
Testing the Usefulness of the Surface Collecting Method in a Vertebrate Microfossil Site from the Barremian of Spain (Los Menires, Mirambel Formation)

 , , ,
, , ,
Abstract
1. Introduction
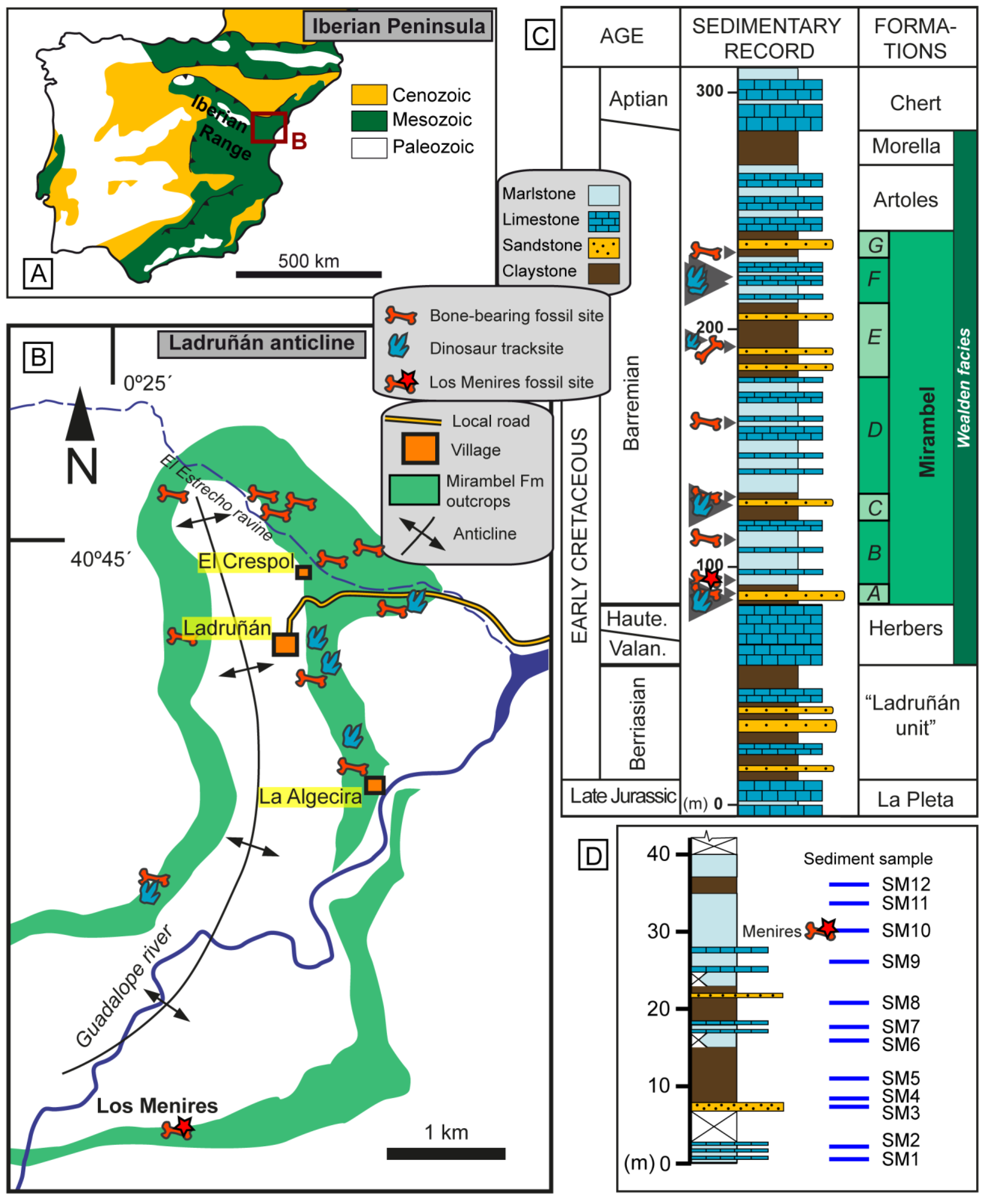
2. Geological and Palaeontological Setting
3. Materials and Methods
3.1. Selection of the Locality for the Case Study
3.2. Sampling Methodology
4. Results
4.1. Vertebrate Record: Sample Obtained by Surface Collection
4.1.1. Osteichthyes
4.1.2. Testudinata
4.1.3. Crocodylomorpha
4.1.4. Dinosaurs
4.1.5. Eggshells
4.1.6. Coprolites
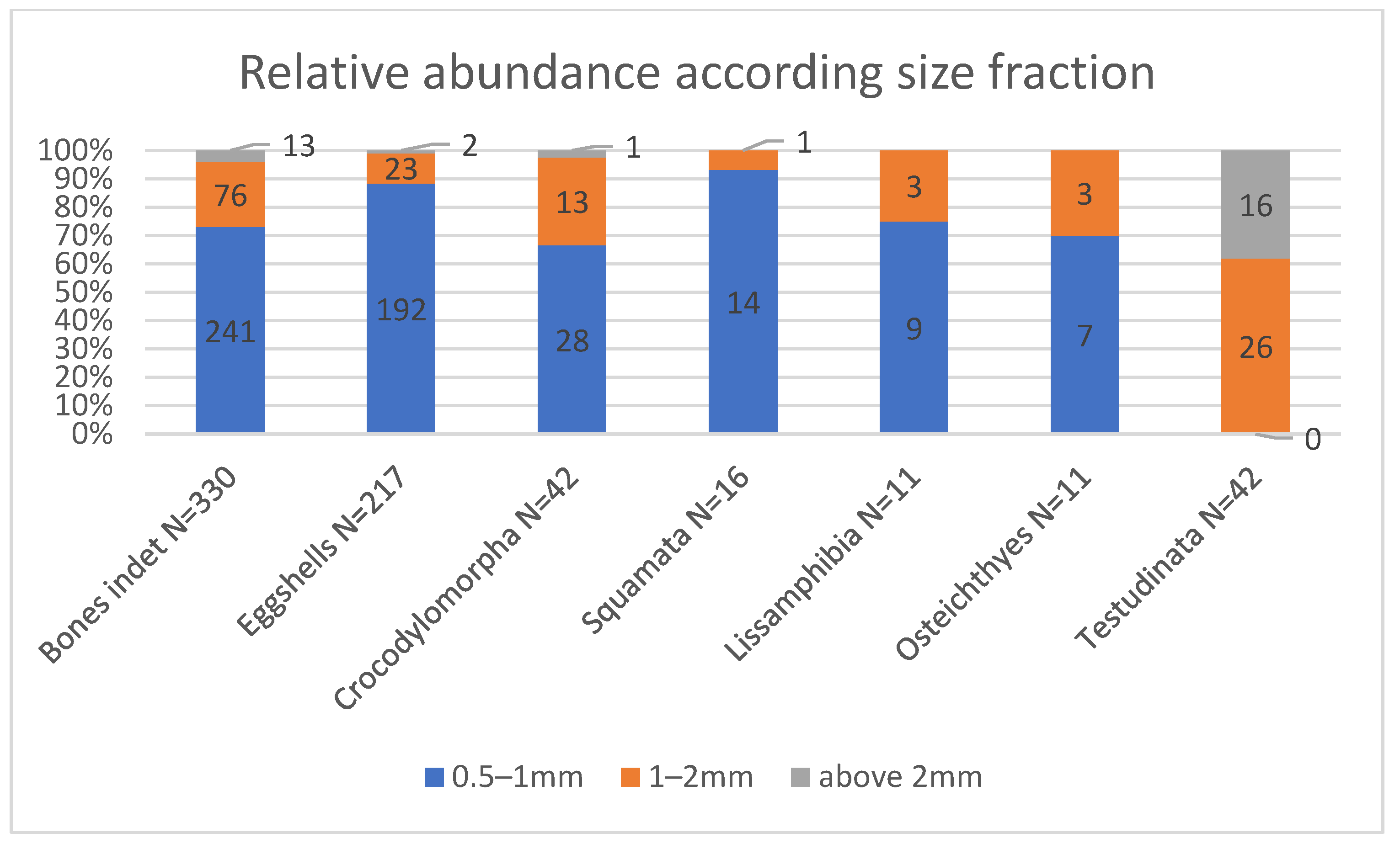
4.2. Screen-Washing Subsampling
5. Discussion
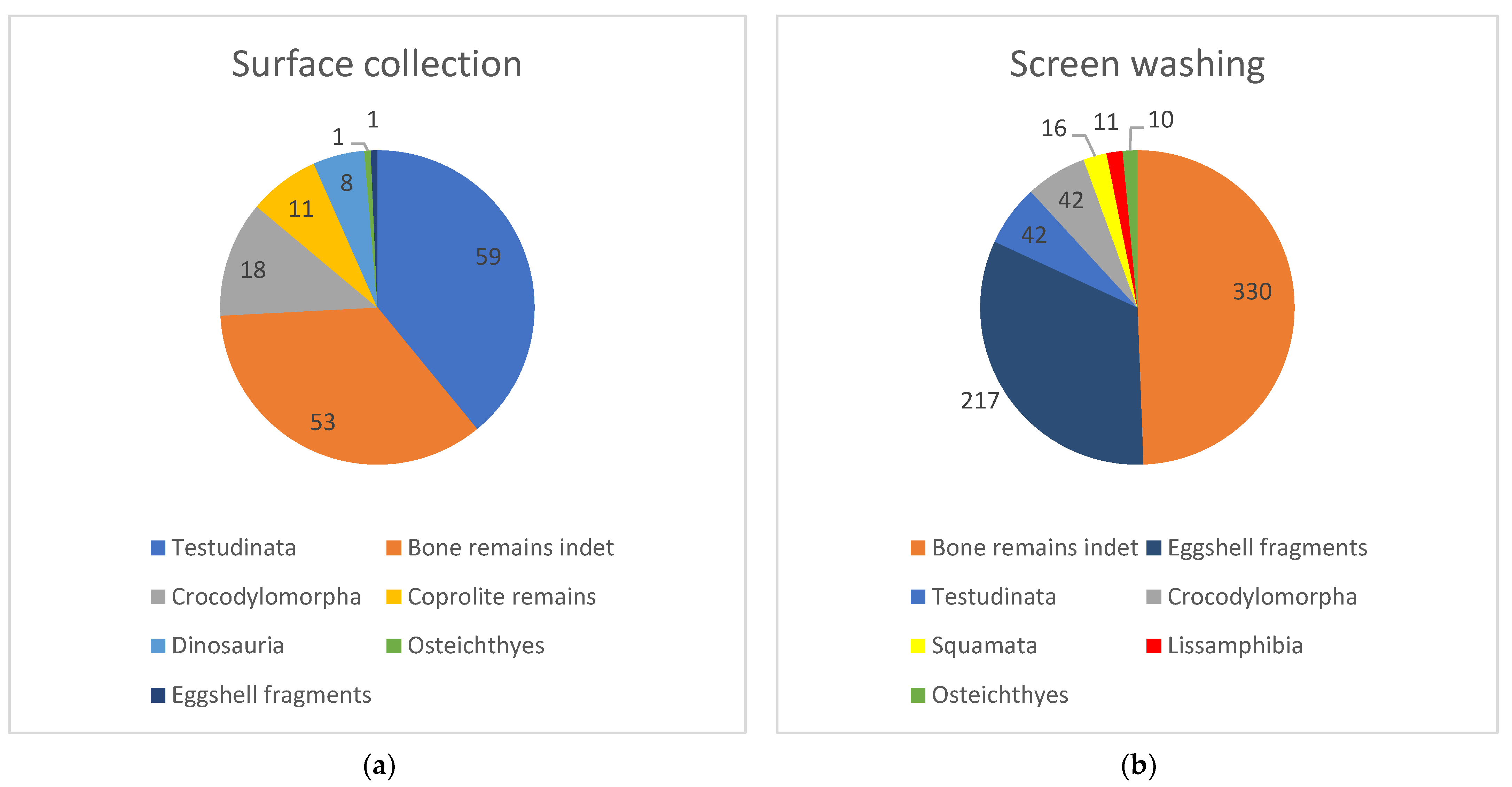
5.1. Comparison between Samples: Surface Collection vs. Screen Washing
5.2. Origin of the Microfossil Site and Palaeoecological Approach to the Biota of Los Menires
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples (2 kg) | Lithology | Stratigraphic Location (m) | Eggshells (Fragments) | Skeletal Remains | Charophytes | Ostracods | Depositional Environment |
|---|---|---|---|---|---|---|---|
| SM1 | Gray and ochre marls | 0.2 | yes (30) | no | yes | yes | Shallow lacustrine |
| SM2 | Gray and ochre marls | 2 | no | no | no | no | Alluvial-palustrine |
| SM3 | Gray sandstones | 7.5 | no | no | no | no | Fluvial channel |
| SM4 | Ochre lutites with red tones | 8.5 | no | no | no | no | Floodplain |
| SM5 | Purplish lutites | 11 | no | no | no | no | Floodplain |
| SM6 | Gray marls with red patches | 16 | no | no | no | no | Floodplain |
| SM7 | Gray marls with ochre patches | 17.5 | no | no | no | no | Floodplain |
| SM8 | Ochre lutites | 21 | yes (12) | no | yes | yes | Palustrine |
| SM9 | Dark-gray marls | 26.5 | yes (50) | yes | yes | yes | Shallow lacustrine |
| SM10 * | Very-dark-gray marls | 29.5 | yes (63) | yes | yes | yes | Shallow lacustrine |
| SM11 | Gray marls with ochre tones | 34 | yes (2) | no | yes | no | Palustrine |
| SM12 | Reddish and ochre marls | 36 | yes (5) | no | yes | no | Palustrine |
Appendix B
| Higher Taxa | Taxa | Los Menires Assemblage | Surface Sample of Los Menires |
|---|---|---|---|
| Chondrichthyes | |||
| Hybodontidae indet. | |||
| Osteichthyes | x | x | |
| Pycnodontiformes indet. | x | ||
| Lepisosteiformes indet. 1 | x | ||
| Amiiformes indet. | x | ||
| Lissamphibia | |||
| Lissamphibia indet. | x | ||
