aboveground biomass of grassland communities, the stability of M. Godron communities, and plant diversity (using the Margalef index, Simpson index, Shannon–Wiener index, and Alatalo evenness index) in the sample plots. We analyzed community stability, plant diversity, and the correlation between community stability, plant diversity, and environmental factors (such as longitude, latitude, altitude, slope, humus thickness, soil thickness, air temperature, precipitation, and soil moisture). The results show that the plant coverage and height of different grassland types in the Habahe Forest area decrease from mountain meadow to desert steppe. The aboveground biomass and species diversity of meadows are significantly higher than those of steppes. The stability of different grassland types is as follows: montane steppe > montane meadow steppe > mountain meadow > desert steppe. The species richness of the four grassland types is positively correlated with community stability, while evenness is negatively correlated with community stability. Dominance in montane steppe and montane meadow steppe is significantly positively correlated with community stability, whereas diversity in desert steppe and montane steppe is significantly negatively correlated with community stability. Precipitation and soil moisture are the main factors affecting species diversity and stability in the Habahe Forest area. Soil nutrients and slope can also impact community stability by affecting species diversity. In summary, the ecological management of grasslands in the Habahe Forest area should consider climate, soil, and terrain factors, and different management measures should be selected to adapt to different grassland types.
1. Introduction
The grassland ecosystem is the largest terrestrial ecosystem in the world, covering about half of the Earth’s land area. It not only provides ecological services and abundant resources for humans but also plays a crucial role in stabilizing the global ecosystem [,]. During the lengthy evolutionary process, grassland ecosystems have adapted to environmental disturbances such as grazing, salinization, and climate extremes. However, intensified climate change and frequent anthropogenic disturbances now severely threaten the diversity, stability, and ecological services provided by grassland organisms. The decline in species diversity may further jeopardize the stability of grassland ecosystems [,]. The Habahe Forest area is situated in the southern foothills of the Altai Mountains in Xinjiang, China. It is abundant in grazing resources and encompasses a vast expanse of grassland, crucial for the development of animal husbandry in Xinjiang. Additionally, it plays a significant role in climate regulation and enhancing the ecological environment [,,]. The fragile ecological environment of the Altai Habahe Forest and the degradation of grasslands may lead to a decline in biodiversity and further threaten ecosystem stability []. Therefore, studying the ecological characteristics, fluctuations in biodiversity, and stability of different grassland types, as well as their interrelationships, is crucial for analyzing the combined effects of environmental factors and anthropogenic disturbances on plants. This research is of profound significance for effectively conducting ecological conservation of grasslands and maintaining ecological balance.
Plant community stability plays a central role in ecosystem sustainability and serves as a key indicator of ecosystem health. Among the various methods used to assess plant community stability, species diversity is considered one of the most important determinants [,]. Currently, there is still no consensus regarding the relationship between plant community stability and species diversity, with three main perspectives: positive correlation, negative correlation, and no clear relationship []. Early scholars like MacArthur [] and Elton [] posited that simpler ecosystems are less stable, as species diversity directly influences the ecosystem’s resilience to external disturbances; thus, higher diversity enhances plant community stability. Later, researchers such as Tilman [,], Naeem [], and McGrady [] provided strong support for the diversity–stability theory through long-term experiments. A comprehensive analysis of 52 studies revealed that over 60% demonstrated a positive correlation between stability and diversity []. Furthermore, a study across different vegetation zones on the Loess Plateau highlighted a significant positive correlation between species diversity and community stability []. However, doubts persist regarding the diversity–stability theory, with some scholars suggesting that higher diversity may disrupt species balance and lead to reduced stability [,]. Lei [] found that different diversity indexes had different effects on community stability in the study of alpine meadow and alpine desert steppe on the Qinghai–Tibet Plateau, but it was generally observed that there was a linear negative correlation between diversity and stability. Li [] and Zheng [] analyzed species diversity and community stability under varying levels of wildfire disturbance and concluded that their relationship is not simply linear after studying the stability of typical forest communities in the Liaohe River area in northern Hebei Province. Yu [] found that the effect of diversity on stability is noticeable only within a certain threshold. Additionally, it has been argued that communities are in a constant state of dynamic change, challenging the existence of a direct relationship between species diversity and community stability []. Studies from different regions have shown that topography [], soil [], and climate [] can be the main environmental factors influencing species diversity and community stability. The interaction between plant community diversity and soil maintains ecosystem stability and may better reflect the ecological adaptability of the community []. In a study of the distribution pattern of plant communities in eastern Tibet, Zhang [] found that elevation, aspect, slope, and soil nutrients determined the distribution of regional plant communities, with elevation, aspect, and slope having significant effects on community species diversity. Studies have shown that higher soil moisture has a significant positive effect on increasing plant species, plant growth rate, and plant diversity index []. Slope and aspect can comprehensively characterize habitat conditions, closely related to light, humidity, temperature, water, and soil factors, directly determining plant growth conditions []. Mendez et al. [] studied seasonal tropical forests and found that slope aspect influenced the relationship between elevation and species richness. Species richness on the southern slope was positively correlated with elevation, while species richness on the northern slope was negatively correlated with elevation. Massante [] showed that community biodiversity increased with increasing rainfall. Recent research has also shown that environmental factors strongly influence community stability. Environmental factors affect the structure and evolution of biological communities, which in turn affects community stability []. Through the above research, it can be seen that there are significant differences in the relationship between plant community distribution patterns, diversity, stability, and environment under different natural conditions. Altitude and precipitation may be key factors in the change in mountain plant communities. Therefore, it is necessary to study the effects of environmental factors on plant community stability and species diversity. Additionally, the influence of scale, environmental factors, disturbance intensity, and community characteristics should be considered [].
The purpose of this study was to explore the differences in community characteristics among different grassland types and the relationship between grassland plant species diversity and community stability in the Habahe Forest area of the Altay Mountains. Additionally, the effects of environmental factors on species diversity and community stability were analyzed. This study specifically validated the following assumptions: (1) In different grassland types in the Habahe Forest area, the species diversity and community stability of meadows are higher than those of other grasslands. (2) There is a positive correlation between grassland plant species diversity and community stability in the Habahe Forest area.
2. Materials and Methods
2.1. Overview of the Study Area
Habahe is situated in the southern foothills of the Altai Mountains and the northern edge of the Junggar Basin in China, located between 85°33′ and 87°18′ E longitude and 47°37′ and 49°07′ N latitude, with elevations ranging from about 410 to 3396 m [] (Figure 1). The region experiences an inland cold continental climate, characterized by an average annual temperature of approximately 5.3 °C. Temperatures can reach up to 30 °C in summer and drop to −19 °C in winter, resulting in significant diurnal temperature variations. Annual rainfall averages 205.6 mm, contributing to a dry climate with high evaporation rates and extensive sunshine hours. Habahe is known for its dry and windy springs, short and hot summers, cool autumns, and cold winters [].
