
The Complete Mitochondrial Genome of Glischropus bucephalus (Vespertilionidae; Chiroptera) Provides New Evidence for Pipistrellus Paraphyly

 , , and
, , and 
Abstract
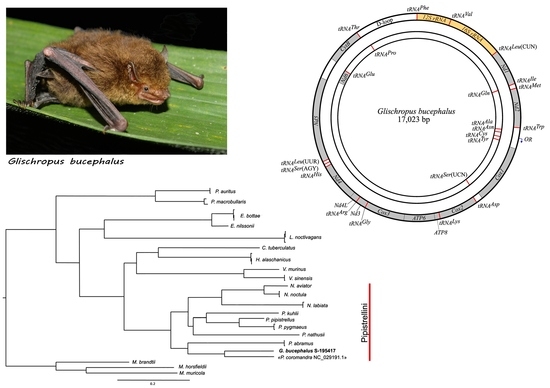
1. Introduction
2. Materials and Methods
2.1. DNA Extraction and Amplification
2.2. Mitogenome Assembling and Annotation
2.3. Phylogenetic Reconstruction
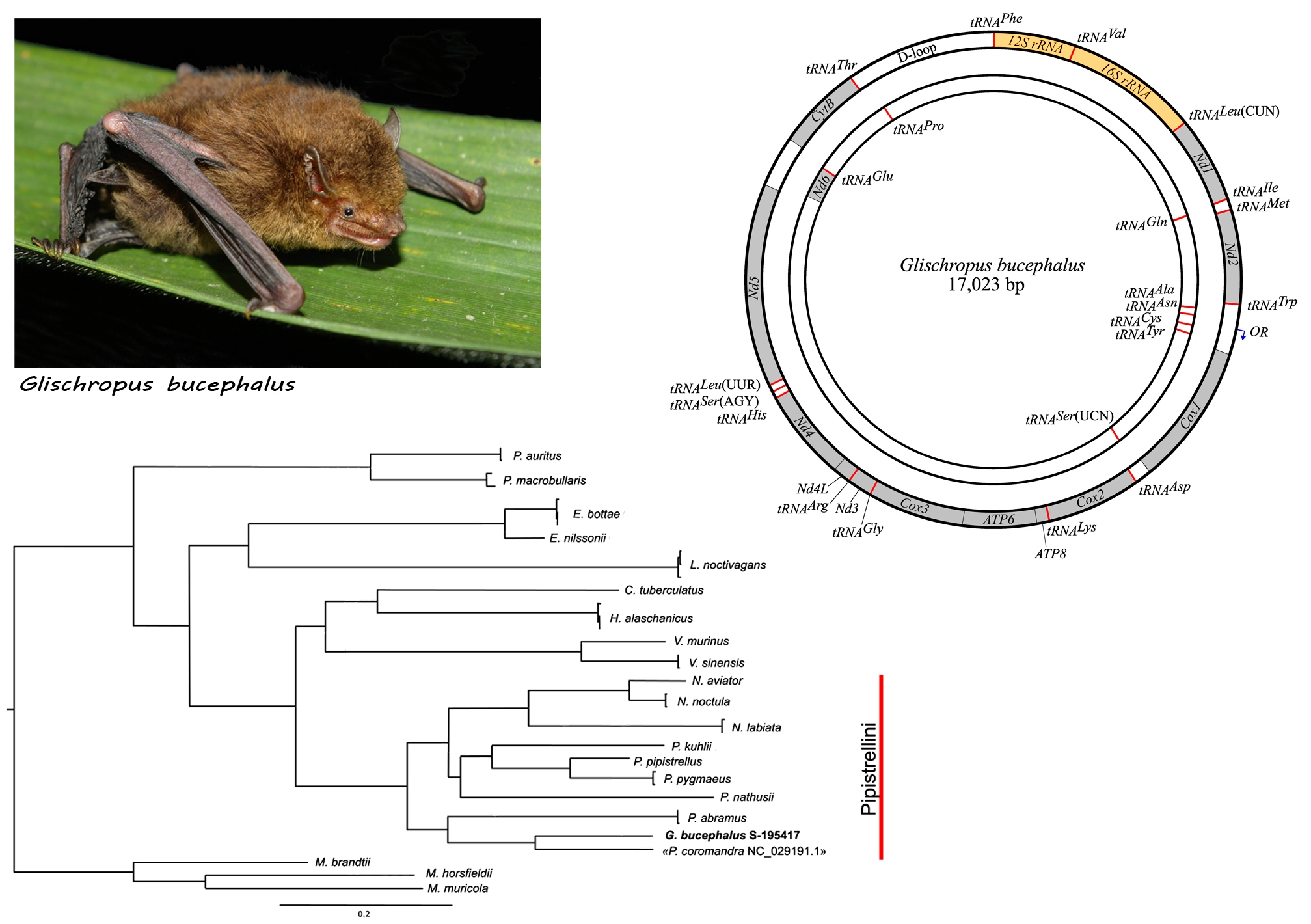
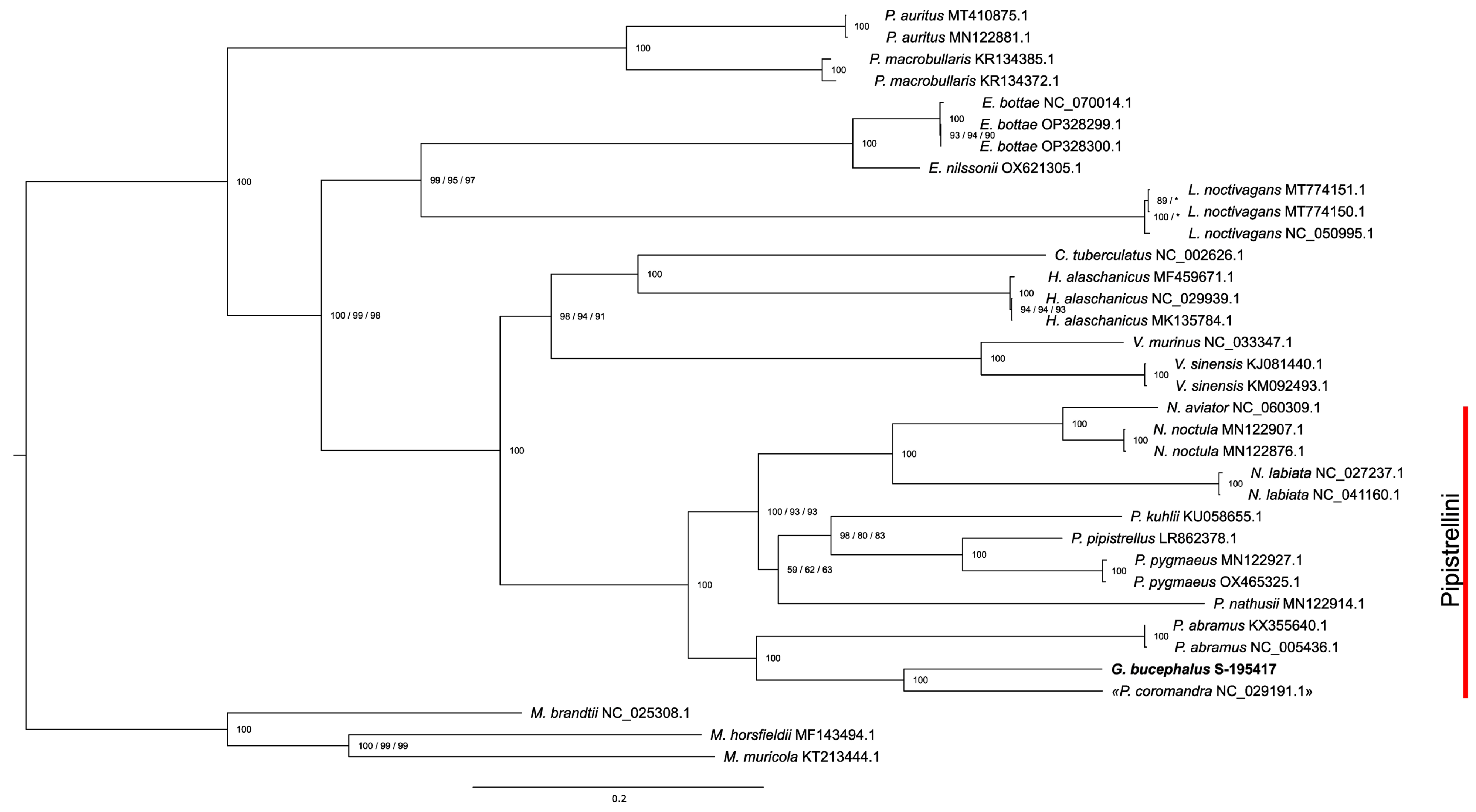
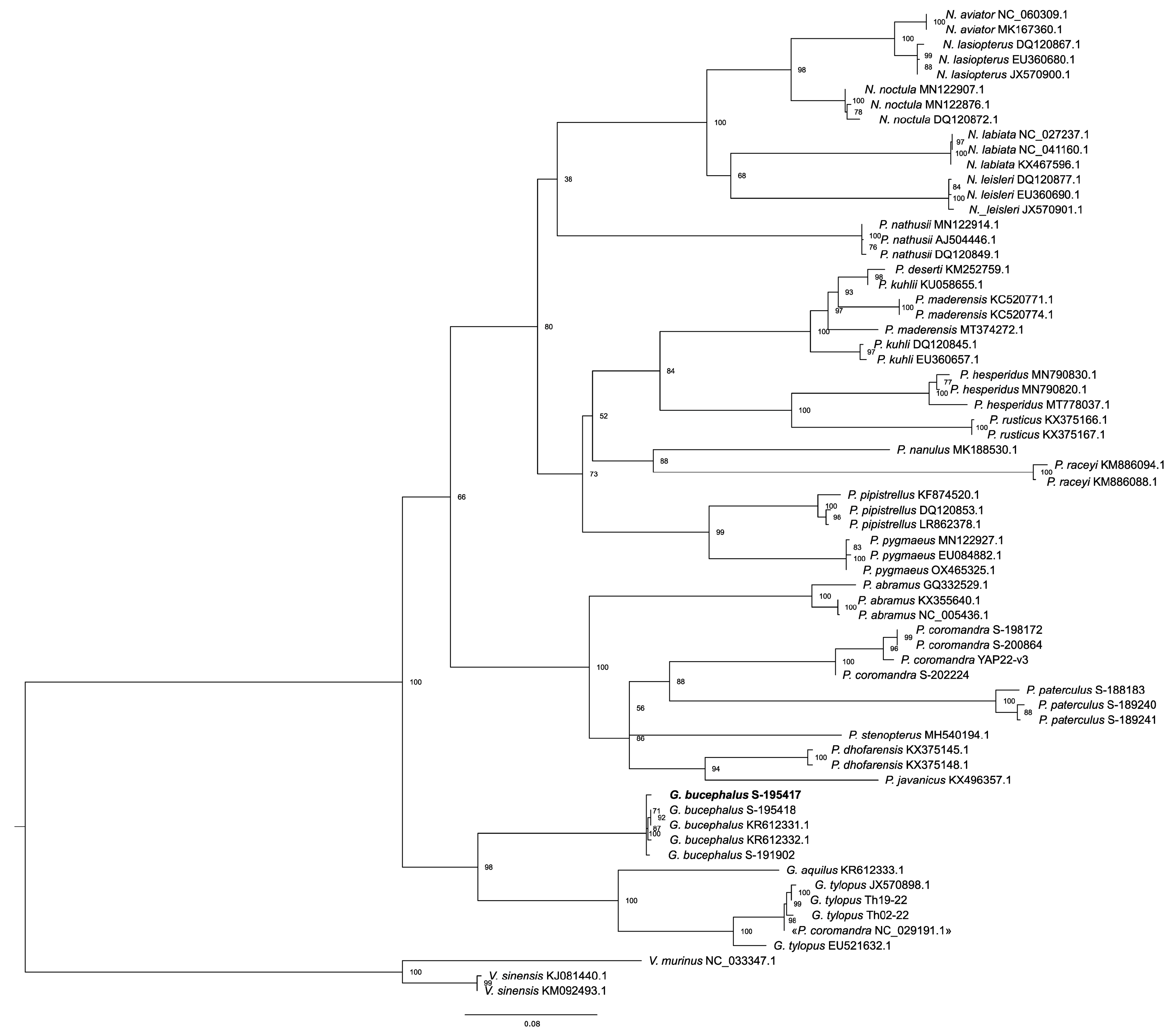
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, G.E. Descriptions of new or little-known species of bats of the genus Vesperugo. Proc. Zool. Soc. Lond. 1875, 470–474. [Google Scholar]
- Tate, G.H.H. Results of the Archbold Expeditions 47. Review of the vespertilionine bats, with special attention to genera and species of the Archbold Expeditions. Bull. Am. Mus. Nat. Hist. 1942, 80, 221–297. [Google Scholar]
- Csorba, G. A new species of Glischropus from the Indochinese Subregion (Mammalia: Chiroptera: Vespertilionidae). Zootaxa 2011, 2925, 41–48. [Google Scholar] [CrossRef]
- Csorba, G.; Görföl, T.; Wiantoro, S.; Kingston, T.; Bates, P.J.J.; Huang, J.C. Thumb-pads up—A new species of thickthumbed bat from Sumatra (Chiroptera: Vespertilionidae: Glischropus). Zootaxa 2015, 3980, 267–278. [Google Scholar] [CrossRef]