| Squamata | |||
| Squamata indet. | x | ||
| Chelonia | |||
| Helochelydridae indet. 2 | x | x | |
| Pleurosternidae indet. | x | x | |
| Crocodylomorpha | |||
| Goniopholididae indet. | x | x | |
| Atoposauridae indet. | x | ||
| Bernissartiidae indet. | x | x | |
| Ornithopoda | |||
| Styracosterna indet. | x | x | |
| Sauropoda | |||
| Titanosauriformes indet. | |||
| Theropoda | ? | ||
| Spinosauridae indet. | x | ||
| Carcharodontosauridae indet. | |||
| Maniraptoriformes indet. | x | ||
| Mammalia | |||
| Spalacotheriidae indet. | x |
References
- Brinkman, D.B. Paleoecology of the Judith River Formation (Campanian) of Dinosaur Provincial Park, Alberta, Canada: Evidence from vertebrate microfossil localities. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 78, 37–54. [Google Scholar] [CrossRef]
- Rogers, R.R.; Brady, M.E. Origins of microfossil bonebeds: Insights from the Upper Cretaceous Judith River Formation of north-central Montana. Paleobiology 2010, 36, 80–112. [Google Scholar] [CrossRef]
- Avrahami, H.M.; Gates, T.A.; Heckert, A.B.; Makovicky, P.J.; Zanno, L.E. A new microvertebrate assemblage from the Mussentuchit Member, Cedar Mountain Formation: Insights into the paleobiodiversity and paleobiogeography of early Late Cretaceous ecosystems in western North America. PeerJ 2018, 6, e5883. [Google Scholar] [CrossRef] [PubMed]
- Badgley, C. Taphonomy of mammalian fossil remains from Siwalik rocks of Pakistan. Paleobiology 1986, 12, 119–142. [Google Scholar] [CrossRef]
- Jamniczky, H.A.; Brinkman, D.B.; Russell, A.P. Vertebrate microsite sampling: How much is enough? J. Vertebr. Paleontol. 2003, 23, 725–734. [Google Scholar] [CrossRef]
- Oreska, M.P.; Carrano, M.T.; Dzikiewicz, K.M. Vertebrate paleontology of the Cloverly Formation (Lower Cretaceous), I: Faunal composition, biogeographic relationships, and sampling. J. Vertebr. Paleontol. 2013, 33, 264–292. [Google Scholar] [CrossRef]
- Forcino, F.L.; Stafford, E.S. The influence of collection method on paleoecological datasets: In-place versus surface-collected fossil samples in the Pennsylvanian Finis Shale, Texas, USA. PLoS ONE 2020, 15, e0228944. [Google Scholar] [CrossRef]
- Rogers, R.R.; Carrano, M.T.; Rogers, K.A.C.; Perez, M.; Regan, A.K. Isotaphonomy in concept and practice: An exploration of vertebrate microfossil bonebeds in the Upper Cretaceous (Campanian) Judith River Formation, north-central Montana. Paleobiology 2017, 43, 248–273. [Google Scholar] [CrossRef]