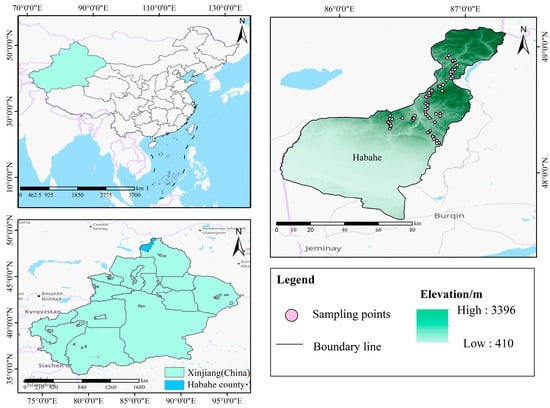
Figure 1.
Overview maps of the study area.
The region hosts a rich diversity of grassland plants, including Festuca ovina, Stipa capillata, Poa annua, Artemisia frigida, Helictochloa hookeri, Geranium wilfordii, Potentilla chinensis, and Taraxacum mongolicum, among others [].
2.2. Experimental Design
The field survey of grassland plant communities was conducted from June to July 2022, during the peak season of pasture growth. Sample strips were set up at different elevations, river valleys, pasture areas, and tourist areas in the natural grasslands of the Habahe Forest area. Based on elevation distribution characteristics, the Habahe Forest area was categorized into four primary types of grasslands: desert steppe (DS), montane steppe (MS), montane meadow steppe (MMS), and mountain meadow (MM). For each grassland type, 55 sample plots and 165 sample squares were randomly distributed. Each sample plot measured 20 m × 20 m, with three 1 m × 1 m grassland sample squares randomly selected from each plot (Figure 2). Detailed recording of plant species and quantitative characteristics such as height, density, cover, biomass, etc., was conducted during the sampling process. The distribution of sample plots across different grassland types was as follows: desert steppe: 11 sample plots and 33 sample squares; mountain steppe: 25 sample plots and 75 sample squares; mountain meadow steppe: 10 sample plots and 30 sample squares; mountain meadow: 9 sample plots and 26 sample squares.

Figure 2.
Schematic diagram of sampling points.
Quantitative plant characteristics were measured as follows: height was measured using a tape measure to determine natural height; density was recorded by directly counting the number of species per square meter; cover was assessed visually; and aboveground biomass was measured using the flush mowing method, where the fresh weight of aboveground biomass for each species was weighed and recorded as biomass.
2.3. Research Methodology
- (1)
- Species diversity was assessed using the Margalef richness index (S), the Simpson dominance index (D), the Shannon–Wiener diversity index (H), and the Alatalo evenness index (Ea) as measures of α-diversity, calculated with the following formulas [,]:
Margalef index: This index reflects species richness within the community; higher values indicate greater species diversity.
Simpson index: This index measures the prominence and role of dominant species in the community; higher values indicate more pronounced dominance and a more uneven distribution of species.
Shannon–Wiener index: This index reflects the overall status of species richness and evenness within the community; higher values indicate greater species richness, higher evenness, and higher community diversity.
Alatalo index: This index reflects the distribution uniformity of species within the community; higher values indicate a more uniform distribution of species.
In the above equations, S represents the total number of species in the sample; N denotes the total number of individuals in the sample; and Pi signifies the ratio of the number of individuals of plant species i to the total number of individuals [].
- (2)
- The M. Godron stability measure was introduced to the study of plant ecology by French ecologists from industrial production. In this study, the stability of grassland communities in the Habahe region was evaluated using the law-of-contribution method proposed by M. Godron []. This method sorts all plant species within the community based on their frequency, calculates their relative frequencies, and then accumulates the reciprocal of the total number of species and their relative frequencies. Subsequently, the cumulative percentages of inverse plant species and relative frequencies are matched and curve-fitted to obtain a smooth curve equation y = ax3 + bx2 + cx + d intersecting with y = 100 − x. The intersection point (x, y) is determined, and the Euclidean distance from this point to (20, 80) is calculated. A smaller distance indicates greater community stability, while a larger distance suggests instability. Zheng [] proposed using species cover instead of frequency to better reflect interaction relationships between plant species, thereby enhancing stability measurement accuracy. Accordingly, this study adopted an improved M. Godron stability measure, using the reciprocal of the Euclidean distance (ESD value) as the M. Godron index to characterize community stability. The equation used for calculation is as follows []:
- (3)
- Plant communities’ similarities were measured using the Jaccard similarity coefficient according to the following formula []:
2.4. Statistical Analysis
Microsoft Excel 2019 was utilized for initial organization and calculation of data. SPSS 26.0 software was employed for statistical analysis, utilizing independent-sample t-tests to assess differences in stability among various grassland community types in the Habahe area. Since the samples obtained were independent random samples, we used one-way analysis of variance (ANOVA) to analyze the variability of vegetation community characteristics and the four diversity indices of different grassland types. We used the least significant difference (LSD) method in the one-way ANOVA for comparison tests, all of which were tested for significance at the 0.05 level. Pearson’s correlation coefficient was used to examine linear relationships between community stability and diversity in different grassland types, as well as correlations between plant community characteristics and environmental factors. ArcGIS 10.8 was used to create an overview map of the study area, while Origin 2021 was employed for additional map creation tasks.
Based on field surveys, the Chinese Vegetation Journal was consulted to identify the Latin names and habitat types of species recorded in the Altai Mountains, and we compiled a list of dominant grassland plant species in the study area (Table 1).

Table 1.
Characteristics of community structure of different grassland types in Habahe.
2.5. Determination of Environmental Factors
The altitude (AL), longitude (EL), and latitude (NL) were measured by GPS. The slope (SL) was measured using a compass. Soil drilling tools were used to sample soil in the quadrat, and the soil thickness (ST) and humus layer thickness (HT) were measured with a scale. Soil moisture (SM) was monitored with a soil detector. Meteorological data, including annual mean precipitation (AMP) and annual mean temperature (AMT), were obtained from the World Climate Database (https://www.worldclim.org/, accessed on 4 August 2024).
3. Results
3.1. Characteristics of Community Structure in Different Grassland Types
There are four primary types of grasslands in the Altai Habahe Forest region: desert steppe, montane steppe, montane meadow steppe, and mountain meadow. Desert grasslands, as integral components of the desert ecosystem, play crucial roles in water conservation, wind and sand control, climate regulation, biodiversity preservation, and ecosystem stability. Dominant species in desert grasslands include Stipa grandis, Festuca valesiaca, and Artemisia frigida. Montane steppe exhibits the highest species richness, totaling 63 species. Montane meadow steppe is characterized by a diverse array of companion species, notably including Achillea millefolium, Phlomoides oreophila, Bromus inermis, Galium verum, Fragaria vesca, and Medicago falcata (Table 1).