- Saikia, U.; Ruedi, M.; Csorba, G. Out of Southeast Asia: A new species of thick-thumbed bat (Chiroptera: Vespertilionidae: Glischropus) from Meghalaya, north-eastern India. Zootaxa 2022, 5154, 355–364. [Google Scholar] [CrossRef]
- Miller, G.S. The Families and Genera of Bats; Government Printing Office: Washington, DC, USA, 1907; pp. 1–282.
- Volleth, M.; Bronner, G.; Göpfert, M.C.; Heller, K.-G.; von Helversen, O.; Yong, H.-S. Karyotype comparison and phylogenetic relationships of Pipistrellus-like bats (Vespertilionidae; Chiroptera; Mammalia). Chromosome Res. 2001, 9, 25–46. [Google Scholar] [CrossRef] [PubMed]
- Heaney, L.R.; Balete, D.S.; Alviola, P.; Rickart, E.A.; Ruedi, M. Nyctalus plancyi and Falsistrellus petersi (Chiroptera: Vespertilionidae) from northern Luzon, Philippines: Ecology, phylogeny, and biogeographic implications. Acta Chiropterol. 2012, 14, 265–278. [Google Scholar] [CrossRef]
- Simmons, N.B. Order Chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; The Johns Hopkins Univ. Press: Baltimore, MD, USA, 2005; pp. 312–529. [Google Scholar]
- Zhukova, S.S.; Solovyeva, E.N.; Artyushin, I.V.; Kruskop, S.V. Paraphyly of the Pipistrelles (Pipistrellus; Vespertilionidae) is confirmed by the analysis of the nuclear gene markers. Doklady Biochem. Biophys. 2022, 507, 302–306. [Google Scholar] [CrossRef]