- Sankey, J.T.; Baszio, S. (Eds.) Vertebrate Microfossil Assemblages: Their Role in Paleoecology and Paleobiogeography. Life of the Past; Indiana University Press: Bloomington, IN, USA, 2008; 296p. [Google Scholar]
- Richter, G.; Teichmüller, R. 1933. Die Entwicklung der Keltiberischen Ketten; Weismannsche Buchhandlung: Berlin, Germany, 1933; 118p. [Google Scholar]
- Salas, R.; Guimerà, J.; Más, R.; Martín-Closas, C.; Meléndez, A.; Alonso, A. Evolution of the Mesozoic central Iberian Rift System and its Cainozoic inversion (Iberian Chain). Mem. Natl. Mus. Nat. Hist. 2001, 186, 145–185. [Google Scholar]
- Nebot, M.; Guimerà, J. Kinematic evolution of a fold-and-thrust belt developed during basin inversion: The Mesozoic Maestrat basin, E Iberian Chain. Geol. Mag. 2018, 155, 630–640. [Google Scholar] [CrossRef]
- García-Penas, A.; Aurell, M.; Zamora, S. Progressive opening of a shallow-marine bay (Oliete Subbasin, Spain) and the record of possible eustatic fall events near the Barremian-Aptian boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 594, 110938. [Google Scholar] [CrossRef]
- Gasca, J.M.; Moreno-Azanza, M.; Bádenas, B.; Díaz-Martínez, I.; Castanera, D.; Canudo, J.I.; Aurell, M. Integrated overview of the vertebrate fossil record of the Ladruñán anticline (Spain): Evidence of a Barremian alluvial-lacustrine system in NE Iberia frequented by dinosaurs. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 472, 192–202. [Google Scholar] [CrossRef]
- Eberth, D.A.; Rogers, R.R.; Fiorillo, A.R. A practical approach to the study of bonebeds. In Bonebeds: Genesis, Analysis, and Paleobiological Significance; Rogers, R.R., Eberth, D.A., Fiorillo, A.R., Eds.; University of Chicago Press: Chicago, IL, USA, 2007; pp. 103–220. [Google Scholar]
- Moreno-Azanza, M.; Canudo, J.I.; Gasca, J.M. Enigmatic Early Cretaceous ootaxa from Western Europe with signals of extrinsic eggshell degradation. Cretac. Res. 2015, 56, 617–627. [Google Scholar] [CrossRef]
- Cifelli, R.L. Techniques for Recovery and Preparation of Microvertebrate Fossils; Oklahoma Geological Survey; Oklahoma Geological Survey Special Publication: Norman, OK, USA, 1996; p. 36. [Google Scholar]
- López-Arbarello, A.; Sferco, E. New semionotiform (Actinopterygii: Neopterygii) from the Late Jurassic of southern Germany. J. Syst. Palaeontol. 2011, 9, 197–215. [Google Scholar] [CrossRef]