Analysis of vegetation community structural characteristics in the grasslands of the Habahe Forest region revealed significant differences among grassland types. Notably, there was a pronounced decreasing trend in area and plant height from mountain meadow to montane steppe to desert steppe (p < 0.05), with the most significant differences observed between desert steppe and mountain meadow (p < 0.01). Aboveground biomass of plants showed a gradual decline from montane meadow steppe through montane steppe to desert steppe, with the difference between desert steppe and montane meadow steppe being highly significant (p < 0.01) (Figure 3). Analysis of species diversity maps across different grassland types in the Habahe Forest area revealed that the Simpson index, Shannon–Wiener index, and Alatalo index were significantly lower in desert steppe compared to the other three grassland types (p < 0.05). The Margalef index and Alatalo index were highest in montane steppe among the four grassland types, indicating richer and more evenly distributed species. Mountain meadow steppe exhibited the highest Shannon–Wiener index and overall species diversity compared to other grassland types. Analysis of Simpson’s index variation among different grassland types indicated that mountain meadow steppe had the highest Simpson’s index, while its Shannon–Wiener and Alatalo indices were relatively lower. In summary, significant differences in diversity levels were observed among the different grassland types in the Habahe Forest region, with the Simpson index showing the largest differences and the Margalef index showing the smallest differences (Figure 4).
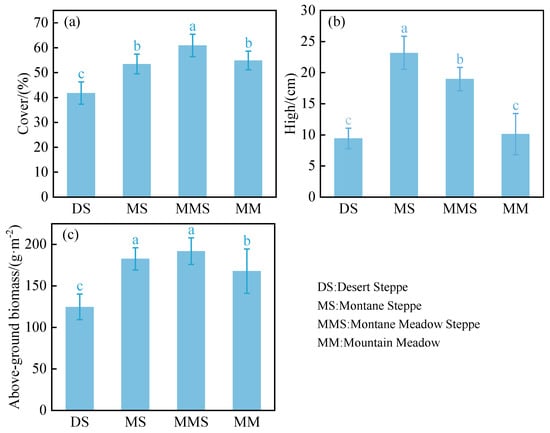
Figure 3.
Characteristics of community cover in different grassland types (a); Characteristics of community height in different grassland types (b); Characteristics of community above-ground biomass in different grassland types (c). Different lowercase letters in the superscripts of the figure columns indicate significant differences between the various grassland types (p < 0.05).
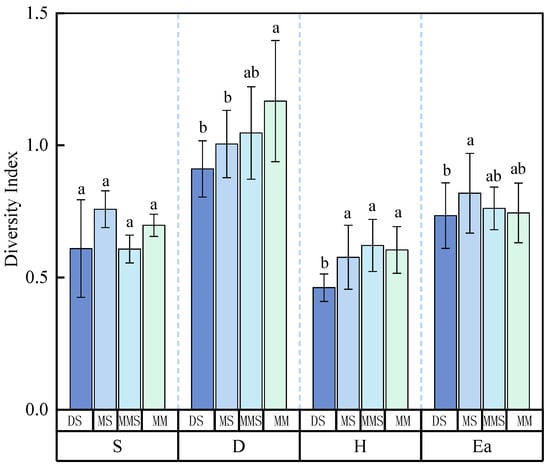
Figure 4.
Changes in species diversity among different grassland types in the Habahe Forest area. Different lowercase letters in the superscripts of the figure columns indicate significant differences between the various grassland types (p < 0.05). S denotes the Margalef index, D denotes the Simpson index, H denotes the Shannon–Wiener index, and Ea denotes the Alatalo index.
3.2. Stability and Similarity Analysis of Communities in Different Grassland Types
The stability of plant communities across different grassland types in the Habahe Forest area of the Altai Mountains was assessed using M. Godron’s stability measure combined with mathematical simulation methods to generate plant species curves and frequency scatter plots (Figure 5, Table 2). The intersection coordinates of these plots closer to the stabilization point (20, 80) indicate greater community stability, whereas those farther away indicate less stability. The results of M. Godron’s stability simulation showed ESD (Euclidean distance) values at the intersection of fitted curves with the y = 100 − x line for desert steppe, mountain steppe, mountain meadow steppe, and mountain meadow plant communities as 4.88, 0.41, 0.59, and 0.88, respectively. Thus, the stability ranking among grassland types was mountain steppe > mountain meadow steppe > mountain meadow > desert steppe. Desert steppe exhibited the highest ESD value, indicating lower stability compared to other grassland types, while mountain steppe showed the lowest ESD value, indicating the highest stability.
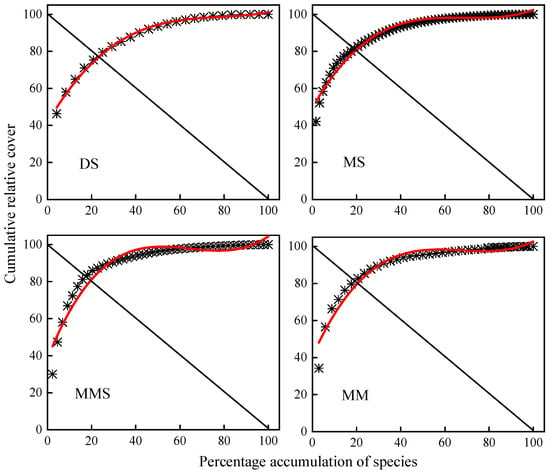
Figure 5.
Simulation curves of M. Godron stability in different grassland types in Habahe Forest area.
Table 2.
Stability characteristics of different grassland communities in Habahe Forest area.
In summary, the analysis revealed variations in stability among the four grassland types in the Habahe Forest area, yet all communities generally maintained stable conditions.
Using Jaccard’s similarity coefficients, Table 3 presents the shared vegetation communities among the four grassland types in the Habahe Forest region. Coefficients between desert steppe and montane steppe, as well as mountain meadow, ranged from 0 to 0.25, indicating very low or no similarity. Similarity coefficients between montane steppe and desert steppe, and mountain meadow, ranged from 0.25 to 0.50, indicating moderate dissimilarity. Montane meadow steppe exhibited similarity coefficients of 0.50 to 0.75 with both montane steppe and mountain meadow, indicating moderate similarity. The lowest similarity coefficient was found between desert steppe and mountain meadow at 0.184, corresponding to only nine common species. Montane steppe and mountain meadow steppe shared the highest number of species at 38, with a similarity coefficient of 0.535, reflecting the highest community similarity. Overall, significant differences in plant communities exist among the different grassland types in the Habahe Forest area. The greatest differences were observed between desert grassland and mountain meadow, while the least differences were noted between montane steppe and montane meadow steppe. The divergence in plant communities among different grassland types generally increases with geographical distance.
Table 3.
Community similarity of different grassland types in Habahe Forest area.
