
On the Diversity of Semiochemicals of the Pygidial Gland Secretions of Subterranean Ground Beetles (Coleoptera: Carabidae)


 ,
,  , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Chemical Analyses of Pygidial Gland Secretion
2.3. Morphological Analyses
3. Results
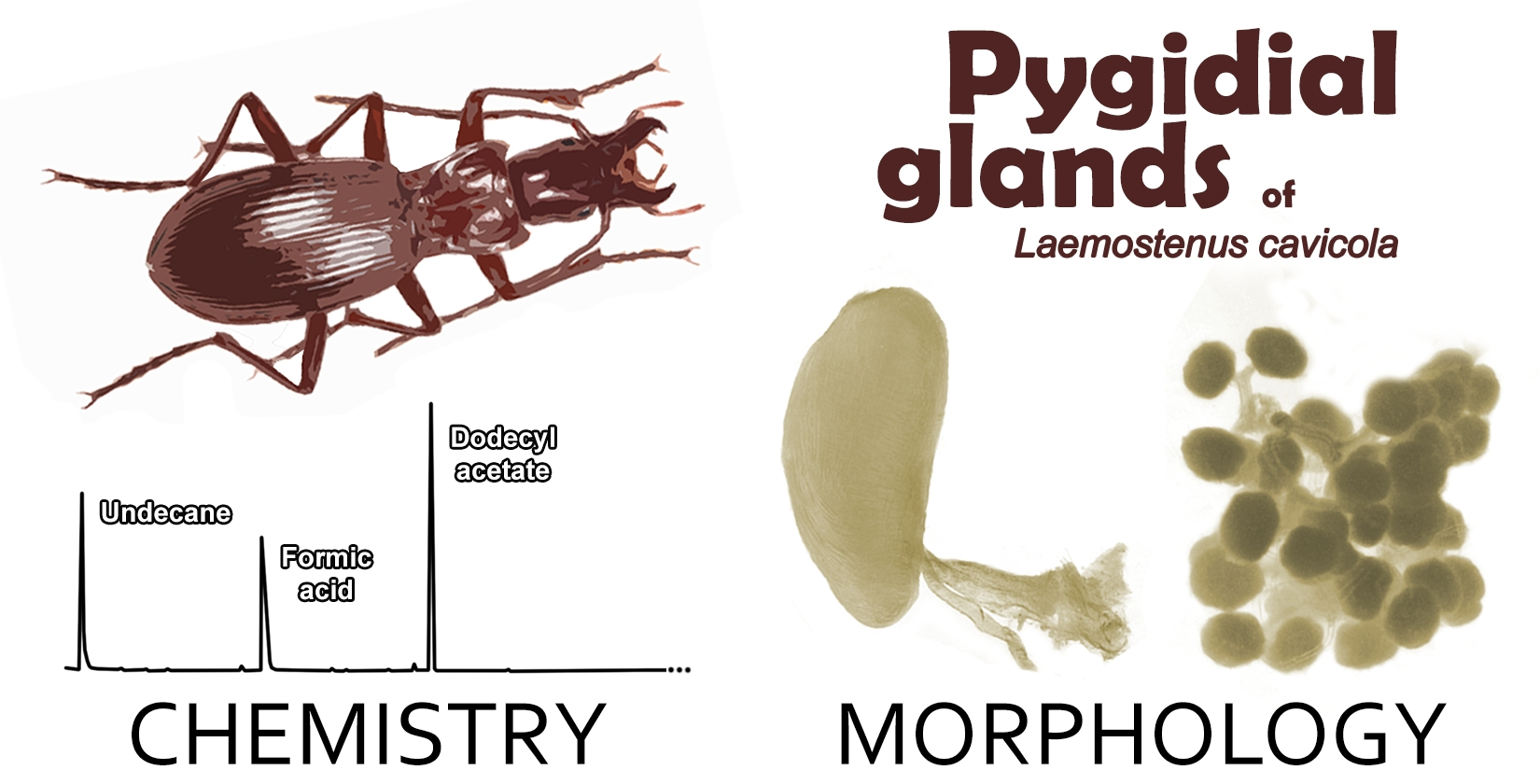
3.1. Chemical Composition of Pygidial Gland Secretion
3.2. Pygidial Gland Morphology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blum, M.S. Biochemical Defenses of Insects; Academic Press: New York, NY, USA, 1978. [Google Scholar] [CrossRef]
- Blum, M.S. Semiochemical parsimony in the Arthropoda. Annu. Rev. Entomol. 1996, 41, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.L.; Eisner, T.; Whittaker, W.H. Allomones and kairomones: Transspecific chemical messengers. BioScience 1970, 20, 21–22. [Google Scholar] [CrossRef]
- Forsyth, D.J. The structure of pygidial defence glands of Carabidae (Coleoptera). Trans. Zool. Soc. London 1972, 32, 249–309. [Google Scholar] [CrossRef]
- Kanehisa, K.; Shiraga, T. Morphological study of the pygidial defensive systems in carabid beetles. Ber. Ohara Inst. Landwirtsch. Biol. Okayama Univ. 1978, 17, 83–94. [Google Scholar]
- Rork, A.M.; Renner, T. Carabidae semiochemistry: Current and future directions. J. Chem. Ecol. 2018, 44, 1069–1083. [Google Scholar] [CrossRef]
- Nenadić, M.; Soković, M.; Glamočlija, J.; Ćirić, A.; Perić-Mataruga, V.; Ilijin, L.; Tešević, V.; Vujisić, L.; Todosijević, M.; Vesović, N.; et al. Antimicrobial activity of the pygidial gland secretion of three ground beetle species (Insecta: Coleoptera: Carabidae). Sci. Nat. 2016, 103, 34. [Google Scholar] [CrossRef]
- Nenadić, M.; Soković, M.; Glamočlija, J.; Ćirić, A.; Perić-Mataruga, V.; Tešević, V.; Vujisić, L.; Todosijević, M.; Vesović, N.; Ćurčić, S. Antimicrobial activity of the pygidial gland secretion of the troglophilic ground beetle Laemostenus (Pristonychus) punctatus (Dejean, 1828) (Insecta: Coleoptera: Carabidae). Bull. Entomol. Res. 2016, 106, 474–480. [Google Scholar] [CrossRef]
- Nenadić, M.; Ljaljević-Grbić, M.; Stupar, M.; Vukojević, J.; Ćirić, A.; Tešević, V.; Vujisić, L.; Todosijević, M.; Vesović, N.; Živković, N.; et al. Antifungal activity of the pygidial gland secretion of Laemostenus punctatus (Coleoptera: Carabidae) against cave-dwelling micromycetes. Sci. Nat. 2017, 104, 52. [Google Scholar] [CrossRef] [PubMed]