- Mackiewicz, P.; Matosiuk, M.; Świsłocka, M.; Zachos, F.E.; Hajji, G.M.; Saveljev, A.P.; Seryodkin, I.V.; Farahvash, T.; Rezaei, H.R.; Torshizi, R.V.; et al. Phylogeny and evolution of the genus Cervus (Cervidae, Mammalia) as revealed by complete mitochondrial genomes. Sci. Rep. 2022, 12, 16381. [Google Scholar] [CrossRef]
- Kruskop, S.V. Bats of Vietnam. Checklist and an Identification Manual, 2nd ed.; Revised and Supplemented; KMK Scientific Press: Moscow, Russia, 2013; pp. 1–300. [Google Scholar]
- Hassanin, A.; Hugot, J.-P.; van Vuuren, B.J. Comparison of mitochondrial genome sequences of pangolins (Mammalia, Pholidota). Comptes Rendus Biol. 2015, 338, 260–265. [Google Scholar] [CrossRef]
- Zhang, Y.-C.; Lwin, Y.H.; Li, R.; Maung, K.-W.; Li, G.-G.; Quan, R.-C. Molecular phylogeny of the genus Muntiacus with special emphasis on the phylogenetic position of Muntiacus gongshanensis. Zool. Res. 2019, 42, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbour Lab Press: New York, NY, USA, 1989; pp. 1–1659. [Google Scholar]
- Rahman, M.M.; Yoon, K.B.; Kim, J.Y.; Hussin, M.Z.; Park, Y.C. Complete mitochondrial genome sequence of the Indian pipistrelle Pipistrellus coromandra (Vespertilioninae). Animal Cells Sys. 2016, 20, 86–94. [Google Scholar] [CrossRef][Green Version]
- Langmead, B.; Wilks, C.; Antonescu, V.; Charles, R. Scaling read aligners to hundreds of threads on general-purpose processors. Bioinformatics 2019, 35, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Locatelli, A.G.; Jebb, D.; Teeling, E.C. The complete mitochondrial genome of Kuhl’s pipistrelle, Pipistrellus kuhlii (Chiroptera: Vespertilionidae). Mitochondrial DNA Part B 2016, 1, 423–424. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2010, 39, D225–D229. [Google Scholar] [CrossRef]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phyl. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Laslett, D.; Canbäck, B. ARWEN: A program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.R.; Lim, S.J.; Kim, H.J.; Cho, J.Y.; Park, Y.C. Complete mitochondrial genome of the house bat Pipistrellus abramus (Mammalia: Chiroptera) from Korea. Mitochondrial DNA Part B 2017, 2, 540–541. [Google Scholar] [CrossRef]
- Kruskop, S.V.; Solovyeva, E.N.; Kaznadzey, A.D. Unusual Pipistrelle: Taxonomic position of the Malayan Noctule (Pipistrellus stenopterus; Vespertilionidae; Chiroptera). Zool. Stud. 2018, 57, 1–15. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Type | Gene | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ND1 | ND2 | COX1 | COX2 | ATP8 | ATP6 | COX3 | ND3 | ND4L | ND4 | ND5 | ND6 | CYTB | |
| GTR+F+G4 | 1st pos | 1st pos | 1st pos | 1st pos | 1st pos | ||||||||
| TPM3u+F+I+G4 | 2nd pos | 2nd pos | 2nd pos | 2nd pos | 2nd pos | 2rd pos | |||||||
| TIM+F+I+G4 | 3rd pos | 3rd pos | 3rd pos | ||||||||||
| TIM2+F+I+G4 | 1st pos | 1st pos, 2nd pos | 1st pos | 1st pos | 1st pos | ||||||||