3.3. Relationship between Species Diversity and Community Stability in Different Grassland Types
To investigate the relationship between species diversity and community stability, this study selected four grassland types in the Habahe Forest area for analysis and employed linear fitting to explore their correlation. Results indicated varied relationships between species diversity and stability across different grassland types, with distinct impacts of different diversity indices within each type. As depicted in Figure 6, the Margalef index exhibited a positive correlation with the community stability index across all four grassland types, particularly showing a highly significant correlation (p < 0.01) in montane meadow steppe. This suggests that community stability increases with greater species richness. Conversely, the correlation between desert steppe and montane steppe was not significant (p > 0.05), indicating that interannual variations in species richness within these communities did not significantly affect stability. The Simpson index showed a positive correlation with the community stability index in montane steppe and montane meadow steppe, with a highly significant correlation (p < 0.01), suggesting that species dominance may better reflect community stability. In contrast, the Shannon–Wiener index exhibited a highly significant negative correlation (p < 0.01) with the community stability index in desert steppe and montane steppe, indicating that community stability decreased with increasing species diversity, particularly showing instability at higher diversity levels. Changes observed in the relationship between the Shannon–Wiener index and community stability index in montane meadow steppe and mountain meadow indicated that as species diversity increased, community stability also increased, emphasizing the role of intra-annual species diversity changes in these grassland types. The Alatalo index showed a negative correlation with plant community stability across all four grassland types, desert steppe, montane steppe, montane meadow steppe, and mountain meadow, indicating that higher species homogeneity can enhance community stability. However, this correlation was not significant (p > 0.05) in mountain meadow, suggesting that species evenness does not strongly predict community stability in this grassland type (Figure 6, Table 4). Overall, these findings highlight the complex relationships between species diversity indices and community stability across diverse grassland ecosystems.
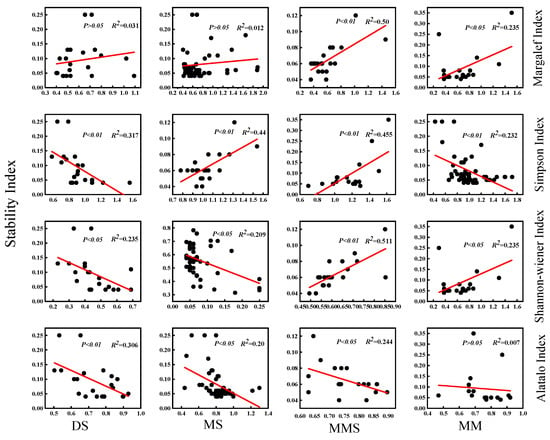
Figure 6.
Relationship between species diversity and community stability of different grassland types in Habahe Forest area.
Table 4.
Fitted equations of species diversity and community stability of plant communities in different grassland types in Habahe Forest area.
3.4. Relationship between Plant Community Characteristics and Environmental Factors
Figure 7 illustrates the correlation analysis between environmental factors and community characteristics, diversity indices, and community stability. The M. Godron index showed a significant positive correlation with precipitation (p < 0.05), indicating that precipitation plays a crucial role in maintaining community stability. The Margalef index exhibited a significant negative correlation (p < 0.05) with latitude and longitude and a significant positive correlation (p < 0.05) with humus thickness. The Simpson index demonstrated significant positive correlations (p < 0.05 and 0.01) with elevation, soil thickness, and slope, suggesting that environmental factors such as elevation, slope, and soil thickness positively influence the dominance of plant communities. The Shannon–Wiener diversity index showed significant positive correlations (p < 0.05 or 0.01) with elevation, soil thickness, and humus thickness, highlighting the importance of elevation and soil factors in promoting plant community diversity within specific ranges. The Alatalo evenness index exhibited a highly significant positive correlation with precipitation (p < 0.01), indicating that grassland community evenness increases under the influence of precipitation. Plant community characteristics showed high sensitivity to environmental factors, with plant cover and aboveground biomass displaying significant positive correlations (p < 0.01) with altitude, longitude, latitude, humus thickness, and soil moisture. Conversely, plant cover, plant height, and aboveground biomass exhibited highly significant negative correlations (p < 0.01) with temperature. These findings suggest that increased soil moisture enhances plant cover and biomass, while lower temperatures correspond to decreased plant cover, height, and aboveground biomass.
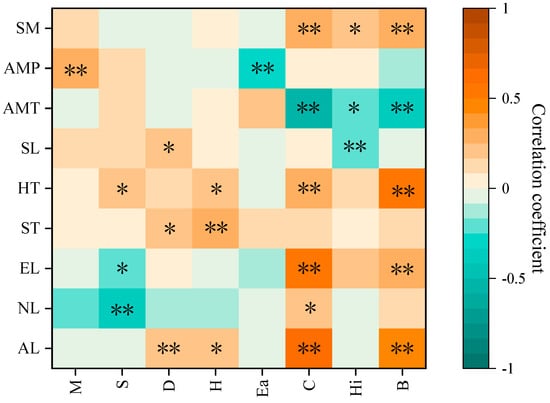
Figure 7.
In the Habahe Forest area, the correlation between plant community characteristics and environmental factors is depicted in the figure. Asterisks (*) denote significant correlations at the p < 0.05 level, while double asterisks (**) indicate highly significant correlations at the p < 0.01 level. Specifically, M represents the stability index; S represents the richness index; D represents the dominance index; H represents the diversity index; Ea represents the evenness index; C represents canopy cover; Hi represents height; B represents biomass; AL represents altitude; NL represents latitude; EL represents longitude; ST represents soil thickness; HT represents humus thickness; SL represents slope; AMT represents annual mean temperature; AMP represents annual mean precipitation; and SM represents soil moisture.
4. Discussion
4.1. Changes in Plant Community Composition under Different Grassland Types
The composition of grassland plant communities is crucial for distinguishing between grassland types and forms the foundation for studying their productivity and stability. Investigating grassland plant communities provides insights into the organization, function, and structure of grassland ecosystems, offering a scientific basis for their management and conservation []. Key characteristics of grassland plant community composition include species diversity and dominance patterns, including dominant, subdominant, companion, and occasional species, alongside other quantitative attributes []. In the study area, 71 plant species belonging to 20 families and 58 genera were identified, predominantly from families such as Poaceae, Asteraceae, Rosaceae, Fabaceae, and Ranunculaceae. These plants are well adapted to the unique ecological conditions of the region, playing a vital role in the Habahe forest area’s grassland ecosystems. Dominant species such as fescue, needle fescue, and morning glory were prominent, consistent with findings from previous studies on typical plant communities in the Altay region of Xinjiang by Su []. Analysis of vegetation coverage and height across different grassland types revealed distinct variations, declining in an orderly manner from mountain meadow to desert steppe, consistent with observations by Li [] in the Tian Shan and Altai mountains. Variations in moisture and climatic conditions among grassland types likely account for differences in vegetation quantity, coverage, and height []. Studies on aboveground biomass indicated that montane meadow steppe exhibited the highest biomass, whereas desert steppe had the lowest. This disparity is closely linked to regional water availability, temperature regimes, and soil conditions. Alpine meadows, situated at higher altitudes and receiving abundant rainfall, experience less continental climatic influence and foster greater organic matter accumulation in soil, promoting vegetation growth. Conversely, desert steppe, characterized by lower altitudes, limited precipitation, and susceptibility to human disturbances such as grazing, exhibits lower biomass due to inhibited vegetation growth [].