- De la Fuente, M.S.; Maniel, I.; Jannello, J.M.; Sterli, J.; Garrido, A.C.; Garcia, R.A.; Salgado, L.; Canudo, J.I.; Bolatti, R. Unusual shell anatomy and osteohistology in a new Late Cretaceous panchelid turtle from northwestern Patagonia, Argentina. Acta Palaeontol. Pol. 2017, 62, 585–601. [Google Scholar] [CrossRef]
- Joyce, W.G.; Rabi, M.; Clark, J.M.; Xu, X. A toothed turtle from the Late Jurassic of China and the global biogeographic history of turtles. BMC Evol. Biol. 2016, 16, 236. [Google Scholar] [CrossRef]
- Joyce, W.G.; Chapman, S.D.; Moody, R.T.; Walker, C.A. The skull of the solemydid turtle Helochelydra nopcsai from the Early Cretaceous of the Isle of Wight (UK) and a review of Solemydidae. Spec. Pap. Palaeontol. 2011, 86, 75–97. [Google Scholar] [CrossRef]
- Scheyer, T.M.; Pérez-García, A.; Murelaga, X. Shell bone histology of solemydid turtles (stem Testudines): Palaeoecological implications. Org. Divers. Evol. 2015, 15, 199–212. [Google Scholar] [CrossRef]
- De la Fuente, M.S.; Barbieri, R.; Chafrat, P. Una tortuga Chelidae (Testudines: Pleurodira) de cuello largo en el Grupo Neuquén, Río Negro, Argentina: Significado cronológico y paleobiogeográfico. Andean Geol. 2010, 37, 398–412. [Google Scholar] [CrossRef]
- Canudo, J.I.; Gasca, J.M.; Aurell, M.; Badiola, A.; Blain, H.-A.; Cruzado-Caballero, P.; Gómez-Fernández, D.; Moreno-Azanza, M.; Parrilla, J.; Rabal-Garcés, R.; et al. La Cantalera: An exceptional window onto the vertebrate biodiversity of the Hauterivian-Barremian transition in the Iberian Peninsula. J. Iber. Geol. 2010, 36, 295–324. [Google Scholar] [CrossRef]
- Puértolas-Pascual, E.; Canudo, J.I.; Sender, L.M. New material from a huge specimen of Anteophthalmosuchus cf. escuchae (Goniopholididae) from the Albian of Andorra (Teruel, Spain): Phylogenetic implications. J. Iber. Geol. 2015, 41, 41–56. [Google Scholar] [CrossRef]
- Buffetaut, E.; Ford, R.L.E. The crocodilian Bernissartia in the Wealden of the Isle of Wight. Palaeontology 1979, 22, 905–912. [Google Scholar]
- Sweetman, S.C.; Pedreira-Segade, U.; Vidovic, S.U. A new bernissartiid crocodyliform from the Lower Cretaceous Wessex Formation (Wealden Group, Barremian) of the Isle of Wight, southern England. Acta Palaeontol. Pol. 2015, 60, 257–268. [Google Scholar] [CrossRef]