| TPM3u+F+I+G4 | 2nd pos | 2nd pos | 2nd pos | 2nd pos | 2nd pos | ||||||||
| TIM2e+I+G4 | 1st pos | 1st pos | |||||||||||
| TIM2+F+I+G4 | 3rd pos | 3rd pos | 3rd pos | 3rd pos | |||||||||
| TN+F+I+G4 | 3rd pos | 3rd pos | 3rd pos | 3rd pos | 3rd pos | ||||||||
| HKY+F+I+G4 | 1st pos, 2nd pos | ||||||||||||
| HKY+F+G4 | 3rd pos | ||||||||||||
| Species | Mitochondrion | cytb |
|---|---|---|
| Glischropus aquilus | KR612333.1 | |
| G. bucephalus | OR667258 | KR612331.1, KR612332.1, OR667259, OR667260, OR667261 |
| G. tylopus | JX570898.1, EU521632.1, OR667262, OR667263 | |
| “Pipistrellus coromandra” | NC_029191.1 | NC_029191.1 |
| Nyctalus aviator | NC_060309.1 | NC_060309.1, MK167360.1 |
| N. labiata | NC_027237.1, NC_041160.1 | NC_027237.1, NC_041160.1, KX467596.1 |
| N. lasiopterus | DQ120867.1, EU360680.1, JX570900.1 | |
| N. leisleri | DQ120877.1, JX570901.1, EU360690.1 | |
| N. noctula | MN122876.1, MN122907.1 | MN122907.1, MN122876.1, DQ120872.1 |
| Pipistrellus abramus | KX355640.1, NC_005436.1 | GQ332529.1, KX355640.1, NC_005436.1 |
| P. deserti | KM252759.1 | |
| P. coromandra | OR667264, OR667265, OR667266, OR667267 | |
| P. dhofarensis | KX375145.1, KX375148.1 | |
| P. hesperidus | MN790830.1, MT778037.1, MN790820.1 | |
| P. javanicus | KX496357.1 | |
| P. kuhlii | KU058655.1 | KU058655.1, DQ120845.1, EU360657.1 |