In this study, diversity indices among the four grassland types exhibited notable variations. Specifically, the Margalef index, Simpson index, Shannon–Wiener index, and Alatalo index were comparatively lower in desert steppe, displaying significant differences compared to the other grassland types. This disparity likely stems from environmental variations and differing vegetation compositions. Desert steppe ecosystems primarily comprise xerophytic plants, with fewer mesophotic species compared to the other three grassland types []. Moreover, desert steppe areas are susceptible to disturbances from both climatic fluctuations and anthropogenic activities, which disrupt habitats and contribute to lower species diversity [,]. Conversely, montane steppe demonstrated significantly higher richness indices compared to each grassland type, consistent with findings from studies in the Bolzin forest area of the Altai Mountains []. Furthermore, this research identified a notably superior Shannon–Wiener diversity index in mountain meadows compared to grasslands, a difference likely influenced by altitude. Elevated terrains offer improved soil conditions and favorable hydrothermal environments conducive to a broader spectrum of forage grasses, thereby reducing interspecific competition within high-altitude meadows and maximizing species diversity indices. This observation aligns with conclusions drawn by Wang et al. [], who noted that species adaptability and environmental suitability increase with altitude from grasslands to meadows, particularly in mid- to high-altitude zones characterized by favorable water and heat conditions.
4.2. Characterization of Community Stability and Its Relationship with Species Diversity in Different Grassland Types
Community stability refers to the capacity of a community to achieve and maintain a state of relative equilibrium, facilitating interactions among species and the ability to self-regulate and return to equilibrium following disturbances from external factors []. In this study, M. Godron’s method for assessing plant community stability was employed to analyze variations in stability among four grassland types in the Habahe Forest area. The findings indicated that the stability of desert steppe communities was significantly lower compared to montane steppe, montane meadow steppe, and mountain meadow, consistent with previous research [,]. Adequate water availability creates favorable conditions for plant growth, supporting species diversity and maintaining stability in a relatively consistent state. Conversely, water scarcity can impede the normal growth of certain species, thereby contributing to decreased community stability [].
Biodiversity plays a crucial role in delivering ecosystem services and maintaining ecosystem functions, with plant species diversity encompassing richness, dominance, diversity, and evenness. It is widely recognized that higher species diversity in grasslands contributes to greater ecosystem stability and overall health [,]. Studies have indicated that communities with low species diversity tend to exhibit higher stability, whereas those with high species diversity often show lower stability []. In this experiment, the relationship between species diversity and stability varied across different grassland types. Furthermore, the stability characteristics within the same grassland type under the influence of different diversity indices also varied. Overall, the findings demonstrated that increasing species richness can enhance community stability, whereas increasing species evenness may decrease it. This observation aligns with previous research by Hu [], Xu [], and Lei [], who similarly found that greater species richness promotes community stability. Conversely, higher evenness levels may reduce the community’s resilience to disturbances, thereby lowering stability []. The study area, situated in an arid zone, features a fragile ecological environment where species are sensitive to external disturbances. Consequently, ecosystem recovery from severe damage is slow, emphasizing the critical role of biome resilience in maintaining stability. These factors contribute to a negative correlation between evenness and stability in the region’s grassland communities.
In addition to biotic factors, abiotic factors also significantly influence species diversity and community stability, complicating predictions regarding the impact of species diversity on community stability []. It has been observed that the relationships between the Simpson index, Shannon–Wiener index, and community stability vary across different grassland types, showing both positive and negative correlations. This inconsistency underscores the need for further investigation into the nuanced interactions between species diversity and community stability in the Habahe Forest region. Thus, it is imprudent to assume a universal positive or negative correlation between diversity and stability. Species diversity reflects only certain aspects of community stability, influenced by external disturbances, environmental conditions, and inherent community characteristics. Moreover, exploring how the scale of study influences the relationship between diversity and stability warrants attention.
4.3. Species Diversity and Community Stability in Relation to Environmental Factors
The influence of ecological factors such as climate, soil, geomorphology, and geography on plant community species diversity has been extensively studied, yielding complex and varied results [,]. Different geographical locations of communities contribute to unique habitat conditions, influencing the environmental characteristics of each community []. Numerous environmental factors have been employed to explain species diversity patterns in mountain ecosystems, with climatic variables related to heat and moisture playing crucial roles in shaping diversity patterns across different regions and scales []. In this study, environmental factors including longitude, latitude, altitude, air temperature, precipitation, and soil moisture were analyzed for their impacts on α-diversity. The findings indicated a positive correlation between community stability in grasslands within the Habahe Forest area and the Margalef index with precipitation. Changes in precipitation can induce temporal and spatial variations in water availability for plants, thereby altering intra- and interspecific relationships and ultimately affecting community stability []. Precipitation plays a multifaceted role in ecosystem dynamics, facilitating seed germination in soil, enhancing solubilization and movement of soil nutrients [], and promoting nutrient uptake by plants, thereby fostering species diversity and ultimately enhancing community stability []. The Shannon–Wiener index and Alatalo index exhibited a positive correlation with elevation in this study. As elevation increased within a specific range, species dominance and diversity also increased, consistent with previous findings [,,,]. This relationship is likely attributed to greater habitat diversity in areas with pronounced habitat heterogeneity, providing a wider array of ecological niches for organisms. Geographical location influences climatic characteristics, particularly the vertical gradients in climate that intensify with altitude, further diversifying habitat types and accommodating species with specific habitat requirements, thereby enhancing species richness and diversity []. Furthermore, soil moisture and temperature emerged as critical factors influencing plant community characteristics in the experiment. Soil moisture positively influenced canopy cover, plant height, and aboveground biomass.
The stability of grassland plant communities is influenced by diverse factors such as species composition, diversity characteristics, internal community dynamics, and broader-scale climate fluctuations []. A thorough understanding of these key mechanisms is essential for studying methods to sustain ecological balance. Additionally, exploring the appropriate scale of study and the selection of ecological factors is crucial to ensure the effectiveness and scientific validity of subsequent research endeavors.