- Brinkmann, W. Die krokodilier-fauna aus der Unter-Kreide (Ober-Barremium) von Uña (Provinz Cuenca, Spanien). Berl. Geowiss. Abh. 1992, 5, 1–123. [Google Scholar]
- Schwarz, D.; Salisbury, S.W. A new species of Theriosuchus (Atoposauridae, Crocodylomorpha) from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal. Geobios 2005, 38, 779–802. [Google Scholar] [CrossRef]
- Gasca, J.M.; Canudo, J.I.; Moreno-Azanza, M. On the diversity of Iberian iguanodont dinosaurs: New fossils from the lower Barremian, Teruel province, Spain. Cretac. Res. 2014, 50, 264–272. [Google Scholar] [CrossRef]
- Medrano-Aguado, E.; Parrilla-Bel, J.; Gasca, J.M.; Alonso, A.; Canudo, J.I. Ornithopod palaeobiodiversity in the Barranco del Hocino-1 site, from the upper Barremian in the Oliete subbasin (Teruel, Spain). Earth Sci. J. Procedia 2021, 1, 58–61. [Google Scholar] [CrossRef]
- Hunt, A.P.; Lucas, S.G. Descriptive terminology of coprolites and recent feces. Vertebrate Coprolites. N. M. Mus. Nat. Hist. Sci. Bull. 2012, 57, 153–160. [Google Scholar]
- Milàn, J.; Rasmussen, E.S.; Dybkjær, K. A crocodilian coprolite from the lower Oligocene Viborg Formation of Sofienlund Lergrav, Denmark. Bull. Geol. Soc. Den. 2018, 66, 181–187. [Google Scholar] [CrossRef]
- Milàn, J. Crocodylian scatology–A look into morphology, internal architecture, inter- and intraspecific variation and prey remains in extant crocodylian feces. New Mex. Mus. Nat. Hist. Sci. Bull. 2012, 57, 65–72. [Google Scholar]
- Schwarz, D.; Raddatz, M.; Wings, O. Knoetschkesuchus langenbergensis gen. nov. sp. nov., a new atoposaurid crocodyliform from the Upper Jurassic Langenberg Quarry (Lower Saxony, northwestern Germany), and its relationships to Theriosuchus. PLoS ONE 2017, 12, e0160617. [Google Scholar] [CrossRef]
- Guillaume, A.R.; Moreno-Azanza, M.; Puértolas-Pascual, E.; Mateus, O. Palaeobiodiversity of crocodylomorphs from the Lourinhã Formation based on the tooth record: Insights into the palaeoecology of the Late Jurassic of Portugal. Zool. J. Linn. Soc. 2020, 189, 549–583. [Google Scholar] [CrossRef]
- Lasseron, M.; Allain, R.; Gheerbrant, E.; Haddoumi, H.; Jalil, N.E.; Métais, G.; Rage, J.-C.; Vullo, R.; Zouhri, S. New data on the microvertebrate fauna from the Upper Jurassic or lowest Cretaceous of Ksar Metlili (Anoual Syncline, eastern Morocco). Geol. Mag. 2020, 157, 367–392. [Google Scholar] [CrossRef]