| P. maderensis | KC520771.1, KC520774.1, MT374272.1 | |
| P. nanulus | MK188530.1 | |
| P. nathusii | MN122914.1 | MN122914.1, AJ504446.1, DQ120849.1 |
| P. paterculus | OR667268, OR667269, OR667270 | |
| P. pipistrellus | LR862378.1 | KF874520.1, DQ120853.1, LR862378.1 |
| P. pygmaeus | MN122927.1, OX465325.1 | MN122927.1, OX465325.1, EU084882.1 |
| P. raceyi | KM886094.1, KM886088.1 | |
| P. rusticus | KX375166.1, KX375167.1 | |
| P. stenopterus | MH540194.1 | |
| Plecotus auritus | MN122881.1, MT410875.1 | |
| P. macrobullaris | KR134372.1, KR134385.1 | |
| Hypsugo alaschanicus | MF459671.1, MK135784.1, NC_029939.1 | |
| Lasionycteris noctivagans | MT774150.1, MT774151.1, NC_050995.1 | |
| Chalinolobus tuberculatus | NC 002626.1 | |
| Eptesicus bottae | NC_070014.1, OP328299.1, OP328300.1 | |
| E. nilssonii | OX621305.1 | |
| Vespertilio murinus | NC_033347.1 | NC_033347.1 |
| V. sinensis | KJ081440.1, KM092493.1. | KJ081440.1, KM092493.1. |
| Myotis brandtii | NC_025308.1 | |
| M. horsfieldii | MF143494.1 | |
| M. muricola | KT213444.1 |
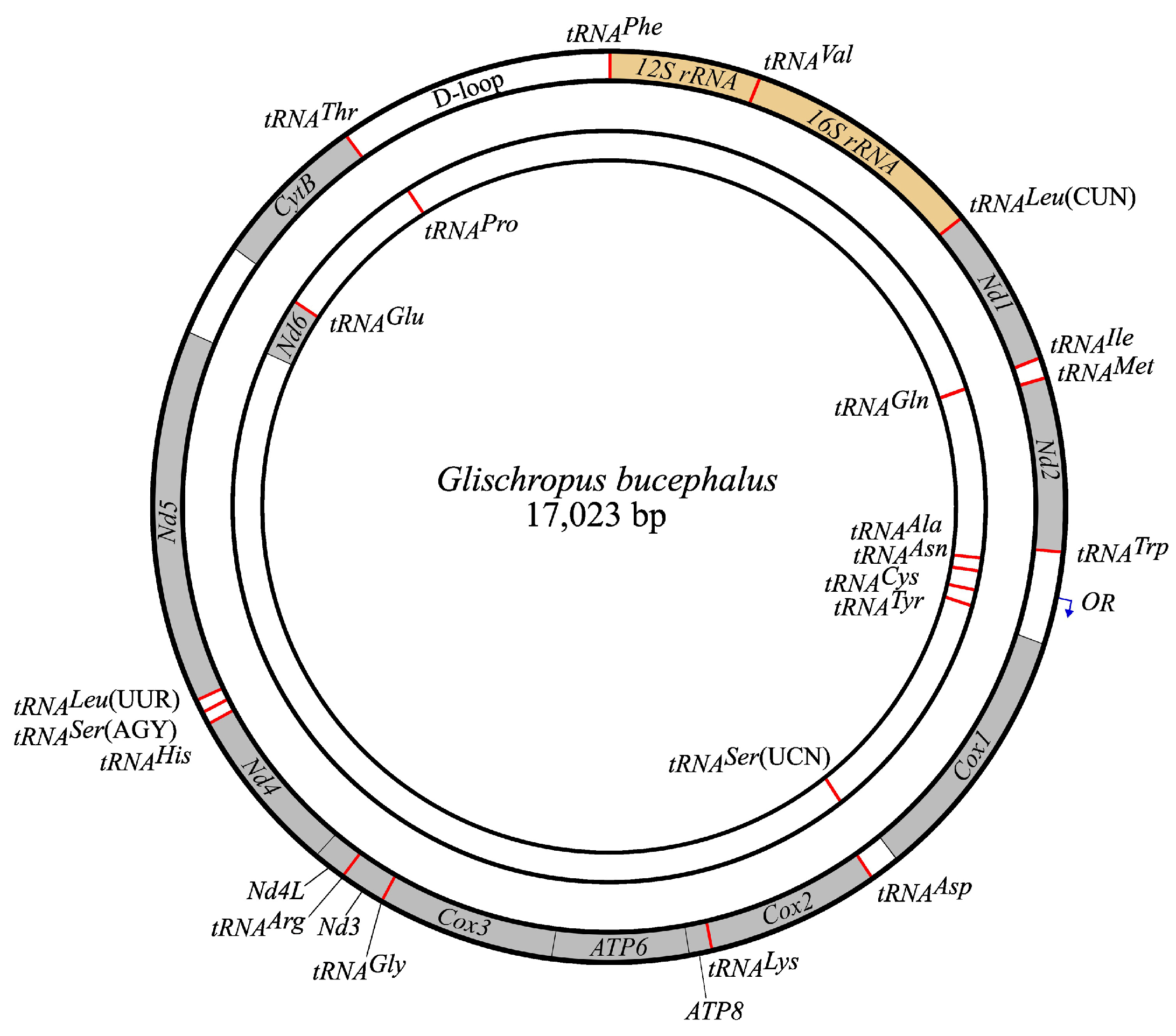
| Start Position | Stop Position | Length (bp) | Anticodon | Start Codon | Stop Codon | Strand | |
|---|---|---|---|---|---|---|---|
| tRNAPhe | 1 | 73 | 73 | GAA | + | ||
| 12S rRNA | 74 | 1010 | 937 | + | |||
| tRNAVal | 1011 | 1078 | 68 | TAC | + | ||
| 16S rRNA | 1079 | 2644 | 1566 | + | |||