5. Conclusions
The vegetation community characteristics of various grassland types in the Habahe Forest area exhibit notable differences. The aboveground biomass, coverage, and plant height decrease progressively from meadow to steppe. Additionally, diversity levels vary across these grassland types, with the Simpson index showing the greatest variation and the Margalef index the least. Desert steppe plant communities have significantly lower stability compared to the other three grassland types, with the highest stability observed in mountain steppes. The relationship between plant community stability and diversity across different grassland types suggests that communities with higher species richness tend to have greater stability, whereas those with higher species evenness tend to have lower stability. Water availability plays a dominant role in constraining the development of species diversity and community stability in these grasslands. To effectively maintain species diversity and stability in the grasslands of the Habahe Forest area, it is essential to consider the composition, characteristics, and influencing factors of grassland plant communities and to develop management strategies tailored to the specific field conditions.
Author Contributions
G.Z.: conceptualization, methodology, software, investigation, formal analysis, writing—original draft; M.Y.: conceptualization, funding acquisition, resources, supervision, writing—review and editing; M.L.: data curation, writing—original draft; W.C.: visualization, investigation; Q.H.: resources, supervision; X.P.: software, validation; X.Z.: visualization, writing—review and editing; J.C.: data curation, investigation; J.Q.: resources, validation; Y.L.: investigation, formal analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ecological Monitoring Analysis of Altai Mountain State Forest Management Bureau (2021), grant no. 3010010251; and the National Natural Science Foundation of China, grant no. 42377449.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors. These data are not publicly available due to ethical restrictions.
Acknowledgments
The principal author of this article is, firstly, grateful for the support from the National Natural Science Foundation of China and NSFC—Xinjiang Joint Fund, secondly grateful to the reviewers for reviewing the manuscript in their busy schedules, and finally grateful to my supervisor, M.Y, for her guidance and support of my research work.
Conflicts of Interest
The funders had no role in the design of this study; in the collection, analyses, or interpretation of the data; in the writing of the manuscript; or in the decision to publish the results.
References
- Stevens, N.; Bond, W.; Feurdean, A.; Lehmann, C.E.R. Grassy ecosystems in the Anthropocene. Annu. Rev. Env. Resour. 2022, 47, 261–289. [Google Scholar] [CrossRef]
- Zhao, G.L. Trends in grassland science: Based on the shift analysis of research themes since the early 1900s. Fundam. Res. 2023, 3, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Chomel, M.; Fry, E.L.; Johnson, D.; Lavallee, J.M.; Png, K.; Bullock, J.M.; Lavorel, S.; Manning, P.; Provost, G.L.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; Luca, E.D.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Chen, C.F. Causes of Grassland Degradation in Altai Mountains and Countermeasures for Management. Prot. For. Sci. Technol. 2022, 215, 57–59. [Google Scholar] [CrossRef]
- Amanguli, M.; Luo, J. Exploring the Causes of Grassland Degradation and Management Countermeasures in Xinjiang. Xinjiang Xu Mu Ye 2019, 34, 37–39. [Google Scholar] [CrossRef]
- Bu, E.J.; Zhao, S.; He, F.; Zhu, X.L.; Xu, D.D.; Li, X.L.; Xin, X.P. Sustainable development strategy study on Xinjiang’s grassland animal husbandry. Chin. J. Agric. Resour. Reg. Plan. 2014, 35, 120–127. [Google Scholar] [CrossRef]
- Zhang, K.L.; Ye, M.; He, Q.Z.; Yin, X.K.; Guo, J.X. Analysis of Species Diversity and VOR index of Different Grassland Types in the Habahe Region of the Alitai Mountains. J. Soil Water Conserv. 2023, 37, 262–271+279. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhao, H.L. Review on the study of vegetation stability. Chin. J. Ecol. 2003, 22, 42–48. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity–stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef]
- MacArthur, R. Fluctuations of animal populations and a measure of community stability. Ecology 1955, 36, 533–536. [Google Scholar] [CrossRef]
- Elton, C.S. The ecology of invasions by animals and plants. J. Range Manag. 1958, 47, 525. [Google Scholar]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Tilman, D. Biodiversity: Population versus ecosystem stability. Ecology 1996, 77, 350–363. [Google Scholar] [CrossRef]
- Naeem, S. Li Biodiversity enhances ecosystem reliability. Nature 1997, 390, 507–509. [Google Scholar] [CrossRef]
- McGrady, S.J.; Morin, P.J. Biodiversity, density compensation, and the dynamics of populations and functional groups. Ecology 2000, 81, 361–373. [Google Scholar] [CrossRef]
- Ives, A.R. Carpenter S R. Stability and diversity of ecosystems. Science 2007, 317, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.Y.; Zhang, L.; Liao, L.R.; Wang, J.; Lei, S.L.; Liu, G.B.; Fang, N.F.; Zhang, C. Relationship between grassland plant diversity and community stability and its driving factors on the Loess Plateau. Acta Ecol. Sin. 2023, 43, 60–69. [Google Scholar] [CrossRef]
- Gardner, M.R.; Ashby, W.B. Connectance of large dynamic (cybernetic) systems: Critical values for stability. Nature 1970, 228, 784. [Google Scholar] [CrossRef]
- Gardner, M.R. Will a large complex system be stable? Nature 1972, 238, 413–414. [Google Scholar] [CrossRef]
- Lei, S.L.; Liao, L.R.; Wang, J.; Zhang, L.; Ye, Z.C.; Liu, G.B.; Zhang, C. The diversity-Godron stability relationship of alpine grassland and its environmental drivers. Acta Pratacult. Sin. 2023, 32, 1–12. [Google Scholar] [CrossRef]