- Gasca, J.M.; Badiola, A.; Canudo, J.I.; Moreno-Azanza, M.; Puértolas, E. The fossil vertebrate assemblage from the Pochancalo 1 site (Valanginian- Hauterivian, Villanueva de Huerva, Zaragoza, Spain). In Actas de V Jornadas Internacionales Sobre Paleontología de Dinosaurios y su Entorno; Colectivo Arquelógico y Paleontológico Salense: Burgos, Spain, 2012; pp. 159–172. [Google Scholar]
- Pochat-Cottilloux, Y.; Allain, R.; Lasseron, M. Microvertebrate fauna from Gadoufaoua (Niger, Aptian, Early Cretaceous). Comptes Rendus Palevol 2022, 21, 901–926. [Google Scholar] [CrossRef]
- Behrensmeyer, A.K. Bonebeds through time. In Bonebeds: Genesis, Analysis, and Paleobiological Significance; Rogers, R.R., Eberth, D.A., Fiorillo, A.R., Eds.; University of Chicago Press: Chicago, IL, USA, 2007; pp. 65–101. [Google Scholar]
- Pérez-García, A. The Iberian fossil record of turtles: An update. J. Iber. Geol. 2017, 43, 155–191. [Google Scholar] [CrossRef]
- Buscalioni, A.D.; Chamero, B. Crocodylomorpha. In Las Hoyas: A Cretaceous Wetland.: A Multidisciplinary Synthesis after 25 Years of Research on an Exceptional Fossil Lagerstätte from Spain; Friedrich Verlag: Hanover, Germany, 2016; pp. 162–169. [Google Scholar]
- Buffetaut, E. Radiation évolutive, paléoécologie et biogéographie des crocodiliens mésosuchiens. Mem. Soc. Géol. France 1982, 142, 1–88. [Google Scholar]
- Pouech, J.; Amiot, R.; Lécuyer, C.; Mazin, J.M.; Martineau, F.; Fourel, F. Oxygen isotope composition of vertebrate phosphates from Cherves-de-Cognac (Berriasian, France): Environmental and ecological significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 410, 290–299. [Google Scholar] [CrossRef]
- Ruiz-Omeñaca, J.I.; Canudo, J.I.; Cuenca-Bescós, G. Primera evidencia de un área de alimentación de dinosaurios herbívoros en el Cretácico Inferior de España (Teruel). Monogr. Acad. Cienc. Exactas Físicas Químicas Nat. Zaragoza 1997, 10, 1–48. [Google Scholar]
- Moreno-Azanza, M.; Canudo, J.I.; Gasca, J.M. Spheroolithid eggshells in the Lower Cretaceous of Europe. Implications for eggshell evolution in ornithischian dinosaurs. Cretac. Res. 2014, 51, 75–87. [Google Scholar] [CrossRef]
- Burton, T.M.; Tiner, R. Ecology of wetlands. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 507–515. [Google Scholar] [CrossRef]