- Holliday, A.E.; Mattingly, T.M.; Toro, A.A.; Donald, L.J.; Holliday, N.J. Age- and sex-related variation in defensive secretions of adult Chlaenius cordicollis and evidence for their role in sexual communication. Chemoecology 2016, 26, 107–119. [Google Scholar] [CrossRef]
- Giglio, A.; Vommaro, M.L.; Brandmayr, P.; Talarico, F. Pygidial glands in Carabidae, an overview of morphology and chemical secretion. Life 2021, 11, 562. [Google Scholar] [CrossRef] [PubMed]
- Vesović, N. Morphological Study of the Pygidial Glands and Analysis of the Chemical Composition of the Secretions of Selected Ground Beetle Species (Insecta: Coleoptera: Carabidae). Ph.D. Thesis, Faculty of Biology, University of Belgrade, Belgrade, Serbia, 12 February 2019. (In Serbian). [Google Scholar]
- Schildknecht, H.; Maschwitz, U.; Winkler, H. Zur Evolution der Carabiden-Wehrdrüsensekrete. Über Arthropoden-Abwehrstoffe XXXII. Naturwissenschaften 1968, 55, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Will, K.W.; Gill, A.S.; Lee, H.; Attygalle, A.B. Quantification and evidence for mechanically metered release of pygidial secretions in formic acid-producing carabid beetles. J. Insect Sci. 2010, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Vesović, N.; Ćurčić, S.; Vujisić, L.; Nenadić, M.; Krstić, G.; Perić-Mataruga, V.; Milosavljević, S.; Antić, D.; Mandić, B.; Petković, M.; et al. Molecular diversity of compounds from pygidial gland secretions of cave-dwelling ground beetles: The first evidence. J. Chem. Ecol. 2015, 41, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Assmann, T.; Casale, A.; Drees, C.; Habel, J.C.; Matern, A.; Schuldt, A. The dark side of relict species biology: Cave animals as ancient lineages. In Relict Species: Phylogeography and Conservation Biology, 1st ed.; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 91–103. [Google Scholar] [CrossRef]
- Dimkić, I.; Stanković, S.; Kabić, J.; Stupar, M.; Nenadić, M.; Ljaljević-Grbić, M.; Žikić, V.; Vujisić, L.; Tešević, V.; Vesović, N.; et al. Bat guano-dwelling microbes and antimicrobial properties of the pygidial gland secretion of a troglophilic ground beetle against them. Appl. Microbiol. Biotechnol. 2020, 104, 4109–4126. [Google Scholar] [CrossRef] [PubMed]