| tRNALeu | 2650 | 2725 | 76 | TAA | + | ||
| Nd1 | 2731 | 3684 | 954 | ATG | TA- | + | |
| tRNAIle | 3687 | 3755 | 69 | GAT | + | ||
| tRNAGln | 3753 | 3827 | 75 | TTG | - | ||
| tRNAMet | 3828 | 3896 | 69 | CAT | + | ||
| Nd2 | 3897 | 4937 | 1041 | ATT | T- | + | |
| tRNATrp | 4939 | 5005 | 67 | TCA | + | ||
| tRNAAla | 5013 | 5081 | 69 | TGC | - | ||
| tRNAAsn | 5082 | 5154 | 73 | GTT | - | ||
| OR | 5155 | 5189 | 35 | ||||
| tRNACys | 5187 | 5252 | 66 | GCA | - | ||
| tRNATyr | 5253 | 5319 | 67 | GTA | - | ||
| Cox1 | 5321 | 6862 | 1542 | ATG | TAA | + | |
| tRNASer | 6869 | 6937 | 69 | TGA | - | ||
| tRNAAsp | 6945 | 7011 | 67 | GTC | + | ||
| Cox2 | 7012 | 7692 | 681 | ATG | TAA | + | |
| tRNALys | 7699 | 7765 | 67 | TTT | + | ||
| ATP8 | 7767 | 7967 | 201 | ATG | TAA | + | |
| ATP6 | 7928 | 8605 | 678 | ATG | TAA | + | |
| Cox3 | 8608 | 9390 | 783 | ATG | TA- | + | |
| tRNAGly | 9392 | 9460 | 69 | TCC | + | ||
| Nd3 | 9461 | 9805 | 345 | ATT | TA- | + | |
| tRNAArg | 9809 | 9878 | 70 | TCG | + | ||
| Nd4L | 9880 | 10,173 | 294 | ATG | TAA | + | |
| Nd4 | 10,170 | 11,546 | 1377 | ATG | T- | + | |
| tRNAHis | 11,548 | 11,616 | 69 | GTG | + | ||
| tRNASer | 11,617 | 11,675 | 59 | GCT | + | ||
| tRNALeu | 11,676 | 11,745 | 70 | TAG | + | ||
| Nd5 | 11,764 | 13,552 | 1789 | ATA | TAA | + | |
| Nd6 | 13,544 | 14,062 | 519 | ATG | TAA | - | |
| tRNAGlu | 14,066 | 14,133 | 68 | TTC | - | ||