- Behrensmeyer, A.K.; Chapman, R.E. Models and simulations of time-averaging in terrestrial vertebrate accumulations. Short Courses Paleontol. 1993, 6, 125–149. [Google Scholar] [CrossRef]
- Martin, R.E. Taphonomy: A Process Approach; Cambridge Paleobiology Series; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar] [CrossRef]
- Castanera, D.; Díaz-Martínez, I.; Moreno-Azanza, M.; Canudo, J.I.; Gasca, J.M. An overview of the Lower Cretaceous dinosaur tracksites from the Mirambel Formation in the Iberian Range (NE Spain). Cretac. Period: Biot. Divers. Biogeogr. 2016, 71, 65–74. [Google Scholar]






| Number | Element | Higher Taxonomic Group | Taxa | Labels | Size (Maximum Dimension) |
|---|---|---|---|---|---|
| 56 | Plate fragment | Testudinata | Pleurosternidae and Helochelydidae | MENI-A-1 to 56 | Up to 20 mm |
| 54 | Undetermined bone fragment | Vertebrata | - | MENI-A-57 to 109 and 148 | From 20 to 41 mm |
| 11 | Coprolite fragment | - | - | MENI-A-110 to 120 | Up to 23 mm |
| 10 | Osteoderm fragment | Crocodylomorpha | - | MENI-A-121 to 130 | Up to 12 mm |
| 7 | Tooth | Ornithopoda | Styracosterna | MENI-A-131 to 137 | More than 9 mm |
| 5 | Tooth | Crocodylomorpha | Goniopholididae | MENI-A-138 to 142 | From 4.6 to 9.7 mm |
| 3 | Tooth | Crocodylomorpha | Bernissartiidae? | MENI-A-143 to 145 | From 3.7 to 4.5 mm |
| 1 | Vertebrae | Crocodylomorpha | - | MENI-A-146 | 8.7 mm |
| 1 | Vertebrae | Testudinata | - | MENI-A-147 | 5.4 mm |
| 1 | Scale | Osteichthyes | - | MENI-A-149 | 7.5 mm |
| 1 | Tooth fragment | Dinosauria | Saurischia | MENI-A-150 | 15.4 mm |
| 1 | Eggshell fragment | - | MENI-A-151 | 6.5 mm |
| Sample and Size (kg) | Mesh Size (mm) | Osteichthyes | Lissamphibia | Squamata | Testudinata | Crocodylomorpha | Bone Remains Indet | Eggshell Fragments |
|---|---|---|---|---|---|---|---|---|
| A (3.372) | 0.5 | 4 | 0 | 0 | 0 | 5 | 31 | 24 |
| 1 | 1 | 2 | 1 | 0 | 7 | 19 | 3 | |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total A | 5 | 1 | 2 | 0 | 13 | 53 | 27 | |
| B (2.911) | 0.5 | 2 | 0 | 3 | 0 | 9 | 55 | 40 |
| 1 | 1 | 1 | 0 | 15 | 4 | 11 | 5 | |
| 2 | 0 | 0 | 0 | 4 | 0 | 1 | 0 | |
| Total B | 3 | 1 | 3 | 19 | 13 | 67 | 45 | |
| C (2.680) | 0.5 | 0 | 2 | 3 | 0 | 3 | 50 | 47 |
| 1 | 0 | 0 | 0 | 2 | 0 | 19 | 3 | |
| 2 | 0 | 0 | 0 | 5 | 0 | 0 | 2 | |
| Total C | 0 | 2 | 3 | 7 | 3 | 69 | 52 | |
| D (3.300) | 0.5 | 1 | 0 | 6 | 0 | 8 | 55 | 43 |