- Casale, A. Subtribe Sphodrina Laporte, 1834. In Catalogue of Palaearctic Coleoptera. Volume 1. Archostemata–Myxophaga–Adephaga. Revised and Updated Edition; Löbl, I., Löbl, D., Eds.; Brill: Leiden, The Netherlands, 2017; pp. 773–790. [Google Scholar]
- Kanehisa, K.; Murase, M. Comparative study of the pygidial defensive systems of carabid beetles. Appl. Entomol. Zool. 1977, 12, 225–235. [Google Scholar] [CrossRef]
- Vranić, S.; Vujisić, L.; Vesović, N.; Jeremić, M.; Pantelić, D.; Todosijević, M.; Pavlović, D.; Ćurčić, N.B.; Radovanović, M.; Petrović, M.D.; et al. Secretions of pygidial defensive glands in three species of the genus Bembidion (Carabidae), and morphology of pygidial glands in B. (Peryphanes) dalmatinum. Ann. Zool. Fenn. 2022, 59, 131–147. [Google Scholar] [CrossRef]
- Blum, M.S.; Jones, T.H.; House, G.J.; Tschinkel, W.R. Defensive secretions of tiger beetles: Cyanogenetic basis. Comp. Biochem. Physiol. B 1981, 69, 903–904. [Google Scholar] [CrossRef]
- Kelley, K.C.; Schilling, A.B. Quantitative variation in chemical defense within and among subgenera of Cicindela. J. Chem. Ecol. 1998, 24, 451–472. [Google Scholar] [CrossRef]
- Pearson, D.L.; Blum, M.S.; Jones, T.H.; Fales, H.M.; Gonda, E.; Witte, B.R. Historical perspective and the interpretation of ecological patterns: Defensive compounds of tiger beetles (Coleoptera: Cicindelidae). Am. Nat. 1988, 132, 404–416. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Rt (min) | Compound | RI | RIlit | Relative Percentage (%) |
|---|---|---|---|---|---|
| 1 | 4.16 | Undecane | 1100 | 1100 | 27.0 |
| 2 | 8.79 | Acetic acid | 1465 | 1400–1498 | 0.5 |
| 3 | 9.38 | Formic acid | 1508 | 1470–1544 | 37.4 |
| 4 | 13.81 | Dodecyl formate | 1858 | - | 0.8 |
| 5 | 14.31 | Dodecyl acetate | 1901 | 1876–1900 | 33.5 |
| 6 | 16.54 | 1-Tetradecyl acetate | 2104 | 2062–2106 | 0.1 |
| 7 | 26.16 | Palmitic acid | 2935 | 2871–2954 | 0.7 |
| Compound | L. (A.) cavicola | L. (P.) punctatus | D. (P.) milutini | P. globiceps |
|---|---|---|---|---|
| Undecane | * | * | * | |
| Tridecane | * | |||
| Acetic acid | * | * | * | |
| Formic acid | * | * | ||
| Benzaldehyde | * | |||
| Propionic acid | * | |||
| Isobutyric acid | * | |||
| Butyric acid | * | |||
| Isovaleric acid | * | |||
| Decyl acetate | * | |||
| Undecyl acetate | * | |||
| Dodecyl formate | * | |||
| Dodecyl acetate | * | * | ||
| 1-Tetradecyl acetate | * | |||
| Isocaproic acid | * | |||
| 1-Dodecanol | * | |||