| CytB | 14,139 | 15,275 | 1137 | ATG | AGA | + | |
| tRNAThr | 15,279 | 15,348 | 70 | TGT | + | ||
| tRNAPro | 15,348 | 15,416 | 69 | TGG | - | ||
| D-loop | 15,416 | 17,023 | 1608 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhukova, S.S.; Speranskaya, A.S.; Lisenkova, A.A.; Kruskop, S.V. The Complete Mitochondrial Genome of Glischropus bucephalus (Vespertilionidae; Chiroptera) Provides New Evidence for Pipistrellus Paraphyly. Diversity 2023, 15, 1085. https://doi.org/10.3390/d15101085
Zhukova SS, Speranskaya AS, Lisenkova AA, Kruskop SV. The Complete Mitochondrial Genome of Glischropus bucephalus (Vespertilionidae; Chiroptera) Provides New Evidence for Pipistrellus Paraphyly. Diversity. 2023; 15(10):1085. https://doi.org/10.3390/d15101085
Chicago/Turabian StyleZhukova, Svetlana S., Anna S. Speranskaya, Alexandra A. Lisenkova, and Sergei V. Kruskop. 2023. "The Complete Mitochondrial Genome of Glischropus bucephalus (Vespertilionidae; Chiroptera) Provides New Evidence for Pipistrellus Paraphyly" Diversity 15, no. 10: 1085. https://doi.org/10.3390/d15101085
APA StyleZhukova, S. S., Speranskaya, A. S., Lisenkova, A. A., & Kruskop, S. V. (2023). The Complete Mitochondrial Genome of Glischropus bucephalus (Vespertilionidae; Chiroptera) Provides New Evidence for Pipistrellus Paraphyly. Diversity, 15(10), 1085. https://doi.org/10.3390/d15101085