| 1 | 1 | 0 | 0 | 4 | 1 | 13 | 3 | |
| 2 | 0 | 0 | 0 | 2 | 0 | 9 | 0 | |
| Total D | 1 | 0 | 6 | 6 | 9 | 77 | 46 | |
| E (2.700) | 0.5 | 0 | 7 | 2 | 0 | 3 | 50 | 38 |
| 1 | 0 | 0 | 0 | 5 | 1 | 14 | 9 | |
| 2 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | |
| Total E | 0 | 7 | 2 | 10 | 4 | 64 | 47 | |
| TOTAL (14.963) | 0.5 | 7 | 9 | 14 | 0 | 28 | 241 | 192 |
| 1 | 3 | 3 | 1 | 26 | 13 | 76 | 23 | |
| 2 | 0 | 0 | 0 | 16 | 1 | 13 | 2 | |
| All remains * | 10 | 12 | 15 | 42 | 42 | 330 | 217 |
| Sampling Methods: | Surface Collection | Screen Washing | ||
|---|---|---|---|---|
| Rank | Fossil Category | Number of Specimens | Fossil Category | Number of Specimens |
| 1 | Testudinata | 59 | Bone remains indet | 330 |
| 2 | Bone remains indet | 53 | Eggshell fragments | 217 |
| 3 | Crocodylomorpha | 18 | Testudinata | 42 |
| 4 | Coprolite remains | 11 | Crocodylomorpha | 42 |
| 5 | Dinosauria | 8 | Squamata | 16 |
| 6 | Osteichthyes | 1 | Lissamphibia | 11 |
| 7 | Eggshell fragments | 1 | Osteichthyes | 10 |
| - | Squamata | 0 | Coprolite remains | 0 |
| - | Lissamphibia | 0 | Dinosauria | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gasca, J.M.; Parrilla-Bel, J.; Moreno-Azanza, M.; Navarro-Lorbés, P.; Canudo, J.I. Testing the Usefulness of the Surface Collecting Method in a Vertebrate Microfossil Site from the Barremian of Spain (Los Menires, Mirambel Formation). Diversity 2023, 15, 135. https://doi.org/10.3390/d15020135
Gasca JM, Parrilla-Bel J, Moreno-Azanza M, Navarro-Lorbés P, Canudo JI. Testing the Usefulness of the Surface Collecting Method in a Vertebrate Microfossil Site from the Barremian of Spain (Los Menires, Mirambel Formation). Diversity. 2023; 15(2):135. https://doi.org/10.3390/d15020135
Chicago/Turabian StyleGasca, José M., Jara Parrilla-Bel, Miguel Moreno-Azanza, Pablo Navarro-Lorbés, and José I. Canudo. 2023. "Testing the Usefulness of the Surface Collecting Method in a Vertebrate Microfossil Site from the Barremian of Spain (Los Menires, Mirambel Formation)" Diversity 15, no. 2: 135. https://doi.org/10.3390/d15020135
APA StyleGasca, J. M., Parrilla-Bel, J., Moreno-Azanza, M., Navarro-Lorbés, P., & Canudo, J. I. (2023). Testing the Usefulness of the Surface Collecting Method in a Vertebrate Microfossil Site from the Barremian of Spain (Los Menires, Mirambel Formation). Diversity, 15(2), 135. https://doi.org/10.3390/d15020135