| Caproic acid | * | * | * | |
| p-Cresol | * | |||
| Pelargonic acid | * | |||
| Capric acid | * | |||
| 7-Hexyldocosane | * | |||
| 9-Methyltetracosane | * | |||
| Benzoic acid | * | * | ||
| Lauric acid | * | |||
| Pentacosane | * | |||
| Pentacosene | * | |||
| 3-Ethyltetracosane | * | |||
| Hexacosane | * | |||
| Myristic acid | * | * | ||
| 11-Methylheptacosane | * | |||
| Heptacosene | * | |||
| Heptacosadiene isomer 1 | * | |||
| Heptacosadiene isomer 2 | * | |||
| Octacosane | * | |||
| Palmitic acid | * | * | * | * |
| Nonacosene | * | |||
| Nonacosadiene isomer 1 | * | |||
| Nonacosadiene isomer 2 | * | |||
| Nonacosadiene isomer 3 | * | |||
| Nonacosatetraene | * | |||
| Nonacosapentaene | * | |||
| Stearic acid | * | * | * | |
| Oleic acid | * | * | * | |
| Linoleic acid | * |
| Species | Gland Structure | |||||
|---|---|---|---|---|---|---|
| Secretory Lobes | Main Collecting Canal | Reservoir | Efferent Duct | |||
| Diameter | Length | Width | Length | Width | Length | |
| L. (A.) cavicola | 0.21–0.30 | 20 | 0.05–0.08 | 1.97 | 0.88 | 1.30 |
| L. (P.) punctatus | 0.15–0.20 | 10–15 | 0.12 | 2.00 | 1.50 | 4.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vranić, S.; Vujisić, L.; Vesović, N.; Todosijević, M.; Pavićević, M.; Radović, D.; Ćurčić, S. On the Diversity of Semiochemicals of the Pygidial Gland Secretions of Subterranean Ground Beetles (Coleoptera: Carabidae). Diversity 2023, 15, 136. https://doi.org/10.3390/d15020136
Vranić S, Vujisić L, Vesović N, Todosijević M, Pavićević M, Radović D, Ćurčić S. On the Diversity of Semiochemicals of the Pygidial Gland Secretions of Subterranean Ground Beetles (Coleoptera: Carabidae). Diversity. 2023; 15(2):136. https://doi.org/10.3390/d15020136
Chicago/Turabian StyleVranić, Sofija, Ljubodrag Vujisić, Nikola Vesović, Marina Todosijević, Miloš Pavićević, Dejan Radović, and Srećko Ćurčić. 2023. "On the Diversity of Semiochemicals of the Pygidial Gland Secretions of Subterranean Ground Beetles (Coleoptera: Carabidae)" Diversity 15, no. 2: 136. https://doi.org/10.3390/d15020136
APA StyleVranić, S., Vujisić, L., Vesović, N., Todosijević, M., Pavićević, M., Radović, D., & Ćurčić, S. (2023). On the Diversity of Semiochemicals of the Pygidial Gland Secretions of Subterranean Ground Beetles (Coleoptera: Carabidae). Diversity, 15(2), 136. https://doi.org/10.3390/d15020136