- Li, W.; Zhou, M.; Zhao, P.W.; Tian, J.L.; Wang, Z.X.; Zhao, W.; Gao, Y. Vegetation Characteristics in the Early Stage of Restoration of Burned Area in Eastern Daxing’an Mountains. J. Northeast For. Univ. 2020, 48, 51–55. [Google Scholar] [CrossRef]
- Zhen, S.Q.; Liu, J.F.; Feng, X.P.; He, Z.S.; Li, W.Z.; Li, L.; Chen, W.W.; Liu, M.X. Species Diversity and Stability of Different Plant Communities in Daiyun Mountain. J. Northwest For. Univ. 2016, 31, 50–57+64. [Google Scholar] [CrossRef]
- Yu, X.W.; Song, X.S.; Kang, F.F.; Han, H.R. Evaluation on the stability of typical forest communities in sourse region of Liaohe river in North Hebei. J. Arid. Land Resour. Environ. 2016, 31, 50–57+64. [Google Scholar] [CrossRef]
- Bi, X.L.; Hong, W.; WU, C.Z.; Yan, S.J.; Feng, L.; Wang, X.G. Reasearch on the Bio-diversity and Stability of the Rare Plant Communities. J. Fujian Coll. For. 2003, 23, 301–304. [Google Scholar] [CrossRef]
- Pinder, J.E.; Kroh, G.C.; White, J.D. Basham May AM. The relationships between vegetation type and topography in Lassen Volcanic National Park. Plant Ecol. 1997, 131, 17–29. [Google Scholar] [CrossRef]
- Zhao, T.T.; Zhao, C.Z.; Kang, M.P.; Li, Q.; Tang, Y.R.; Zeng, H.X.; Wang, Y.F. Numerical classification and ordination of shrub communities on the north slope of Qilian Mountains. Chin. J. Ecol. 2021, 40, 731–739. [Google Scholar] [CrossRef]
- O’Brien, E.M.; Field, R.; Whittaker, R.J. Climatic gradients in woody plant (tree and shrub) diversity: Water-energy dynamics, residual variation, and topography. Oikos 2000, 89, 588–600. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–259. [Google Scholar] [CrossRef]
- Zhang, H.G.; Zou, W.; Chen, Z.; He, L.J.; Peng, X.F.; Wang, G.Y.; Peng, P.H.; Li, J.J.; Shi, S.L. Distribution pattern of plant community and its relationship with environmental factors in eastern Tibet, China. Chin. J. Appl. Environ. Biol. 2023, 29, 1289–1297. [Google Scholar] [CrossRef]
- Sun, Q.Y.; Zhang, W.Q.; Zhao, L.Q.; Chi, M.F.; Luo, G.S.; Zheng, X.M.; Jia, Z.K.; Wang, Q.C. Variation of understory plant diversity and its relationship with soil moisture in Pinus tabulaeformis plantation before and after clearcutting. J. Cent. South. Univ. For. Technol. 2020, 40, 119–129. [Google Scholar] [CrossRef]
- Tan, S.S.; Ye, Z.L.; Yuan, L.B.; Zhou, R.F.; Hu, G.; Jin, X.F.; Yu, M.J. Beta diversity of plant communities in Baishanzu Nature Reserve. Acta Ecol. Sin. 2013, 33, 6944–6956. [Google Scholar] [CrossRef]
- Mendez, T.M.; Meave, J.A.; Zermeno, H.I.; Ibarra, M.G. Effects of slope aspect and topographic position on environmental variables, disturbance regime and tree community attributes in a seasonal tropical dry forest. J. Veg. Sci. 2016, 27, 1094–1103. [Google Scholar] [CrossRef]
- Shall, J.M. Influence of topography, bare sand, and soil pH on the occurrence and distribution of plant species in a lacustrine dune ecosystem. J. Torrey Bot. Soc. 2014, 141, 29–38. [Google Scholar] [CrossRef]
- Grace, J.B.; Michael, A.T.; Smith, M.D.; Seabloom, E.; Andelman, S.J.; Meche, G.; Weiher, E.; Allain, L.K.; Jutila, H.; Sankaran, M.; et al. Does species diversity limit productivity in natural grassland communities? Ecol. Lett. 2007, 10, 680–689. [Google Scholar] [CrossRef] [PubMed]
- He, Z.S.; Liu, J.F.; Zhang, S.Q.; Houng, W.; Wu, Z.Y.; Xu, D.W.; Wu, C.Z. Effects of Forest Gap Disturbance on Plant Species Diversity and Stability in Regeneration Layers of Castanopsis kawakamii Natural Forests. Plant Sci. J. 2012, 30, 133–140. [Google Scholar] [CrossRef]
- Qiao, M.; Chen, Y.N.; Zhao, X.Y. Analysis and evaluation on the agricultural morphological conditions in Haba river basin, Xinjiang. Geogr. Aird Area 1997, 2, 67–72. [Google Scholar] [CrossRef]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part one). Chin. Biodivers. 1994, 2, 229–239. [Google Scholar]
- Ma, K.P. Measuring methods of biodiversity i. Measuring methods of α diversity (part two). Chin. Biodivers. 1994, 2, 231–239. [Google Scholar]
- Pan, X.; Ye, M.; Xu, X.; He, Q.; Gu, X.; Zeng, G.; Chen, W.; Li, M. Characteristics and Relationships between Species Diversity and Productivity of Different Grassland Types in the Burqin Forest Region of the Altai Mountains. Forests 2023, 14, 1829. [Google Scholar] [CrossRef]
- Godron, M. Some aspects of heterogeneity in grasslands of Cantal. Stat. Ecol. 1972, 3, 397–415. [Google Scholar]
- Zheng, Y.R. Comparison of methods for studying stability of forest community. Sci. Silvae Sin. 2000, 5, 28–32. [Google Scholar]
- Xue, C.Y.; Li, X.G.; Tan, Z.Q.; Li, Z. Plant community stability and its relationship with species diversity of typical wetland in Poyang Lake, China. Acta Ecol. Sin. 2022, 41, 1–10. [Google Scholar] [CrossRef]
- He, F.L.; Jin, H.X.; Guo, C.X.; Ma, J.M.; Wu, H. Vegetation Composition and Community Similarity of Haloxylon ammodendron Plantations at Different Degree of Degradation on the Edge of the Minqin Oasis. J. Desert Res. 2017, 37, 1135–1141. [Google Scholar] [CrossRef]
- Ma, Q.L.; Lu, Q.; Wei, L.Y.; Jin, H.J. Varying characteristics of soil seed banks during the succession process of Nitraria tangutorum vegetation in an arid desert area. Acta Ecol. Sin. 2015, 35, 2285–2294. [Google Scholar] [CrossRef]
- Zhao, W.; Yi, Y.L.; Li, S.X.; Liu, J.J.; Dong, Y.L.; Su, S.F. Study on Vegetation Communities and Forage Nutrition Characteristics of Different Types of Grassland in Qilian Mountains. Acta Agrestia Sin. 2022, 30, 1328–1335. [Google Scholar] [CrossRef]
- Chang, H.; Sun, H.L.; Liu, Y.H.; Qiu, X.; Shi, L.; Wen, C. Community Structure and Diversity under Different Degrees of Degraded Grassland in East Wuzhumuqin Meadow. Acta Agrestia Sin. 2020, 28, 184–192. [Google Scholar] [CrossRef]
- Su, J.J.; Liu, Y.P.; Liu, L.Y.; Wu, T.Z.; Wang, M. Quantitative classification and ordination analysis of typical plant communities in Altay Prefecture of Xinjiang. Acta Pratacult. Sin. 2023, 32, 50–67. [Google Scholar] [CrossRef]
- Li, X.M.; Song, Y.B.; Jiang, W.L. Tianshan Mountains, Altai Different Types of Grass Lawn Nutrient Evaluation Group. Caoye Yu Xumu 2015, 219, 44–48. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.H. Dynamic Analysis of Community Characteristics and Diversity of Grassland in the Northern Slope of Tianshan Mountain. Xinjiang Agric. Sci. 2017, 54, 148–155. [Google Scholar] [CrossRef]
- Wang, D.W.; Sheng, J.D.; Liu, Y.H.; Zha, X.H.; Men, X.H. Analysis of grassland biomass characteristics and its impact factors in north shop of Tianshan. Pratac. Sci. 2014, 31, 125–131. [Google Scholar] [CrossRef]
- Yang, Z.P.; Ouyang, H.; Song, M.H.; Zhou, C.P.; Yang, W.B.; Liu, X.P. Species diversity and above-ground biomass of alpine vegetation in permafrost region of Qinghai–Tibetan Plateau. Chin. J. Ecol. 2010, 29, 617–623. [Google Scholar] [CrossRef]
- Huang, X.B.; Su, J.R.; Li, S.F.; Liu, W.D.; Lang, X.D. Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Sci. Rep. 2019, 9, 6979. [Google Scholar] [CrossRef]
- Zhang, J.G.; Wang, L.D.; Yao, T.; Li, H.Y.; Gao, Y.M.; Yan, X.M.; Li, C.N.; Li, Q.; Fen, Y.; Hu, Y.T. Plant community structure and species diversity differences in alpine grassland in the Qilian Mountains with different levels of degradation. Acta Pratacult. Sin. 2019, 28, 15–25. [Google Scholar] [CrossRef]
- Yin, X.K.; Ye, M.; Guo, J.X.; Zhang, K.L.; Zhao, F.F. Relationship Between Species Diversity and Productivity of Different Grassland Types in Buerjin Forest Area of Altai Mountains. J. Soil Water Conserv. 2022, 36, 110–115. [Google Scholar] [CrossRef]
- Wang, W.Y.; Wang, Q.J.; Li, S.X.; Wang, G. Distribution and species diversity of plant communities along transect on the northeastern Tibetan Plateau. Biodivers. Conserv. 2006, 15, 1811–1828. [Google Scholar] [CrossRef]
- Liu, P.X.; Lin, X.L.; Chen, L.Q. Phylogenetic structure and species diversity pattern of plant community under flooding disturbance in the riparian zone of reservoir. Acta Ecol. Sin. 2023, 43, 4700–4711. [Google Scholar] [CrossRef]
- Hu, D.; Lü, G.H.; Wang, H.F.; Yang, Q.; Cai, Y. Response of desert plant diversity and stability to soil factors based on water gradient. Acta Ecol. Sin. 2021, 41, 6738–6748. [Google Scholar] [CrossRef]
- Wu, C.H.; Cai, J.J.; Li, H.; Zhang, Y.; Kang, Y.X. Studies on diversity and stability of the Fagaceae community in the Zibai mountain. J. Cent. South Univ. For. Technol. 2021, 41, 108–115. [Google Scholar] [CrossRef]
- Gao, R.F.; Zhang, T.Y.; Bai, Y.; Cui, H.; Liu, Z.Y. Effects of Different Improvement Measures on Species Diversity and Community Productivity of Degraded Mowing Grassland. Chin. J. Grassl. 2019, 41, 98–104. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.L.; Yan, L.; Cao, X.J. Responses of alpine vegetation and soils to the disturbance of plateau pika (Ochotona curzoniae) at burrow level on the Qinghai–Tibetan Plateau of China. Ecol. Eng. 2016, 88, 232–236. [Google Scholar] [CrossRef]
- Tan, C.; Liu, T.; Liu, X.L.; Liu, C. Study on species diversity and stability of Haloxylon ammoderon community in the southwestern edge of Gurbantonggut desert. J. Arid Land Resour. Environ. 2010, 24, 148–158. [Google Scholar] [CrossRef]
- Xu, H.P.; Yu, C.; Shu, C.C.; Jing, S.H.; Pan, X.L.; Guo, Z.G. The effect of plateau pika disturbance on plant community diversity and stability in an alpine meadow. Acta Pratacult. Sin. 2019, 28, 90–99. [Google Scholar] [CrossRef]
- Hillebrand, H.; Bennett, D.B.; Cadotte, M.W. Consequences of dominance: A review of evenness effects on local and regional ecosystem processes. Ecology 2008, 89, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Huang, Y.M. Biodiversity and stability mechanisms: Understanding and future research. Acta Ecol. Sin. 2016, 36, 3859–3870. [Google Scholar] [CrossRef]
- Zhu, G.D.; Guo, N.; Han, Y.J.; Lv, G.Y.; Wang, Z.Y.; Wang, C.J. Effects of Extreme Drought on Plant Diversity and Soil Properties of Inner Mongolian Desert Steppe. Chin. J. Grassl. 2021, 43, 52–59. [Google Scholar] [CrossRef]
- Guo, J.X.; Luo, T.F.; Ye, M.; Yin, X.K.; Zhang, K.L.; Zhao, F.F. Relationship between Grassland Species Diversity and Topographical Factors in Fuyun Mining Area of Altai Mountain, Xinjiang Province. Chin. Wild Plant Resour. 2022, 41, 89–94. [Google Scholar] [CrossRef]
- Jia, Z.T.; Yang, J.Y.; Sun, Y.X.; Chen, Q.; Yan, R.L.; Li, N.N. Analysis of Species Diversity and Regulation Factors of Salsola passerine Community in Alxa Plateau. Chin. J. Grassl. 2021, 43, 1–9. [Google Scholar] [CrossRef]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guégan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; Brien, E.O.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Fan, L.L.; Li, Y.M.; Nataliia, T.; Ma, X.X.; Ma, J. Response of Herbaceous Plant Quantity to Different Water Input and Meteorological Factors in a Cold Desert. Arid Zone Res. 2019, 36, 139–146. [Google Scholar] [CrossRef]
- Du, Z.Y.; An, H.; Wang, Z.L.; Wang, B.; Zhang, X.W. Response of plant community structure and its stability to water and nitrogen addition in desert grassland. Acta Ecol. Sin. 2021, 41, 2359–2371. [Google Scholar] [CrossRef]
- Yang, H.J.; Wu, M.Y.; Liu, W.X.; Zhang, Z.; Zhang, N.L.; Wan, S.Q. Community structure and composition in response to climate change in a temperate steppe. Glob. Change Biol. 2015, 17, 452–465. [Google Scholar] [CrossRef]
- Shimono, A.; Zhou, H.; Shen, H.; Hirota, M.; Ohtsuka, T.; Tang, Y. Patterns of plant diversity at high altitudes on the Qinghai–Tibetan Plateau. J. Plant Ecol. 2010, 3, 1–7. [Google Scholar] [CrossRef]
- Dorji, T.; Moe, S.R.; Klein, J.A.; Totland, Ø. Plant species richness, evenness, and composition along environmental gradients in an alpine meadow grazing ecosystem in central Tibet, China. Arct. Antarct. Alp. Res. 2014, 46, 308–326. [Google Scholar] [CrossRef]
- Ye, P.C.; Chen, H.; Wu, J.Y.; Zhang, G.F. Distribution Pattern and Correlation with Main Environmental Factors of Higher Plant Diversity in North–West Yunnan. J. Ecol. Rural Environ. 2020, 36, 89–94. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).